
PPIC, EMP3 and CHI3L1 Are Novel Prognostic Markers for High Grade Glioma

Abstract
:1. Introduction
2. Results
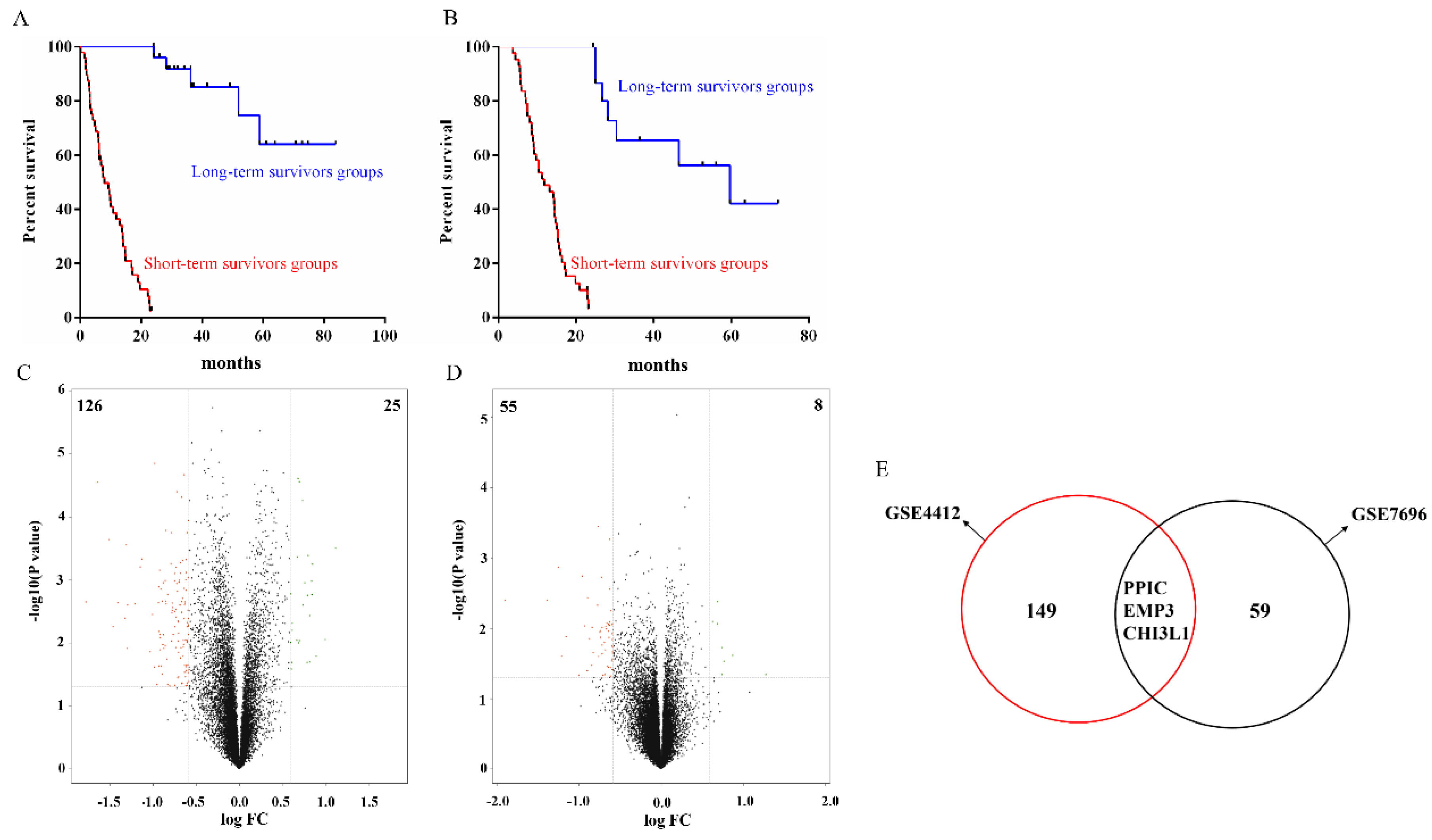
2.1. Identification of DEPGs
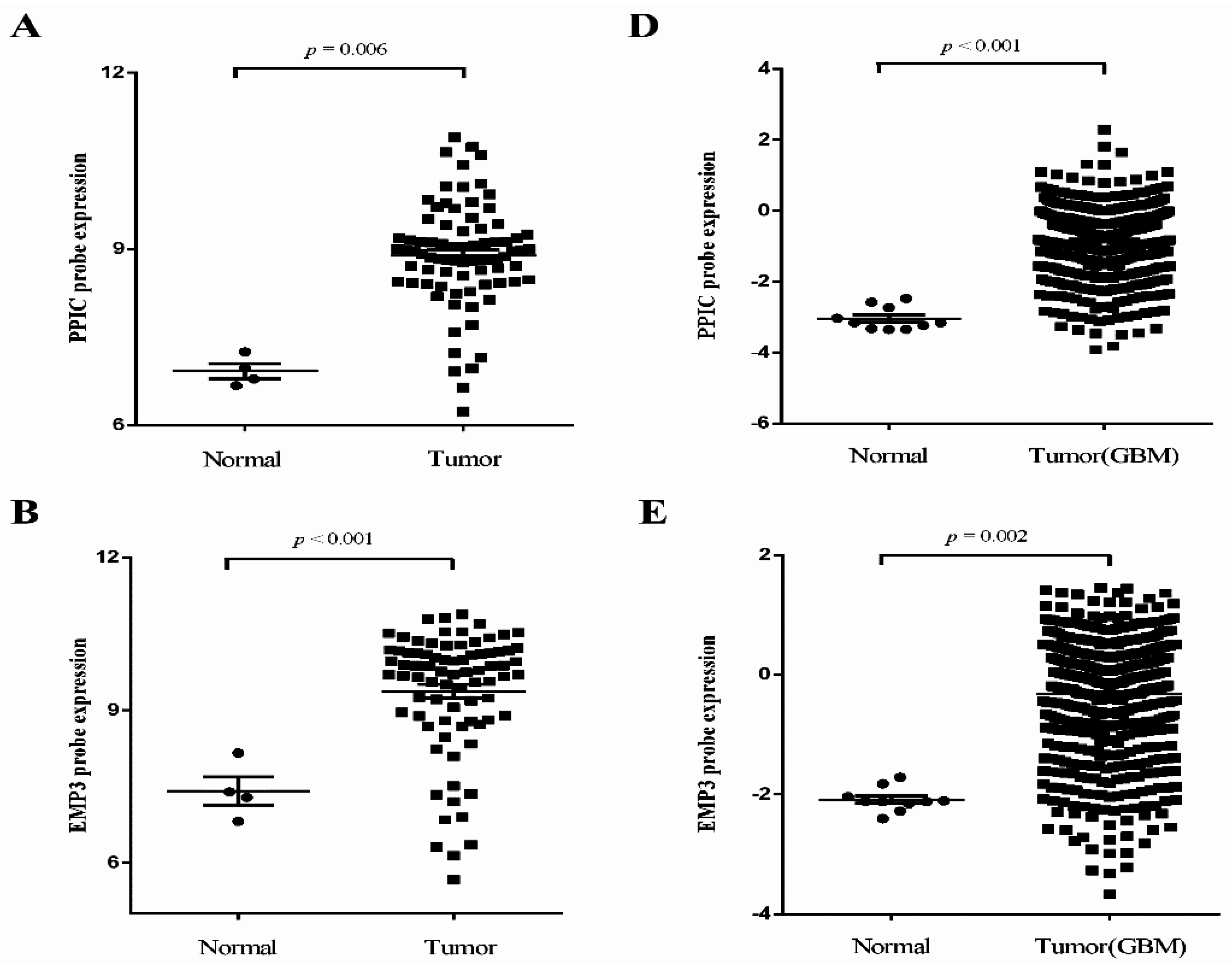
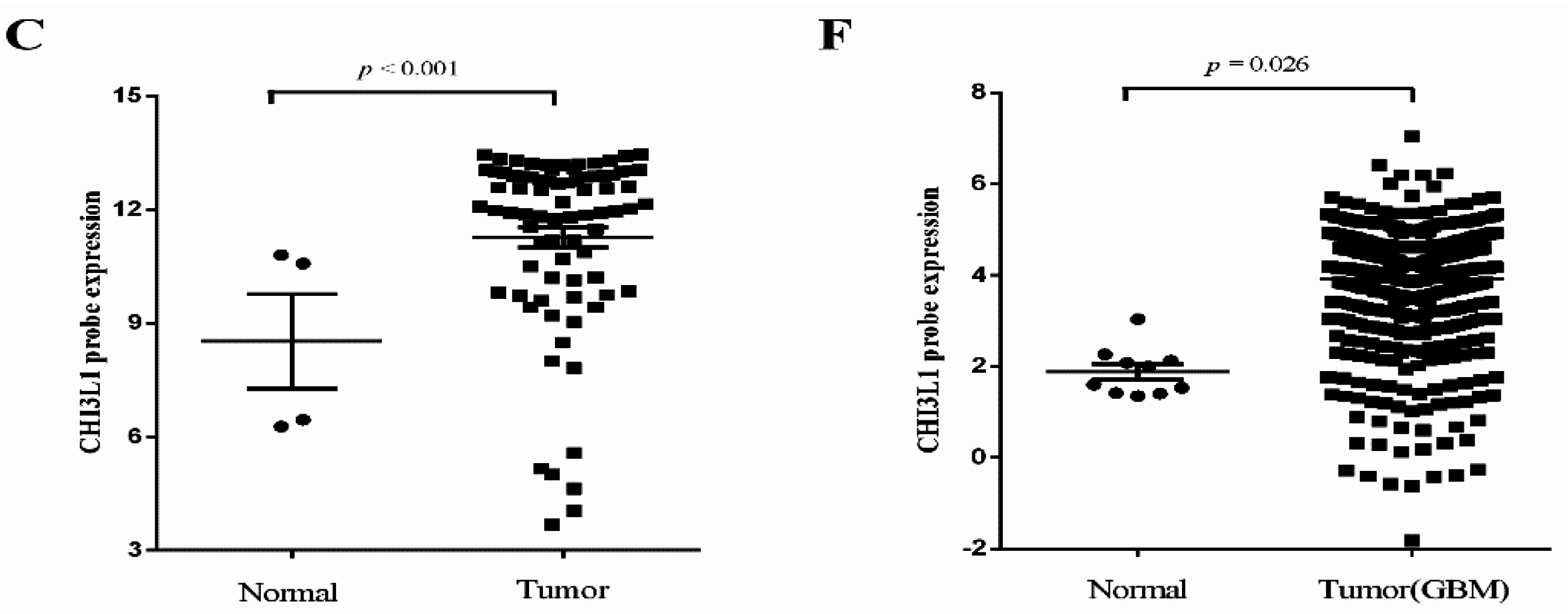
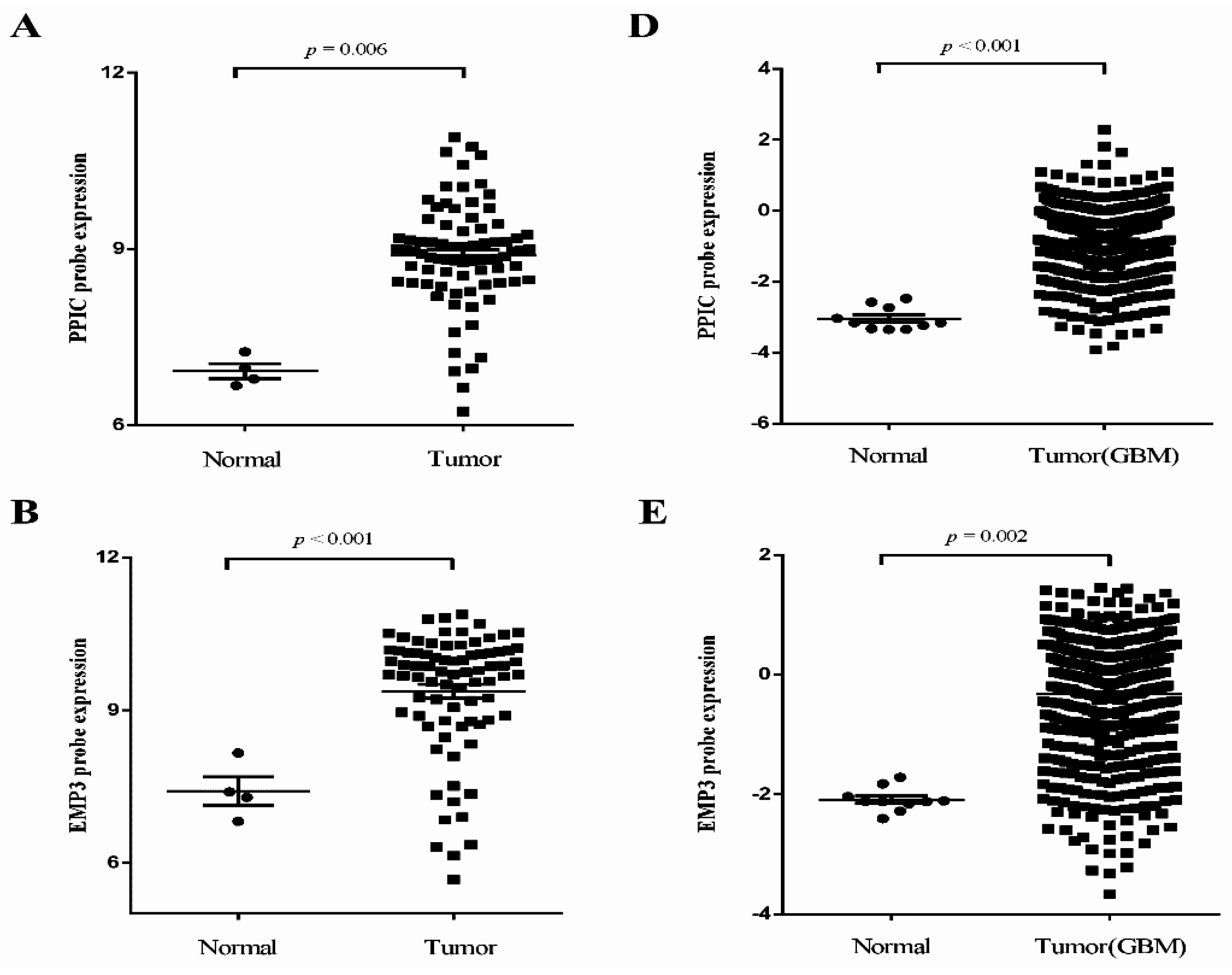
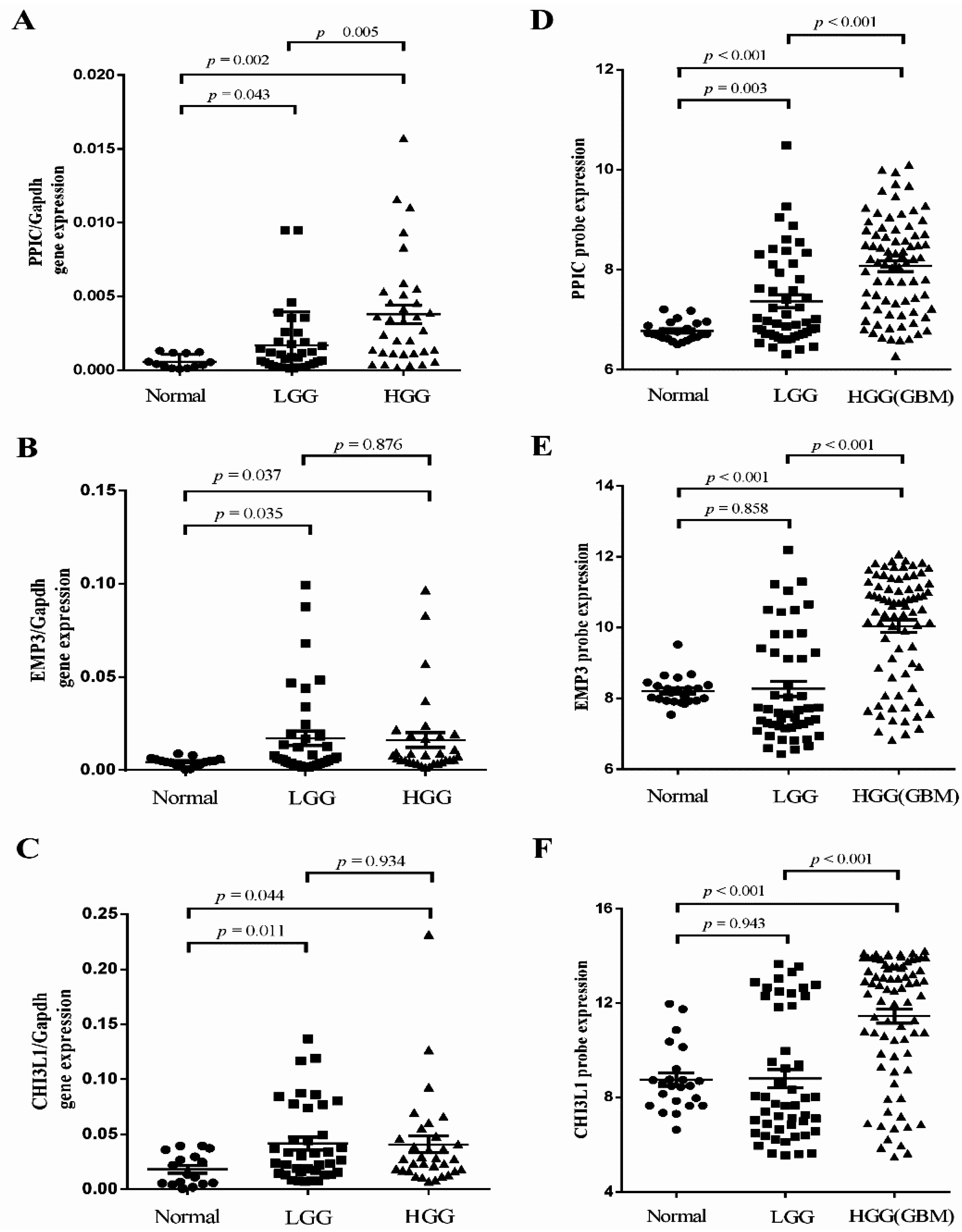
2.2. Independent Validation of Glioma-Specific Markers
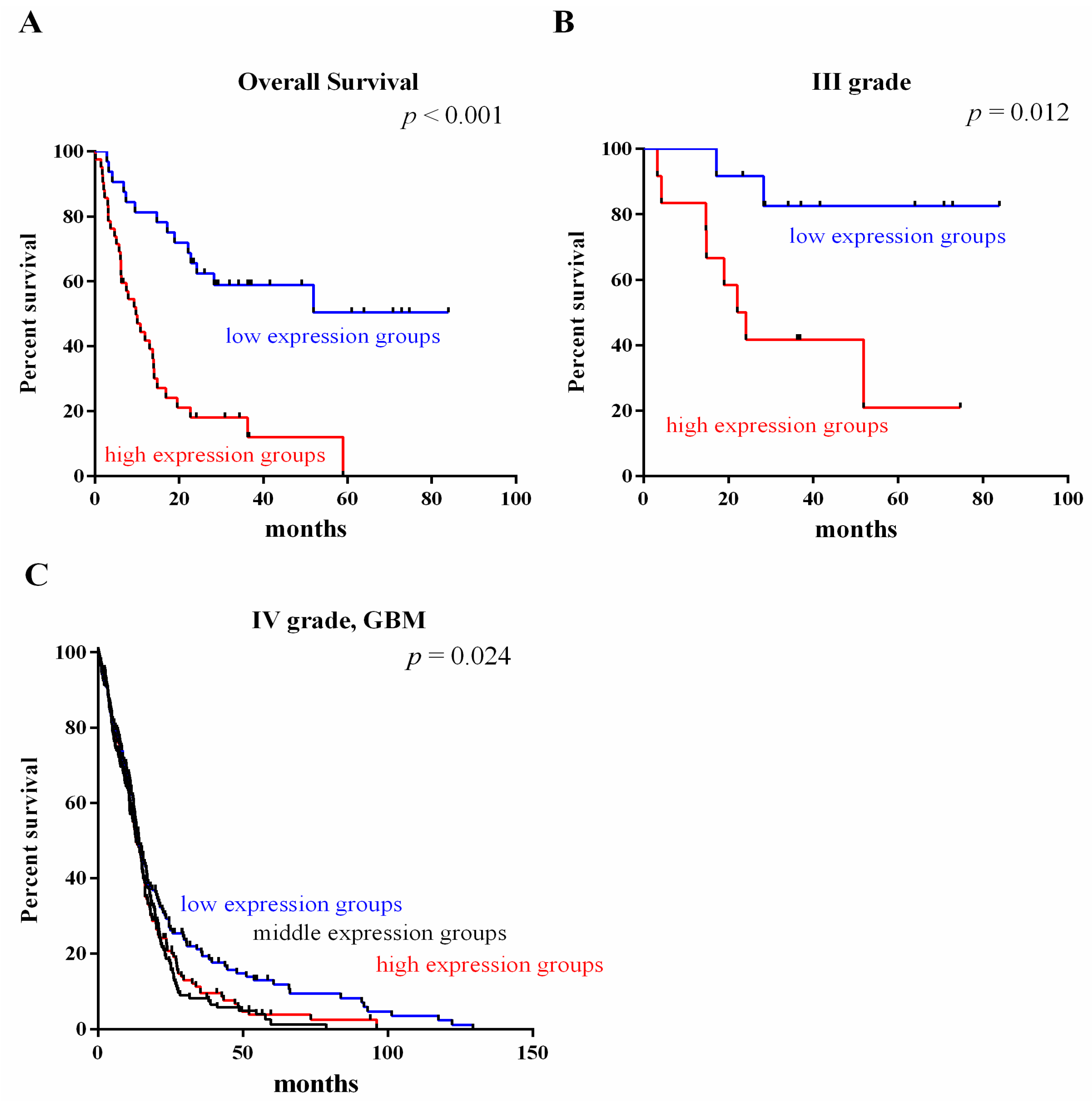
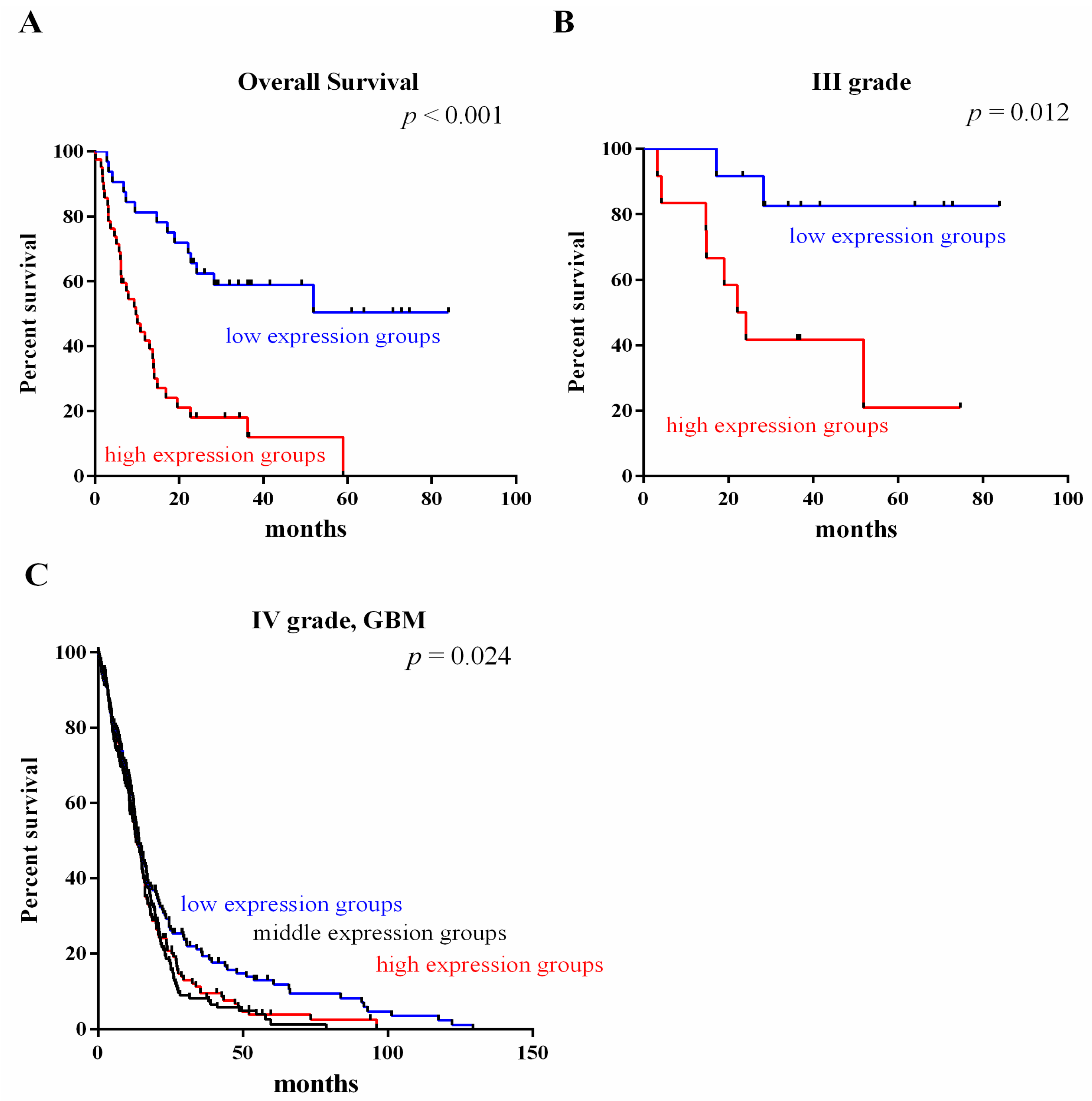
2.3. Survival Value Validation of Patients with Grades III and IV Gliomas by the Three-Gene Signature
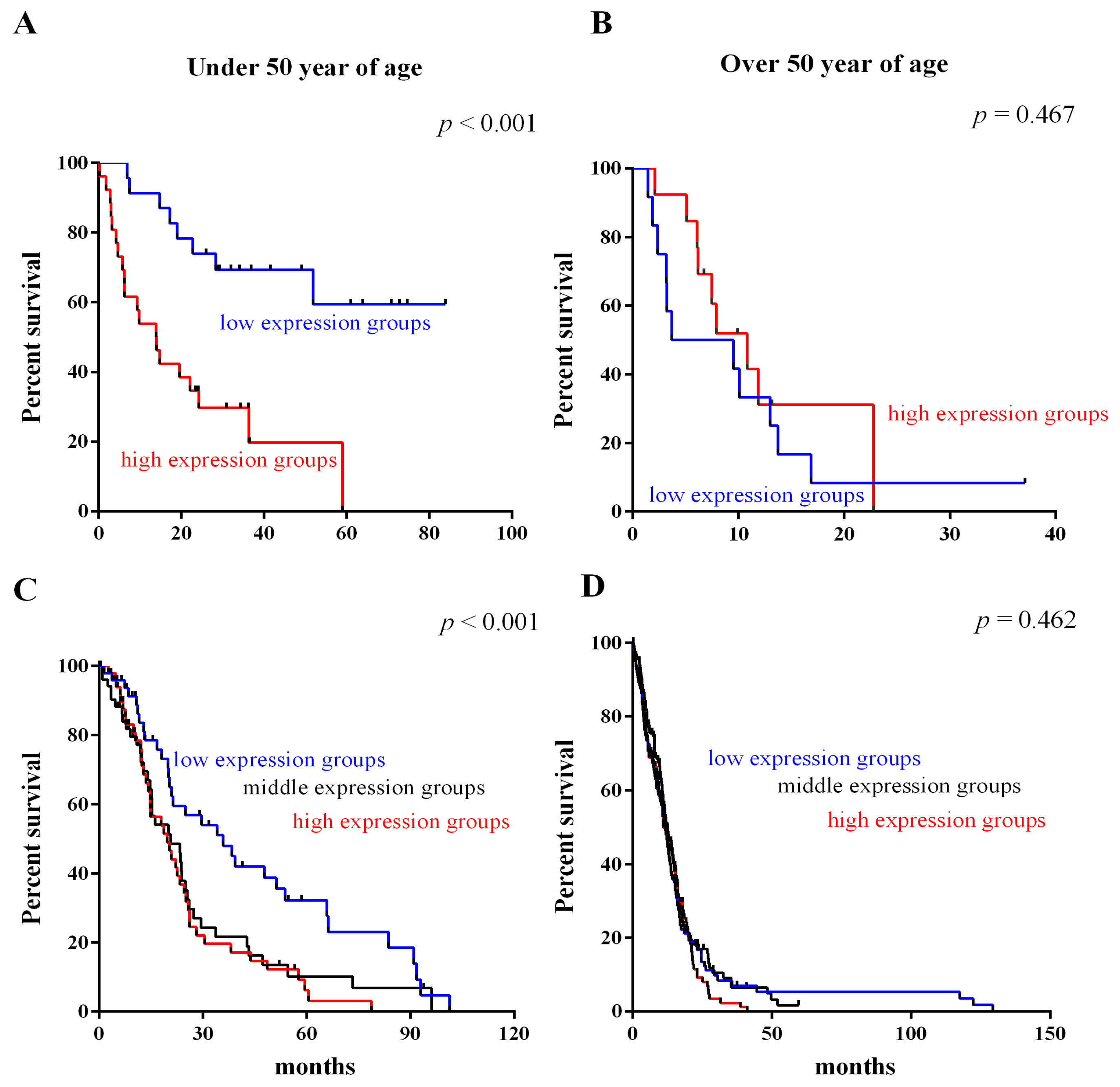
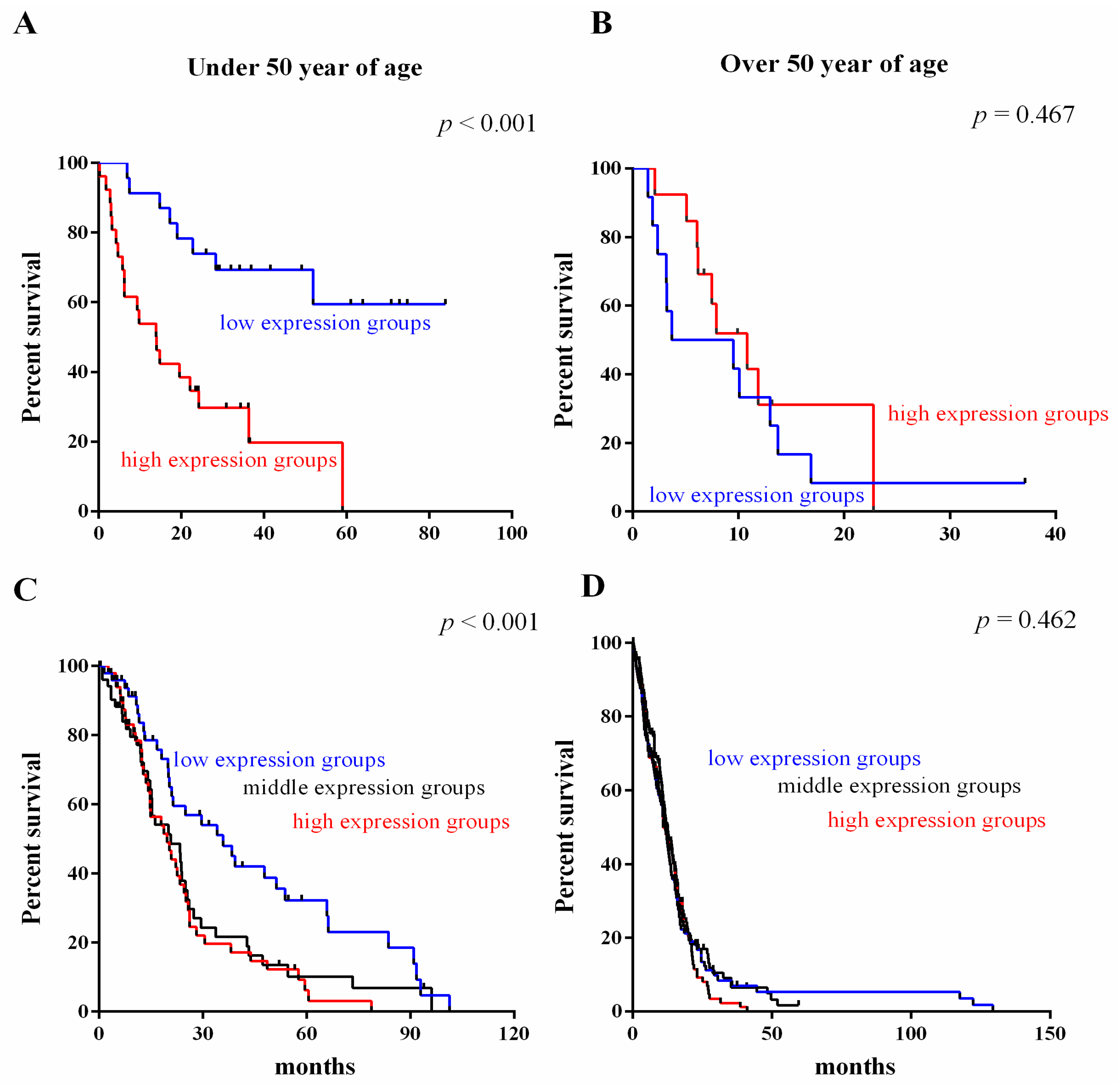
2.4. Survival Value Validation of Patients with Age by the Three-Gene Signature
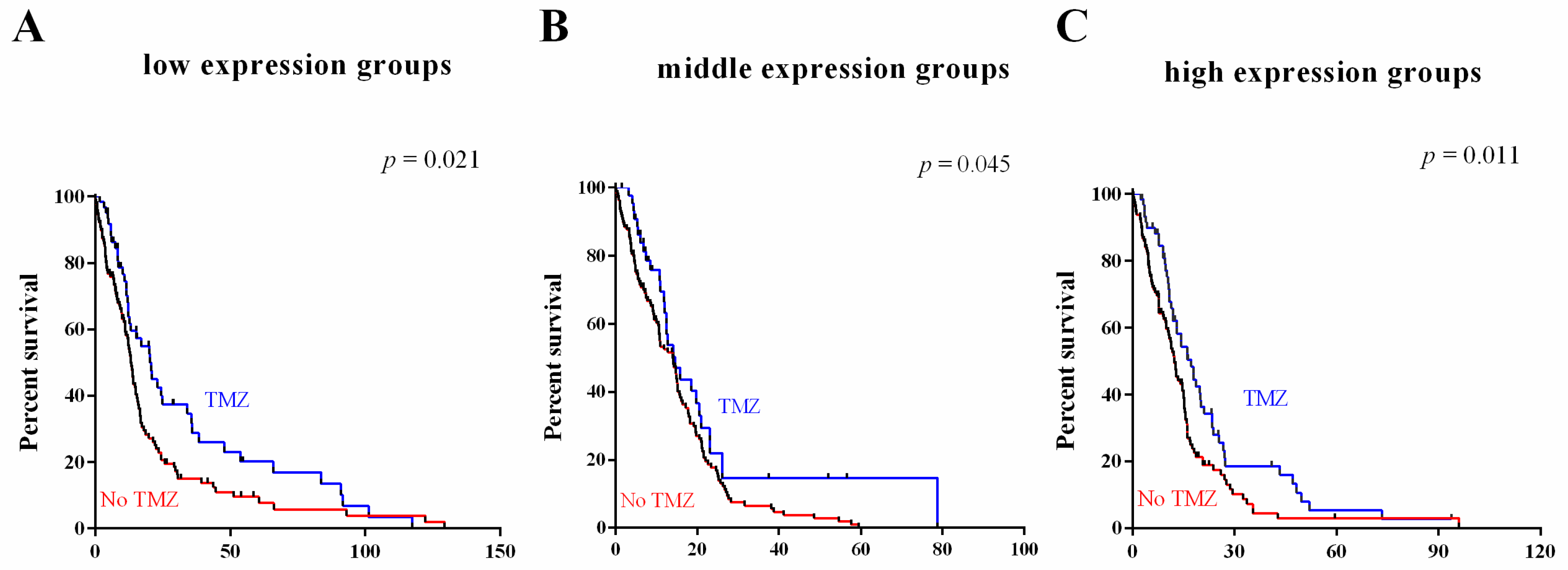
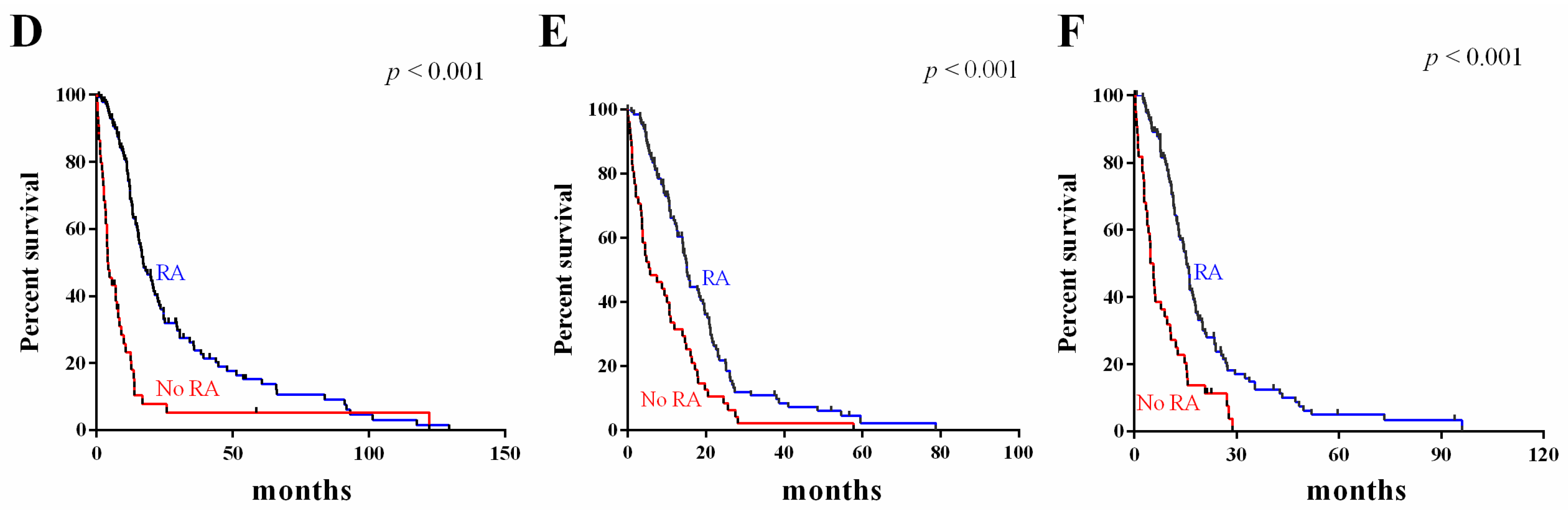
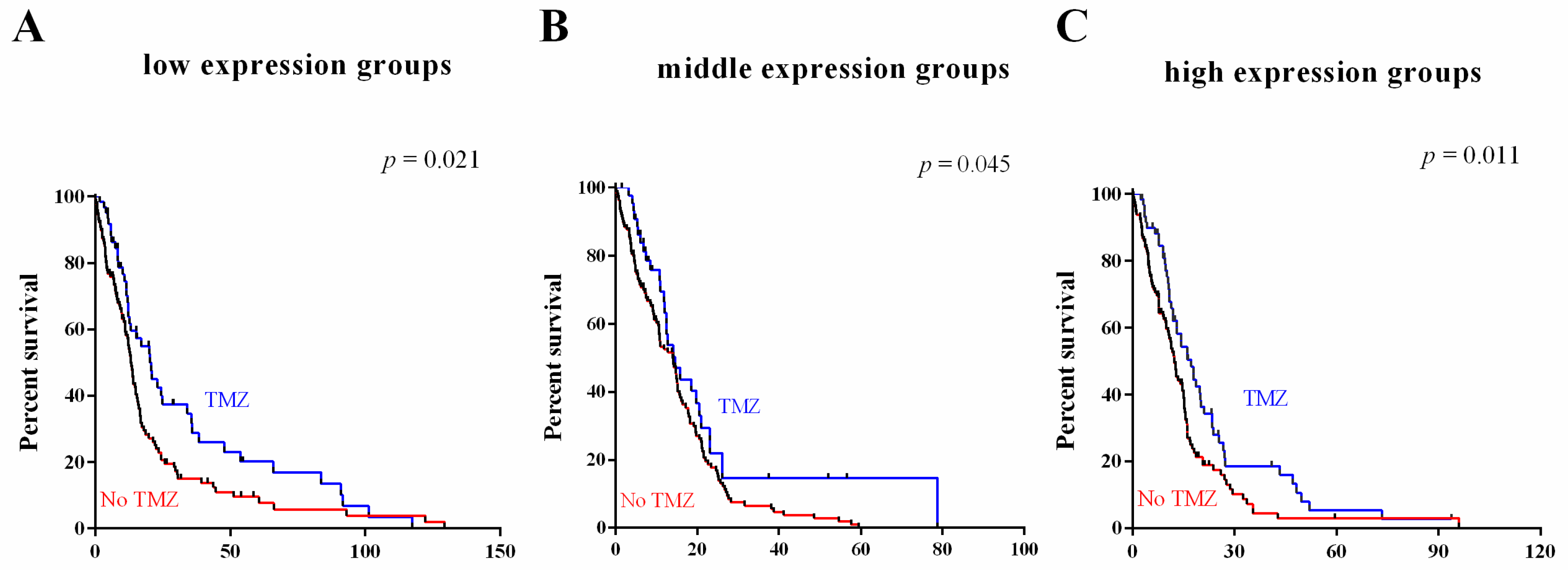
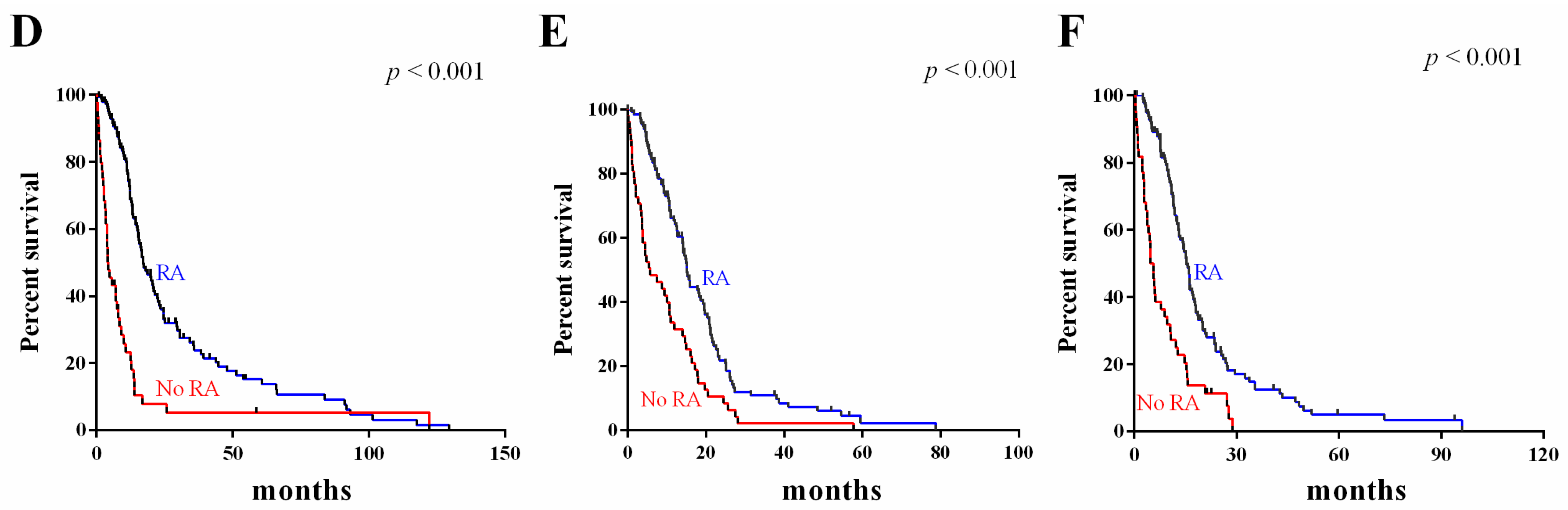
2.5. Survival Value Validation of Patients with Temozolomide (TMZ) and Radiotherapy by the Three-Gene Signature
3. Discussion
4. Materials and Methods
4.1. Affymetrix Microarray Data
4.2. Differentially Expressed Prognostic Genes (DEPGs) Analysis
4.3. Patients and Tissue Samples
4.4. RNA Extraction and Reverse-Transcription PCR from Human Tissue
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Charles, N.A.; Holland, E.C.; Gilbertson, R.; Glass, R.; Kettenmann, H. The brain tumor microenvironment. Glia 2012, 60, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Goebell, E.; Paustenbach, S.; Vaeterlein, O.; Ding, X.Q.; Heese, O.; Fiehler, J.; Kucinski, T.; Hagel, C.; Westphal, M.; Zeumer, H. Low-grade and anaplastic gliomas: Differences in architecture evaluated with diffusion-tensor MR imaging. Radiology 2006, 239, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Buckner, J.C. Factors influencing survival in high-grade gliomas. Semin. Oncol. 2003, 30, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Tan, F.; Hess, K.R.; Yung, W.K. The expression of PAX6, PTEN, vascular endothelial growth factor, and epidermal growth factor receptor in gliomas: Relationship to tumor grade and survival. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 3369–3375. [Google Scholar]
- Chakravarti, A.; Noll, E.; Black, P.M.; Finkelstein, D.F.; Finkelstein, D.M.; Dyson, N.J.; Loeffler, J.S. Quantitatively determined survivin expression levels are of prognostic value in human gliomas. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 1063–1068. [Google Scholar] [CrossRef]
- Nutt, C.L.; Mani, D.R.; Betensky, R.A.; Tamayo, P.; Cairncross, J.G.; Ladd, C.; Pohl, U.; Hartmann, C.; McLaughlin, M.E.; Batchelor, T.T.; et al. Gene expression-based classification of malignant gliomas correlates better with survival than histological classification. Cancer Res. 2003, 63, 1602–1607. [Google Scholar] [PubMed]
- Fuller, G.N.; Hess, K.R.; Rhee, C.H.; Yung, W.K.; Sawaya, R.A.; Bruner, J.M.; Zhang, W. Molecular classification of human diffuse gliomas by multidimensional scaling analysis of gene expression profiles parallels morphology-based classification, correlates with survival, and reveals clinically-relevant novel glioma subsets. Brain Pathol. 2002, 12, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Sallinen, S.L.; Sallinen, P.K.; Haapasalo, H.K.; Helin, H.J.; Helen, P.T.; Schraml, P.; Kallioniemi, O.P.; Kononen, J. Identification of differentially expressed genes in human gliomas by DNA microarray and tissue chip techniques. Cancer Res. 2000, 60, 6617–6622. [Google Scholar] [PubMed]
- Shai, R.; Shi, T.; Kremen, T.J.; Horvath, S.; Liau, L.M.; Cloughesy, T.F.; Mischel, P.S.; Nelson, S.F. Gene expression profiling identifies molecular subtypes of gliomas. Oncogene 2003, 22, 4918–4923. [Google Scholar] [CrossRef] [PubMed]
- Laenen, G.; Thorrez, L.; Bornigen, D.; Moreau, Y. Finding the targets of a drug by integration of gene expression data with a protein interaction network. Mol. Biosyst. 2013, 9, 1676–1685. [Google Scholar] [CrossRef] [PubMed]
- Morine, M.J.; Toomey, S.; McGillicuddy, F.C.; Reynolds, C.M.; Power, K.A.; Browne, J.A.; Loscher, C.; Mills, K.H.; Roche, H.M. Network analysis of adipose tissue gene expression highlights altered metabolic and regulatory transcriptomic activity in high-fat-diet-fed IL-1RI knockout mice. J. Nutr. Biochem. 2013, 24, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Chao, Y.; Tong, W.; Yu, S. Sugar coordinately and differentially regulates growth- and stress-related gene expression via a complex signal transduction network and multiple control mechanisms. Plant Physiol. 2001, 125, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Van den Boom, J.; Wolter, M.; Kuick, R.; Misek, D.E.; Youkilis, A.S.; Wechsler, D.S.; Sommer, C.; Reifenberger, G.; Hanash, S.M. Characterization of gene expression profiles associated with glioma progression using oligonucleotide-based microarray analysis and real-time reverse transcription-polymerase chain reaction. Am. J. Pathol. 2003, 163, 1033–1043. [Google Scholar] [CrossRef]
- Wei, B.; Wang, L.; Du, C.; Hu, G.; Wang, L.; Jin, Y.; Kong, D. Identification of differentially expressed genes regulated by transcription factors in glioblastomas by bioinformatics analysis. Mol. Med. Rep. 2015, 11, 2548–2554. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Diehn, M.; Watson, N.; Bollen, A.W.; Aldape, K.D.; Nicholas, M.K.; Lamborn, K.R.; Berger, M.S.; Botstein, D.; Brown, P.O.; et al. Gene expression profiling reveals molecularly and clinically distinct subtypes of glioblastoma multiforme. Proc. Natl. Acad. Sci. USA 2005, 102, 5814–5819. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Charara, N.; Schmitz, R.; Wehrli, S.; Mikol, V.; Zurini, M.G.; Quesniaux, V.F.; Movva, N.R. Human cyclophilin C: Primary structure, tissue distribution, and determination of binding specificity for cyclosporins. Biochemistry 1994, 33, 8218–8224. [Google Scholar] [CrossRef] [PubMed]
- Barik, S. Immunophilins: For the love of proteins. Cell. Mol. Life Sci. CMLS 2006, 63, 2889–2900. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Heitman, J. The cyclophilins. Genome Biol. 2005, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, D.C.; Stocki, P.; Williams, D.B. Cyclophilin C participates in the US2-mediated degradation of major histocompatibility complex class I molecules. PLoS ONE 2015, 10, e0145458. [Google Scholar] [CrossRef] [PubMed]
- Chicheportiche, Y.; Vassalli, P. Cloning and expression of a mouse macrophage cDNA coding for a membrane glycoprotein of the scavenger receptor cysteine-rich domain family. J. Biol. Chem. 1994, 269, 5512–5517. [Google Scholar] [PubMed]
- Trahey, M.; Weissman, I.L. Cyclophilin C-associated protein: A normal secreted glycoprotein that down-modulates endotoxin and proinflammatory responses in vivo. Proc. Natl. Acad. Sci. USA 1999, 96, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.; Trahey, M.; Weissman, I. Cloning and characterization of cyclophilin C-associated protein: A candidate natural cellular ligand for cyclophilin C. Proc. Natl. Acad. Sci. USA 1993, 90, 6815–6819. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Imai, H.; Seki, K.; Tomizawa, S.; Nakamura, M.; Honda, F.; Kawahara, N.; Saito, N. Cyclophilin C-associated protein and cyclophilin C mRNA are upregulated in penumbral neurons and microglia after focal cerebral ischemia. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2005, 25, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Li, S.; Longaker, M.T.; Lorenz, H.P. Cyclophilin C-associated protein is up-regulated during wound healing. J. Cell. Physiol. 2007, 210, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Longaker, M.T.; Lorenz, H.P. Cyclophilin C-associated protein is a mediator for fibronectin fragment-induced matrix metalloproteinase-13 expression. J. Biol. Chem. 2004, 279, 55334–55340. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Lin, B.W.; Li, S.; Longaker, M.T.; Lorenz, H.P. Cyclophilin C-associated protein/Mac-2 binding protein colocalizes with calnexin and regulates the expression of tissue transglutaminase. J. Cell. Physiol. 2010, 223, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.; Suter, U. Epithelial membrane protein-2 and epithelial membrane protein-3: Two novel members of the peripheral myelin protein 22 gene family. Gene 1996, 175, 115–120. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Benvenisty, N. Characterization of a tumor-associated gene, a member of a novel family of genes encoding membrane glycoproteins. Gene 1996, 183, 69–75. [Google Scholar] [CrossRef]
- Alaminos, M.; Davalos, V.; Ropero, S.; Setien, F.; Paz, M.F.; Herranz, M.; Fraga, M.F.; Mora, J.; Cheung, N.K.; Gerald, W.L.; et al. EMP3, a myelin-related gene located in the critical 19q13.3 region, is epigenetically silenced and exhibits features of a candidate tumor suppressor in glioma and neuroblastoma. Cancer Res. 2005, 65, 2565–2571. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.; Cheung, N.K.; Chen, L.; Qin, J.; Gerald, W. Loss of heterozygosity at 19q13.3 is associated with locally aggressive neuroblastoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2001, 7, 1358–1361. [Google Scholar]
- Reifenberger, G.; Collins, V.P. Pathology and molecular genetics of astrocytic gliomas. J. Mol. Med. 2004, 82, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Von Deimling, A.; Fimmers, R.; Schmidt, M.C.; Bender, B.; Fassbender, F.; Nagel, J.; Jahnke, R.; Kaskel, P.; Duerr, E.M.; Koopmann, J.; et al. Comprehensive allelotype and genetic anaysis of 466 human nervous system tumors. J. Neuropathol. Exp. Neurol. 2000, 59, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, M.K.; Gilbert, M.R.; Holland, E.C. Gene expression microarray analysis reveals YKL-40 to be a potential serum marker for malignant character in human glioma. Cancer Res. 2002, 62, 4364–4368. [Google Scholar] [PubMed]
- Shao, R.; Taylor, S.L.; Oh, D.S.; Schwartz, L.M. Vascular heterogeneity and targeting: The role of YKL-40 in glioblastoma vascularization. Oncotarget 2015, 6, 40507–40518. [Google Scholar] [PubMed]
- Iwamoto, F.M.; Hottinger, A.F.; Karimi, S.; Riedel, E.; Dantis, J.; Jahdi, M.; Panageas, K.S.; Lassman, A.B.; Abrey, L.E.; Fleisher, M.; et al. Serum YKL-40 is a marker of prognosis and disease status in high-grade gliomas. Neuro-Oncol. 2011, 13, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Hormigo, A.; Gu, B.; Karimi, S.; Riedel, E.; Panageas, K.S.; Edgar, M.A.; Tanwar, M.K.; Rao, J.S.; Fleisher, M.; DeAngelis, L.M.; et al. YKL-40 and matrix metalloproteinase-9 as potential serum biomarkers for patients with high-grade gliomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 5698–5704. [Google Scholar] [CrossRef] [PubMed]
- Serao, N.V.; Delfino, K.R.; Southey, B.R.; Beever, J.E.; Rodriguez-Zas, S.L. Cell cycle and aging, morphogenesis, and response to stimuli genes are individualized biomarkers of glioblastoma progression and survival. BMC Med. Genom. 2011, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Steponaitis, G.; Skiriute, D.; Kazlauskas, A.; Golubickaite, I.; Stakaitis, R.; Tamasauskas, A.; Vaitkiene, P. High CHI3L1 expression is associated with glioma patient survival. Diagn. Pathol. 2016, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Larsson, O.; Wahlestedt, C.; Timmons, J.A. Considerations when using the significance analysis of microarrays (SAM) algorithm. BMC Bioinform. 2005, 6, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N% | PPIC Expression Levels | EMP3 Expression Levels | CHI3L1 Expression Levels | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| High | Low | Ratio | p | High | Low | Ratio | p | High | Low | Ratio | p | ||
| Expression | Expression | (High/Low) | Expression | Expression | (High/Low) | Expression | Expression | (High/Low) | |||||
| Sex | |||||||||||||
| Male | 48 (68.57) | 13 | 35 | 0.371 | 0.184 | 16 | 32 | 0.5 | 0.835 | 15 | 33 | 0.454 | 0.441 |
| Female | 22 (31.42) | 10 | 12 | 0.833 | 5 | 17 | 0.294 | 7 | 15 | 0.467 | |||
| Age, year | |||||||||||||
| <45 | 45 (64.28) | 13 | 32 | 0.406 | 0.068 | 14 | 31 | 0.452 | 0.762 | 13 | 32 | 0.406 | 0.965 |
| ≥45 | 25 (35.72) | 11 | 14 | 0.786 | 7 | 18 | 0.389 | 9 | 16 | 0.563 | |||
| Grade | |||||||||||||
| Low (I + II) | 38 (54.28) | 7 | 31 | 0.226 | 0.005 | 11 | 27 | 0.407 | 0.876 | 13 | 25 | 0.52 | 0.935 |
| High (III + IV) | 32 (45.71) | 16 | 16 | 0.500 | 10 | 22 | 0.454 | 9 | 23 | 0.391 | |||
| Sequence | GSE4412 | GSE7696 | GSE4290 | TCGA |
|---|---|---|---|---|
| Patients (n) | 74 | 60 | 157 | 567 |
| Male | 28 | 43 | 347 | |
| Female | 46 | 17 | 220 | |
| Age (years) | 46 (18–82) | 48 (33–70) | 58 (10–89) | |
| Grade (n) | ||||
| I | ||||
| II | 45 | |||
| III | 24 | 31 | ||
| IV | 50 | 60 | 81 | 567 |
| Temozolomide (TMZ) (n) | ||||
| Yes | 166 | |||
| No | 401 | |||
| Radiotherapy Z (n) | ||||
| Yes | 422 | |||
| No | 145 |
| Gene | Sequence | Base |
|---|---|---|
| PPIC | F: AGCAAGTTTCATCGTGTCATCA | 22 |
| R: TGGAAATGTCTCACCATAGATGC | 23 | |
| EMP3 | F: GGAGGTCTCTTCTATGCCACC | 21 |
| R: AGGATCTCCTCGGCGTGAAT | 20 | |
| CHI3L1 | F: GTGAAGGCGTCTCAAACAGG | 20 |
| R: GAAGCGGTCAAGGGCATCT | 19 | |
| GAPDH | F: GAGTCAACGGATTTGGTCGT | 20 |
| R: TTGATTTTGGAGGGATCTCG | 20 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.-F.; Zhu, T.; Mao, C.-X.; Liu, Z.-X.; Wang, Z.-B.; Mao, X.-Y.; Li, L.; Yin, J.-Y.; Zhou, H.-H.; Liu, Z.-Q. PPIC, EMP3 and CHI3L1 Are Novel Prognostic Markers for High Grade Glioma. Int. J. Mol. Sci. 2016, 17, 1808. https://doi.org/10.3390/ijms17111808
Gao Y-F, Zhu T, Mao C-X, Liu Z-X, Wang Z-B, Mao X-Y, Li L, Yin J-Y, Zhou H-H, Liu Z-Q. PPIC, EMP3 and CHI3L1 Are Novel Prognostic Markers for High Grade Glioma. International Journal of Molecular Sciences. 2016; 17(11):1808. https://doi.org/10.3390/ijms17111808
Chicago/Turabian StyleGao, Yuan-Feng, Tao Zhu, Chen-Xue Mao, Zhi-Xiong Liu, Zhi-Bin Wang, Xiao-Yuan Mao, Ling Li, Ji-Ye Yin, Hong-Hao Zhou, and Zhao-Qian Liu. 2016. "PPIC, EMP3 and CHI3L1 Are Novel Prognostic Markers for High Grade Glioma" International Journal of Molecular Sciences 17, no. 11: 1808. https://doi.org/10.3390/ijms17111808