
Cre Fused with RVG Peptide Mediates Targeted Genome Editing in Mouse Brain Cells In Vivo

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
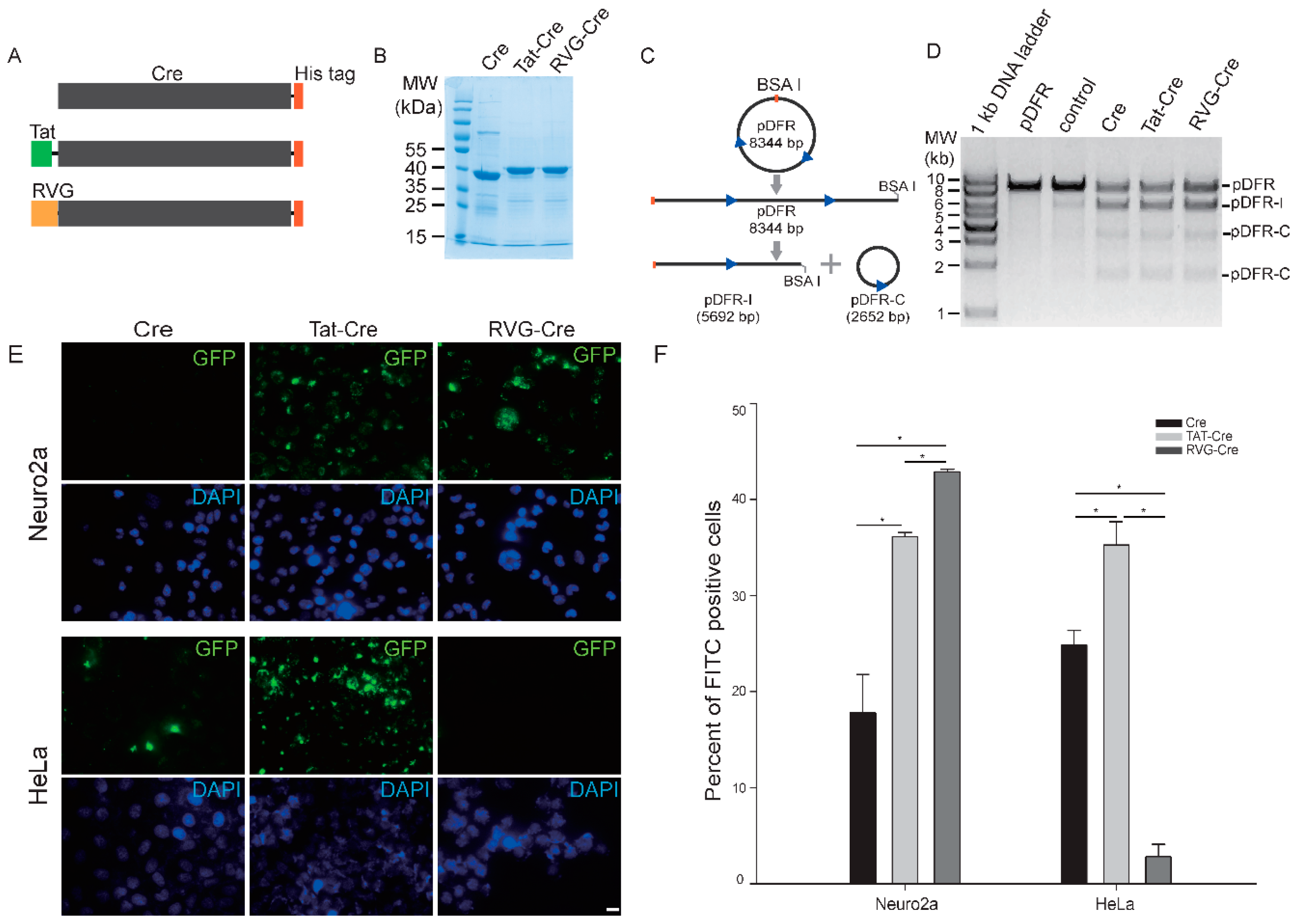
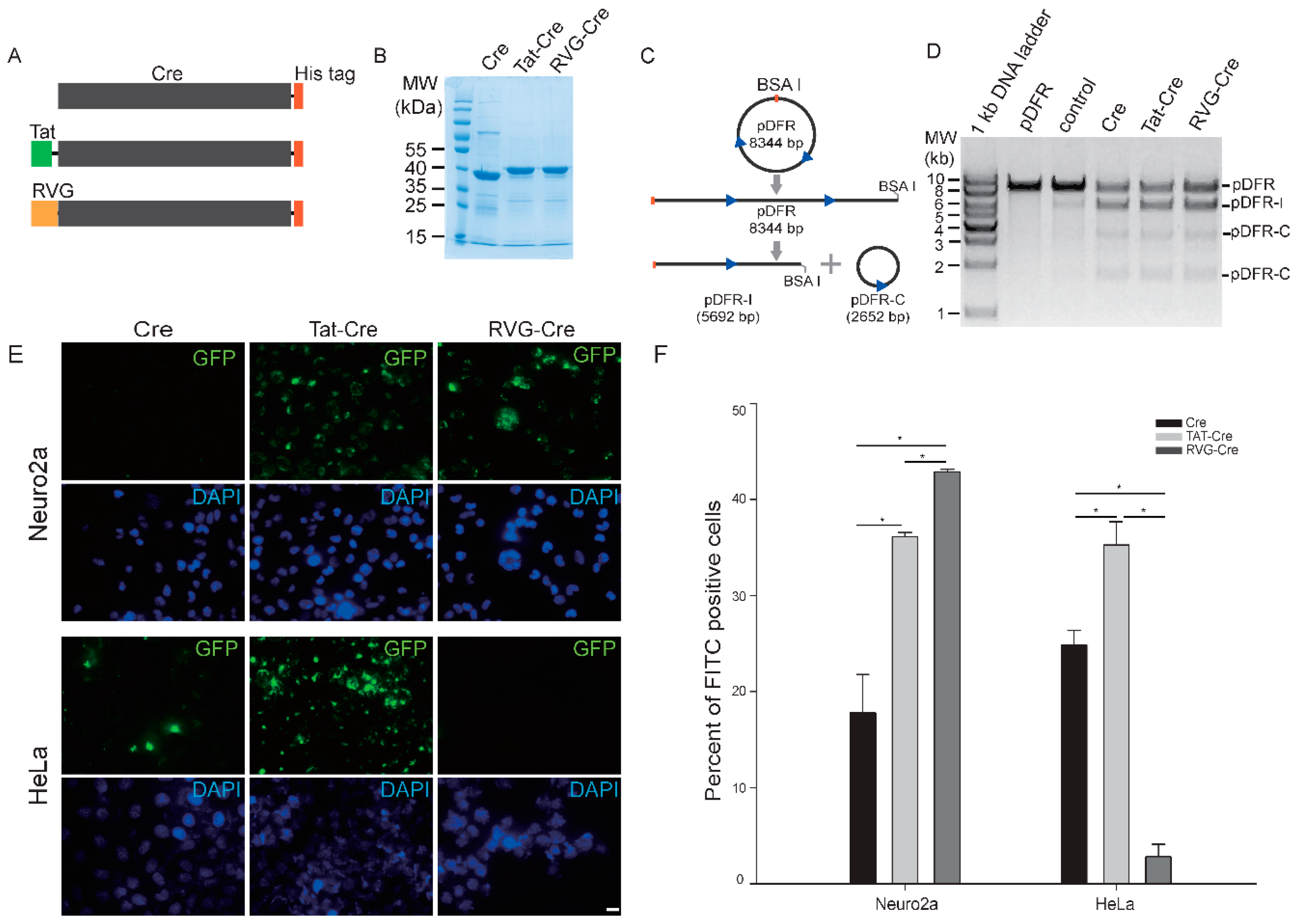
2.1. RVG-Cre Properties In Vitro
2.2. RVG-Cre Cell Line Preference
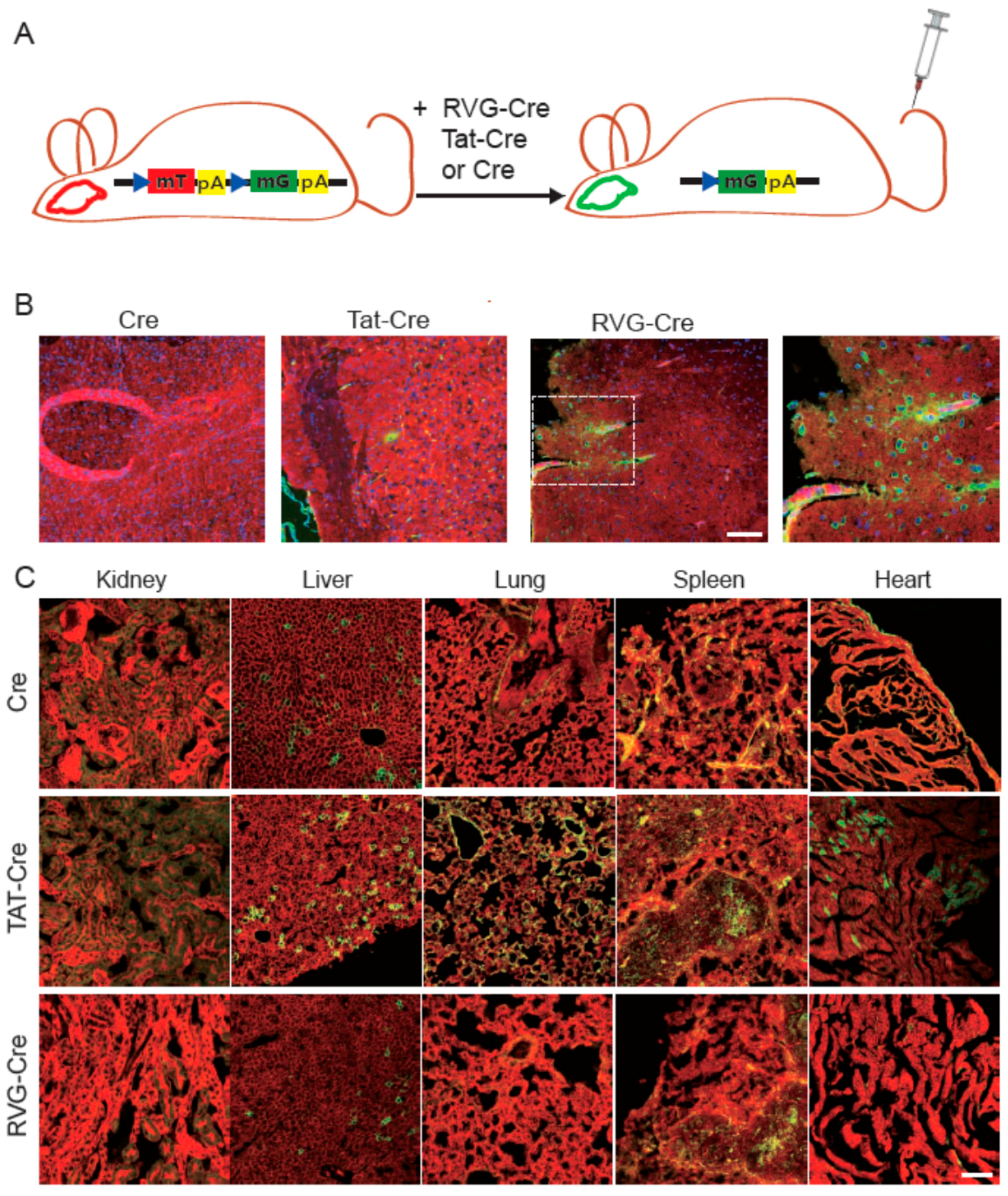
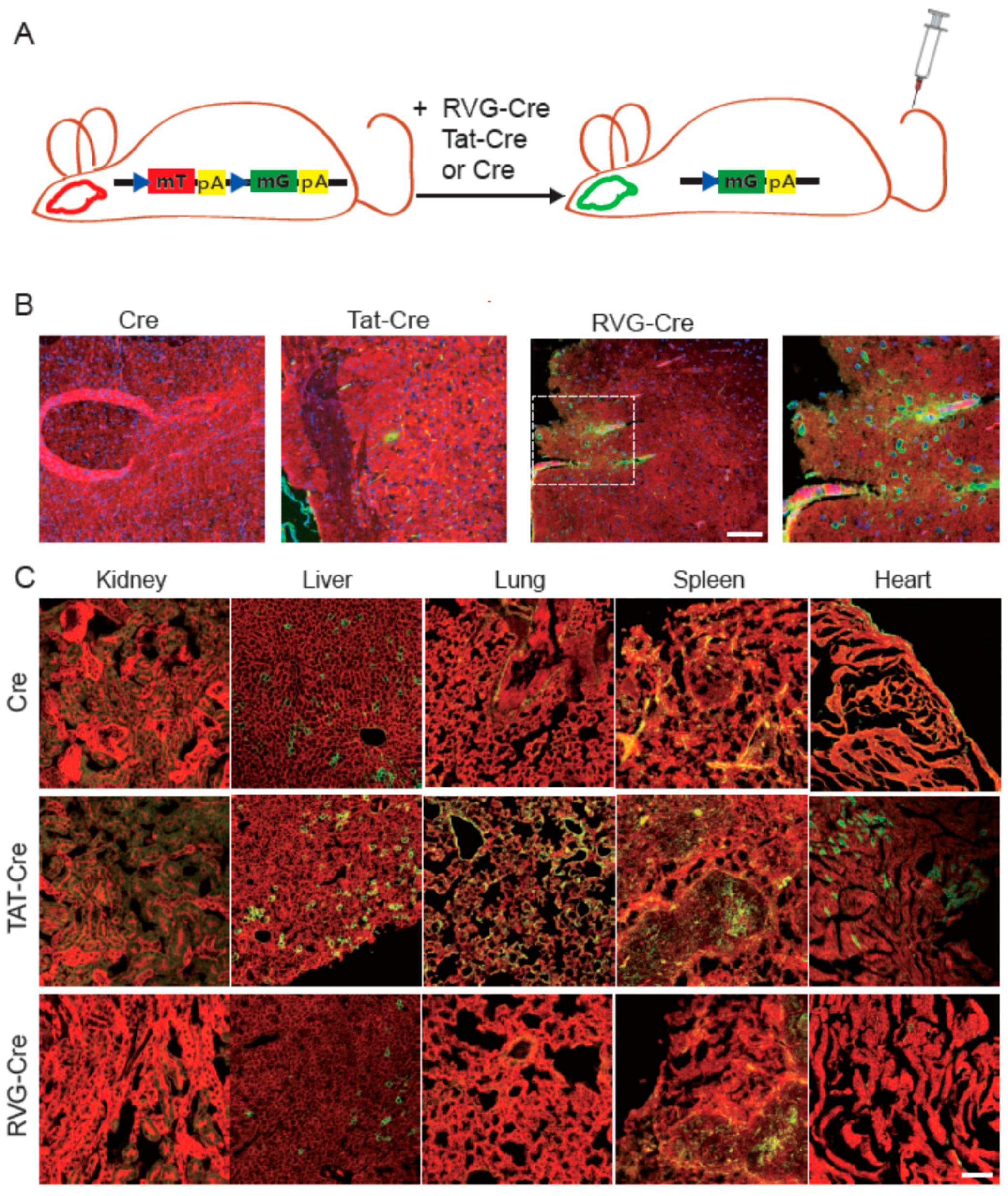
2.3. RVG-Cre Functions in mTmG Mice
2.4. Tissue Specificity of RVG-Cre In Vivo
2.5. Confirmation of Cre Targeting and Activity in Rosa26lacZ Mice
3. Discussion
4. Materials and Methods
4.1. Cloning, Overexpression and Purification of WT Cre, TAT-Cre and RVG-Cre
4.2. Enzyme Activity Assay of Three Cre Fusion Proteins In Vitro
4.3. Transduction of FITC-Labeled Recombinant Proteins into Cells
4.4. In Vivo Test of Cre Proteins with mTmG and Rosa26lacZ Mouse Lines
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CPP | Cell penetrating peptide |
| BBB | blood-brain barrier |
| RVG | rabies virus glycoprotein |
| wt | wild type |
| mT | a membrane form tdTomato fluorescent protein |
| mG | a membrane form GFP fluorescent protein |
References
- Milletti, F. Cell-penetrating peptides: Classes, origin, and current landscape. Drug Discov. Today 2012, 17, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Z.Y.; Zhang, J.; Zhang, Y.; Huo, H.; Wang, T.; Jiang, T.; Wang, S. RVG-peptide-linked trimethylated chitosan for delivery of siRNA to the brain. Biomacromolecules 2014, 15, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, M.A. Peptide-mediated cell delivery: Application in protein target validation. Curr. Opin. Pharmacol. 2002, 2, 587–594. [Google Scholar] [CrossRef]
- Schwarze, S.R.; Ho, A.; Vocero-Akbani, A.; Dowdy, S.F. In vivo protein transduction: Delivery of a biologically active protein into the mouse. Science 1999, 285, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Kondo, E.; Saito, K.; Tashiro, Y.; Kamide, K.; Uno, S.; Furuya, T.; Mashita, M.; Nakajima, K.; Tsumuraya, T.; Kobayashi, N.; et al. Tumour lineage-homing cell-penetrating peptides as anticancer molecular delivery systems. Nat. Commun. 2012, 3, 951. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Liu, B.; Kodadek, T. A high-throughput assay for assessing the cell permeability of combinatorial libraries. Nat. Biotechnol. 2005, 23, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Wu, H.; McBride, J.L.; Jung, K.E.; Kim, M.H.; Davidson, B.L.; Lee, S.K.; Shankar, P.; Manjunath, N. Transvascular delivery of small interfering RNA to the central nervous system. Nature 2007, 448, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Tuffereau, C.; Schmidt, K.; Langevin, C.; Lafay, F.; Dechant, G.; Koltzenburg, M. The rabies virus glycoprotein receptor p75NTR is not essential for rabies virus infection. J. Virol. 2007, 81, 13622–13630. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Zhou, R.; Fu, A.; Xu, X.; Huang, Y.; Hu, C. Targeted delivery of large fusion protein into hippocampal neurons by systemic administration. J. Drug Target. 2011, 19, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, R.; Han, L.; Ke, W.; Shao, K.; Ye, L.; Lou, J.; Jiang, C. Brain-targeting gene delivery and cellular internalization mechanisms for modified rabies virus glycoprotein RVG29 nanoparticles. Biomaterials 2009, 30, 4195–4202. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.L.; Abremski, K. Site-specific recombination by the bacteriophage P1 lox-Cre system. Cre-mediated synapsis of two lox sites. J. Mol. Biol. 1984, 178, 481–486. [Google Scholar] [CrossRef]
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis 2000, 26, 99–109. [Google Scholar] [CrossRef]
- Muzumdar, M.D.; Tasic, B.; Miyamichi, K.; Li, L.; Luo, L. A global double-fluorescent Cre reporter mouse. Genesis 2007, 45, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Soriano, P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat. Genet. 1999, 21, 70–71. [Google Scholar] [CrossRef] [PubMed]
- Jo, D.; Nashabi, A.; Doxsee, C.; Lin, Q.; Unutmaz, D.; Chen, J.; Ruley, H.E. Epigenetic regulation of gene structure and function with a cell-permeable Cre recombinase. Nat. Biotechnol. 2001, 19, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Chien, W.M.; Liu, Y.; Chin, M.T. Genomic DNA recombination with cell-penetrating peptide-tagged Cre protein in mouse skeletal and cardiac muscle. Genesis 2014, 52, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Cronican, J.J.; Thompson, D.B.; Beier, K.T.; McNaughton, B.R.; Cepko, C.L.; Liu, D.R. Potent delivery of functional proteins into Mammalian cells in vitro and in vivo using a supercharged protein. ACS Chem. Biol. 2010, 5, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Gitton, Y.; Tibaldi, L.; Dupont, E.; Levi, G.; Joliot, A. Efficient CPP-mediated Cre protein delivery to developing and adult CNS tissues. BMC Biotechnol. 2009, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Zuris, J.A.; Thompson, D.B.; Shu, Y.; Guilinger, J.P.; Bessen, J.L.; Hu, J.H.; Maeder, M.L.; Joung, J.K.; Chen, Z.Y.; Liu, D.R. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.P.; Melikov, K.; Brooks, H.; Prevot, P.; Lebleu, B.; Chernomordik, L.V. Cellular uptake of unconjugated TAT peptide involves clathrin-dependent endocytosis and heparan sulfate receptors. J. Biol. Chem. 2005, 280, 15300–15306. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, Z.; Sun, Z.; Li, P.; Feng, T.; Wu, S. Cre Fused with RVG Peptide Mediates Targeted Genome Editing in Mouse Brain Cells In Vivo. Int. J. Mol. Sci. 2016, 17, 2104. https://doi.org/10.3390/ijms17122104
Zou Z, Sun Z, Li P, Feng T, Wu S. Cre Fused with RVG Peptide Mediates Targeted Genome Editing in Mouse Brain Cells In Vivo. International Journal of Molecular Sciences. 2016; 17(12):2104. https://doi.org/10.3390/ijms17122104
Chicago/Turabian StyleZou, Zhiyuan, Zhaolin Sun, Pan Li, Tao Feng, and Sen Wu. 2016. "Cre Fused with RVG Peptide Mediates Targeted Genome Editing in Mouse Brain Cells In Vivo" International Journal of Molecular Sciences 17, no. 12: 2104. https://doi.org/10.3390/ijms17122104