
Ginsenoside-Rd Promotes Neurite Outgrowth of PC12 Cells through MAPK/ERK- and PI3K/AKT-Dependent Pathways

Abstract
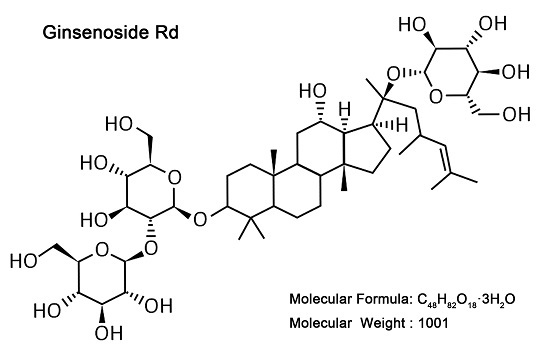
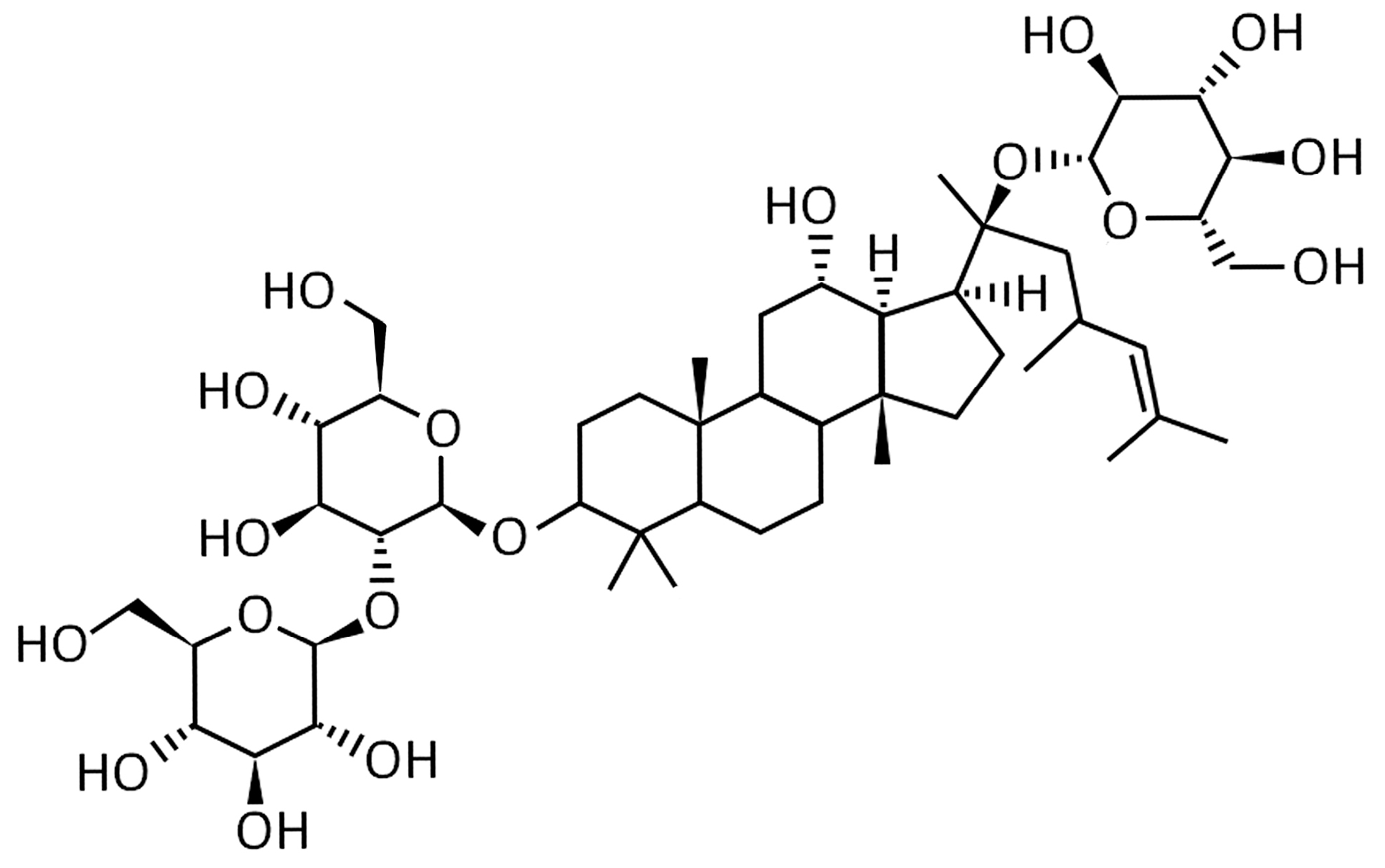
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction

2. Results
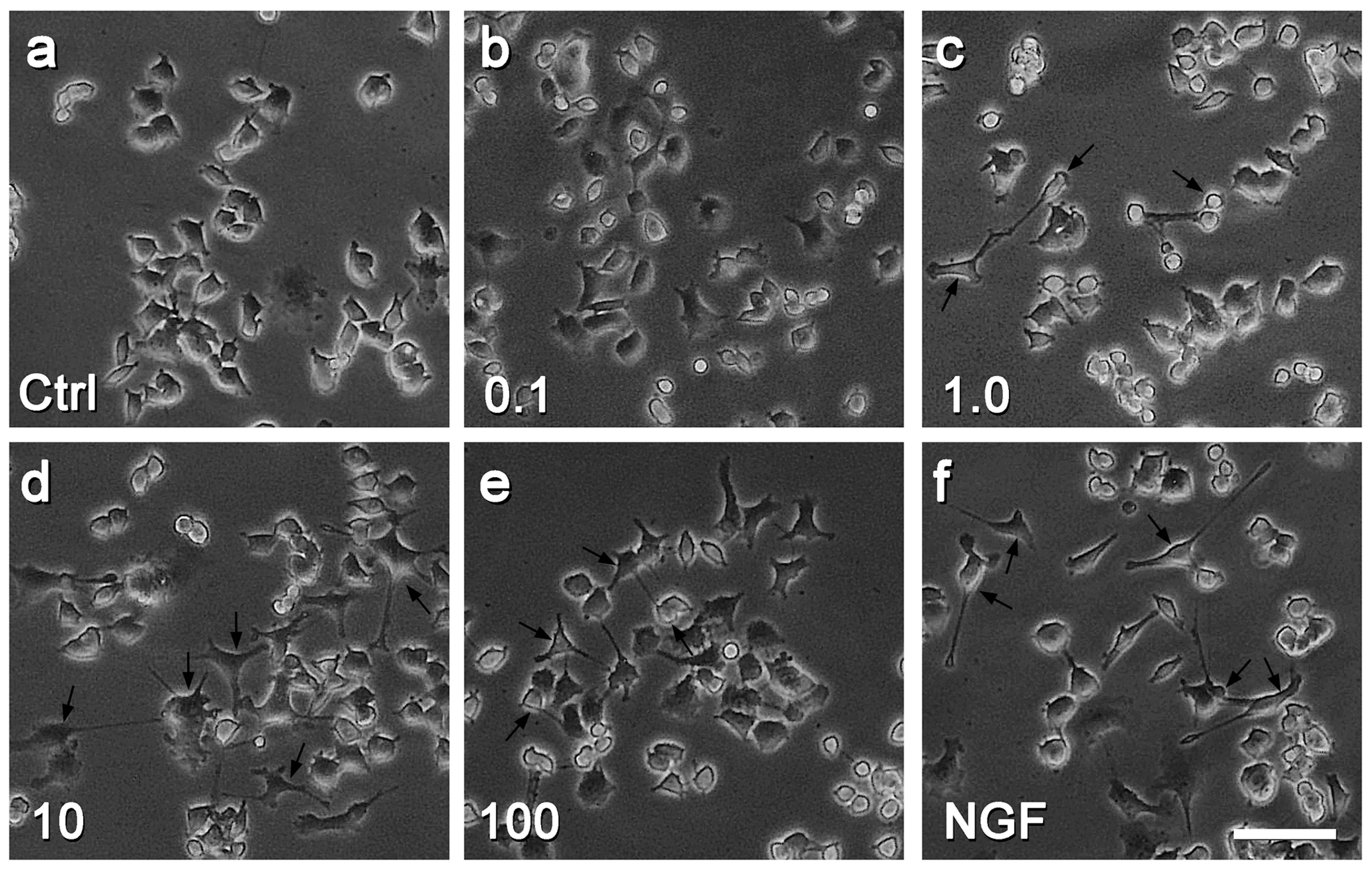
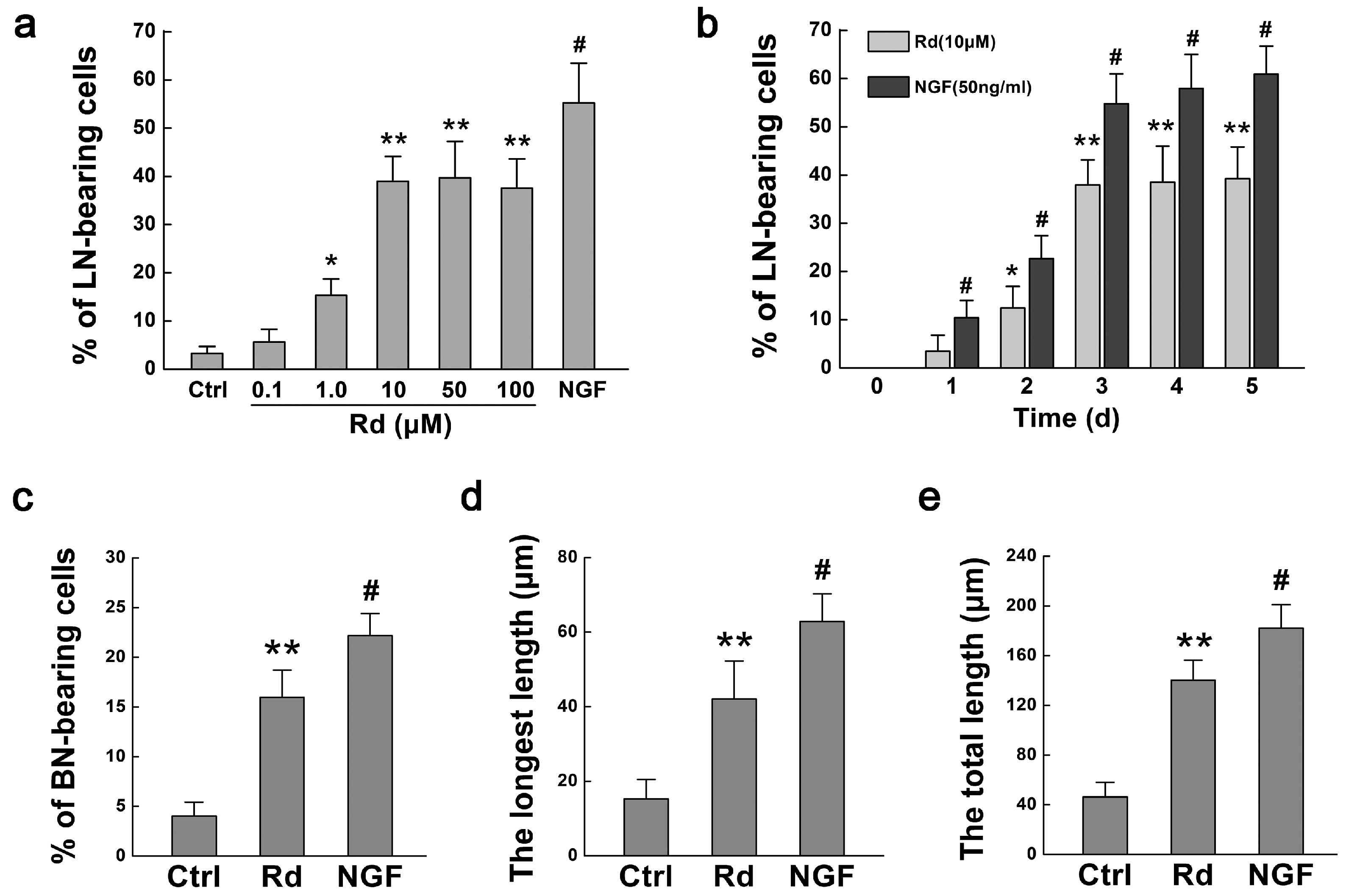
2.1. Rd Promotes Neurite Outgrowth of PC12 Cells


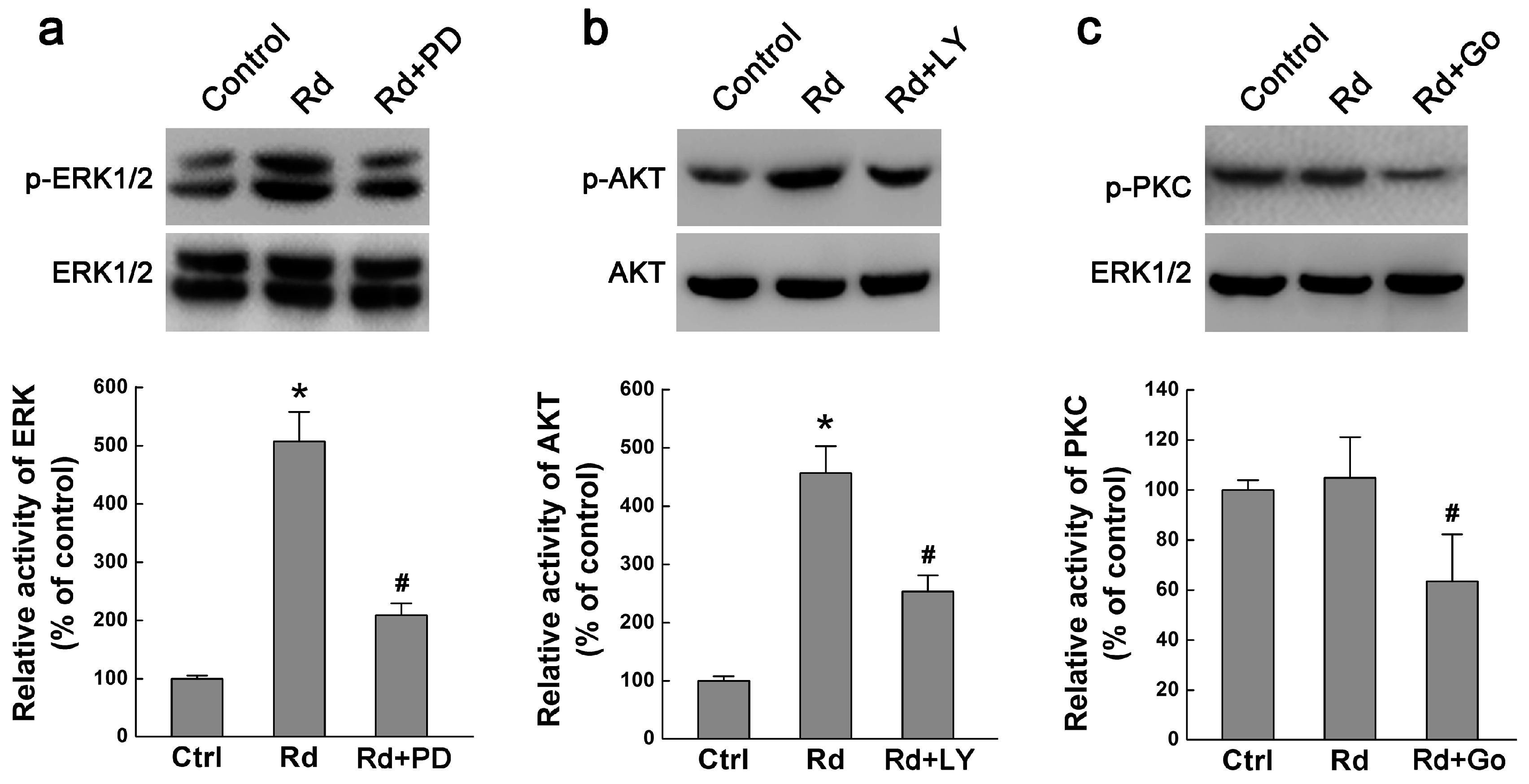
2.2. Rd Activates MAPK/ERK and PI3K/AKT but Not PKC Signaling Pathways

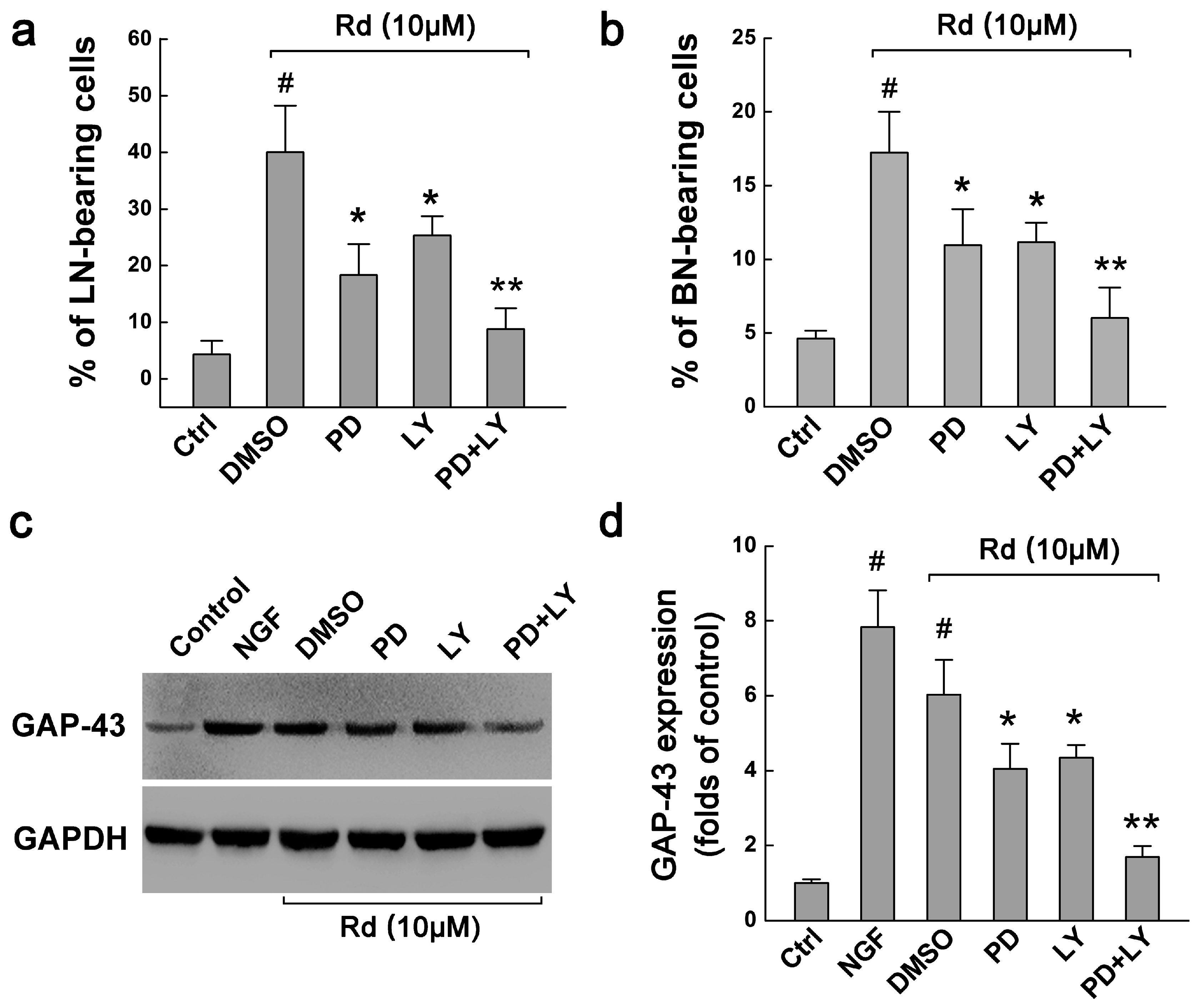
2.3. ERK and ARK Are Involved in Rd-Induced Neurite Outgrowth of PC12 Cells

2.4. Rd Upregulates ERK- and AKT-Mediated GAP-43 Expression to Promote Neurite Outgrowth
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Grouping
4.4. Evaluation of Neurite Outgrowth
4.5. Western Blot
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, C.F.; Chiou, W.F.; Zhang, J.T. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin. 2008, 29, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B. Pharmacological activity of sanchi ginseng (Panax notoginseng). J. Pharm. Pharmacol. 2006, 58, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99. [Google Scholar] [PubMed]
- Liu, X.; Wang, L.; Wen, A.; Yang, J.; Yan, Y.; Song, Y.; Ren, H.; Wu, Y.; Li, Z.; Chen, W.; et al. Ginsenoside-Rd improves outcome of acute ischaemic stroke—A randomized, double-blind, placebo-controlled, multicenter trial. Eur. J. Neurol. 2012, 19, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, J.; Wang, L.; Song, Y.; Yang, J.; Yan, Y.; Ren, H.; Zhao, G. Efficacy and safety of ginsenoside-Rd for acute ischaemic stroke: A randomized, double-blind, placebo-controlled, phase II multicenter trial. Eur. J. Neurol. 2009, 16, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Yang, Q.; Kong, X.; Han, J.; Zhang, X.; Zhang, Y.; Li, P.; Liu, J.; Shi, M.; Xiong, L.; et al. Ginsenoside Rd attenuates early oxidative damage and sequential inflammatory response after transient focal ischemia in rats. Neurochem. Int. 2011, 58, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Zhang, X.; Kong, X.; Han, J.; Yang, Q.; Zhang, Y.; Chen, Y.; Li, P.; Liu, J.; Shi, M.; et al. Ginsenoside Rd attenuates mitochondrial dysfunction and sequential apoptosis after transient focal ischemia. Neuroscience 2011, 178, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Kong, X.; Yang, Q.; Zhang, Y.; Han, J.; Zhao, G. Ginsenoside Rd attenuates redox imbalance and improves stroke outcome after focal cerebral ischemia in aged mice. Neuropharmacology 2011, 61, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Kong, X.; Yang, Q.; Zhang, Y.; Han, J.; Li, P.; Xiong, L.; Zhao, G. Ginsenoside Rd in experimental stroke: Superior neuroprotective efficacy with a wide therapeutic window. Neurotherapeutics 2011, 8, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Wu, Z.; Yang, F.; Zhao, H.; Liu, X.; Deng, Y.; Shi, M.; Zhao, G. Ginsenoside Rd blocks AIF mitochondrio-nuclear translocation and NF-κB nuclear accumulation by inhibiting poly(ADP-ribose) polymerase-1 after focal cerebral ischemia in rats. Neurol. Sci. 2013, 34, 2101–2106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, M.; Bjoras, M.; Wang, W.; Zhang, G.; Han, J.; Liu, Z.; Zhang, Y.; Wang, B.; Chen, J.; et al. Ginsenoside Rd promotes glutamate clearance by up-regulating glial glutamate transporter GLT-1 via PI3K/AKT and ERK1/2 pathways. Front. Pharmacol. 2013, 4, 152. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Han, J.; Kong, X.; Zhao, L.; Cao, R.; Rao, Z.; Zhao, G. Protective effects of ginsenoside Rd on PC12 cells against hydrogen peroxide. Biol. Pharm. Bull. 2008, 31, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Li, N.; Han, J.; Kong, X.; Cao, R.; Rao, Z.; Zhao, G. Neuroprotective effects of ginsenoside Rd against oxygen-glucose deprivation in cultured hippocampal neurons. Neurosci. Res. 2009, 64, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Du, F.; Shi, M.; Ye, R.; Cheng, H.; Han, J.; Ma, L.; Cao, R.; Rao, Z.; Zhao, G. Ginsenoside Rd protects neurons against glutamate-induced excitotoxicity by inhibiting Ca2+ influx. Cell. Mol. Neurobiol. 2012, 32, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.E.; Aminoltejari, K.; Erb, H.; Winship, I.R.; Murphy, T.H. In vivo voltage-sensitive dye imaging in adult mice reveals that somatosensory maps lost to stroke are replaced over weeks by new structural and functional circuits with prolonged modes of activation within both the peri-infarct zone and distant sites. J. Neurosci. 2009, 29, 1719–1734. [Google Scholar] [CrossRef] [PubMed]
- Dancause, N.; Barbay, S.; Frost, S.B.; Plautz, E.J.; Chen, D.; Zoubina, E.V.; Stowe, A.M.; Nudo, R.J. Extensive cortical rewiring after brain injury. J. Neurosci. 2005, 25, 10167–10179. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Overman, J.J.; Katsman, D.; Kozlov, S.V.; Donnelly, C.J.; Twiss, J.L.; Giger, R.J.; Coppola, G.; Geschwind, D.H.; Carmichael, S.T. An age-related sprouting transcriptome provides molecular control of axonal sprouting after stroke. Nat. Neurosci. 2010, 13, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Overman, J.J.; Clarkson, A.N.; Wanner, I.B.; Overman, W.T.; Eckstein, I.; Maguire, J.L.; Dinov, I.D.; Toga, A.W.; Carmichael, S.T. A role for ephrin-A5 in axonal sprouting, recovery, and activity-dependent plasticity after stroke. Proc. Natl. Acad. Sci. USA. 2012, 109, E2230–E2239. [Google Scholar] [CrossRef] [PubMed]
- Bouquet, C.; Nothias, F. Molecular mechanisms of axonal growth. Adv. Exp. Med. Biol. 2007, 621, 1–16. [Google Scholar] [PubMed]
- Schuldiner, O.; Yaron, A. Mechanisms of developmental neurite pruning. Cell. Mol. Life Sci. 2015, 72, 101–119. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.Y.; Tang, B.L. Axonal regeneration in adult CNS neurons-signaling molecules and pathways. J. Neurochem. 2006, 96, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.D.; Mackinnon, S.E. Pathways regulating modality-specific axonal regeneration in peripheral nerve. Exp. Neurol. 2015, 265, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, L.I.; Routtenberg, A. GAP-43: An intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997, 20, 84–91. [Google Scholar] [CrossRef]
- Korshunova, I.; Mosevitsky, M. Role of the growth-associated protein GAP-43 in NCAM-mediated neurite outgrowth. Adv. Exp. Med. Biol. 2010, 663, 169–182. [Google Scholar] [PubMed]
- Cui, Q. Actions of neurotrophic factors and their signaling pathways in neuronal survival and axonal regeneration. Mol. Neurobiol. 2006, 33, 155–179. [Google Scholar] [CrossRef]
- Hinman, J.D. The back and forth of axonal injury and repair after stroke. Curr. Opin. Neurol. 2014, 27, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Raivich, G.; Makwana, M. The making of successful axonal regeneration: Genes, molecules and signal transduction pathways. Brain Res. Rev. 2007, 53, 287–311. [Google Scholar] [CrossRef] [PubMed]
- Heiser, J.H.; Schuwald, A.M.; Sillani, G.; Ye, L.; Muller, W.E.; Leuner, K. TRPC6 channel-mediated neurite outgrowth in PC12 cells and hippocampal neurons involves activation of RAS/MEK/ERK, PI3K, and CAMKIV signaling. J. Neurochem. 2013, 127, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Sato, A.; Yamashita, T. Brimonidine promotes axon growth after optic nerve injury through ERK phosphorylation. Cell Death Dis. 2013, 4, e763. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, C.; Li, D.; Wang, Y.; Shao, W.; You, Y.; Peng, J.; Zhang, X.; Lu, L.; Shen, X. BIG1, a brefeldin A-inhibited guanine nucleotide-exchange protein regulates neurite development via PI3K-AKT and ERK signaling pathways. Neuroscience 2013, 254, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Cheng, Y.Y.; Fan, W.; Yang, C.B.; Ye, S.F.; Cui, W.; Wei, W.; Lao, L.X.; Cai, J.; Han, Y.F.; et al. Botanical drug puerarin coordinates with nerve growth factor in the regulation of neuronal survival and neuritogenesis via activating ERK1/2 and PI3K/AKT signaling pathways in the neurite extension process. CNS Neurosci. Ther. 2015, 21, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Dong, X.; Huang, X.; Huang, X.J.; Liu, H.; Wang, Y.; Ye, W.C.; Shi, L. A natural diarylheptanoid promotes neuronal differentiation via activating ERK and PI3K-AKT dependent pathways. Neuroscience 2015, 303, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lam, J.S.; Zhao, H.; Wang, J.; Chan, S.O. Localization of protein kinase c isoforms in the optic pathway of mouse embryos and their role in axon routing at the optic chiasm. Brain Res. 2014, 1575, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Bjornstrom, K.; Turina, D.; Strid, T.; Sundqvist, T.; Eintrei, C. Orexin A inhibits propofol-induced neurite retraction by a phospholipase D/protein kinase Cε-dependent mechanism in neurons. PLoS ONE 2014, 9, e97129. [Google Scholar] [CrossRef] [PubMed]
- Greer, Y.E.; Fields, A.P.; Brown, A.M.; Rubin, J.S. Atypical protein kinase Cι is required for Wnt3a-dependent neurite outgrowth and binds to phosphorylated Dishevelled 2. J. Biol. Chem. 2013, 288, 9438–9446. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Cui, X.; Sakaguchi, N.; Akashi, M. Ginsenoside Rd prevents and rescues rat intestinal epithelial cells from irradiation-induced apoptosis. Food Chem. Toxicol. 2008, 46, 3080–3089. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Wang, X.; Lau, W.; Wang, Y.; Xing, Y.; Zhang, X.; Ma, X.; Gao, F. Ginsenoside Rd attenuates myocardial ischemia/reperfusion injury via AKT/GSK-3β signaling and inhibition of the mitochondria-dependent apoptotic pathway. PLoS ONE 2013, 8, e70956. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, M.; Ye, R.; Wang, W.; Liu, X.; Zhang, G.; Han, J.; Zhang, Y.; Wang, B.; Zhao, J.; et al. Ginsenoside Rd attenuates Tau protein phosphorylation via the PI3K/AKT/GSK-3β pathway after transient forebrain ischemia. Neurochem. Res. 2014, 39, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.S.; Wang, J.F.; He, B.R.; Cui, Y.S.; Fang, X.Y.; Ni, J.L.; Chen, J.; Wang, K.Z. Ginsenoside Rd attenuates mitochondrial permeability transition and cytochrome c release in isolated spinal cord mitochondria: Involvement of kinase-mediated pathways. Int. J. Mol. Sci. 2014, 15, 9859–9877. [Google Scholar] [CrossRef] [PubMed]
- Yankner, B.A.; Benowitz, L.I.; Villa-Komaroff, L.; Neve, R.L. Transfection of PC12 cells with the human GAP-43 gene: Effects on neurite outgrowth and regeneration. Brain Res. Mol. Brain Res. 1990, 7, 39–44. [Google Scholar] [CrossRef]
- Shea, T.B.; Perrone-Bizzozero, N.I.; Beermann, M.L.; Benowitz, L.I. Phospholipid-mediated delivery of anti-GAP-43 antibodies into neuroblastoma cells prevents neuritogenesis. J. Neurosci. 1991, 11, 1685–1690. [Google Scholar] [PubMed]
- Huffaker, T.; Corcoran, T.; Wagner, J.A. Adenosine inhibits cell division and promotes neurite extension in PC12 cells. J. Cell. Physiol. 1984, 120, 188–196. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.-D.; Xia, F.; Lin, X.-M.; Duan, K.-L.; Wang, F.; Lu, Q.-L.; Cao, H.; Qian, Y.-H.; Shi, M. Ginsenoside-Rd Promotes Neurite Outgrowth of PC12 Cells through MAPK/ERK- and PI3K/AKT-Dependent Pathways. Int. J. Mol. Sci. 2016, 17, 177. https://doi.org/10.3390/ijms17020177
Wu S-D, Xia F, Lin X-M, Duan K-L, Wang F, Lu Q-L, Cao H, Qian Y-H, Shi M. Ginsenoside-Rd Promotes Neurite Outgrowth of PC12 Cells through MAPK/ERK- and PI3K/AKT-Dependent Pathways. International Journal of Molecular Sciences. 2016; 17(2):177. https://doi.org/10.3390/ijms17020177
Chicago/Turabian StyleWu, Song-Di, Feng Xia, Xue-Mei Lin, Kang-Li Duan, Fang Wang, Qing-Li Lu, Huan Cao, Yi-Hua Qian, and Ming Shi. 2016. "Ginsenoside-Rd Promotes Neurite Outgrowth of PC12 Cells through MAPK/ERK- and PI3K/AKT-Dependent Pathways" International Journal of Molecular Sciences 17, no. 2: 177. https://doi.org/10.3390/ijms17020177