
Mitochondria Related Pathway Is Essential for Polysaccharides Purified from Sparassis crispa Mediated Neuro-Protection against Glutamate-Induced Toxicity in Differentiated PC12 Cells

Abstract
:1. Introduction
2. Results
2.1. Purification and Characterization of Sparassis crispa Polysaccharides


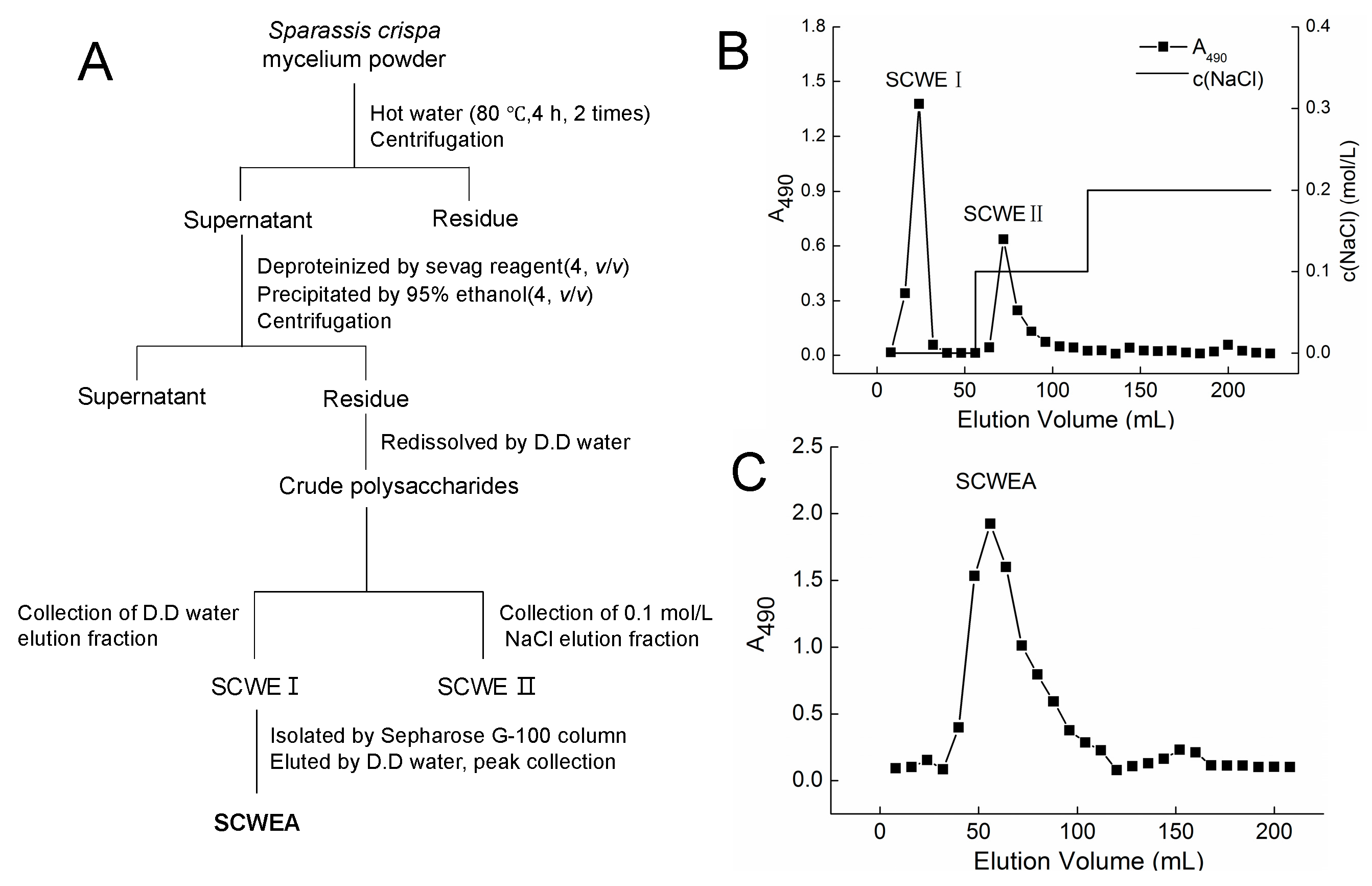{kind=link}
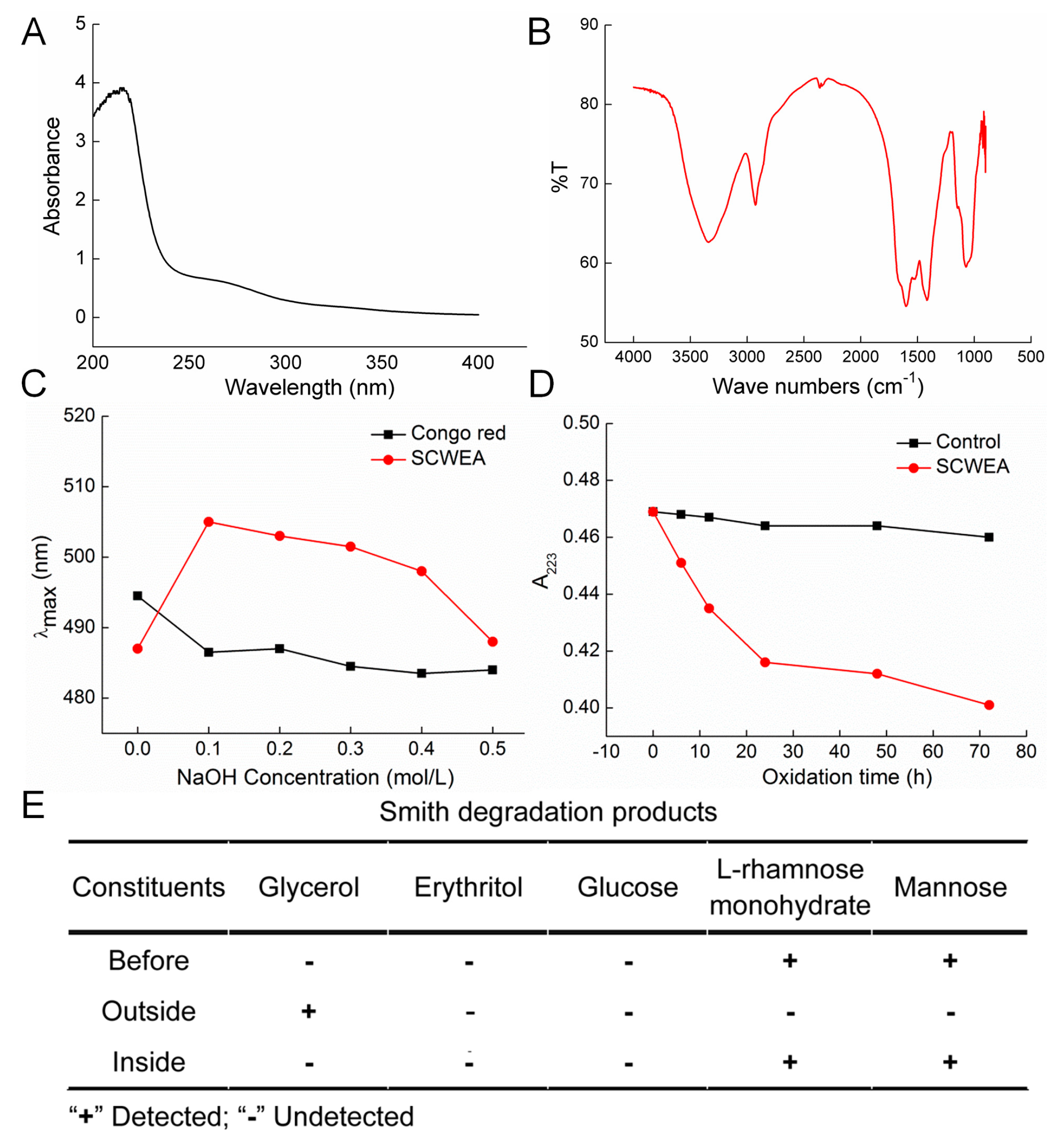{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular weight 1 | Mr (kDa) | molar ratio, mol % |
| 75 | 96.7 | |
| Particle size 2 | Z-Average (r.nm) 3 | PdI 4 |
| 44.45 | 0.306 |
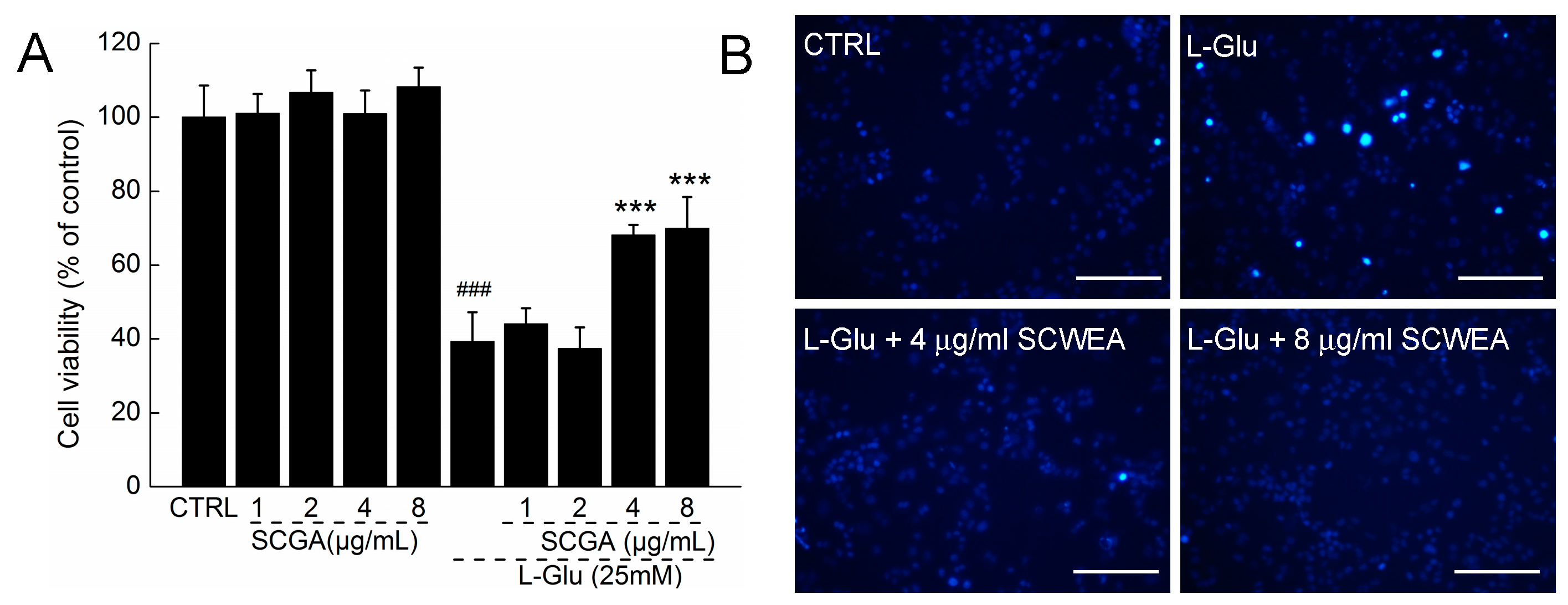
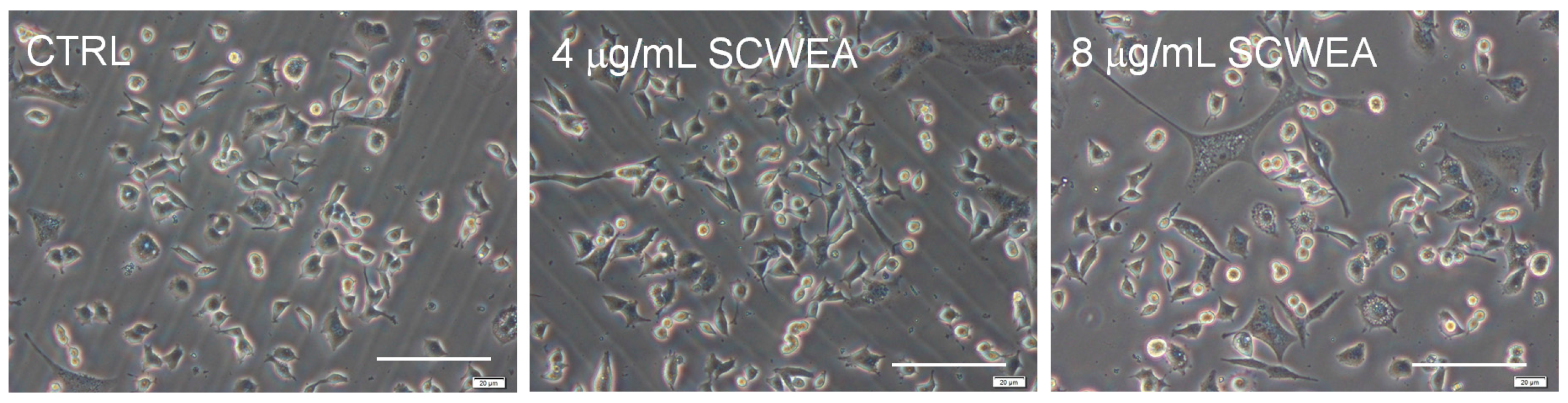
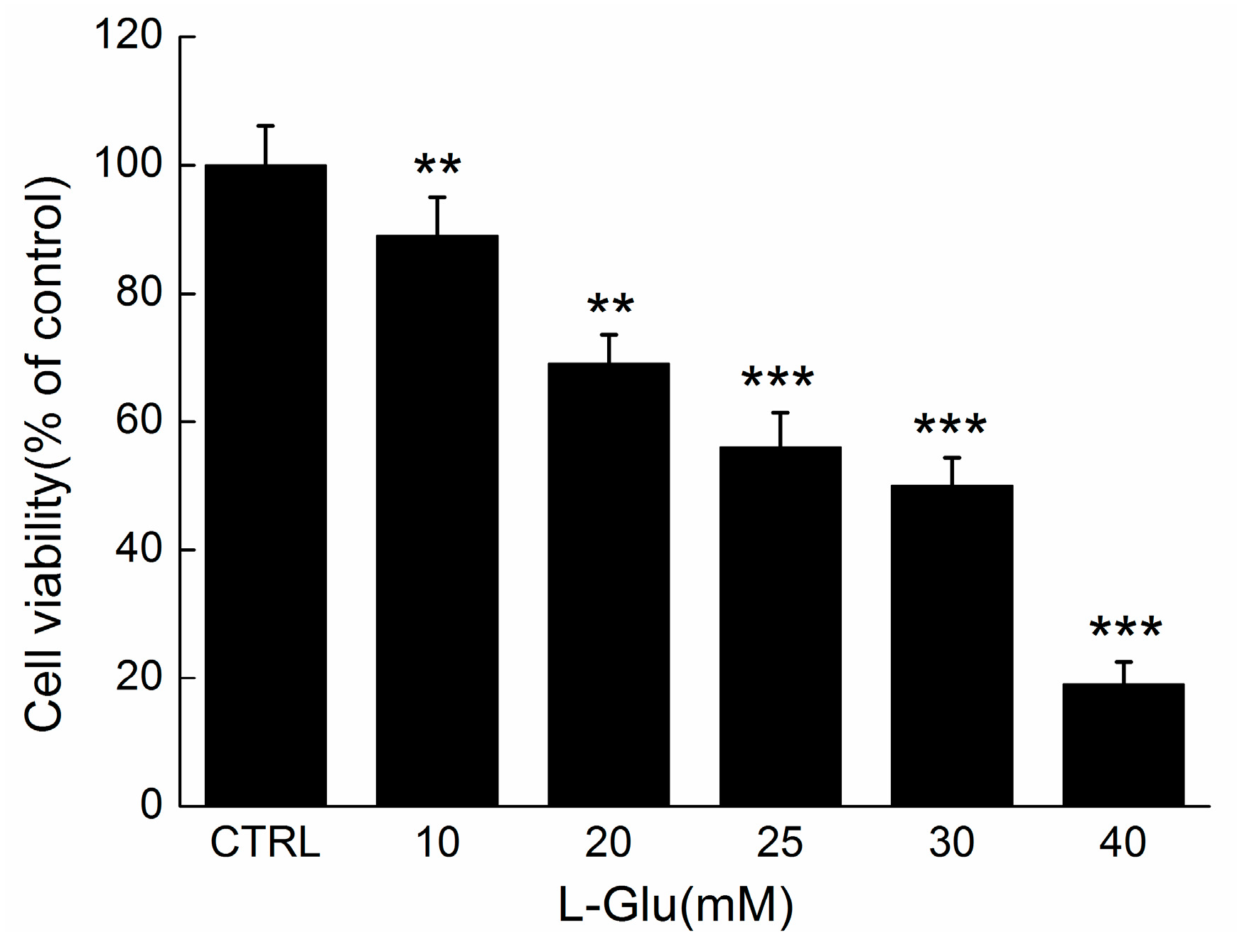
2.2. The Effect of SCWEA against l-Glu Induced Cell Damage in DPC12 Cells
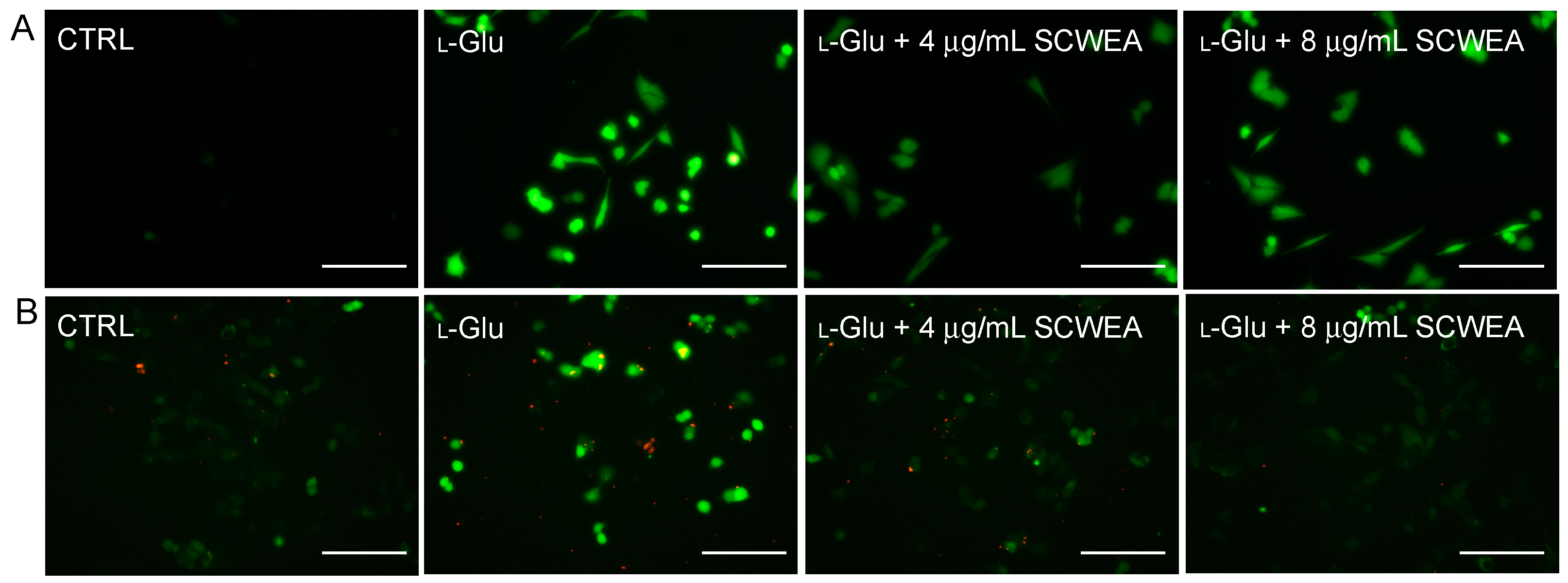
2.3. The Effects of SCWEA on Intracellular Calcium Concentration and Reactive Oxygen Species (ROS) Levels


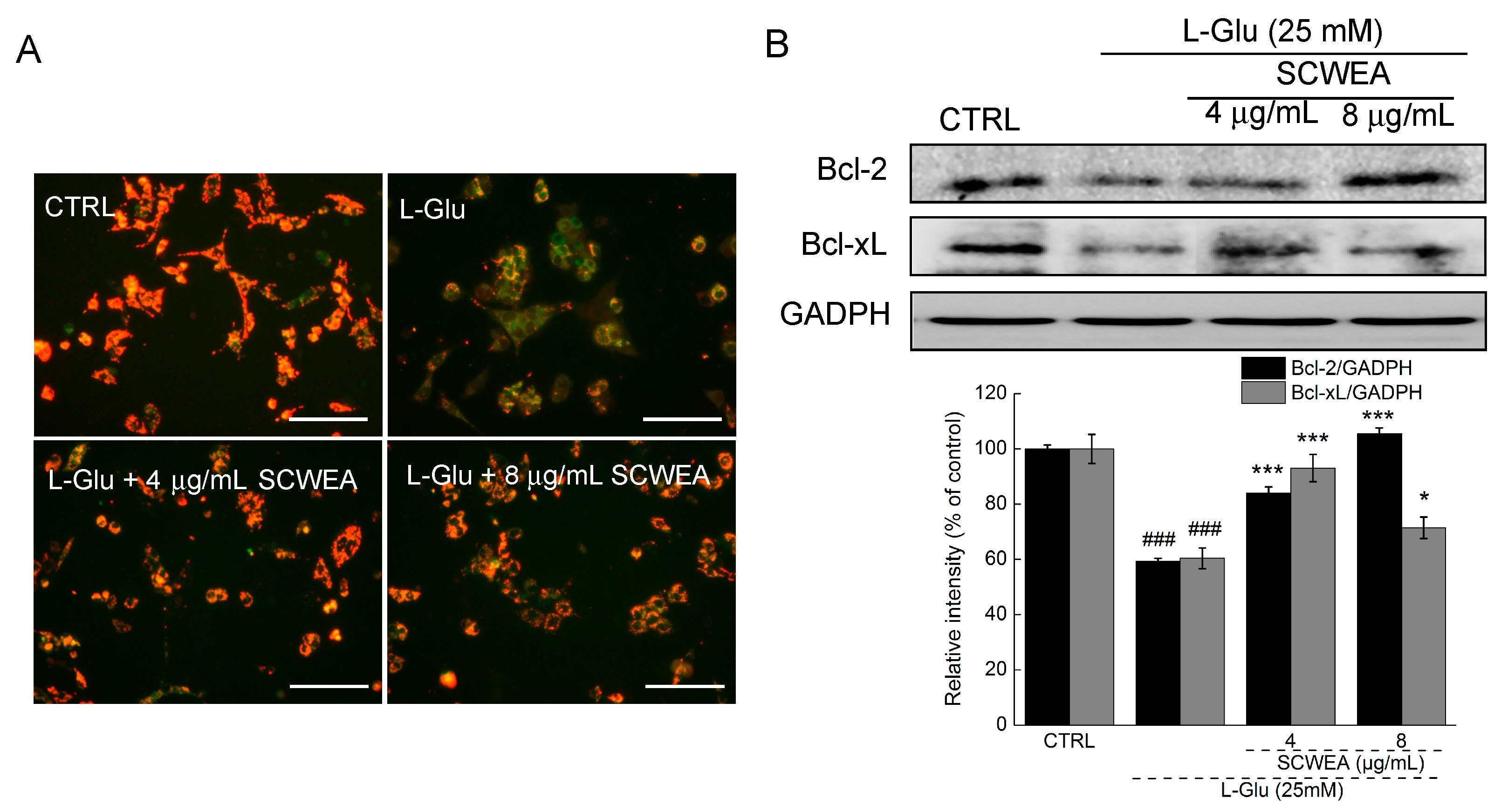
2.4. The Effects of SCWEA on Mitochondrial Function


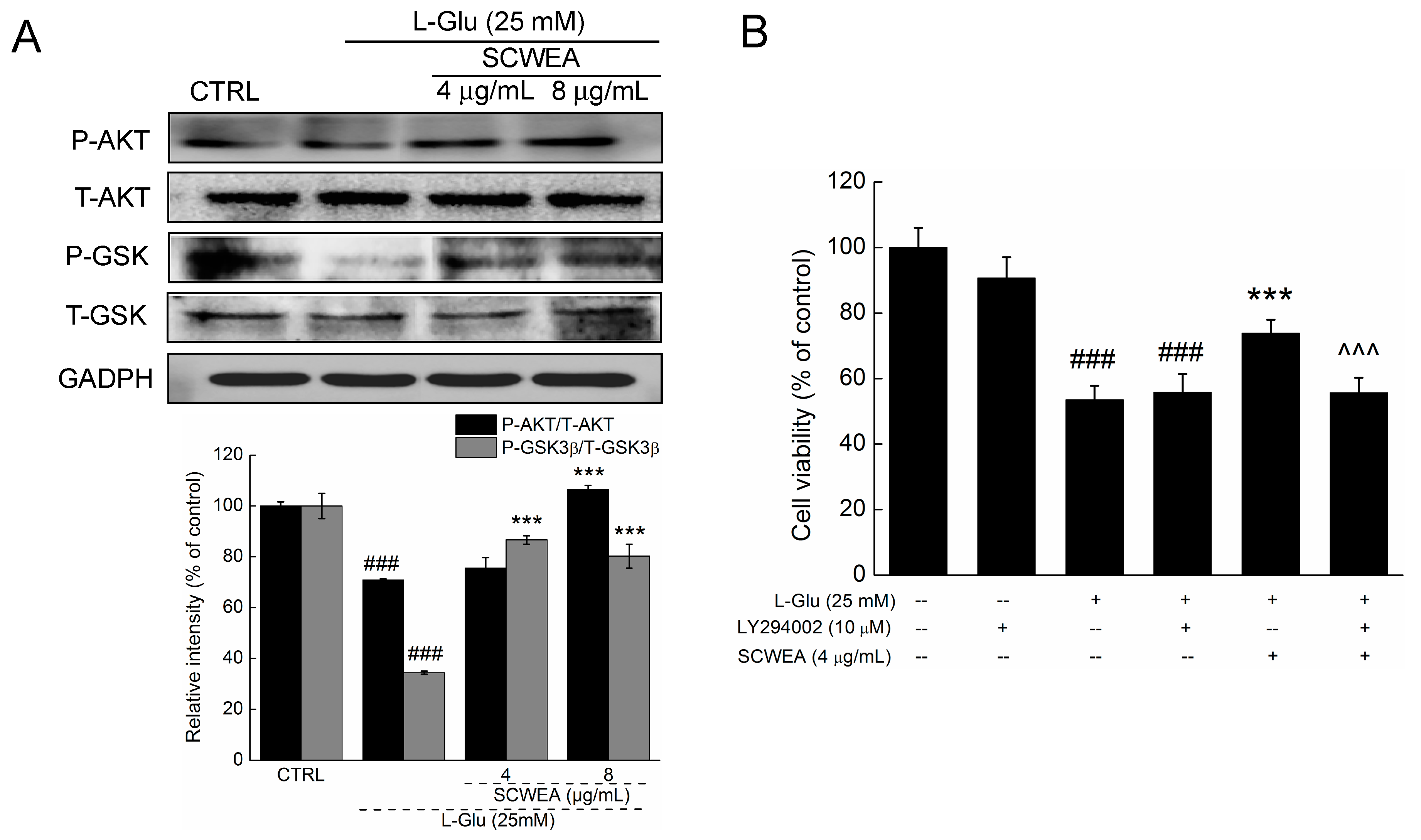
2.5. Protein Kinase B (AKT) Contributing to SCWEA-Mediated Neuro-Protective Effect

3. Discussion
4. Materials and Methods
4.1. Submerge Fermentation of Sparassis crispa and Crude Extract Preparation
4.2. Polysaccharide Purification and Characterization
4.2.1. Purification of Sparassis crispa Polysaccharides
4.2.2. Molecular Weight and Particle Size Measurements
4.2.3. Ultraviolet (UV) Spectra Measurement
4.2.4. Fourier Transform Infrared Spectroscopy (FTIR) Determination
4.2.5. Congo Red Test
4.2.6. Monosaccharides Analysis
4.2.7. Periodate Oxidation-Smith Degradation Reaction of Polysaccharides
4.3. Cell Culture
4.4. Cell Viability Analysis
4.5. Cellular Morphology Analysis
4.6. Measurement of ROS
4.7. Measurement of Intracellular Calcium Concentration ([Ca2+]i)
4.8. Mitochondrial Membrane Potential (MMP) Analysis
4.9. Western Blot
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix


References
- Karbowski, M.; Neutzner, A. Neurodegeneration as a consequence of failed mitochondrial maintenance. Acta Neuropathol. 2012, 123, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Rosello, A.; Warnes, G.; Meier, U.C. Cell death pathways and autophagy in the central nervous system and its involvement in neurodegeneration, immunity and central nervous system infection: To die or not to die—That is the question. Clin. Exp. Immunol. 2012, 168, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, H.; Xu, A.; Ross, T.; Bowler, E.; Hu, Y.; Lesnefsky, E.J. Inhibition of Bcl-2 sensitizes mitochondrial permeability transition pore (MPTP) opening in ischemia-damaged mitochondria. PLoS ONE 2015, 10, e0118834. [Google Scholar] [CrossRef] [PubMed]
- Fayard, E.; Xue, G.; Parcellier, A.; Bozulic, L.; Hemmings, B.A. Protein kinase B (PKB/AKT), a key mediator of the PI3K signaling pathway. Curr. Top. Microbiol. Immunol. 2010, 346, 31–56. [Google Scholar] [PubMed]
- McManus, M.J.; Murphy, M.P.; Franklin, J.L. Mitochondria-derived reactive oxygen species mediate caspase-dependent and -independent neuronal deaths. Mol. Cell. Neurosci. 2014, 63, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Erdmann, N.; Zhao, J.; Cao, Z.; Peng, H.; Zheng, J. HIV-infected macrophages mediate neuronal apoptosis through mitochondrial glutaminase. J. Neurochem. 2008, 105, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.S.; Hossain, M.A.; Park, S.C. Toxicological profiles of poisonous, edible, and medicinal mushrooms. Mycobiology 2014, 42, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.S.; Lai, S.W.; Lin, K.F.; Fang, J.N.; Yuen, W.H.; Chang, R.C. Characterization of polysaccharides from the flowers of nerium indicum and their neuroprotective effects. Int. J. Mol. Med. 2004, 14, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.S.; Wong, A.Y.; So, K.F.; Fang, J.N.; Yuen, W.H.; Chang, R.C. New polysaccharide from nerium indicum protects neurons via stress kinase signaling pathway. Brain Res. 2007, 1153, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.J.; Cheong, K.L.; Zhao, J.; Li, S.P. Chromatography in characterization of polysaccharides from medicinal plants and fungi. J. Sep. Sci. 2013, 36, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ohno, N.; Miura, N.N.; Nakajima, M.; Yadomae, T. Antitumor 1,3-β-glucan from cultured fruit body of sparassis crispa. Biol. Pharm. Bull. 2000, 23, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.H.; Kawaminami, H.; Ishibashi, K.; Miura, N.N.; Adachi, Y.; Ohno, N. Oral administration of soluble β-glucan preparation from the cauliflower mushroom, Sparassis crispa (higher basidiomycetes) modulated cytokine production in mice. Int. J. Med. Mushrooms 2013, 15, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Nameda, S.; Harada, T.; Miura, N.N.; Adachi, Y.; Yadomae, T.; Nakajima, M.; Ohno, N. Enhanced cytokine synthesis of leukocytes by a β-glucan preparation, SCG, extracted from a medicinal mushroom, sparassis crispa. Immunopharmacol. Immunotoxicol. 2003, 25, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Miura, N.N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. IFN-γ induction by SCG, 1,3-β-d-glucan from sparassis crispa, in DBA/2 mice in vitro. J. Interferon Cytokine Res. 2002, 22, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Lee, S.; Singh, T.S.; Choi, J.K.; Shin, T.Y.; Kim, S.H. Sparassis crispa suppresses mast cell-mediated allergic inflammation: Role of calcium, mitogen-activated protein kinase and nuclear factor-κB. Int. J. Mol. Med. 2012, 30, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Iwaoka, E.; Kubo, M.; Shibata, M.; Gao, M. Beneficial effect of Sparassis crispa on stroke through activation of AKT/eNOS pathway in brain of SHRSP. J. Nat. Med. 2011, 65, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Chaoui, D.; Faussat, A.M.; Majdak, P.; Tang, R.; Perrot, J.Y.; Pasco, S.; Klein, C.; Marie, J.P.; Legrand, O. Jc-1, a sensitive probe for a simultaneous detection of p-glycoprotein activity and apoptosis in leukemic cells. Cytom. Part B Clin. Cytom. 2006, 70, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Belagodu, A.; Reiter, R.J.; Ray, S.K.; Banik, N.L. Cytoprotective effects of melatonin on C6 astroglial cells exposed to glutamate excitotoxicity and oxidative stress. J. Pineal Res. 2008, 45, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Feissner, R.F.; Skalska, J.; Gaum, W.E.; Sheu, S.S. Crosstalk signaling between mitochondrial Ca2+ and ROS. Front. Biosci. 2009, 14, 1197–1218. [Google Scholar] [CrossRef]
- Bernardi, P.; Rasola, A. Calcium and cell death: The mitochondrial connection. Subcell. Biochem. 2007, 45, 481–506. [Google Scholar] [PubMed]
- Christophe, M.; Nicolas, S. Mitochondria: A target for neuroprotective interventions in cerebral ischemia-reperfusion. Curr. Pharm. Des. 2006, 12, 739–757. [Google Scholar] [CrossRef] [PubMed]
- Wallgren, M.; Lidman, M.; Pedersen, A.; Brannstrom, K.; Karlsson, B.G.; Grobner, G. Reconstitution of the anti-apoptotic Bcl-2 protein into lipid membranes and biophysical evidence for its detergent-driven association with the pro-apoptotic bax protein. PLoS ONE 2013, 8, e61452. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.M.; Tabellini, G.; Bressanin, D.; Ognibene, A.; Goto, K.; Cocco, L.; Evangelisti, C. The emerging multiple roles of nuclear AKT. Biochim. Biophys. Acta 2012, 1823, 2168–2178. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Preiss, F.; Brauns-Schubert, P.; Schlicher, L.; Charvet, C. GSK-3—At the crossroads of cell death and survival. J. Cell Sci. 2014, 127, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.S.; Zhu, B.; Guo, Z.K.; Huang, G.Z.; Wang, Z.; Chen, J.; Wei, X.J.; Li, Q. Bone marrow mesenchymal stem cell-derived microvesicles protect rat pheochromocytoma PC12 cells from glutamate-induced injury via a PI3K/AKT dependent pathway. Neurochem. Res. 2014, 39, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Y.; Xing, D. LPLI inhibits apoptosis upstream of Bax translocation via a GSK-3β-inactivation mechanism. J. Cell. Physiol. 2010, 224, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Petit-Paitel, A.; Brau, F.; Cazareth, J.; Chabry, J. Involvment of cytosolic and mitochondrial GSK-3β in mitochondrial dysfunction and neuronal cell death of MPTP/MPP-treated neurons. PLoS ONE 2009, 4, e5491. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Kim, Y.S.; Choi, I.Y.; Kim, S.W.; Kim, W.K. 25-hydroxycholesterol induces mitochondria-dependent apoptosis via activation of glycogen synthase kinase-3β in PC12 cells. Free Radic. Res. 2008, 42, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Spreux-Varoquaux, O.; Bensimon, G.; Lacomblez, L.; Salachas, F.; Pradat, P.F.; le Forestier, N.; Marouan, A.; Dib, M.; Meininger, V. Glutamate levels in cerebrospinal fluid in amyotrophic lateral sclerosis: A reappraisal using a new HPLC method with coulometric detection in a large cohort of patients. J. Neurol Sci. 2002, 193, 73–78. [Google Scholar] [CrossRef]
- Wang, D.; Tan, Q.R.; Zhang, Z.J. Neuroprotective effects of paeoniflorin, but not the isomer albiflorin, are associated with the suppression of intracellular calcium and calcium/calmodulin protein kinase II in PC12 cells. J. Mol. Neurosci. 2013, 51, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cheong, Y.K.; Wang, H.; Ren, G.; Yang, Z. Neuroprotective effects of etidronate and 2,3,3-trisphosphonate against glutamate-induced toxicity in PC12 cells. Neurochem. Res. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhu, D.J.; Xu, D.B.; Wu, J.; Bian, X.Y. A study on cordyceps militaris polysaccharide purification, composition and activity analysis. Afr. J. Biotechnol. 2008, 7, 4004–4009. [Google Scholar]
- Zhang, D.Y.; Li, S.J.; Xiong, Q.P.; Jiang, C.X.; Lai, X.P. Extraction, characterization and biological activities of polysaccharides from amomum villosum. Carbohydr. Polym. 2013, 95, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Hu, S.; Liu, C.G.; Meng, Q.F.; Song, J.; Lu, J.H.; Cheng, Y.K.; Gao, C.H.; Liu, Y.; Wang, D.; et al. Purification of polysaccharides from cordyceps militaris and their anti-hypoxic effect. Mol. Med. Rep. 2015, 11, 1312–1317. [Google Scholar] [PubMed]
- Wang, Z.B.; Pei, J.J.; Ma, H.L.; Cai, P.F.; Yan, J.K. Effect of extraction media on preliminary characterizations and antioxidant activities of phellinus linteus polysaccharides. Carbohydr. Polym. 2014, 109, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Linker, A.; Evans, L.R.; Impallomeni, G. The structure of a polysaccharide from infectious strains of Burkholderia cepacia. Carbohydr. Res. 2001, 335, 45–54. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Wang, D.; Zhang, J.; Du, M.; Cheng, Y.; Liu, Y.; Zhang, N.; Wang, D.; Wu, Y. Mitochondria Related Pathway Is Essential for Polysaccharides Purified from Sparassis crispa Mediated Neuro-Protection against Glutamate-Induced Toxicity in Differentiated PC12 Cells. Int. J. Mol. Sci. 2016, 17, 133. https://doi.org/10.3390/ijms17020133
Hu S, Wang D, Zhang J, Du M, Cheng Y, Liu Y, Zhang N, Wang D, Wu Y. Mitochondria Related Pathway Is Essential for Polysaccharides Purified from Sparassis crispa Mediated Neuro-Protection against Glutamate-Induced Toxicity in Differentiated PC12 Cells. International Journal of Molecular Sciences. 2016; 17(2):133. https://doi.org/10.3390/ijms17020133
Chicago/Turabian StyleHu, Shuang, Di Wang, Junrong Zhang, Mengyan Du, Yingkun Cheng, Yan Liu, Ning Zhang, Di Wang, and Yi Wu. 2016. "Mitochondria Related Pathway Is Essential for Polysaccharides Purified from Sparassis crispa Mediated Neuro-Protection against Glutamate-Induced Toxicity in Differentiated PC12 Cells" International Journal of Molecular Sciences 17, no. 2: 133. https://doi.org/10.3390/ijms17020133