
Inhibition of IFN-γ-Induced Nitric Oxide Dependent Antimycobacterial Activity by miR-155 and C/EBPβ

Abstract
:
1. Introduction
2. Results
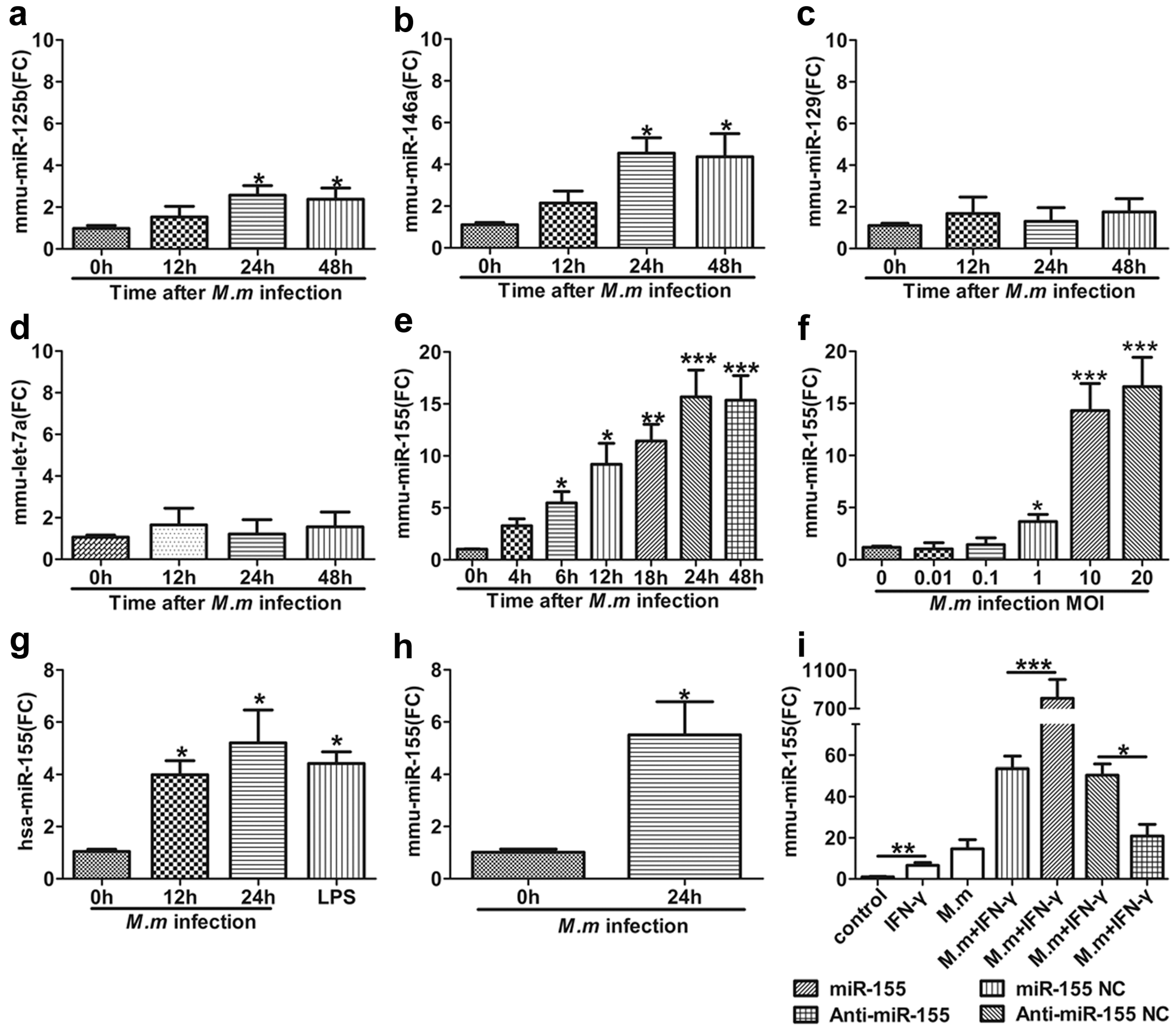
2.1. M.m-Mediated Induction of miR-155 (MicroRNA-155) in Macrophage
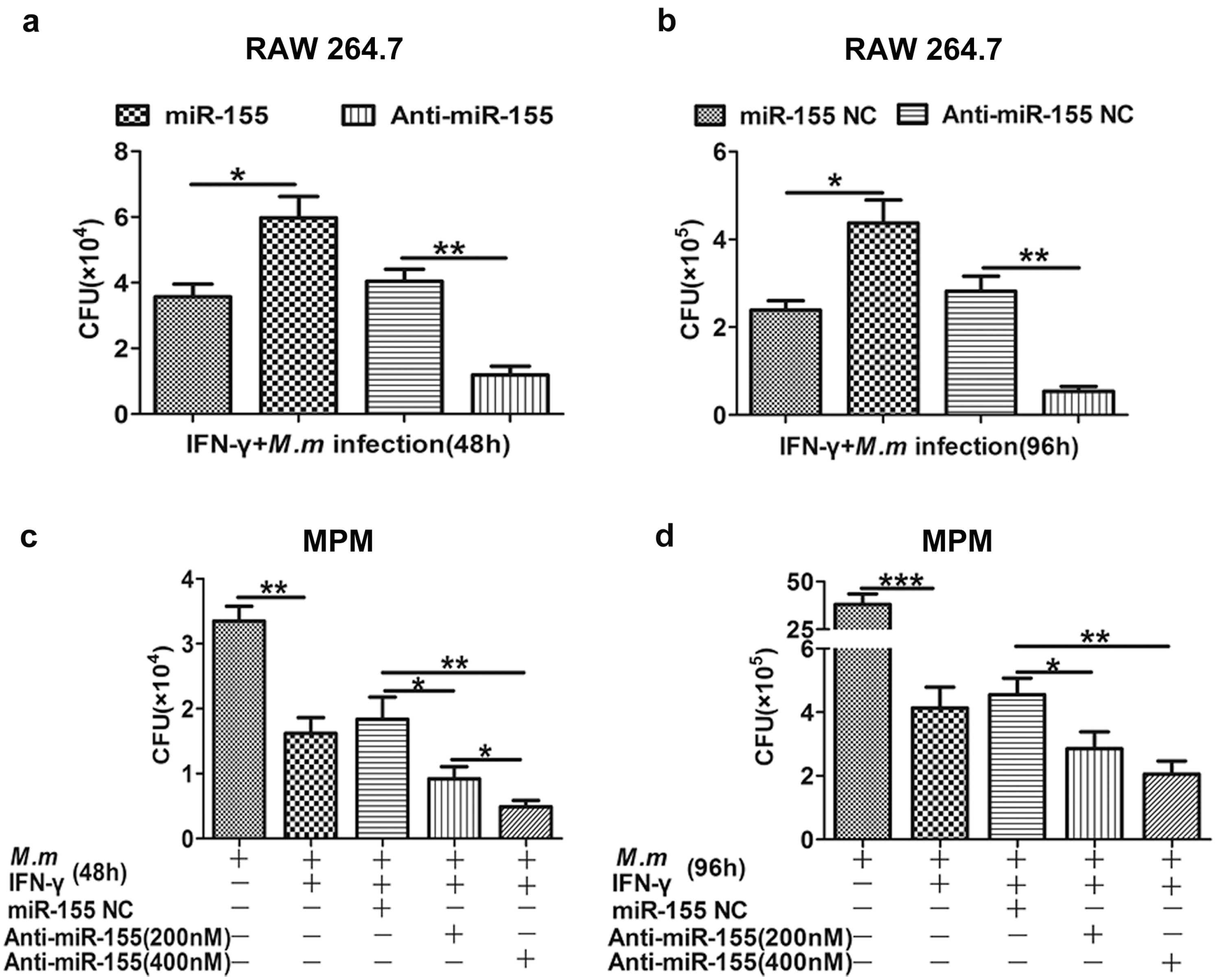
2.2. Effects of miR-155 on Innate Bactericidal Activity
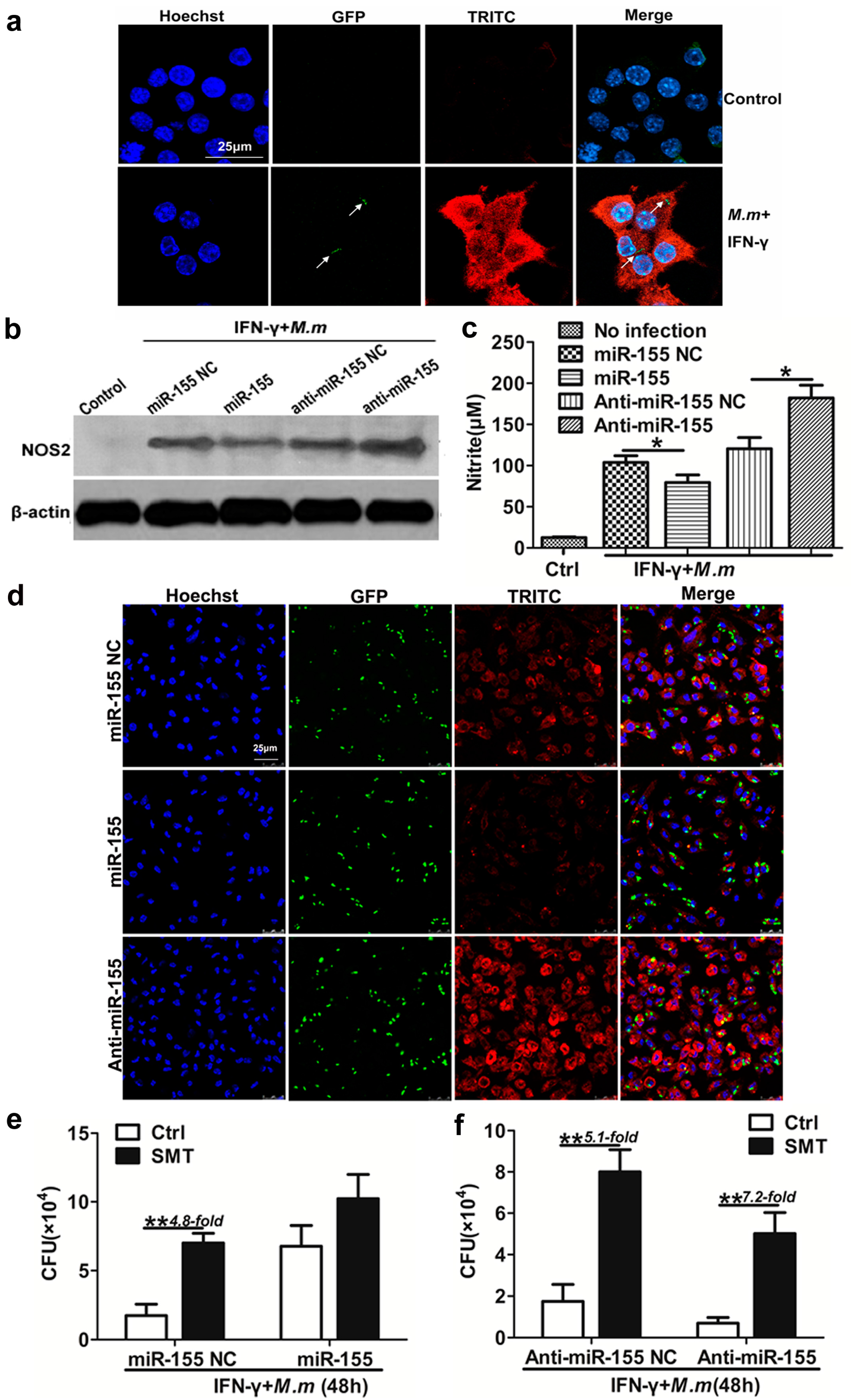
2.3. Role of Nitric Oxide (NO) in miR-155 Promotes the Survival of Intracellular Mycobacteria
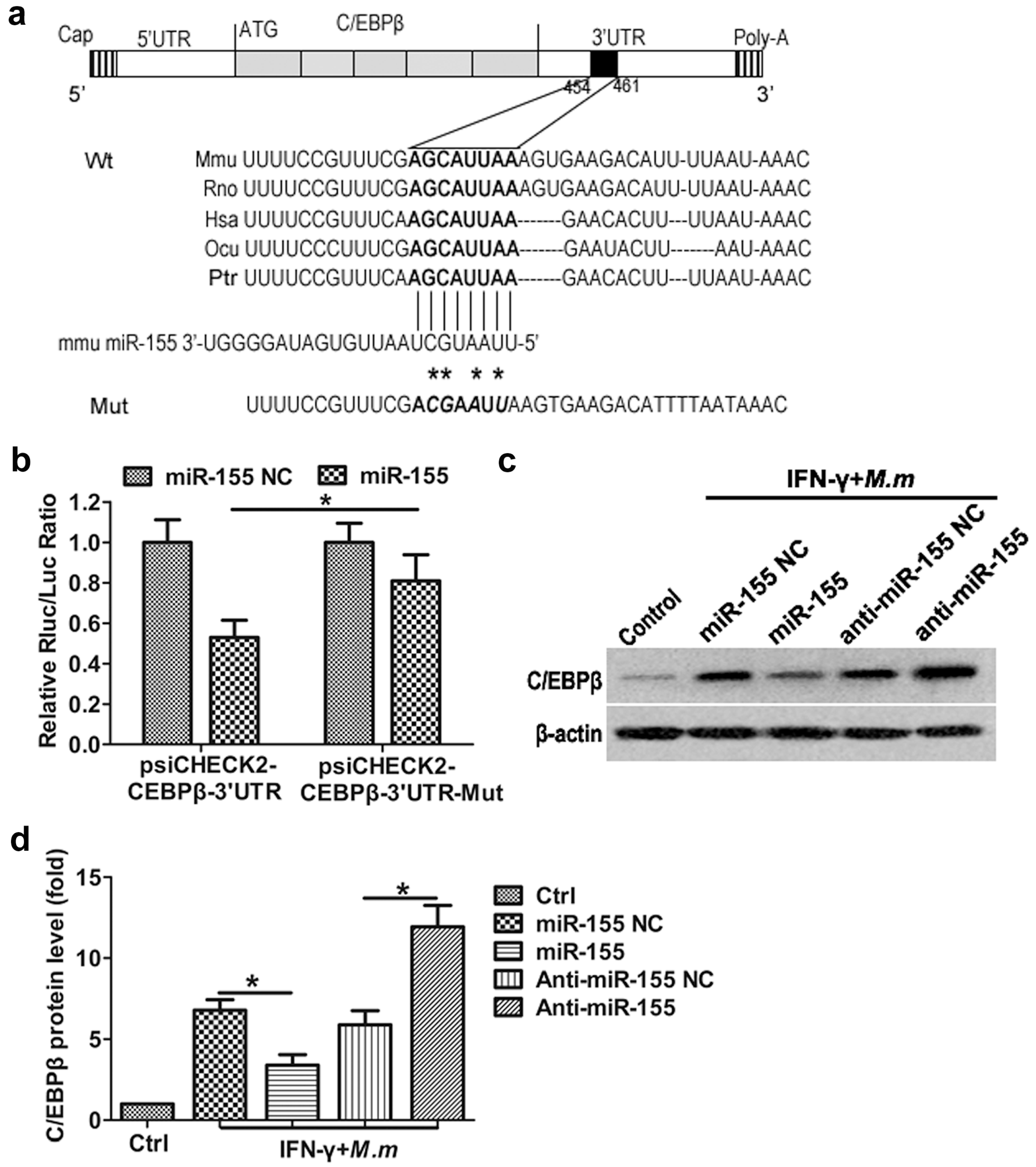
2.4. Regulation of the CCAAT/Enhancer Binding Protein β (C/EBPβ) by miR-155
2.5. Role of C/EBPβ in Bactericidal Activities by Macrophages
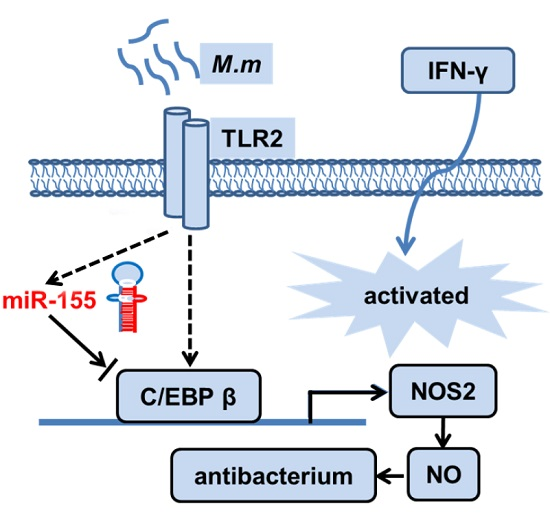
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Bacterial Strains
4.2. Knock down of C/EBPβ
4.3. RNA Isolation and qRT-PCR
4.4. Protein Expression
4.5. Immunofluorescence
4.6. 3′UTR Luciferase Reporter Assays
4.7. Griess Test
4.8. Intracellular Survival Assays
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Russell, D.G. Mycobacterium tuberculosis: Here today, and here tomorrow. Nat. Rev. Mol. Cell Biol. 2001, 2, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Herget, T.; Klebl, B.; Ullrich, A. Interplay between mycobacteria and host signalling pathways. Nat. Rev. Microbiol. 2004, 2, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Krutzik, S.R.; Modlin, R.L. The role of Toll-like receptors in combating mycobacteria. Semin. Immunol. 2004, 16, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Schulte, L.; Vogel, J. The mammalian microRNA response to bacterial infections. RNA Biol. 2012, 9, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, D.; Boldin, M.P.; O’Connell, R.M.; Rao, D.S.; Taganov, K.D. MicroRNAs: New regulators of immune cell development and function. Nat. Immunol. 2008, 9, 839–845. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Chaudhuri, A.A.; Rao, D.S.; Baltimore, D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc. Natl. Acad. Sci. USA 2009, 106, 7113–7118. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Kang, Y.; Zhang, H.; Piao, Z.; Yin, H.; Diao, R.; Xia, J.; Shi, L. Akt1-mediated regulation of macrophage polarization in a murine model of Staphylococcus aureus pulmonary infection. J. Infect. Dis. 2013, 208, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.J.; Ravneberg, D.H.; Clay, C.D.; Piper-Hunter, M.G.; Marsh, C.B.; Elton, T.S.; Gunn, J.S.; Amer, A.; Kanneganti, T.D.; Schlesinger, L.S.; et al. MiR-155 induction by F. novicida but not the virulent F. tularensis results in SHIP down-regulation and enhanced pro-inflammatory cytokine response. PLoS ONE 2009, 4, e8508. [Google Scholar] [CrossRef] [PubMed]
- Oertli, M.; Engler, D.B.; Kohler, E.; Koch, M.; Meyer, T.F.; Muller, A. MicroRNA-155 is essential for the T cell-mediated control of Helicobacter pylori infection and for the induction of Chronic Gastritis and Colitis. J. Immunol. 2011, 187, 3578–3586. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Liu, Z.; Li, B.S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.D.; et al. Induction of microRNA-155 during Helicobacter pylori infection and its negative regulatory role in the inflammatory response. J. Infect. Dis. 2009, 200, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Okoye, I.S.; Czieso, S.; Ktistaki, E.; Roderick, K.; Coomes, S.M.; Pelly, V.S.; Kannan, Y.; Perez-Lloret, J.; Zhao, J.L.; Baltimore, D.; et al. Transcriptomics identified a critical role for Th2 cell-intrinsic miR-155 in mediating allergy and antihelminth immunity. Proc. Natl. Acad. Sci. USA 2014, 111, E3081–E3090. [Google Scholar] [CrossRef] [PubMed]
- Ghorpade, D.S.; Leyland, R.; Kurowska-Stolarska, M.; Patil, S.A.; Balaji, K.N. MicroRNA-155 is required for Mycobacterium bovis BCG-mediated apoptosis of macrophages. Mol. Cell. Biol. 2012, 32, 2239–2253. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Halder, P.; Sahu, S.K.; Kumar, M.; Kumari, M.; Jana, K.; Ghosh, Z.; Sharma, P.; Kundu, M.; Basu, J. Identification of a novel role of ESAT-6-dependent miR-155 induction during infection of macrophages with Mycobacterium tuberculosis. Cell. Microbiol. 2012, 14, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Holla, S.; Kurowska-Stolarska, M.; Bayry, J.; Balaji, K.N. Selective inhibition of IFNG-induced autophagy by Mir155- and Mir31-responsive WNT5A and SHH signaling. Autophagy 2014, 10, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Cambier, C.J.; Takaki, K.K.; Larson, R.P.; Hernandez, R.E.; Tobin, D.M.; Urdahl, K.B.; Cosma, C.L.; Ramakrishnan, L. Mycobacteria manipulate macrophage recruitment through coordinated use of membrane lipids. Nature 2014, 505, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Sun, X.; Shao, X.; Cheng, C.; Feng, J.; Sun, W.; Gu, D.; Liu, W.; Xu, F.; Duan, Y. Macrophage-microglia networks drive M1 microglia polarization after mycobacterium infection. Inflammation 2015, 38, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, M.V.; Ni, B.; Morris, J.D.; Brooks, M.N.; Carlson, T.K.; Bakthavachalu, B.; Schoenberg, D.R.; Torrelles, J.B.; Schlesinger, L.S. Mycobacterium tuberculosis lipomannan blocks TNF biosynthesis by regulating macrophage MAPK-activated protein kinase 2 (MK2) and microRNA miR-125b. Proc. Natl. Acad. Sci. USA 2011, 108, 17408–17413. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xiao, B.; Tang, B.; Li, B.; Li, N.; Zhu, E.; Guo, G.; Gu, J.; Zhuang, Y.; Liu, X.; et al. Up-regulated microRNA-146a negatively modulate Helicobacter pylori-induced inflammatory response in human gastric epithelial cells. Microbes Infect. 2010, 12, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.N.; Eulalio, A.; Mollenkopf, H.J.; Reinhardt, R.; Vogel, J. Analysis of the host microRNA response to Salmonella uncovers the control of major cytokines by the let-7 family. Embo J. 2011, 30, 1977–1989. [Google Scholar] [CrossRef] [PubMed]
- Hasler, R.; Jacobs, G.; Till, A.; Grabe, N.; Cordes, C.; Nikolaus, S.; Lao, K.; Schreiber, S.; Rosenstiel, P. Microbial pattern recognition causes distinct functional micro-RNA signatures in primary human monocytes. PLoS ONE 2012, 7, e31151. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Santosuosso, M.; Ngai, P.; Zganiacz, A.; Xing, Z. Activation of CD8 T cells by mycobacterial vaccination protects against pulmonary tuberculosis in the absence of CD4 T cells. J. Immunol. 2004, 173, 4590–4597. [Google Scholar] [CrossRef] [PubMed]
- Tascon, R.E.; Stavropoulos, E.; Lukacs, K.V.; Colston, M.J. Protection against Mycobacterium tuberculosis infection by CD8+ T cells requires the production of γ interferon. Infect. Immun. 1998, 66, 830–834. [Google Scholar] [PubMed]
- Cooper, A.M.; Dalton, D.K.; Stewart, T.A.; Griffin, J.P.; Russell, D.G.; Orme, I.M. Disseminated tuberculosis in interferon γ gene-disrupted mice. J. Exp. Med. 1993, 178, 2243–2247. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.L.; Abel, L. Genetic dissection of immunity to mycobacteria: The human model. Annu. Rev. Immunol. 2002, 20, 581–620. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Tanaka, K.; Carroll, D.; Flynn, J.; Bloom, B.R. Effects of nitric oxide synthase inhibitors on murine infection with Mycobacterium tuberculosis. Infect. Immun. 1995, 63, 736–740. [Google Scholar] [PubMed]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D.; North, R.J.; LaCourse, R.; Mudgett, J.S.; Shah, S.K.; Nathan, C.F. Identification of nitric oxide synthase as a protective locus against tuberculosis. Proc. Natl. Acad. Sci. USA 1997, 94, 5243–5248. [Google Scholar] [CrossRef] [PubMed]
- Thoma-Uszynski, S.; Stenger, S.; Takeuchi, O.; Ochoa, M.T.; Engele, M.; Sieling, P.A.; Barnes, P.F.; Rollinghoff, M.; Bolcskei, P.L.; Wagner, M.; et al. Induction of direct antimicrobial activity through mammalian toll-like receptors. Science 2001, 291, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- Poli, V. The role of C/EBP isoforms in the control of inflammatory and native immunity functions. J. Biol. Chem. 1998, 273, 29279–29282. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, H.; Schwarz, P.M.; Forstermann, U. Regulation of the expression of inducible nitric oxide synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, W.; Pluss, C.; Hummel, R.; Pfeilschifter, J. Molecular mechanisms of inducible nitric oxide synthase gene expression by IL-1β and cAMP in rat mesangial cells. J. Immunol. 1998, 160, 4961–4969. [Google Scholar] [PubMed]
- Gupta, A.K.; Kone, B.C. CCAAT/enhancer binding protein-β trans-activates murine nitric oxide synthase 2 gene in an MTAL cell line. Am. J. Physiol. 1999, 276, F599–F605. [Google Scholar] [PubMed]
- Dlaska, M.; Weiss, G. Central role of transcription factor NF-IL6 for cytokine and iron-mediated regulation of murine inducible nitric oxide synthase expression. J. Immunol. 1999, 162, 6171–6177. [Google Scholar] [PubMed]
- Tan, C.K.; Tan, E.H.; Luo, B.; Huang, C.L.; Loo, J.S.; Choong, C.; Tan, N.S. SMAD3 deficiency promotes inflammatory aortic aneurysms in angiotensin II-infused mice via activation of iNOS. J. Am. Heart Assoc. 2013, 2, e269. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Diaz, R.A.; Higham, S.; Kone, B.C. alpha-MSH inhibits induction of C/EBPβ-DNA binding activity and NOS2 gene transcription in macrophages. Kidney Int. 2000, 57, 2239–2248. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Akira, S.; Yoshida, K.; Umemoto, M.; Yoneda, Y.; Shirafuji, N.; Fujiwara, H.; Suematsu, S.; Yoshida, N.; Kishimoto, T. Targeted disruption of the NF-IL6 gene discloses its essential role in bacteria killing and tumor cytotoxicity by macrophages. Cell 1995, 80, 353–361. [Google Scholar] [CrossRef]
- Screpanti, I.; Romani, L.; Musiani, P.; Modesti, A.; Fattori, E.; Lazzaro, D.; Sellitto, C.; Scarpa, S.; Bellavia, D.; Lattanzio, G.; et al. Lymphoproliferative disorder and imbalanced T-helper response in C/EBP β-deficient mice. Embo J. 1995, 14, 1932–1941. [Google Scholar] [PubMed]
- Nathan, C.F.; Hibbs, J.J. Role of nitric oxide synthesis in macrophage antimicrobial activity. Curr. Opin. Immunol. 1991, 3, 65–70. [Google Scholar] [CrossRef]
- Lowenstein, C.J.; Alley, E.W.; Raval, P.; Snowman, A.M.; Snyder, S.H.; Russell, S.W.; Murphy, W.J. Macrophage nitric oxide synthase gene: Two upstream regions mediate induction by interferon γ and lipopolysaccharide. Proc. Natl. Acad. Sci. USA 1993, 90, 9730–9734. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, K.; Zhou, L.; Wu, M.; Wu, Y.; Zhu, M.; Lai, X.; Chen, T.; Feng, L.; Li, M.; et al. MicroRNA-155 promotes autophagy to eliminate intracellular mycobacteria by targeting Rheb. PLoS Pathog. 2013, 9, e1003697. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; de Haro, S.A.; Master, S.S.; Keane, J.; Roberts, E.A.; Delgado, M.; Deretic, V. T helper 2 cytokines inhibit autophagic control of intracellular Mycobacterium tuberculosis. Immunity 2007, 27, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Jouanguy, E.; Doffinger, R.; Dupuis, S.; Pallier, A.; Altare, F.; Casanova, J.L. IL-12 and IFN-γ in host defense against mycobacteria and salmonella in mice and men. Curr. Opin. Immunol. 1999, 11, 346–351. [Google Scholar] [CrossRef]
- Sakowski, E.T.; Koster, S.; Celhay, C.P.; Park, H.S.; Shrestha, E.; Hetzenecker, S.E.; Maurer, K.; Cadwell, K.; Philips, J.A. Ubiquilin 1 Promotes IFN-γ-induced xenophagy of Mycobacterium tuberculosis. PLoS Pathog. 2015, 11, e1005076. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Hendriks, W.; Althage, A.; Hemmi, S.; Bluethmann, H.; Kamijo, R.; Vilcek, J.; Zinkernagel, R.M.; Aguet, M. Immune response in mice that lack the interferon-γ receptor. Science 1993, 259, 1742–1745. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Lerena, M.C.; Colombo, M.I. Mycobacterium marinum induces a marked LC3 recruitment to its containing phagosome that depends on a functional ESX-1 secretion system. Cell. Microbiol. 2011, 13, 814–835. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Wang, D.; Li, M.; Wang, H.; Yu, J.; Wang, C.; Liu, J.; Gao, Q. PPE38 modulates the innate immune response and is required for Mycobacterium marinum virulence. Infect. Immun. 2012, 80, 43–54. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Use | Sequences (5′–3′) |
|---|---|
| C/EBPβ 3′UTR | Forward: CCGCTCGAGGCCCTGAGTAATCACTTAAAGATGTTC |
| Revese: ATAAGAATGCGGCCGCAATGTCTTCACTTTAATGCTCGAAA | |
| C/EBPβ 3′UTR-mut Mutant | Forward: AAGTGAAGACATTGCGGCCGCTGG |
| Reverse: AATTCGTCGAAACGGAAAAGGTTCTCAAAATATACA | |
| C/EBPβ shRNA | Forward: gatccCACCCTGCGGAACTTGTTCAATTCAAGAGATTGAACAAGTTCCGCAGGGTGtttttg |
| Reverse: aattcaaaaaCACCCTGCGGAACTTGTTCAATCTCTTGAATTGAACAAGTTCCGCAGGGTGg | |
| Scramble shRNA | Forward: GATCCTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTG |
| Reverse: aattcaaaaaTTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAAg | |
| mmu-miR-155 | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACCCCT |
| Forward: CGCCTGTTAATGCTAATTGTGA | |
| Revese: AGTGCAGGGTCCGAGGTAT | |
| hsa-miR-155 | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACCCCT |
| Forward: CGGTTAATGCTAATCGTGATAGG | |
| Reverse: CAGTGCAGGGTCCGAGGTAT | |
| mmu-miR-146a | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAACCCA |
| Forward: GCCTGAGAACTGAATTCCATG | |
| Revese: AGTGCAGGGTCCGAGGTAT | |
| mmu-miR-125b | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCACAA |
| Forward: TCCCTGAGACCCTAACTTGTGAG | |
| Revese: CAGTGCAGGGTCCGAGGTAT | |
| mmu-miR-129 | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGCAAGC |
| Forward: CTTTTTGCGGTCTGGGCTT | |
| Revese: CAGTGCAGGGTCCGAGGTAT | |
| Let-7 | RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAACTAT |
| Forward: TGAGGTAGTAGGTTGTATAGTTGTCGTAT | |
| Revese: CAGTGCAGGGTCCGAGGTAT | |
| RNU6-1 | RT: AACGCTTCACGAATTTGCGT |
| Forward: CTCGCTTCGGCAGCACA | |
| Revese: AACGCTTCACGAATTTGCGT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Wang, Q.; Zhou, Y.; Duan, Y.; Gao, Q. Inhibition of IFN-γ-Induced Nitric Oxide Dependent Antimycobacterial Activity by miR-155 and C/EBPβ. Int. J. Mol. Sci. 2016, 17, 535. https://doi.org/10.3390/ijms17040535
Qin Y, Wang Q, Zhou Y, Duan Y, Gao Q. Inhibition of IFN-γ-Induced Nitric Oxide Dependent Antimycobacterial Activity by miR-155 and C/EBPβ. International Journal of Molecular Sciences. 2016; 17(4):535. https://doi.org/10.3390/ijms17040535
Chicago/Turabian StyleQin, Yongwei, Qinglan Wang, Youlang Zhou, Yinong Duan, and Qian Gao. 2016. "Inhibition of IFN-γ-Induced Nitric Oxide Dependent Antimycobacterial Activity by miR-155 and C/EBPβ" International Journal of Molecular Sciences 17, no. 4: 535. https://doi.org/10.3390/ijms17040535