
Low Expression of CAPON in Glioma Contributes to Cell Proliferation via the Akt Signaling Pathway

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
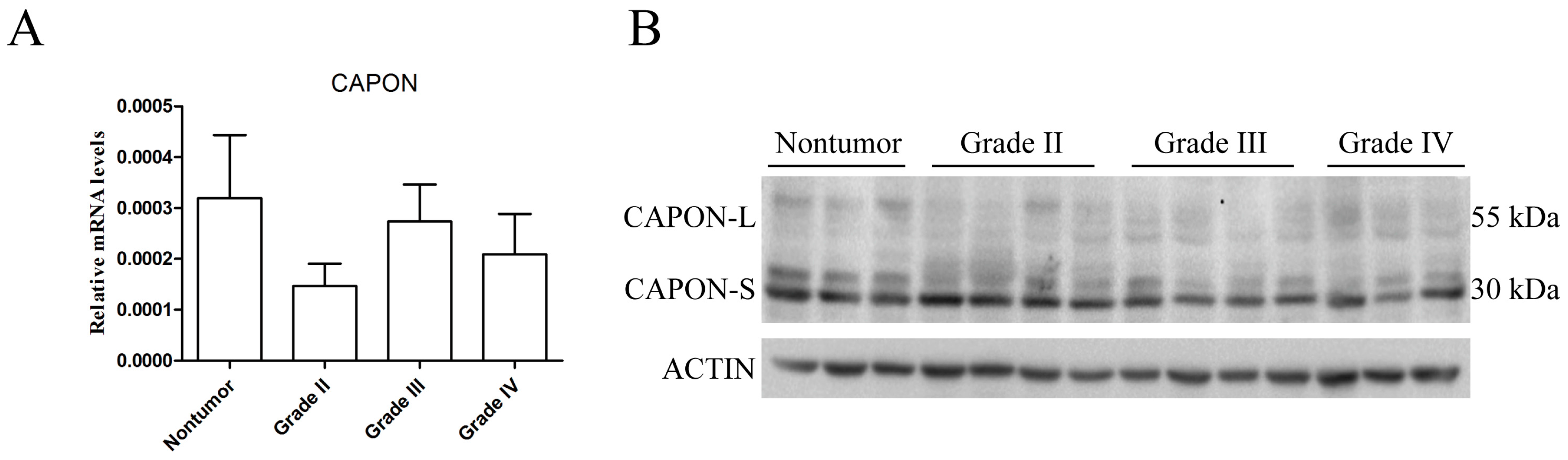
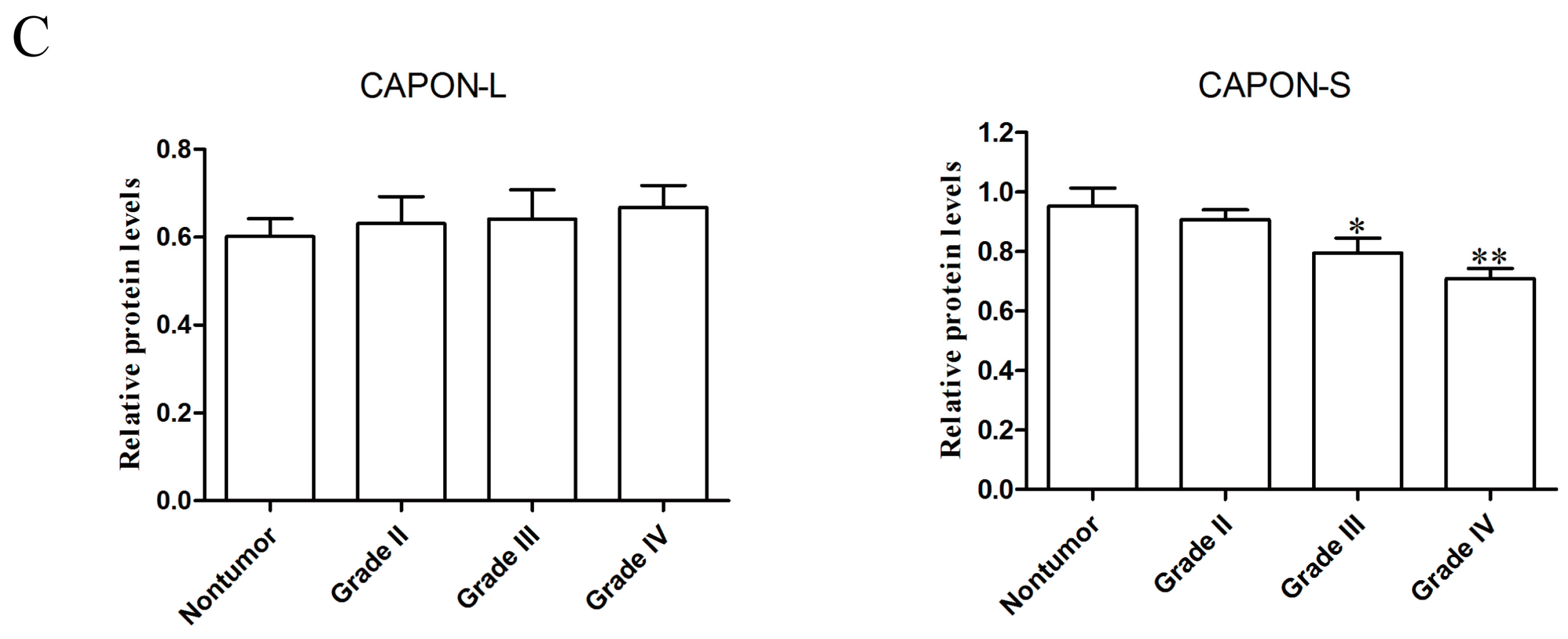
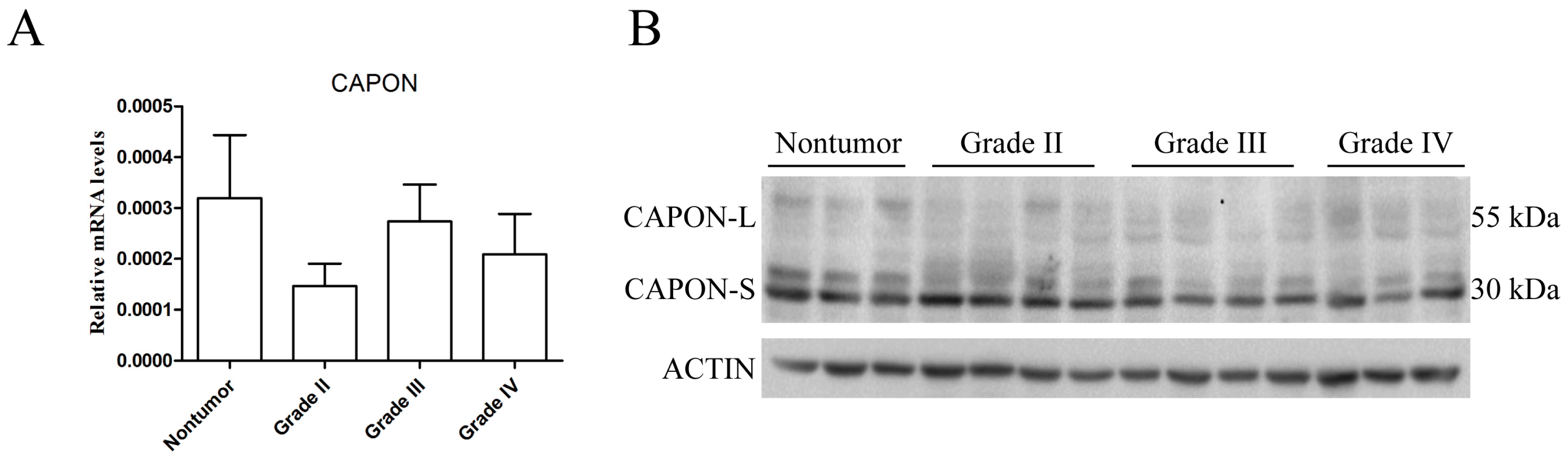
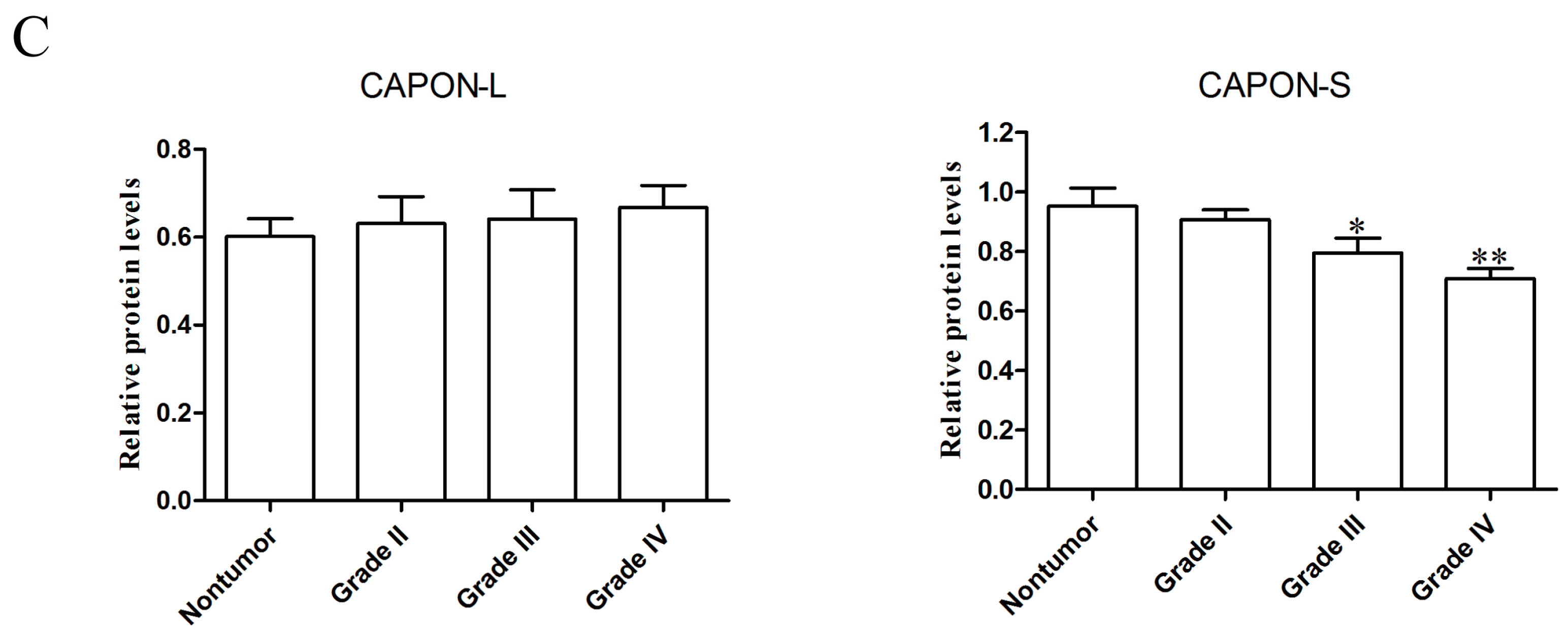
2.1. Expression Changes of CAPON in Human Glioma Tissues
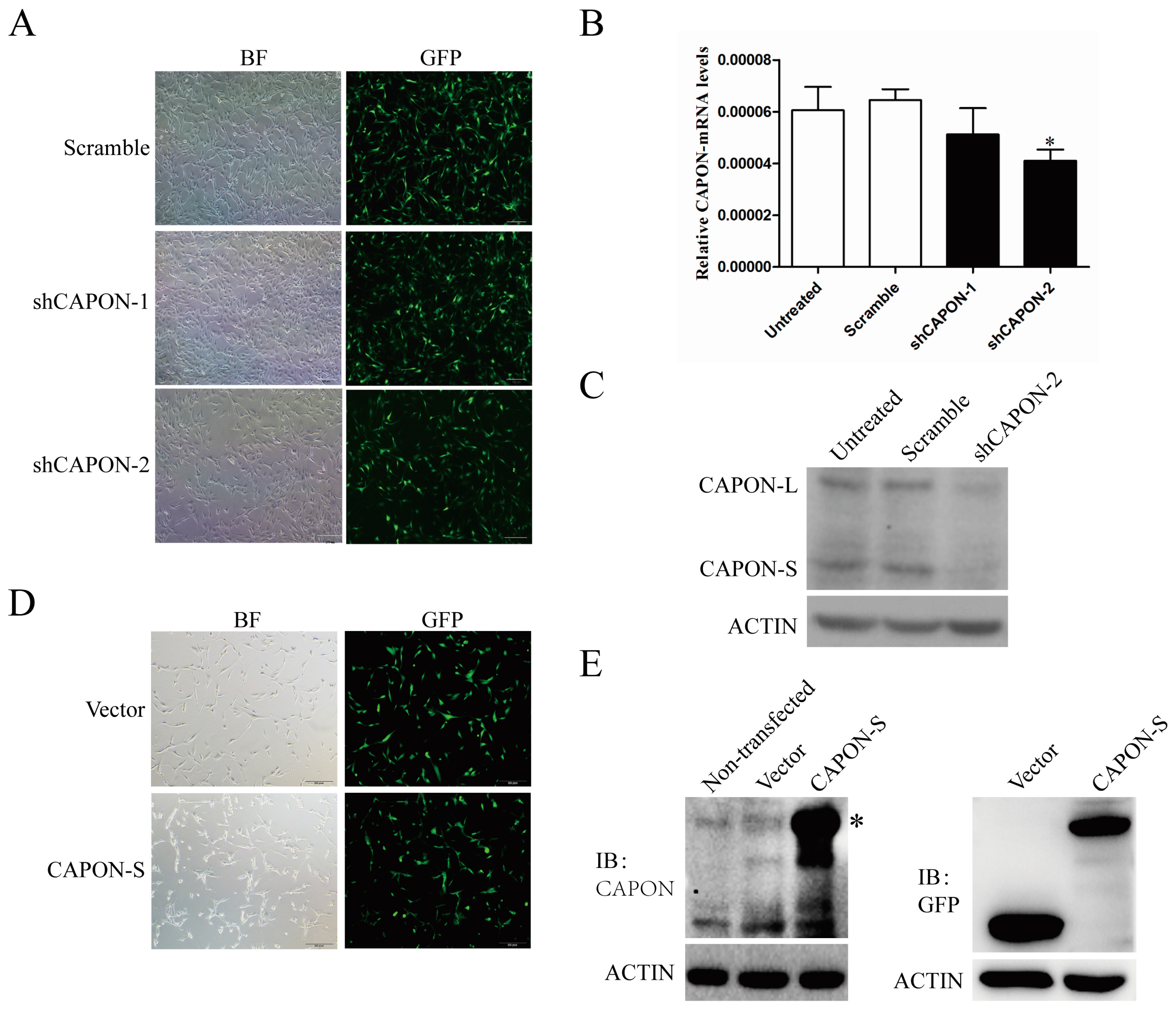
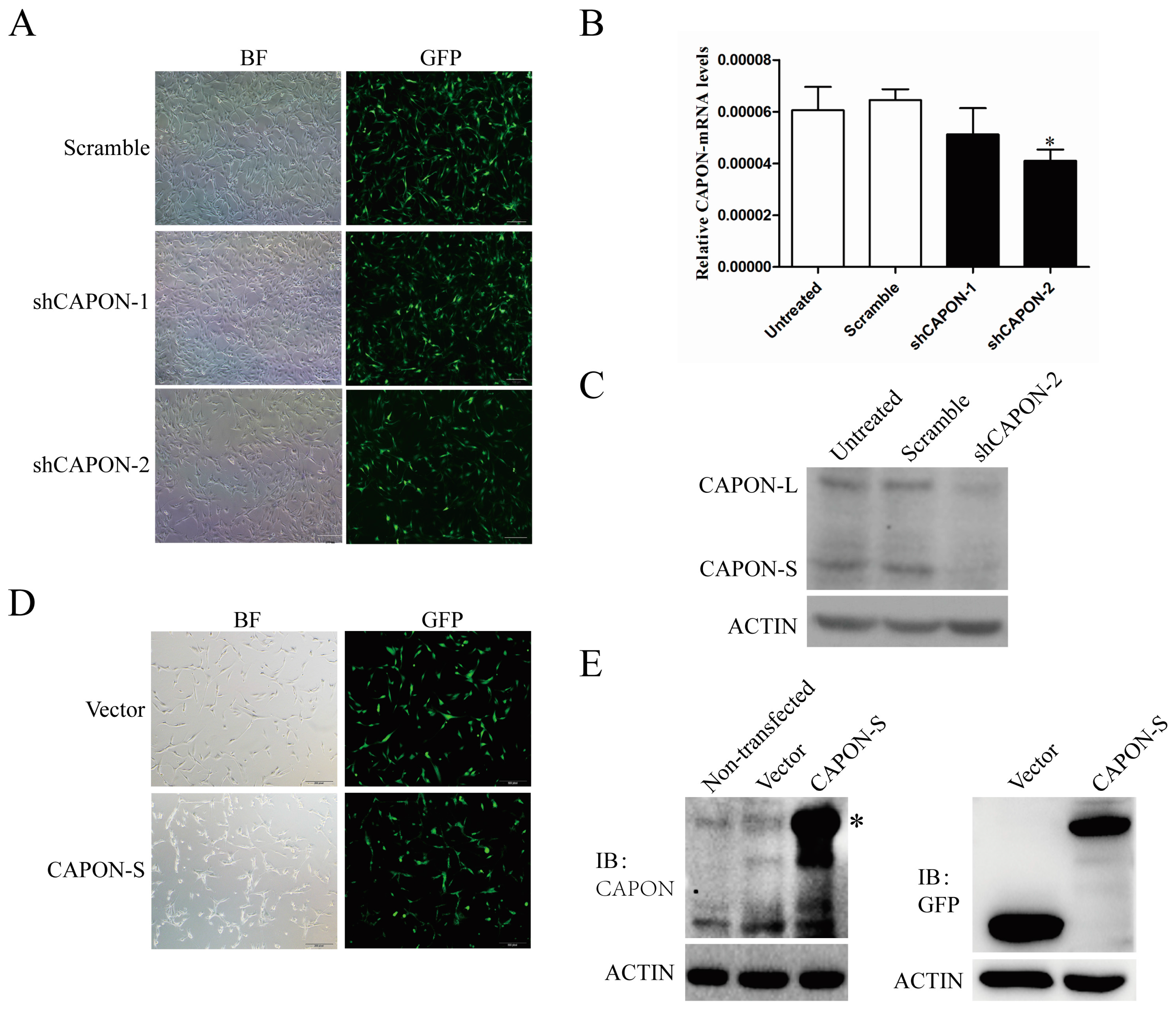
2.2. CAPON-S Overexpression Efficiency and CAPON Down-Regulation Efficiency in Glioma Cells
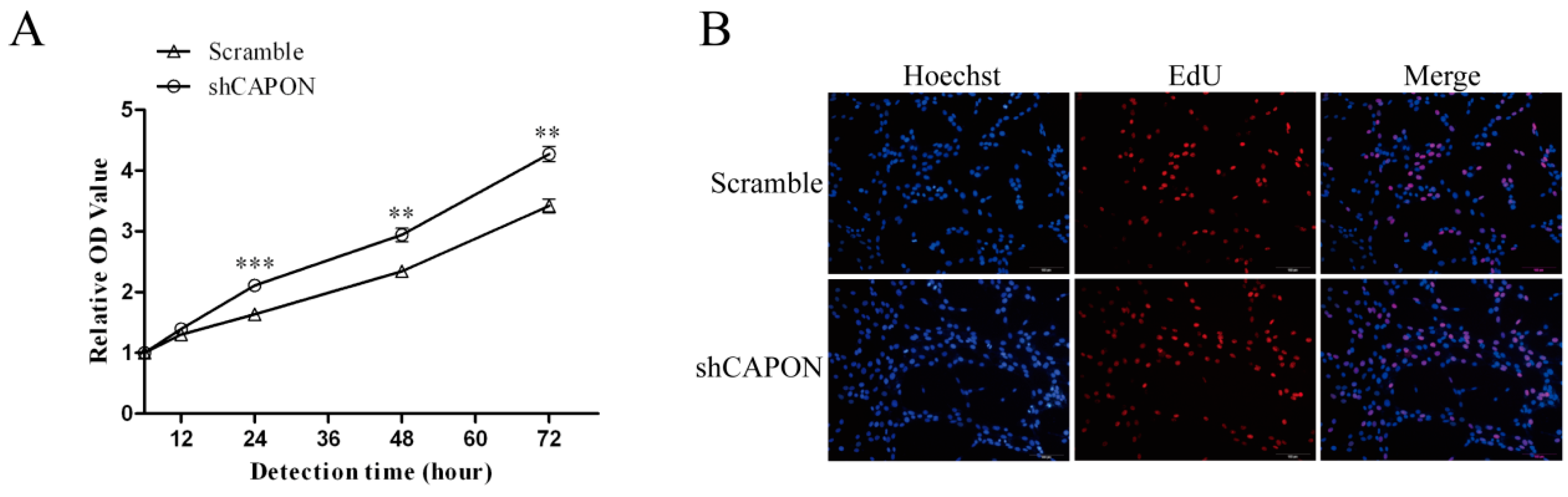
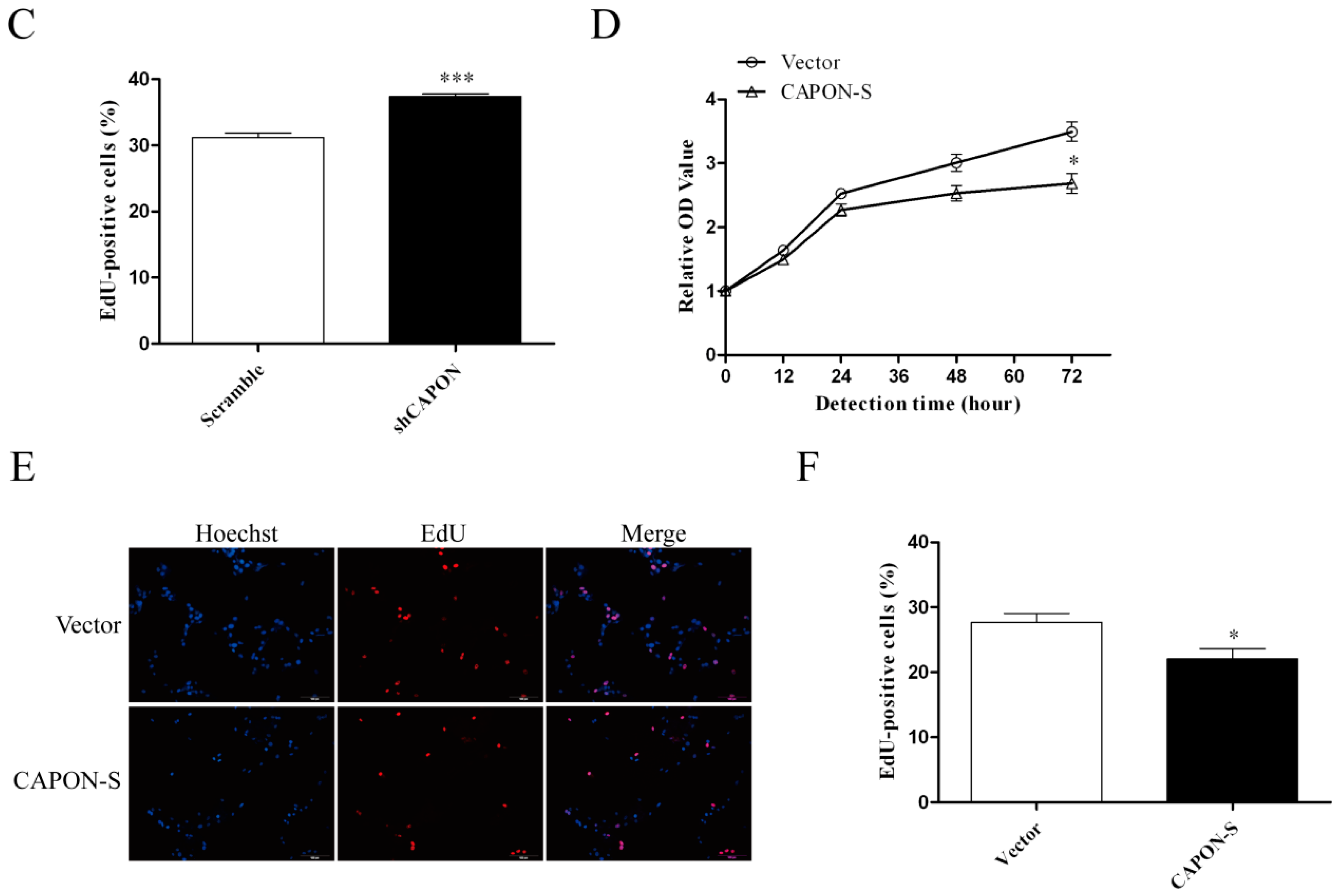
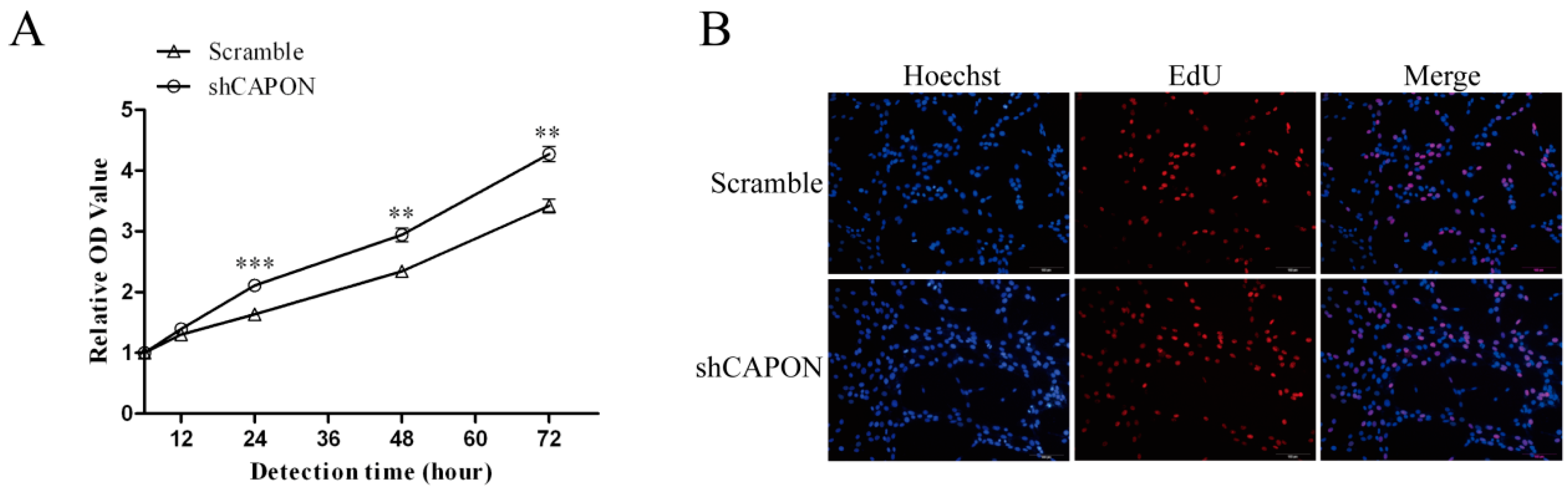
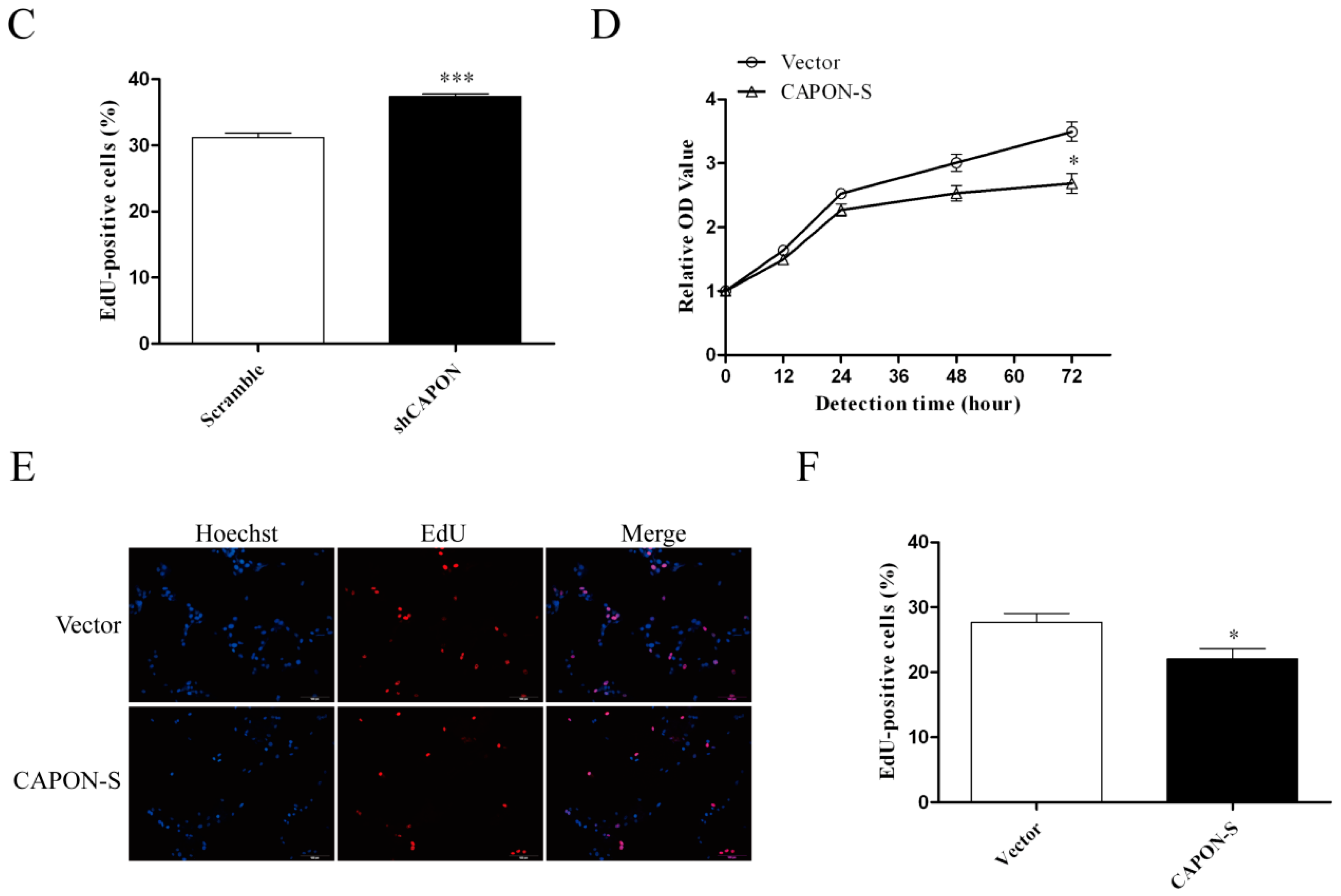
2.3. Effects of CAPON-S Overexpression or CAPON Down-Regulation on the Proliferation of Glioma Cells
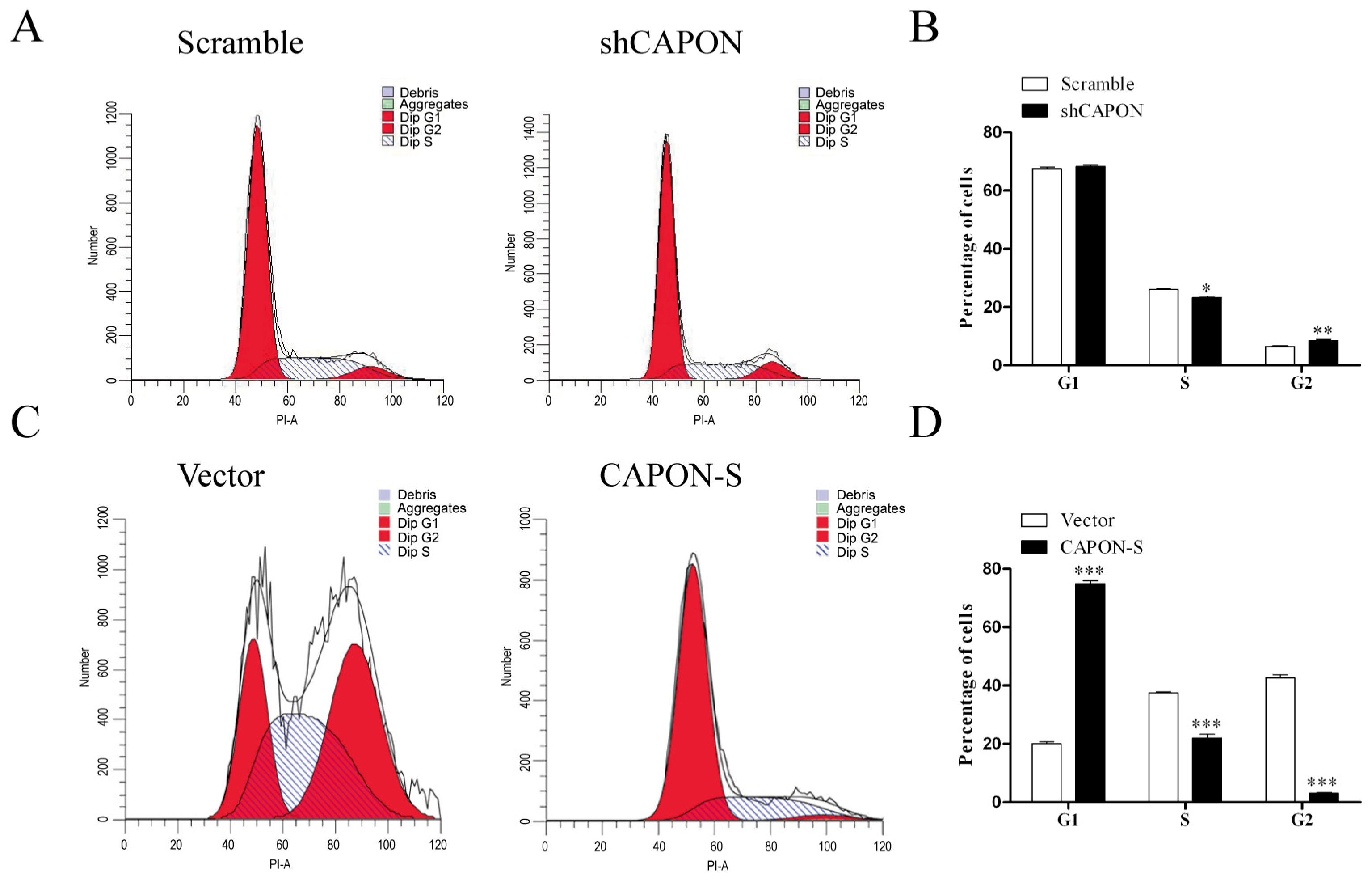
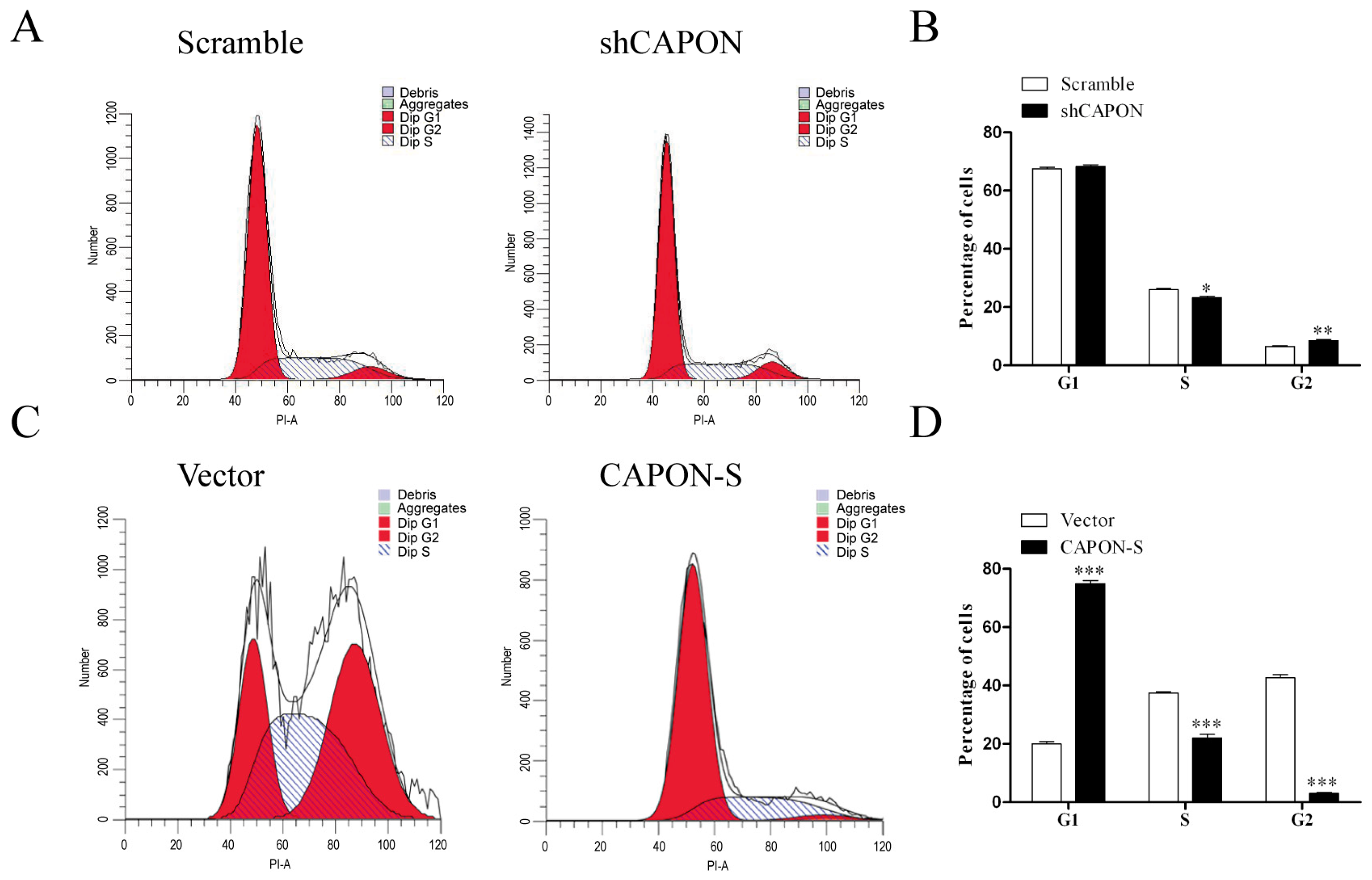
2.4. Effects of CAPON-S Overexpression or CAPON Down-Regulation on the Cell Cycle Progression of Glioma Cells
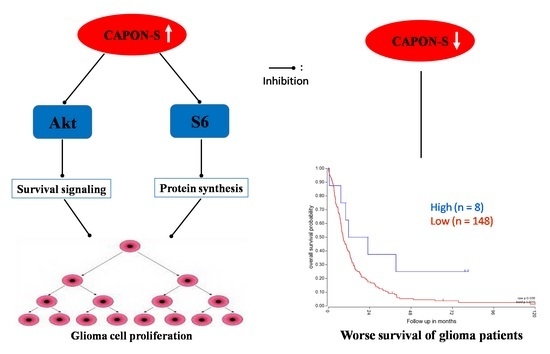
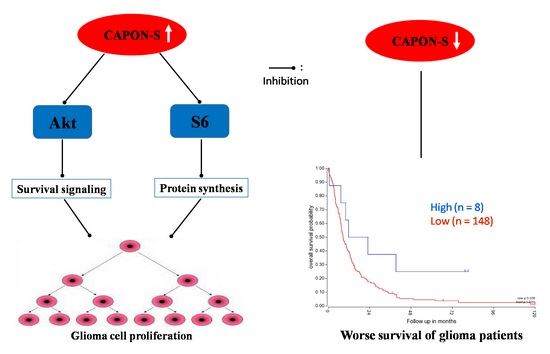
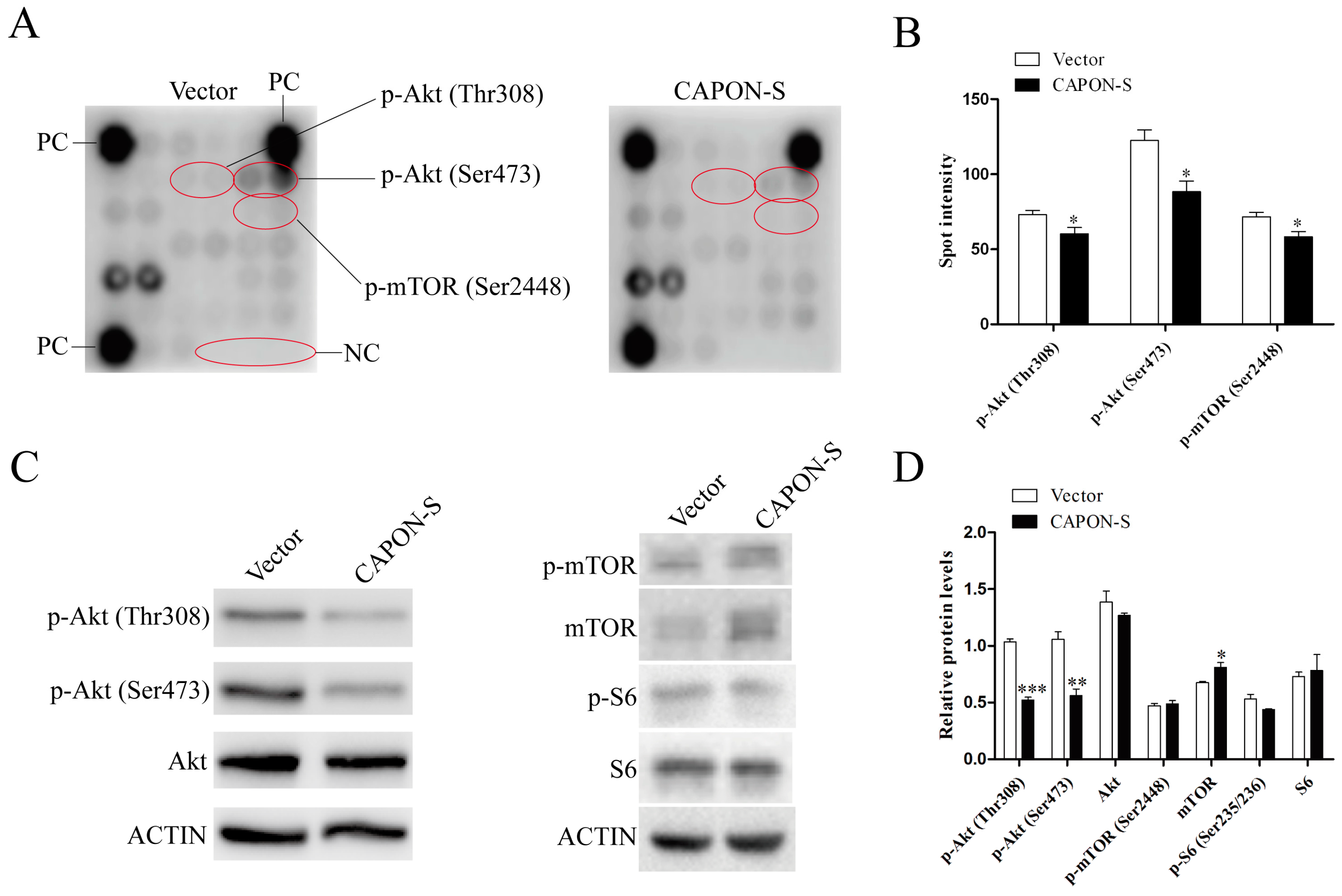
2.5. Effects of CAPON-S Overexpression on the Important Intracellular Signaling Molecules in Glioma Cells
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Glioma and Nontumorous Human Brain Tissues
4.3. Antibodies
4.4. RNA Extraction and Quantitative Real Time-PCR (qRT-PCR)
4.5. Western Blot Analysis
4.6. Lentivirus Construction, Production, and Infection
4.7. Cell Viability Detection
4.8. EdU Assay
4.9. Flow Cytometry
4.10. Antibody Array
4.11. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.G.; Candolfi, M.; Kroeger, K.M.; King, G.D.; Curtin, J.F.; Yagiz, K.; Mineharu, Y.; Assi, H.; Wibowo, M.; Muhammad, A.K.M.G.; et al. Gene therapy and targeted toxins for glioma. Curr. Gene Ther. 2011, 11, 155–180. [Google Scholar] [CrossRef] [PubMed]
- Bourkoula, E.; Mangoni, D.; Ius, T.; Pucer, A.; Isola, M.; Musiello, D.; Marzinotto, S.; Toffoletto, B.; Sorrentino, M.; Palma, A.; et al. Glioma-associated stem cells: A novel class of tumor-supporting cells able to predict prognosis of human low-grade gliomas. Stem Cells 2014, 32, 1239–1253. [Google Scholar] [PubMed]
- Cheng, L.; Huang, Z.; Zhou, W.; Wu, Q.; Donnola, S.; Liu, J.K.; Fang, X.; Sloan, A.E.; Mao, Y.; Lathia, J.D.; et al. Glioblastoma stem cells generate vascular pericytes to support vessel function and tumor growth. Cell 2013, 153, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.B.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Massion, P.B.; Feron, O.; Dessy, C.; Balligand, J.L. Nitric oxide and cardiac function: Ten years after, and continuing. Circ. Res. 2003, 93, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Rand, M.J.; Li, C.G. Nitric oxide as a neurotransmitter in peripheral nerves: Nature of transmitter and mechanism of transmission. Annu. Rev. Physiol. 1995, 57, 659–682. [Google Scholar] [CrossRef] [PubMed]
- Lala, P.K.; Chakraborty, C. Role of nitric oxide in carcinogenesis and tumour progression. Lancet Oncol. 2001, 2, 149–156. [Google Scholar] [CrossRef]
- Torre, D.; Pugliese, A.; Speranza, F. Role of nitric oxide in HIV-1 infection: Friend or foe? Lancet Infect. Dis. 2002, 2, 273–280. [Google Scholar] [CrossRef]
- Griffith, O.W.; Stuehr, D.J. Nitric oxide synthases: Properties and catalytic mechanism. Annu. Rev. Physiol. 1995, 57, 707–736. [Google Scholar] [CrossRef] [PubMed]
- Jaffrey, S.R.; Snowman, A.M.; Eliasson, M.J.; Cohen, N.A.; Snyder, S.H. CAPON: A protein associated with neuronal nitric oxide synthase that regulates its interactions with PSD95. Neuron 1998, 20, 115–124. [Google Scholar] [CrossRef]
- Arking, D.E.; Pfeufer, A.; Post, W.; Kao, W.H.; Newton-Cheh, C.; Ikeda, M.; West, K.; Kashuk, C.; Akyol, M.; Perz, S.; et al. A common genetic variant in the NOS1 regulator NOS1AP modulates cardiac repolarization. Nat. Genet. 2006, 38, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Segalat, L.; Grisoni, K.; Archer, J.; Vargas, C.; Bertrand, A.; Anderson, J.E. CAPON expression in skeletal muscle is regulated by position, repair, NOS activity, and dystrophy. Exp. Cell Res. 2005, 302, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Wang, C.; Zhang, R.; Ng, M.C.; Bao, Y.; Wang, C.; So, W.Y.; Ma, R.C.; Ma, X.; Chan, J.C.; et al. Association of genetic variants of NOS1AP with type 2 diabetes in a chinese population. Diabetologia 2010, 53, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Wratten, N.; Charych, E.I.; Buyske, S.; Firestein, B.L.; Brzustowicz, L.M. Increased expression in dorsolateral prefrontal cortex of CAPON in schizophrenia and bipolar disorder. PLoS Med. 2005, 2, e263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Jin, L.; Zhu, Y.; Zhou, X.; Yu, R.; Gao, S. Research progress in NOS1AP in neurological and psychiatric diseases. Brain Res. Bull. 2016, 125, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Clattenburg, L.; Wigerius, M.; Qi, J.; Rainey, J.K.; Rourke, J.L.; Muruganandan, S.; Sinal, C.J.; Fawcett, J.P. NOS1AP functionally associates with YAP to regulate Hippo signaling. Mol. Cell. Biol. 2015, 35, 2265–2277. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, S.I.; Iden, S.; Collard, J.G. Cell polarity proteins and cancer. Semin. Cancer Biol. 2012, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.; Sun, J.; Lau, A.; Curtis, S.; Goldsmith, J.; Fox, V.L.; Wei, C.; Frazier, M.; Samson, O.; Wong, K.K.; et al. A genetic screen identifies an LKB1-MARK signalling axis controlling the Hippo-YAP pathway. Nat. Cell Biol. 2014, 16, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Humbert, P.O.; Grzeschik, N.A.; Brumby, A.M.; Galea, R.; Elsum, I.; Richardson, H.E. Control of tumourigenesis by the Scribble/Dlg/Lgl polarity module. Oncogene 2008, 27, 6888–6907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Qi, H.X.; Hu, Z.M.; Chang, Y.N.; Shi, Z.M.; Han, X.H.; Han, Y.W.; Zhang, R.X.; Zhang, Z.; Chen, T.; et al. YAP and TAZ take center stage in cancer. Biochemistry 2015, 54, 6555–6566. [Google Scholar] [CrossRef] [PubMed]
- Anastas, J.N.; Biechele, T.L.; Robitaille, M.; Muster, J.; Allison, K.H.; Angers, S.; Moon, R.T. A protein complex of SCRIB, NOS1AP and VANGL1 regulates cell polarity and migration, and is associated with breast cancer progression. Oncogene 2012, 31, 3696–3708. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yan, M.; Lv, Q.; Cheng, C.; Li, X.; Guo, Z.; Tao, T.; Shen, A. Inhibition of nitric oxide-induced nuclear localization of CAPON by NMDA receptor antagonist in cultured rat primary astrocytes. Neurochem. Int. 2010, 56, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Ginet, V.; Liu, X.; Vergun, O.; Tuittila, M.; Mathieu, M.; Bonny, C.; Puyal, J.; Truttmann, A.C.; Courtney, M.J. The nNOS-p38MAPK pathway is mediated by nos1ap during neuronal death. J. Neurosci. 2013, 33, 8185–8201. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Melero-Fernandez de Mera, R.M.; Chen, J.; Ba, W.; Kasri, N.N.; Zhang, M.; Courtney, M.J. Unexpected heterodivalent recruitment of NOS1AP to nNOS reveals multiple sites for pharmacological intervention in neuronal disease models. J. Neurosci. 2015, 35, 7349–7364. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.F.; Qi, X.R.; Zhao, J.; Balesar, R.; Bao, A.M.; Swaab, D.F. Decreased NOS1 expression in the anterior cingulate cortex in depression. Cereb. Cortex 2013, 23, 2956–2964. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Gao, S.; Ben, Z.; Wang, H.; Jia, J.; Tao, T.; Niu, S.; Li, X.; Cheng, C. Identification and potential role of PSD-95 in Schwann cells. Neurol. Sci. 2008, 29, 321–330. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Wang, J.; Zhang, T.; Liu, G.; Jin, L.; Ji, D.; Wang, P.; Meng, Q.; Zhu, Y.; Yu, R. Low Expression of CAPON in Glioma Contributes to Cell Proliferation via the Akt Signaling Pathway. Int. J. Mol. Sci. 2016, 17, 1859. https://doi.org/10.3390/ijms17111859
Gao S, Wang J, Zhang T, Liu G, Jin L, Ji D, Wang P, Meng Q, Zhu Y, Yu R. Low Expression of CAPON in Glioma Contributes to Cell Proliferation via the Akt Signaling Pathway. International Journal of Molecular Sciences. 2016; 17(11):1859. https://doi.org/10.3390/ijms17111859
Chicago/Turabian StyleGao, Shangfeng, Jie Wang, Tong Zhang, Guangping Liu, Lei Jin, Daofei Ji, Peng Wang, Qingming Meng, Yufu Zhu, and Rutong Yu. 2016. "Low Expression of CAPON in Glioma Contributes to Cell Proliferation via the Akt Signaling Pathway" International Journal of Molecular Sciences 17, no. 11: 1859. https://doi.org/10.3390/ijms17111859