
Identification of Potential Biomarkers for Rhegmatogenous Retinal Detachment Associated with Choroidal Detachment by Vitreous iTRAQ-Based Proteomic Profiling

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Demographics and Clinical Variables
2.2. Protein Concentration
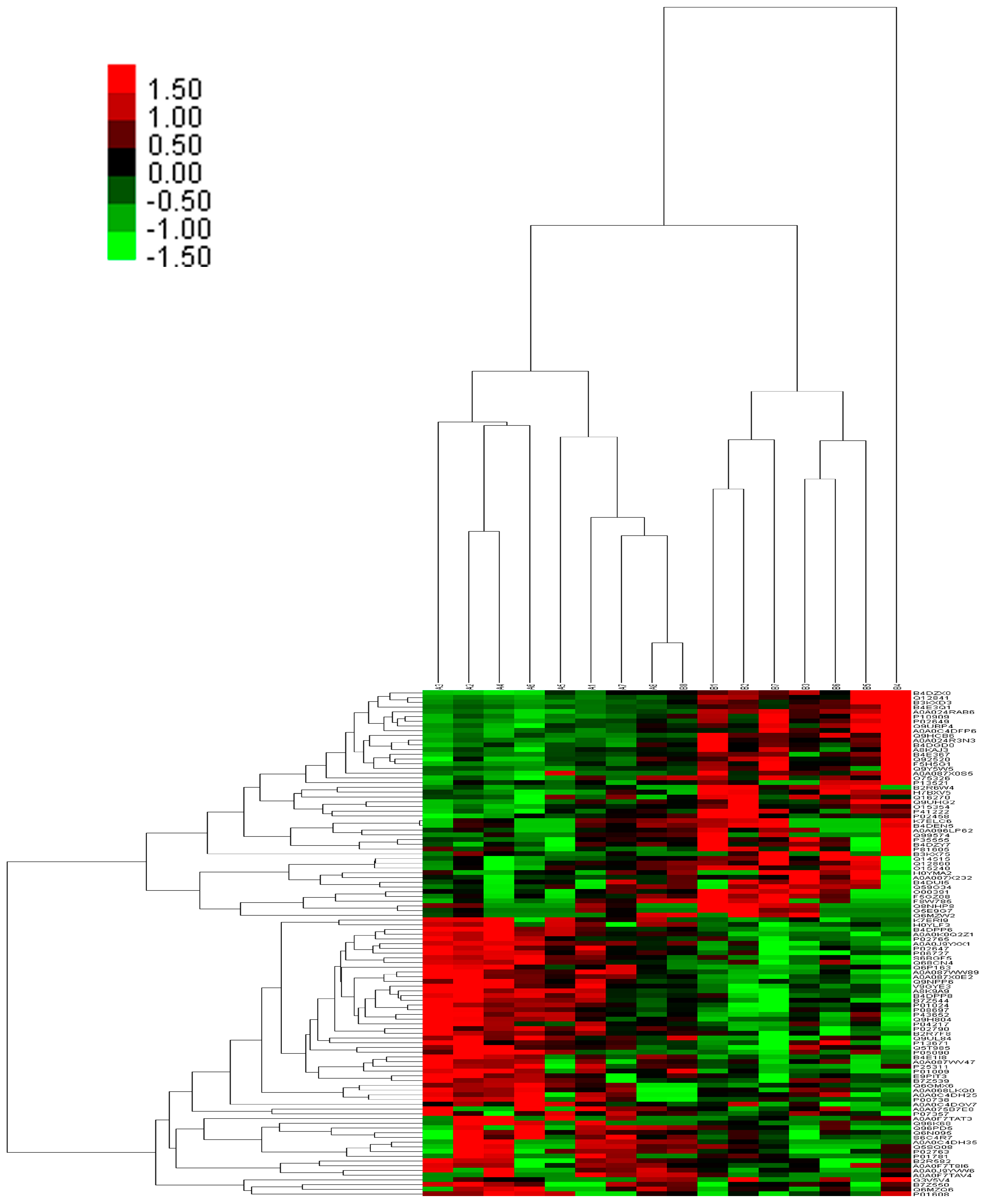
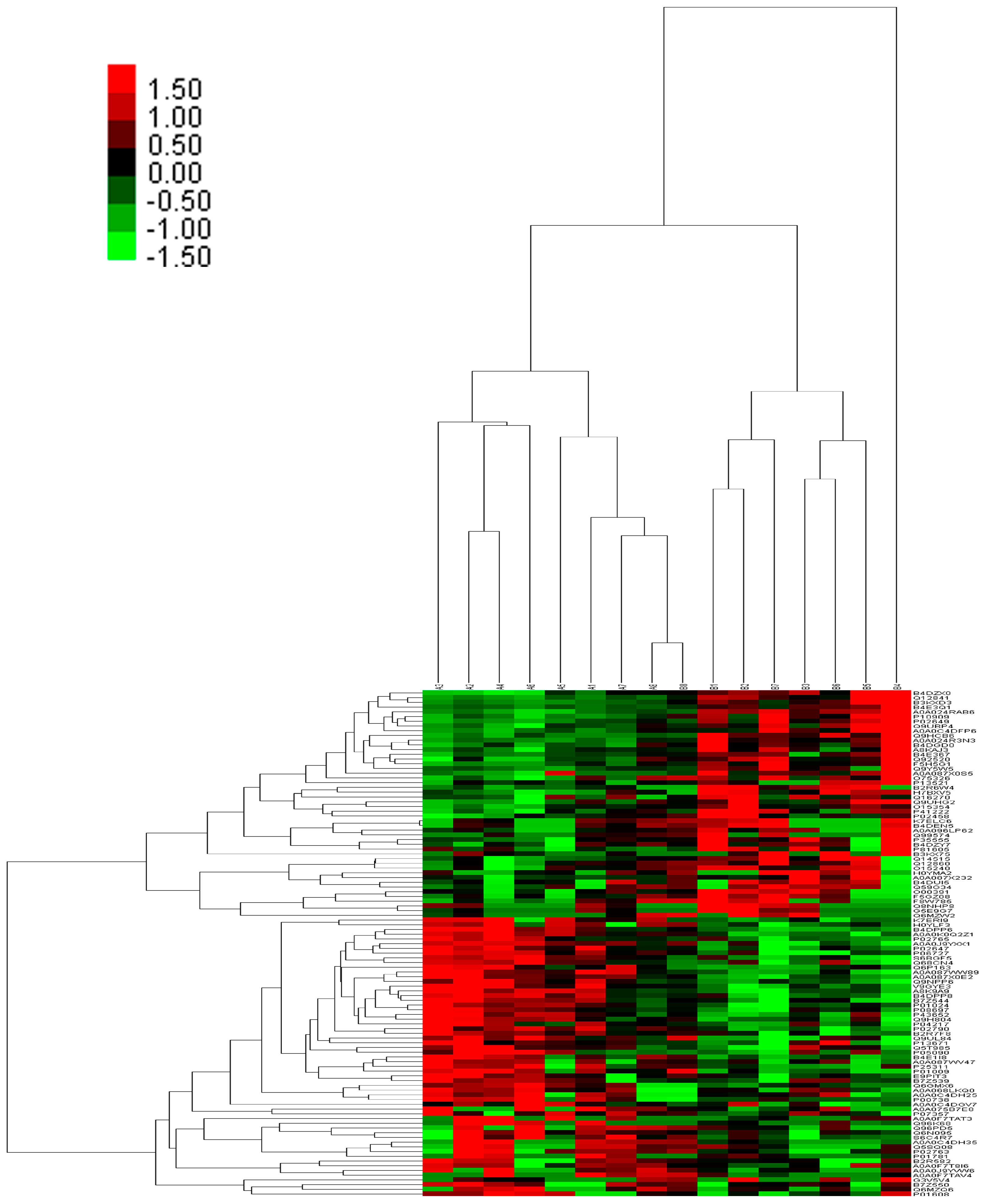
2.3. Integrated Proteome Information
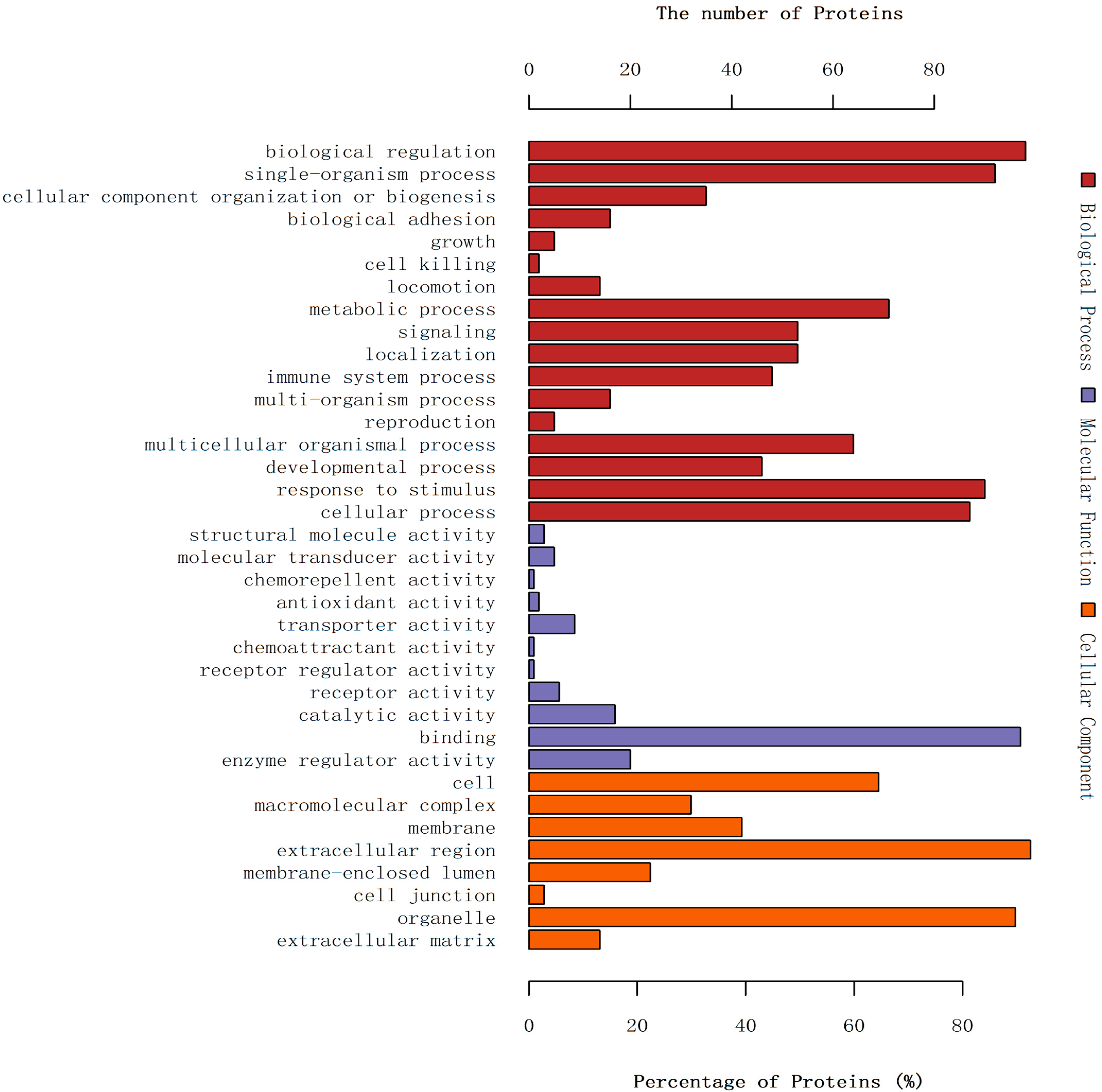
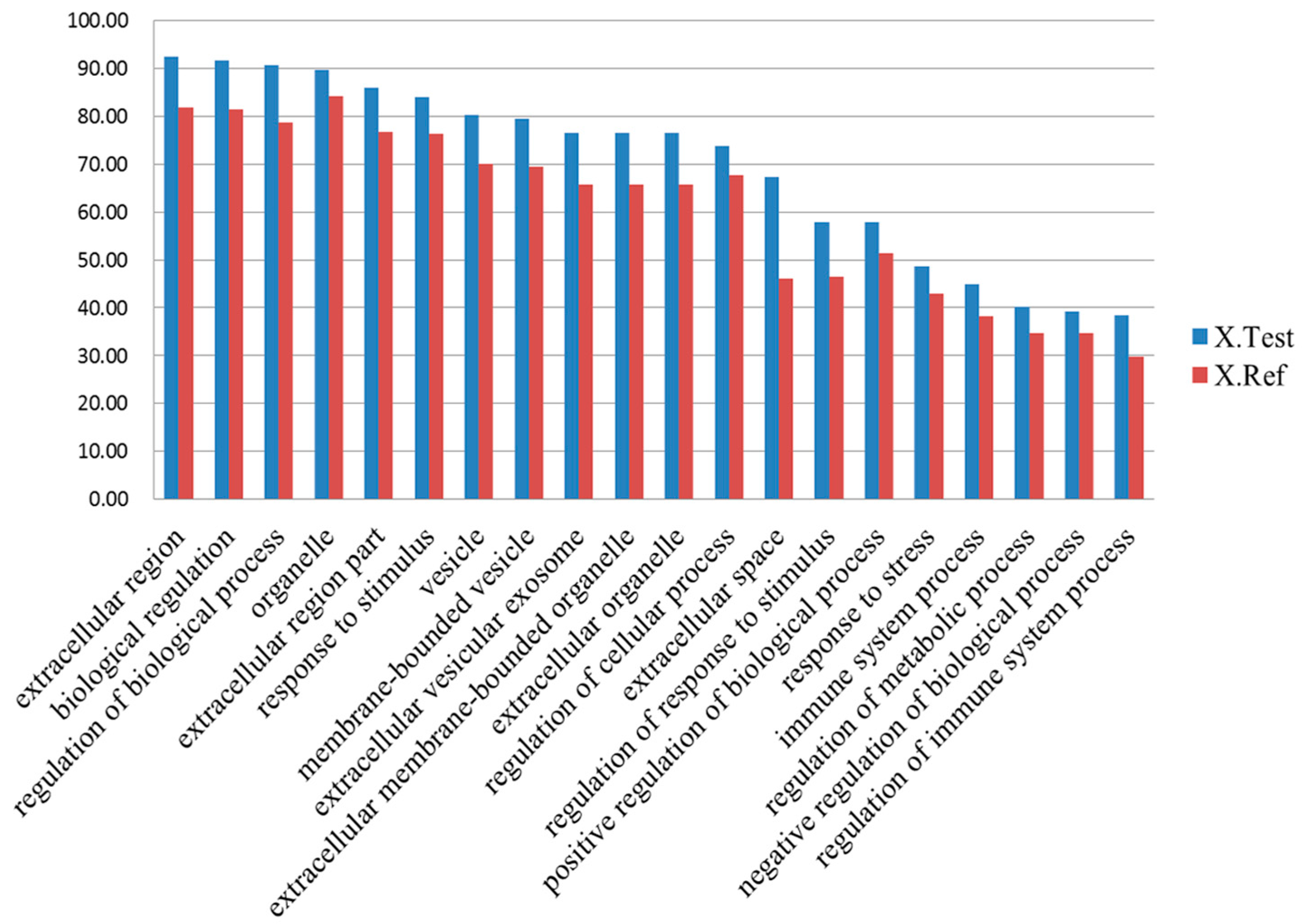
2.4. Gene Ontology Annotation of Differentially Expressed Proteins
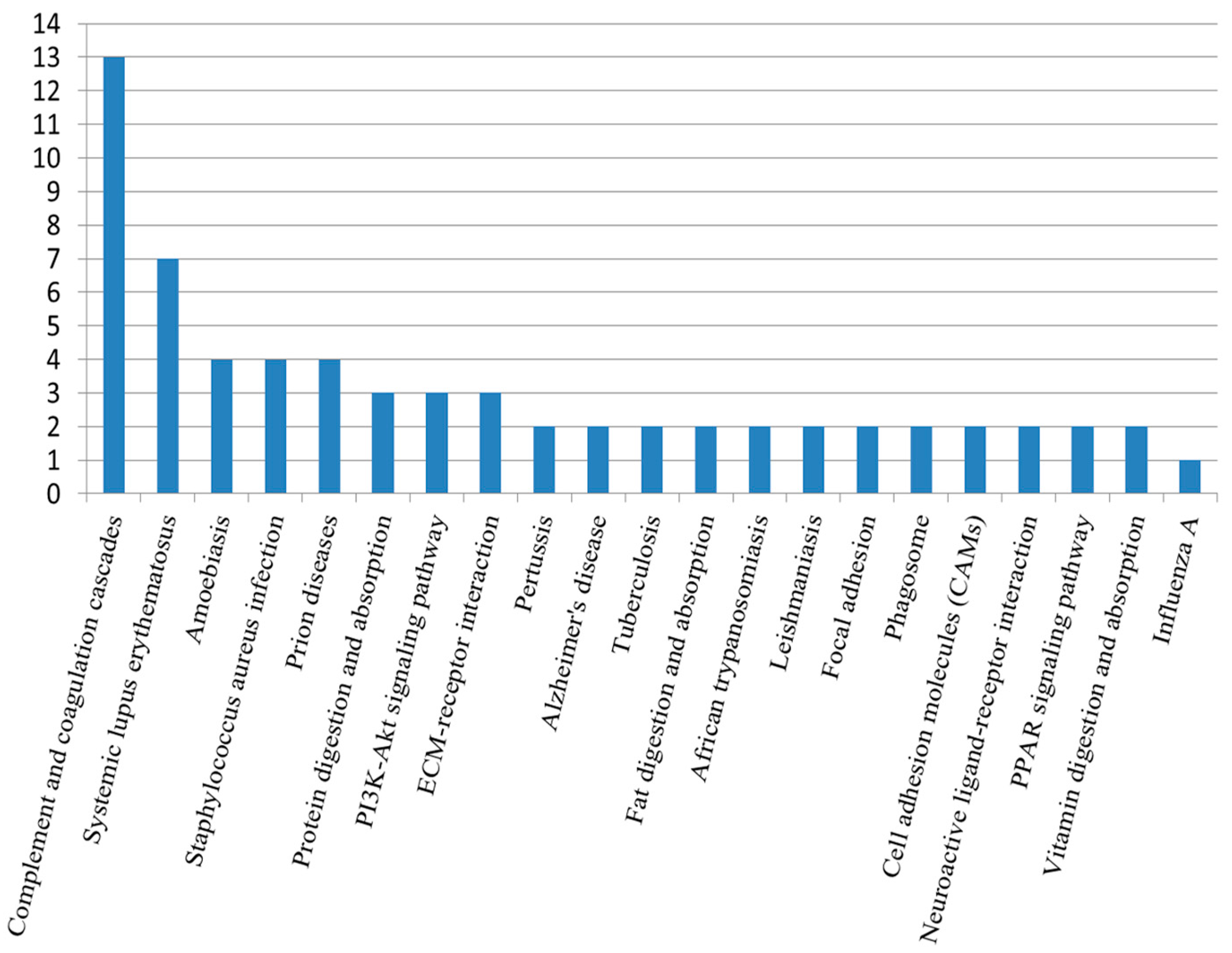
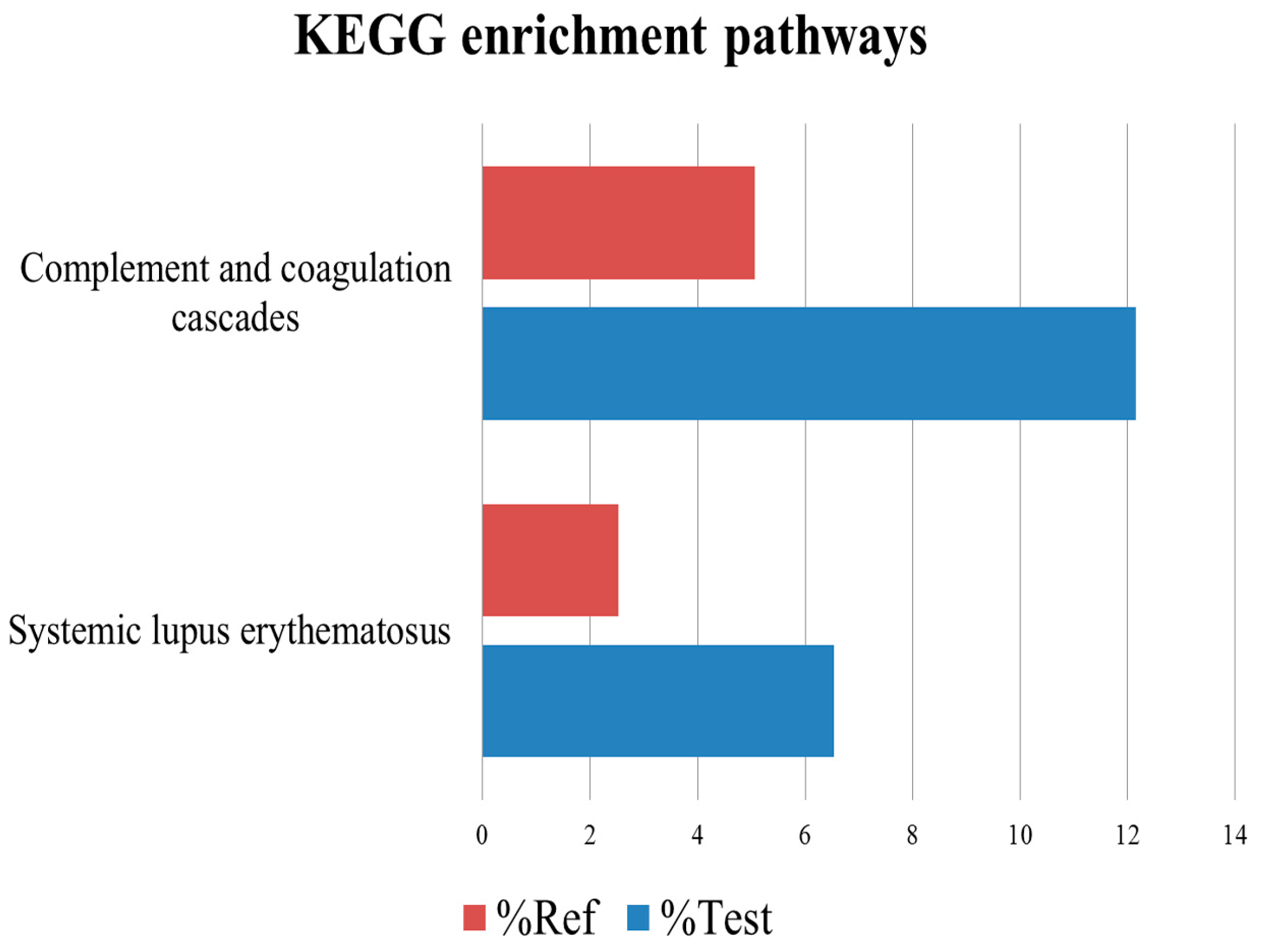
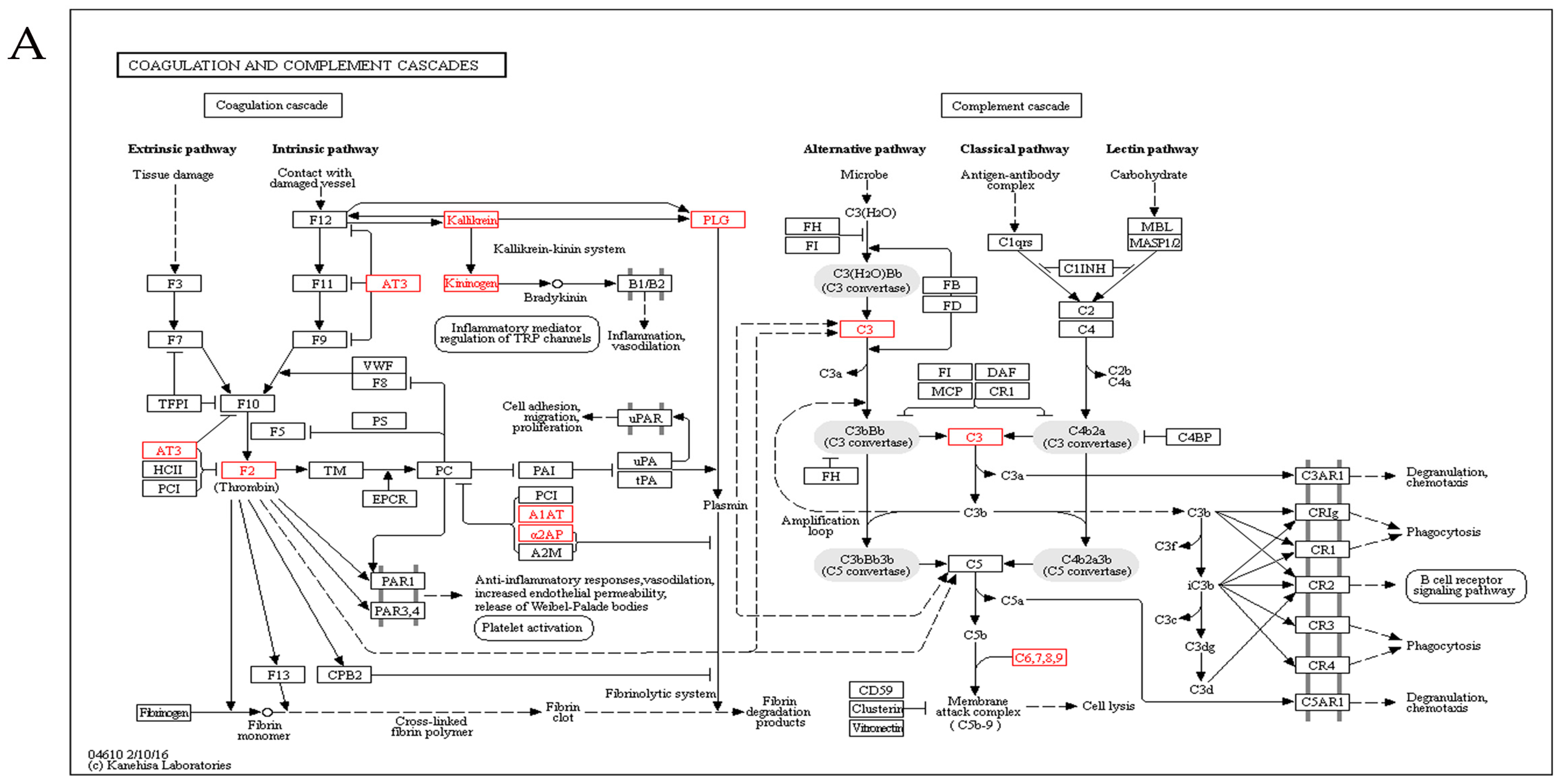
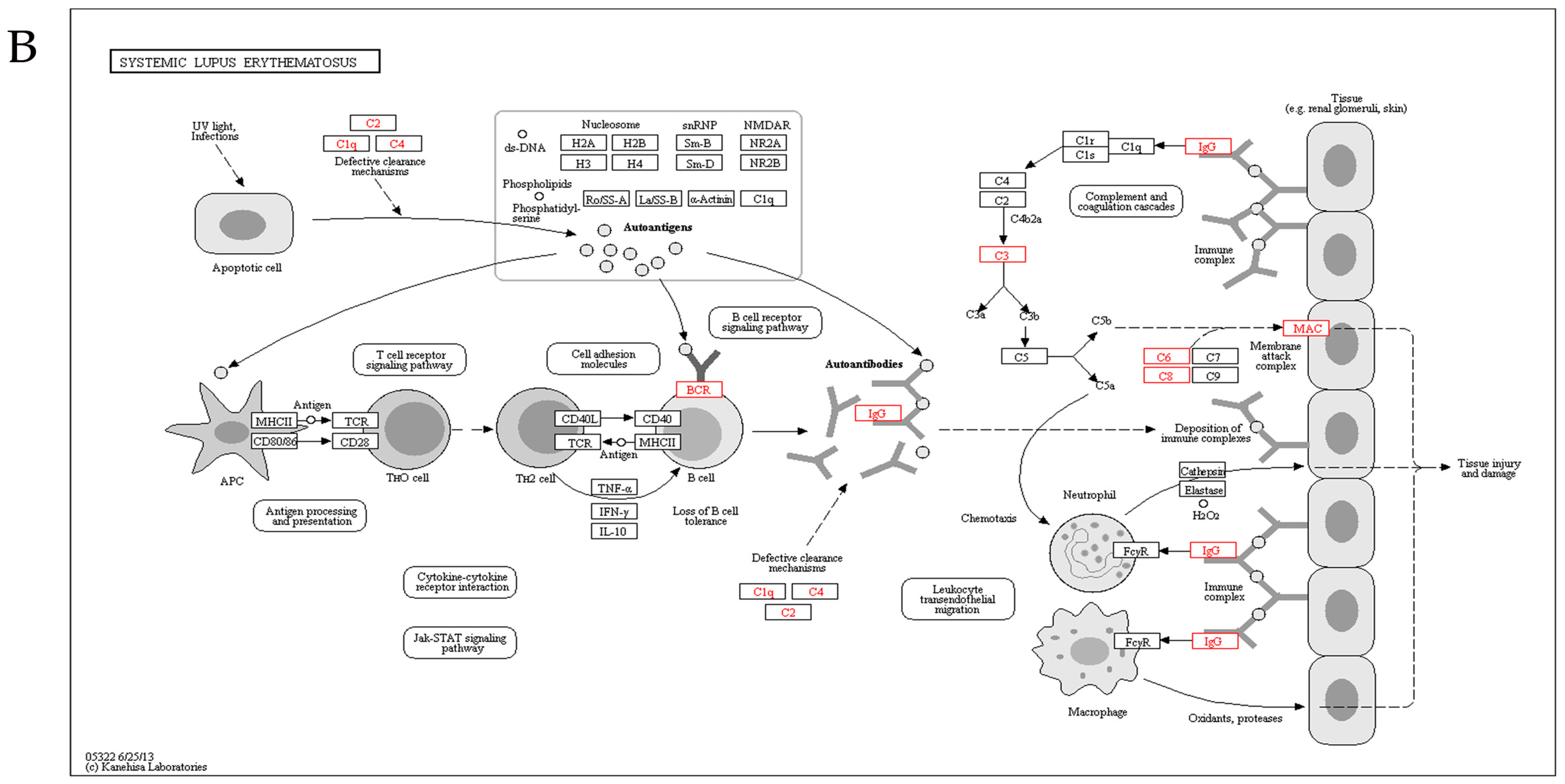
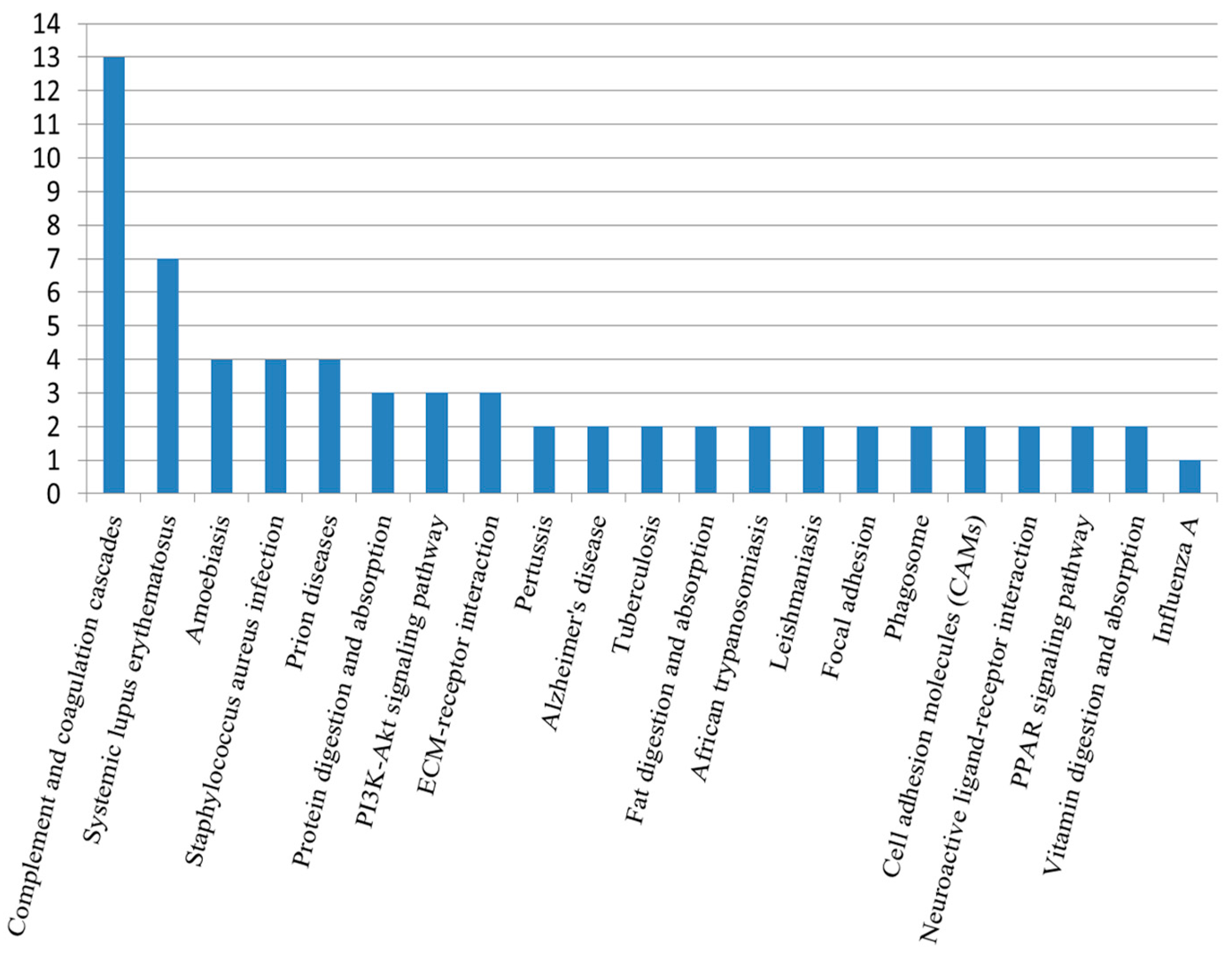
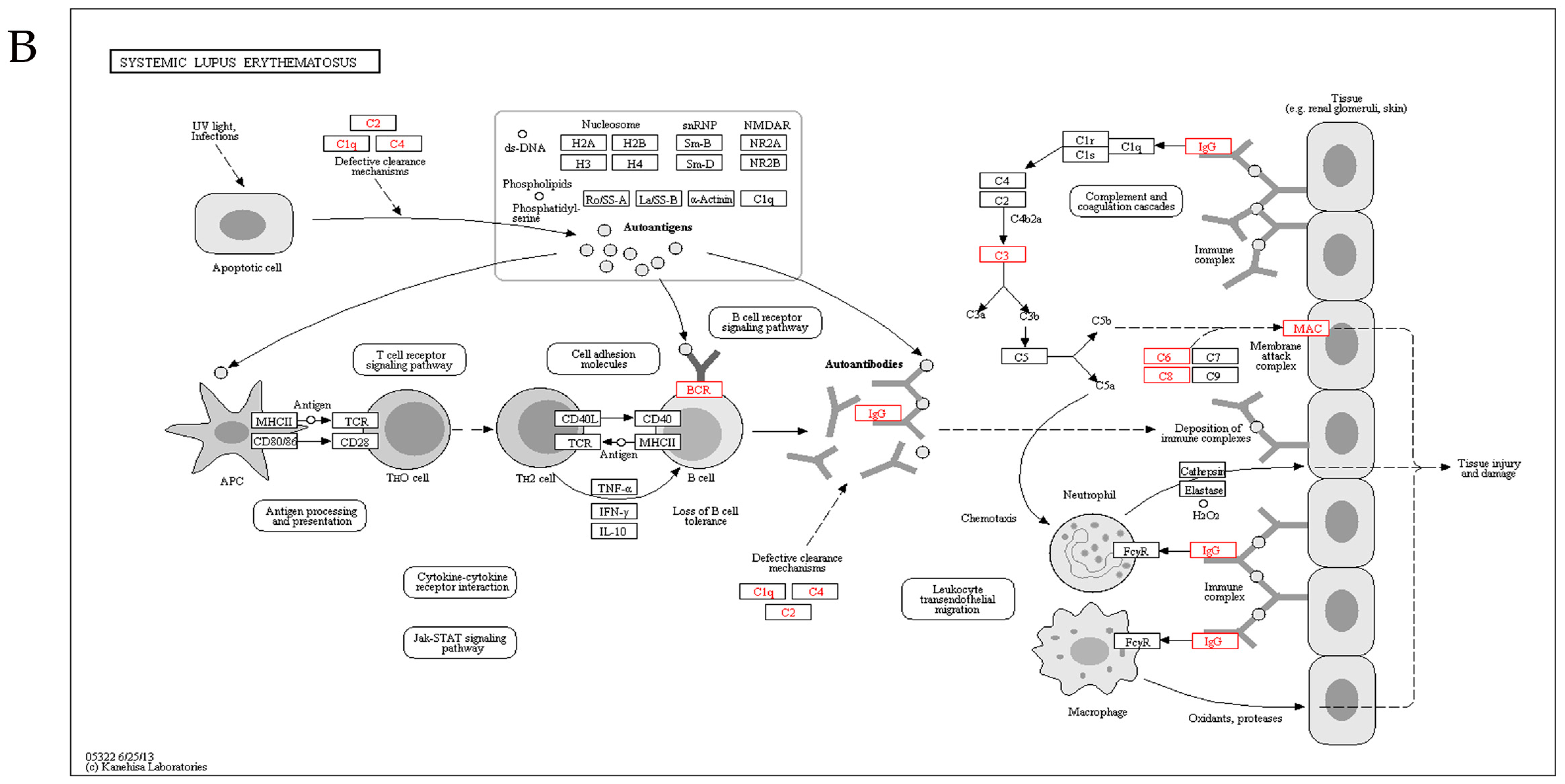
2.5. Kyoto Encyclopedia of Genes and Genomes Analysis
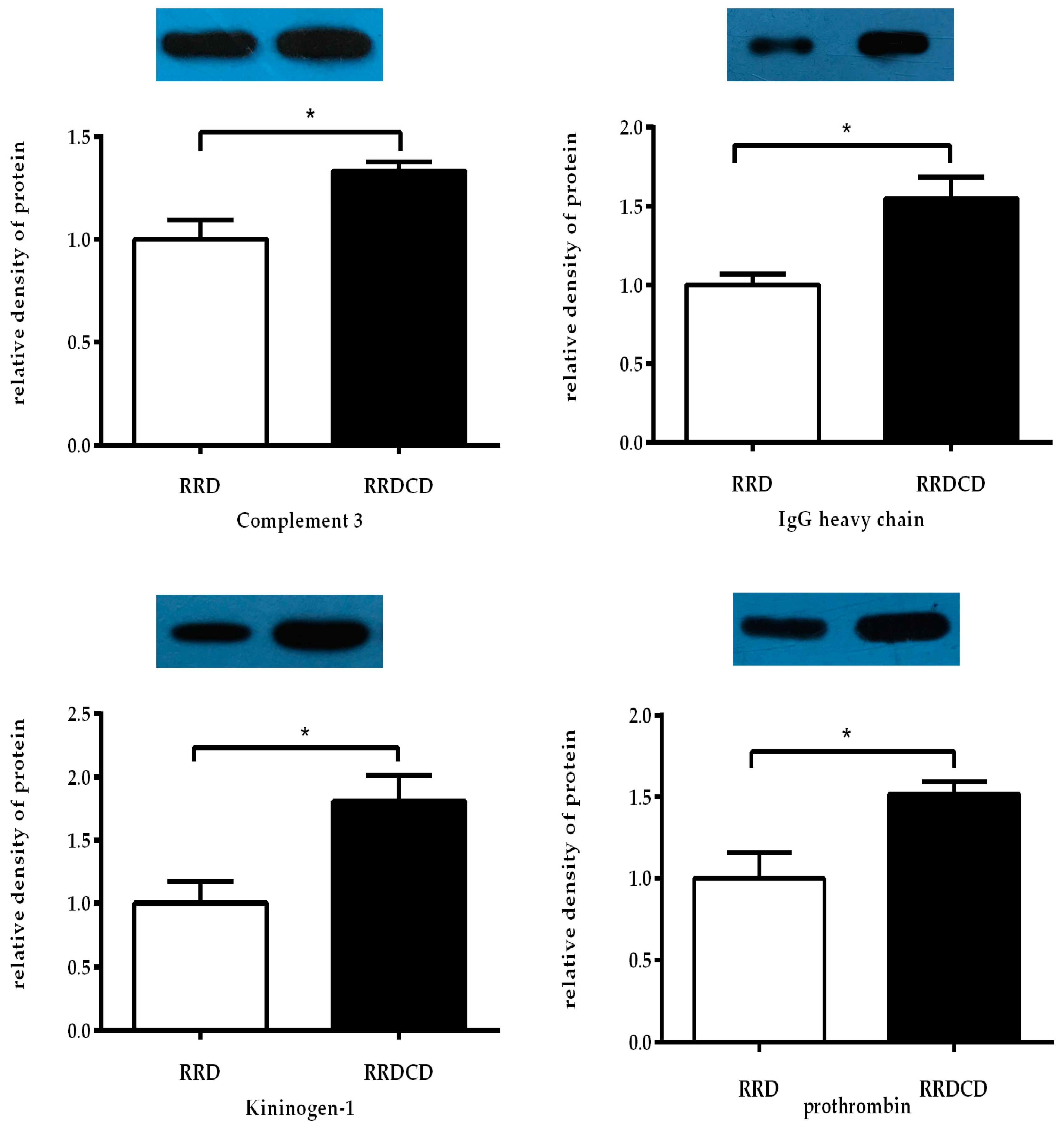
2.6. Protein Verification by Western Blotting Analysis
3. Discussion
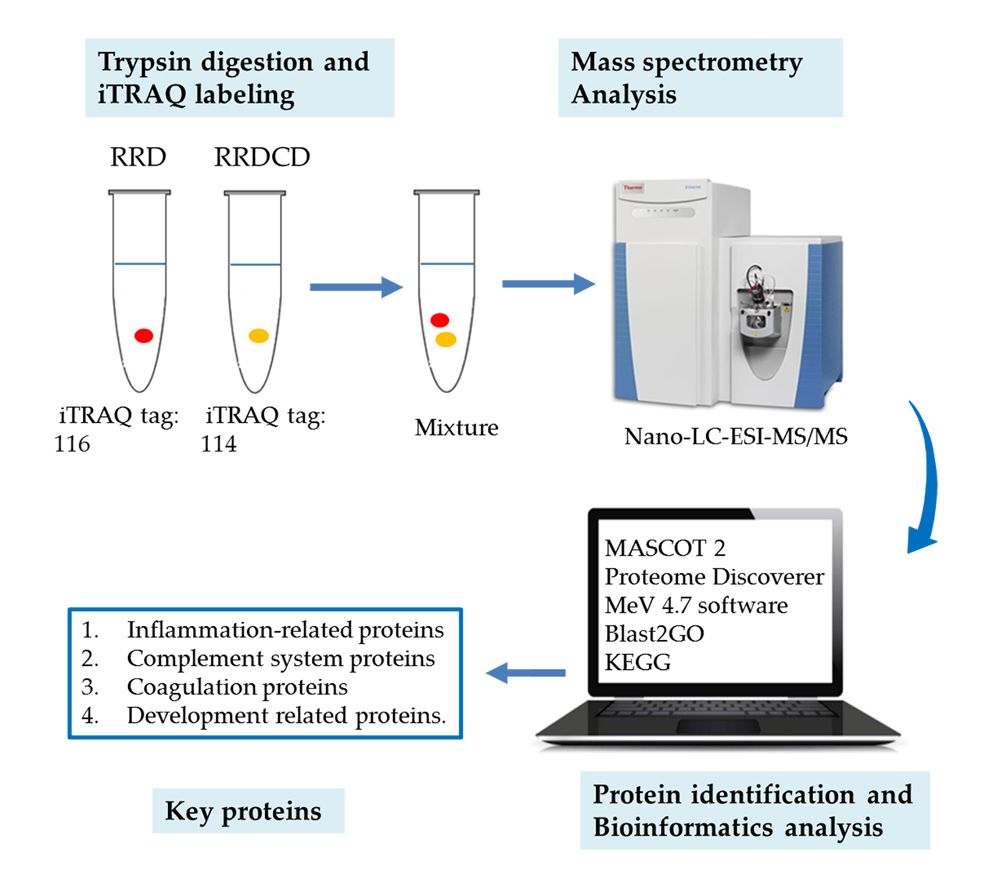
4. Materials and Methods
4.1. Patients
4.2. Ethics Statement
4.3. Sample Preparation
4.4. Protein Digestion and iTRAQ Labeling
4.5. Strong Cationic-Exchange (SCX) Chromatography Separation
4.6. Nano-LC-ESI-MS/MS Analysis
4.7. Protein Identification and Quantification
4.8. Bioinformatic Analysis
4.9. Western Blotting
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| BP | Biological process |
| BRB | Blood-retinal barrier |
| CC | Cellular component |
| GO | Gene ontology |
| iTRAQ | Isobaric tags for relative and absolute quantitation |
| KEGG | Kyoto encyclopedia of genes and genomes |
| MAC | Membrane attack complex |
| MF | Molecular function |
| Nano-LC-ESI-MS/MS | Nano-liquid chromatography-electrospray ion trap-mass spectrometry-mass spectrometry |
| PTGDS | Prostaglandin-H2 d-isomerase |
| PVR | Proliferative vitreoretinopathy |
| RRD | Rhegmatogenous retinal detachment |
| RRDCD | Rhegmatogenous retinal detachment associated with choroidal detachment |
| SLE | Systemic lupus erythematosus |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Coverage | Unique Peptides | Peptides | Fold Change | t Test p Value |
|---|---|---|---|---|---|---|
| K7ERI9 | Apolipoprotein C-I | 23.38 | 3 | 3 | 2.72074 | 6.32 × 10−5 |
| A0A087X0E2 | Serum amyloid A protein | 22.95 | 3 | 3 | 2.548367 | 0.000442 |
| A0A0F7TAV4 | IGHV5-51 protein (Fragment) | 16.36 | 2 | 2 | 1.935654 | 0.032955 |
| P00738 | Haptoglobin | 30.05 | 15 | 15 | 1.925942 | 0.021916 |
| Q6P163 | APOC2 protein | 25 | 2 | 2 | 1.803961 | 0.0011 |
| Q9NPP6 | Immunoglobulin heavy chain variant | 17.79 | 2 | 9 | 1.799359 | 0.001327 |
| A0A075B7E8 | Protein IGHV3OR16-13 | 11.97 | 2 | 2 | 1.789869 | 0.010504 |
| H0YLF3 | β-2-Microglobulin (Fragment) | 28.17 | 2 | 2 | 1.782909 | 0.00658 |
| P05090 | Apolipoprotein D | 30.16 | 6 | 6 | 1.763858 | 0.045205 |
| B4DPP6 | Serum albumin | 53.88 | 5 | 49 | 1.748857 | 0.000246 |
| V9GYE3 | Apolipoprotein A-II | 42.31 | 5 | 5 | 1.740055 | 0.000297 |
| A8K9A9 | Kallikrein B, plasma | 12.54 | 8 | 8 | 1.719625 | 0.000228 |
| P02790 | Hemopexin | 19.26 | 12 | 12 | 1.622425 | 0.000326 |
| Q9H804 | Completement C1R | 7.31 | 2 | 3 | 1.580432 | 0.006066 |
| P02763 | α-1-Acid glycoprotein 1 | 16.42 | 2 | 6 | 1.572967 | 0.009053 |
| P02647 | Apolipoprotein A-I | 46.44 | 17 | 17 | 1.56536 | 0.00079 |
| A0A0K0Q2Z1 | Serpin peptidase inhibitor clade C member 1 | 19.83 | 13 | 13 | 1.562564 | 0.000278 |
| B7Z544 | Inter-α-trypsin inhibitor heavy chain H4 | 11.71 | 11 | 11 | 1.546281 | 0.003001 |
| Q96PD5 | N-acetylmuramoyl-l-alanine amidase | 7.29 | 3 | 3 | 1.545521 | 0.00139 |
| B4DPP8 | Kininogen-1 | 30.84 | 17 | 17 | 1.48514 | 0.000647 |
| B4E1I8 | Leucine-rich α-2-glycoprotein | 15.15 | 6 | 6 | 1.458316 | 0.006872 |
| E9PIT3 | Prothrombin | 13.04 | 8 | 8 | 1.455448 | 0.013628 |
| P43652 | Afamin | 25.04 | 17 | 17 | 1.453959 | 0.003423 |
| P08697 | α-2-Antiplasmin | 12.83 | 7 | 7 | 1.450495 | 0.008491 |
| B2R582 | C-type lectin domain family 3, member B | 21.78 | 4 | 4 | 1.447796 | 0.002065 |
| P06727 | Apolipoprotein A-IV | 42.17 | 21 | 21 | 1.44284 | 0.000649 |
| Q5SQ08 | Complement component C8 γ chain | 11.59 | 2 | 2 | 1.431101 | 0.018442 |
| Q9UL84 | Myosin-reactive immunoglobulin heavy chain variable region | 14.75 | 2 | 2 | 1.387229 | 0.017727 |
| P04217 | α-1B-Glycoprotein | 13.74 | 7 | 7 | 1.372869 | 0.014165 |
| B7Z539 | Inter-α-trypsin inhibitor heavy chain H1 | 7.91 | 7 | 7 | 1.363604 | 0.027945 |
| B2R7F8 | Plasminogen | 24.2 | 22 | 22 | 1.360977 | 0.007811 |
| A0A0C4DGV7 | Retinol-binding protein 4 | 14.07 | 4 | 4 | 1.359868 | 0.04222 |
| P01024 | Complement C3 | 24.95 | 54 | 54 | 1.341747 | 0.002478 |
| P02765 | α-2-HS-glycoprotein | 26.16 | 10 | 10 | 1.333365 | 0.00478 |
| B7Z550 | Complement component 8, β polypeptide | 7.56 | 4 | 4 | 1.329365 | 0.007697 |
| P07357 | Complement component C8 α chain | 11.47 | 7 | 7 | 1.320078 | 0.04949 |
| Q5T985 | Inter-α-trypsin inhibitor heavy chain H2 | 15.51 | 17 | 17 | 1.310572 | 0.030167 |
| P25311 | Zinc-α-2-glycoprotein | 30.2 | 10 | 10 | 1.307167 | 0.031312 |
| P01009 | α-1-Antitrypsin | 27.51 | 14 | 14 | 1.303075 | 0.020778 |
| P13671 | Component C6 | 6.64 | 7 | 7 | 1.224111 | 0.03757 |
| Accession | Description | Coverage | Unique Peptides | Peptides | Fold Change | t Test p Value |
|---|---|---|---|---|---|---|
| Q16270 | Insulin-like growth factor-binding protein 7 | 28.72 | 7 | 7 | 0.795347 | 0.048896 |
| H7BXV5 | Collagen α-1(XVIII) chain | 6.84 | 4 | 4 | 0.72897 | 0.007309 |
| B4DEN5 | N-acetyllactosaminide β-1,3-N-acetylglucosaminyltransferase | 14.71 | 6 | 6 | 0.697059 | 0.007153 |
| A8KAJ3 | EGF-containing fibulin-like extracellular matrix protein 1 | 22.31 | 10 | 10 | 0.694626 | 0.020864 |
| O75326 | Semaphorin-7A | 14.56 | 9 | 9 | 0.688332 | 0.008646 |
| Q12860 | Contactin-1 | 8.35 | 8 | 8 | 0.687135 | 0.010164 |
| P41222 | Prostaglandin-H2 d-isomerase | 17.89 | 7 | 7 | 0.673721 | 0.012421 |
| Q9UHG2 | ProSAAS | 15 | 3 | 3 | 0.67195 | 0.006702 |
| Q8NHP8 | Putative phospholipase B-like 2 | 3.57 | 2 | 2 | 0.669177 | 0.017856 |
| Q92520 | Protein FAM3C | 24.23 | 6 | 6 | 0.668184 | 0.00515 |
| O15240 | Neurosecretory protein VGF | 14.15 | 8 | 8 | 0.661762 | 0.010221 |
| B4DZY7 | Growth-arrest-specific protein 6 | 5.72 | 3 | 3 | 0.650294 | 0.033296 |
| B4DUI5 | Triosephosphate isomerase | 25.82 | 5 | 5 | 0.642348 | 0.009858 |
| A0A0C4DFP6 | Cartilage acidic protein 1 | 9.73 | 7 | 7 | 0.636797 | 0.005166 |
| P10909 | Clusterin | 25.84 | 6 | 14 | 0.629457 | 0.000292 |
| B4DZX0 | Nucleobindin-1 | 14.51 | 7 | 7 | 0.627294 | 0.000278 |
| A0A087X0S5 | Collagen α-1(VI) chain | 8.09 | 8 | 8 | 0.617461 | 0.013995 |
| Q9UBP4 | Dickkopf-related protein 3 | 18.57 | 6 | 6 | 0.603169 | 0.004542 |
| O00391 | Sulfhydryl oxidase 1 | 9.37 | 7 | 7 | 0.598255 | 0.003939 |
| P35555 | Fibrillin-1 | 1.99 | 7 | 7 | 0.597125 | 0.005083 |
| F8W785 | Golgi integral membrane protein 4 | 6.59 | 4 | 4 | 0.592385 | 0.022707 |
| B4DGD0 | Amyloid β A4 protein | 11.34 | 8 | 9 | 0.586216 | 0.009153 |
| F5GZ08 | Amyloid-like protein 1 | 15.71 | 7 | 10 | 0.582642 | 0.011373 |
| O15354 | Prosaposin receptor | 2.77 | 2 | 2 | 0.567766 | 0.016087 |
| P02458 | Collagen α-1(II) chain | 3.36 | 5 | 5 | 0.564508 | 0.025358 |
| A0A087X232 | Complement C1s subcomponent | 6.45 | 5 | 5 | 0.539272 | 0.021601 |
| A0A096LP62 | Inter-α-trypsin inhibitor heavy chain H5 | 7.28 | 5 | 5 | 0.53903 | 0.024978 |
| P81605 | Dermcidin | 27.27 | 4 | 4 | 0.537086 | 0.029414 |
| F5H5G1 | Limbic system-associated membrane protein | 6.6 | 2 | 2 | 0.528697 | 0.007773 |
| Q12841 | Follistatin-related protein 1 | 28.9 | 9 | 9 | 0.495666 | 0.000459 |
| Q14515 | SPARC-like protein 1 | 11.6 | 7 | 7 | 0.494821 | 0.001398 |
| Q99574 | Neuroserpin | 7.8 | 4 | 4 | 0.493937 | 0.026239 |
| Q6MZW2 | Follistatin-related protein 4 | 4.51 | 2 | 3 | 0.485352 | 0.028432 |
| P02649 | Apolipoprotein E | 63.72 | 20 | 22 | 0.480489 | 0.00203 |
| A0A024RAB6 | Heparan sulfate proteoglycan 2 (Perlecan) | 3.89 | 17 | 17 | 0.479996 | 6.82E-05 |
| P13521 | Secretogranin-2 | 6.97 | 4 | 4 | 0.470064 | 0.028148 |
| B4E3Q1 | Calsyntenin-1 | 10.71 | 13 | 13 | 0.440009 | 0.008494 |
| B3KX75 | Neural cell adhesion molecule L1-like protein | 3.14 | 4 | 4 | 0.431103 | 0.04737 |
| Q9HCB6 | Spondin-1 | 11.77 | 10 | 10 | 0.412771 | 0.000529 |
| B3KXD3 | Carboxypeptidase E | 20.43 | 10 | 10 | 0.376381 | 0.001059 |
| A0A024R3N3 | Amyloid β (A4)-like protein 2 | 12.68 | 8 | 9 | 0.37116 | 0.001787 |
| B4E367 | Plexin domain-containing protein 2 | 3.69 | 2 | 2 | 0.363499 | 0.006849 |
| B2R6W4 | Frizzled-related protein (FRZB) | 11.69 | 4 | 4 | 0.291076 | 0.000102 |
| Q9Y5W5 | Wnt inhibitory factor 1 | 10.82 | 4 | 4 | 0.290436 | 0.009571 |
References
- Li, Z.; Li, Y.; Huang, X.; Cai, X.Y.; Chen, X.; Li, S.; Huang, Y.S.; Lu, L. Quantitative analysis of rhegmatogenous retinal detachment associated with choroidal detachment in Chinese using UBM. Retina 2012, 32, 2020–2025. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, F. Combined choroidal and retinal detachment. Arch. Ophthalmol. 1972, 88, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, W.H., 2nd. Rhematogenous retinal detachment complicated by severe intraocular inflammation, hypotony, and choroidal detachment. Trans. Am. Ophthalmol. Soc. 1981, 79, 664–683. [Google Scholar] [PubMed]
- Zhu, J.; Xu, X.; Zhang, X. Surgical therapeutic results of rhegmatogenous retinal detachment associated with choroidal detachment. Zhonghua Yan Ke Za Zhi 2002, 38, 135–139. [Google Scholar] [PubMed]
- Seelenfreund, M.H.; Kraushar, M.F.; Schepens, C.L.; Freilich, D.B. Choroidal detachment associated with primary retinal detachment. Arch. Ophthalmol. 1974, 91, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.; Harris, G.S. Choroidal detachment associated with retinal detachment as a presenting finding. Can. J. Ophthalmol. 1992, 27, 245–248. [Google Scholar] [PubMed]
- Kang, J.H.; Park, K.A.; Shin, W.J.; Kang, S.W. Macular hole as a risk factor of choroidal detachment in rhegmatogenous retinal detachment. Korean J. Ophthalmol. 2008, 22, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M. Pars plana vitrectomy in the treatment of combined rhegmatogenous retinal detachment and choroidal detachment in aphakic or pseudophakic patients. Ophthalmic Surg. Lasers 1997, 28, 288–293. [Google Scholar] [PubMed]
- Dai, Y.; Wu, Z.; Sheng, H.; Zhang, Z.; Yu, M.; Zhang, Q. Identification of inflammatory mediators in patients with rhegmatogenous retinal detachment associated with choroidal detachment. Mol. Vis. 2015, 21, 417–427. [Google Scholar] [PubMed]
- Yu, M.; Wu, Z.; Zhang, Z.; Huang, X.; Zhang, Q. Metabolomic analysis of human vitreous in rhegmatogenous retinal detachment associated with choroidal detachment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5706–5713. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Yadav, A.K. Dissecting the iTRAQ data analysis. Methods Mol. Biol. 2016, 1362, 277–291. [Google Scholar] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The gene ontology consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Janciauskiene, S.M.; Bals, R.; Koczulla, R.; Vogelmeier, C.; Köhnlein, T.; Welte, T. The discovery of α-antitrypsin and its role in health and disease. Respir. Med. 2011, 105, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, M.R. Immune-modulating effects of α-1 antitrypsin. Biol. Chem. 2014, 395, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Grisanti, S.; Wiedemann, P.; Weller, M.; Heimann, K.; Zilles, K. The significance of complement in proliferative vitreoretinopathy. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2711–2717. [Google Scholar]
- Baudouin, C.; Gordon, W.C.; Baudouin, F.; Peyman, G.; Lapalus, P.; Gastaud, P.; Bazan, N.G.; Fredj-Reygrobellet, D. Immunohistologic study of epiretinal membranes in proliferative vitreoretinopathy. Am. J. Ophthalmol. 1990, 110, 593–598. [Google Scholar] [CrossRef]
- Walport, M.J. Complement. First of two parts. N. Engl. J. Med. 2001, 344, 1058–1066. [Google Scholar] [PubMed]
- Klos, A.; Tenner, A.J.; Johswich, K.O.; Ager, R.R.; Reis, E.S.; Köhl, J. The role of the anaphylatoxins in health and disease. Mol. Immunol. 2009, 46, 2753–2766. [Google Scholar] [CrossRef] [PubMed]
- Silasi-Mansat, R.; Zhu, H.; Popescu, N.I.; Peer, G.; Sfyroera, G.; Magotti, P.; Ivanciu, L.; Lupu, C.; Mollnes, T.E.; Taylor, F.B.; et al. Complement inhibition decreases the procoagulant response and confers organ protection in a baboon model of Escherichia coli sepsis. Blood 2010, 116, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Hack, C.E.; Nuijens, J.H.; Felt-Bersma, R.J.; Schreuder, W.O.; Eerenberg-Belmer, A.J.; Paardekooper, J.; Bronsveld, W.; Thijs, L.G. Elevated plasma levels of the anaphylatoxins C3a and C4a are associated with a fatal outcome in sepsis. Am. J. Med. 1989, 86, 20–26. [Google Scholar] [CrossRef]
- Schuerholz, T.; Leuwer, M.; Cobas-Meyer, M.; Vangerow, B.; Kube, F.; Kirschfink, M.; Marx, G. Terminal complement complex in septic shock with capillary leakage: Marker of complement activation? Eur. J. Anaesthesiol. 2005, 22, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Norris, L.A. Blood coagulation. Best Pract. Res. Clin. Obstet. Gynaecol. 2003, 17, 369–383. [Google Scholar] [CrossRef]
- De Witt, S.M.; Verdoold, R.; Cosemans, J.M.; Heemskerk, J.W. Insights into platelet-based control of coagulation. Thromb. Res. 2014, 133, S139–S148. [Google Scholar] [CrossRef]
- O'Brien, M. The reciprocal relationship between inflammation and coagulation. Top. Companion Anim. Med. 2012, 27, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Mackman, N. The many faces of tissue factor. J. Thromb. Haemost. 2009, 7, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Torzewski, M.; Bhakdi, S. Complement and atherosclerosis-united to the point of no return? Clin. Biochem. 2013, 46, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Loukovaara, S.; Loukovaara, S.; Nurkkala, H.; Tamene, F.; Gucciardo, E.; Liu, X.; Repo, P.; Lehti, K.; Varjosalo, M. Quantitative proteomics analysis of vitreous humor from diabetic retinopathy patients. J. Proteome Res. 2015, 14, 5131–5143. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Liu, F.; Cui, S.J.; Liu, Y.; Song, Z.Y.; Cao, H.; Chen, F.E.; Wang, W.J.; Sun, T.; Wang, F. Vitreous proteomic analysis of proliferative vitreoretinopathy. Proteomics 2008, 8, 3667–3678. [Google Scholar] [CrossRef] [PubMed]
- Pathak, M.; Wong, S.S.; Dreveny, I.; Emsley, J. Structure of plasma and tissue kallikreins. Thromb. Haemost. 2013, 110, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Schmaier, A.H. The plasma kallikrein-kinin system counterbalances the renin-angiotensin system. J. Clin. Investig. 2002, 109, 1007–1009. [Google Scholar] [CrossRef] [PubMed]
- Diamandis, E.P.; Yousef, G.M.; Clements, J.; Ashworth, L.K.; Yoshida, S.; Egelrud, T.; Nelson, P.S.; Shiosaka, S.; Little, S.; Lilja, H.; et al. New nomenclature for the human tissue kallikrein gene family. Clin. Chem. 2000, 46, 1855–1858. [Google Scholar] [PubMed]
- Campbell, D.J. The kallikrein-kinin system in humans. Clin. Exp. Pharmacol. Physiol. 2001, 28, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.X.; Song, Q.; Hatcher, H.C.; Crouch, R.K.; Chao, L.; Chao, J. Expression and cellular localization of the kallikrein-kinin system in human ocular tissues. Exp. Eye Res. 1996, 63, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clermont, A.C.; Gao, B.B.; Feener, E.P. Intraocular hemorrhage causes retinal vascular dysfunction via plasma kallikrein. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Phipps, J.A.; Clermont, A.C.; Sinha, S.; Chilcote, T.J.; Bursell, S.E.; Feener, E.P. Plasma kallikrein mediates angiotensin II type 1 receptor-stimulated retinal vascular permeability. Hypertension 2009, 53, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Peng, R.; Chen, H.; Cui, C.; Ba, J.; Wang, F. Kininogen 1 and insulin-like growth factor binding protein 6: Candidate serum biomarkers of proliferative vitreoretinopathy. Clin. Exp. Optom. 2014, 97, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, C.L.; Dobri, N.; Freeman, E.E.; Conlon, M.P.; Chen, P.; Stafford, D.G.; Schwarz, D.M.C.; Golden, K.C.; Zhu, L.; Kitchen, D.B.; et al. Design, synthesis, and evaluation of nonretinoid retinol binding protein 4 antagonists for the potential treatment of atrophic age-related macular degeneration and Stargardt disease. J. Med. Chem. 2014, 57, 7731–7757. [Google Scholar] [CrossRef] [PubMed]
- Retina Society Terminology Committee. The classification of retinal detachment with proliferative vitreoretinopathy. Ophthalmology 1983, 90, 121–125. [Google Scholar]
- Chiu, K.H.; Chang, Y.H.; Wu, Y.S.; Lee, S.H.; Liao, P.C. Quantitative secretome analysis reveals that COL6A1 is a metastasis-associated protein using stacking gel-aided purification combined with iTRAQ labeling. J. Proteome Res. 2011, 10, 1110–1125. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]


| Clinical Characteristics | RRDCD | RRD | p Values |
|---|---|---|---|
| N = 8 | N = 8 | ||
| Sex, n (%) | |||
| Males | 3 (37.5) | 3 (37.5) | 1.0 |
| Females | 5 (62.5) | 5 (62.5) | |
| Age, y | |||
| Median (range) | 60 (46–65) | 59.5 (39–65) | 0.959 |
| Duration of detachments, d | |||
| Median (range) | 8.5 (5–13) | 13 (7–20) | 0.065 |
| PVR grade | |||
| Mild, A, B | 1 | 8 | 0.001 |
| Heavy, C, D | 7 | 0 | |
| IOP, mmHg | |||
| Median (range) | 7.75 (6.8–13.4) | 14 (11.9–17.2) | 0.002 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Ding, N.; Yu, M.; Wang, K.; Luo, S.; Zou, W.; Zhou, Y.; Yan, B.; Jiang, Q. Identification of Potential Biomarkers for Rhegmatogenous Retinal Detachment Associated with Choroidal Detachment by Vitreous iTRAQ-Based Proteomic Profiling. Int. J. Mol. Sci. 2016, 17, 2052. https://doi.org/10.3390/ijms17122052
Wu Z, Ding N, Yu M, Wang K, Luo S, Zou W, Zhou Y, Yan B, Jiang Q. Identification of Potential Biomarkers for Rhegmatogenous Retinal Detachment Associated with Choroidal Detachment by Vitreous iTRAQ-Based Proteomic Profiling. International Journal of Molecular Sciences. 2016; 17(12):2052. https://doi.org/10.3390/ijms17122052
Chicago/Turabian StyleWu, Zhifeng, Nannan Ding, Mengxi Yu, Ke Wang, Shasha Luo, Wenjun Zou, Ying Zhou, Biao Yan, and Qin Jiang. 2016. "Identification of Potential Biomarkers for Rhegmatogenous Retinal Detachment Associated with Choroidal Detachment by Vitreous iTRAQ-Based Proteomic Profiling" International Journal of Molecular Sciences 17, no. 12: 2052. https://doi.org/10.3390/ijms17122052