Dose- and Ion-Dependent Effects in the Oxidative Stress Response to Space-Like Radiation Exposure in the Skeletal System

Abstract
:1. Introduction
2. Results
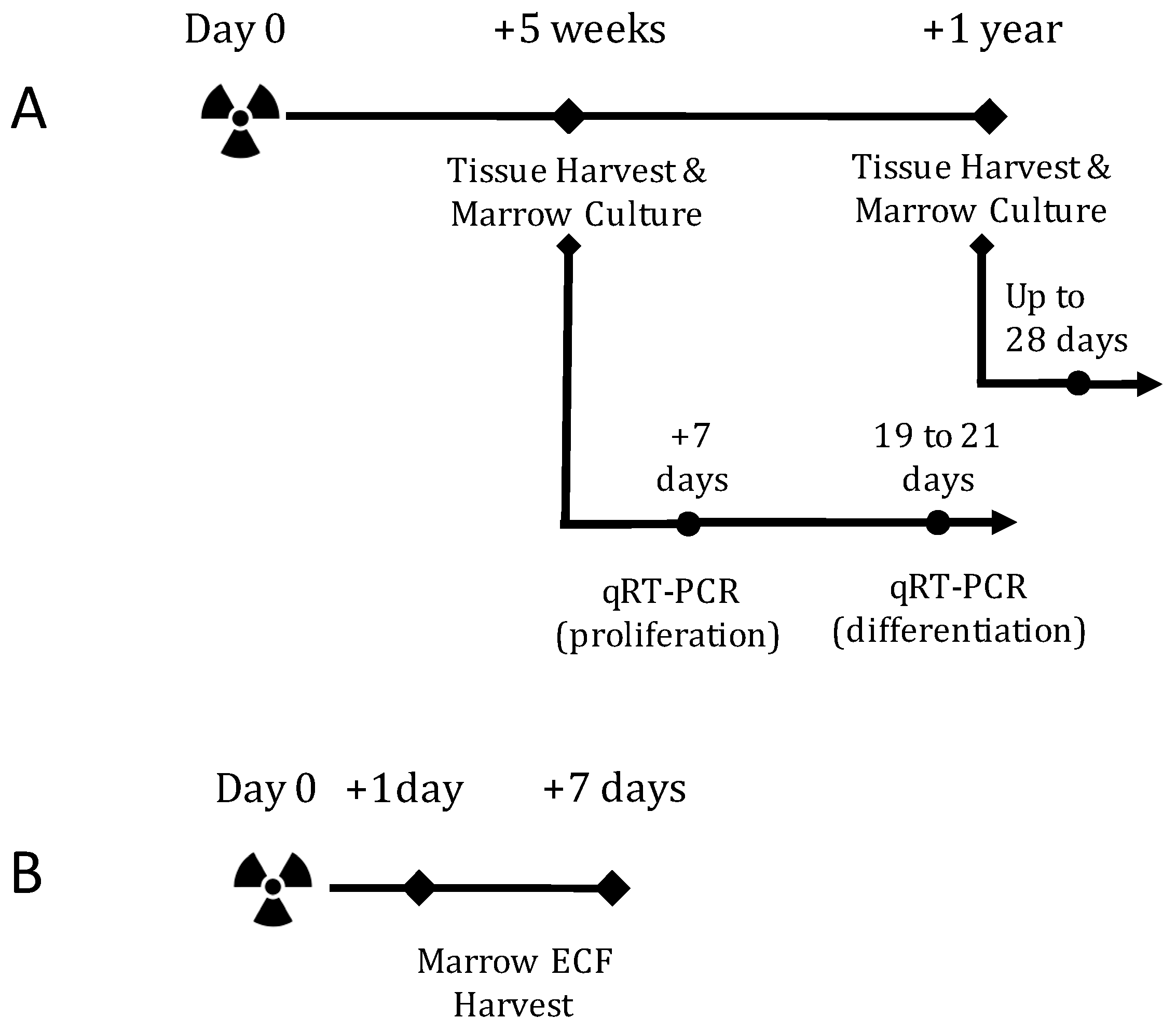
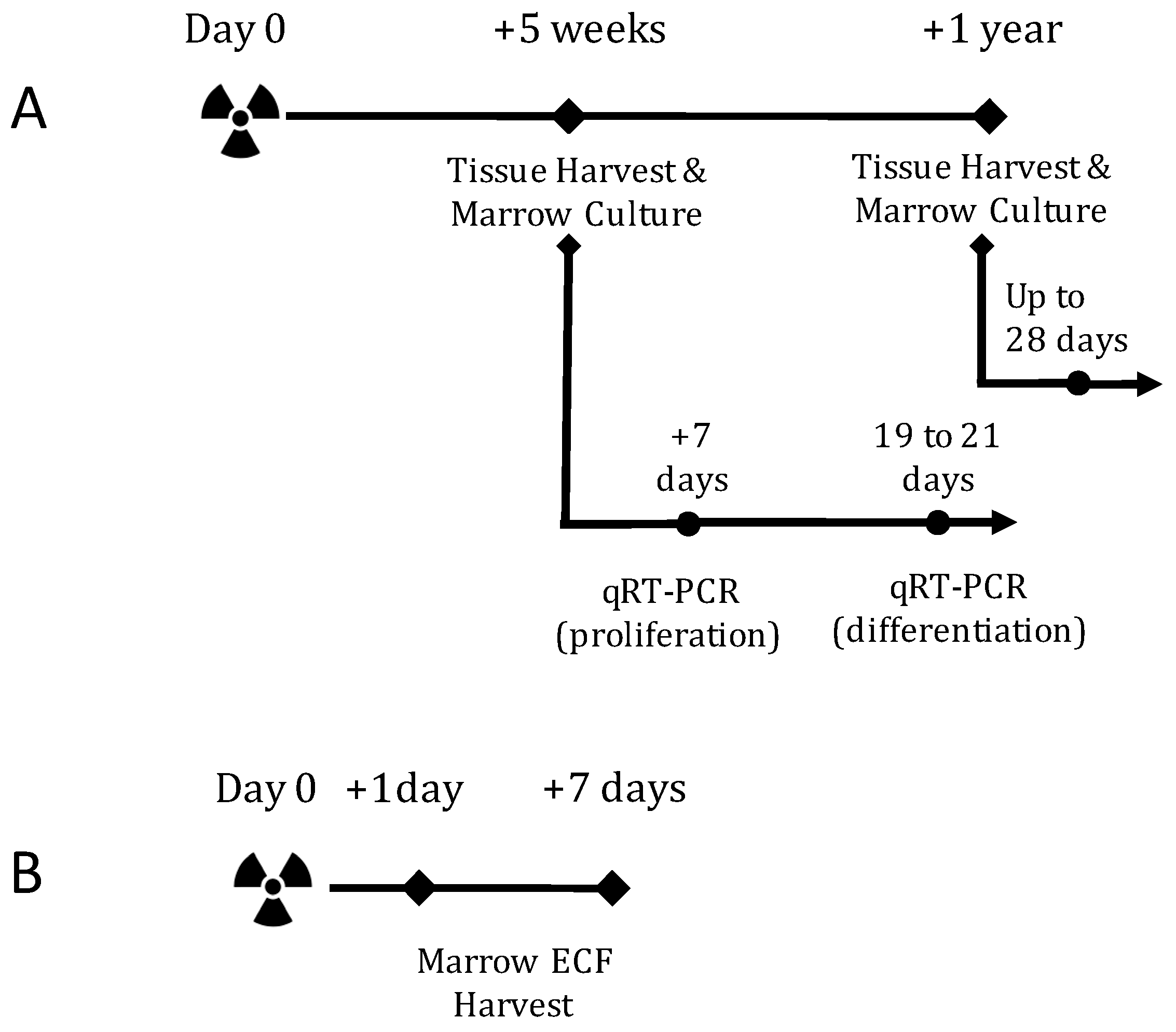
2.1. TBI Experiment Design
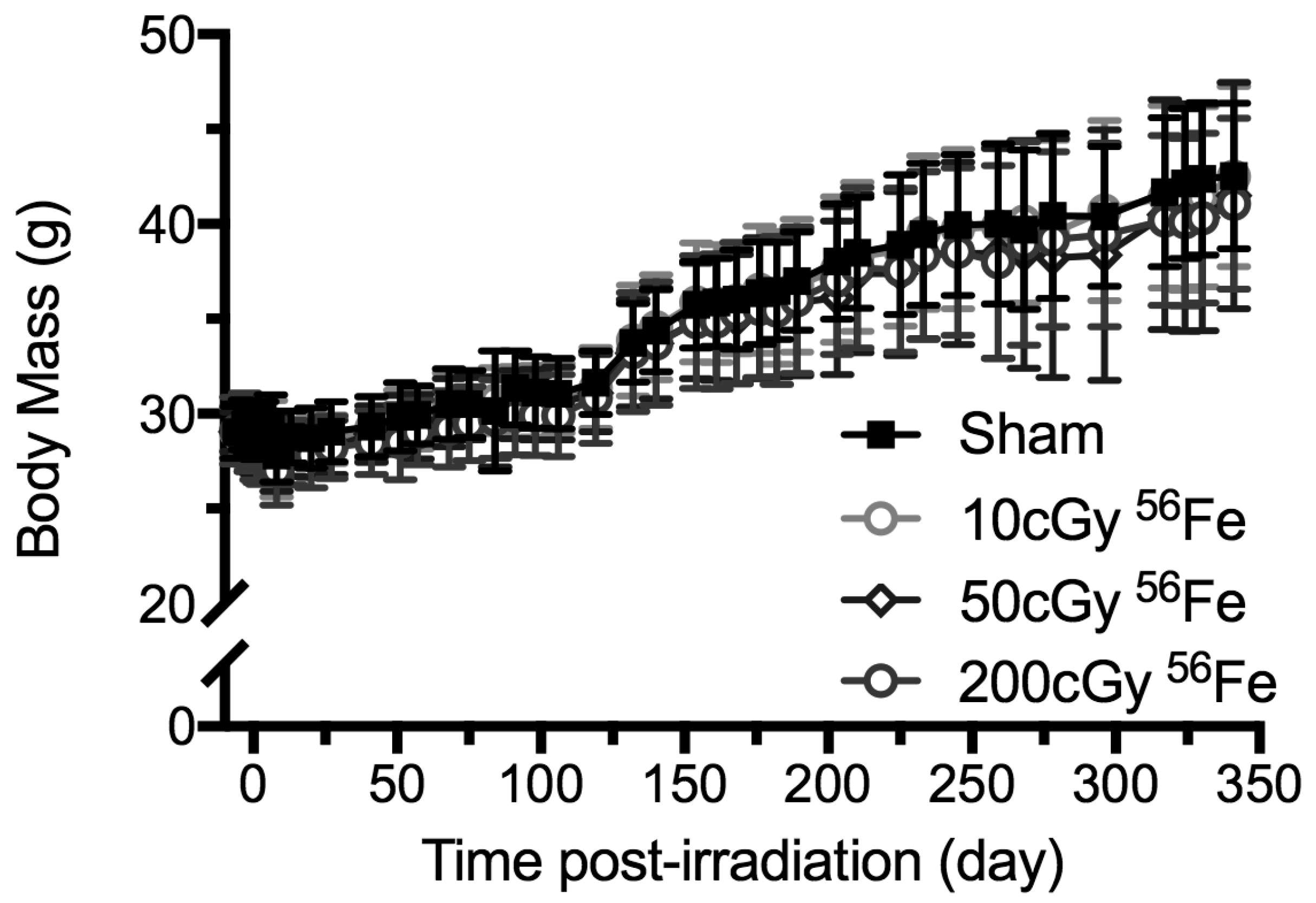
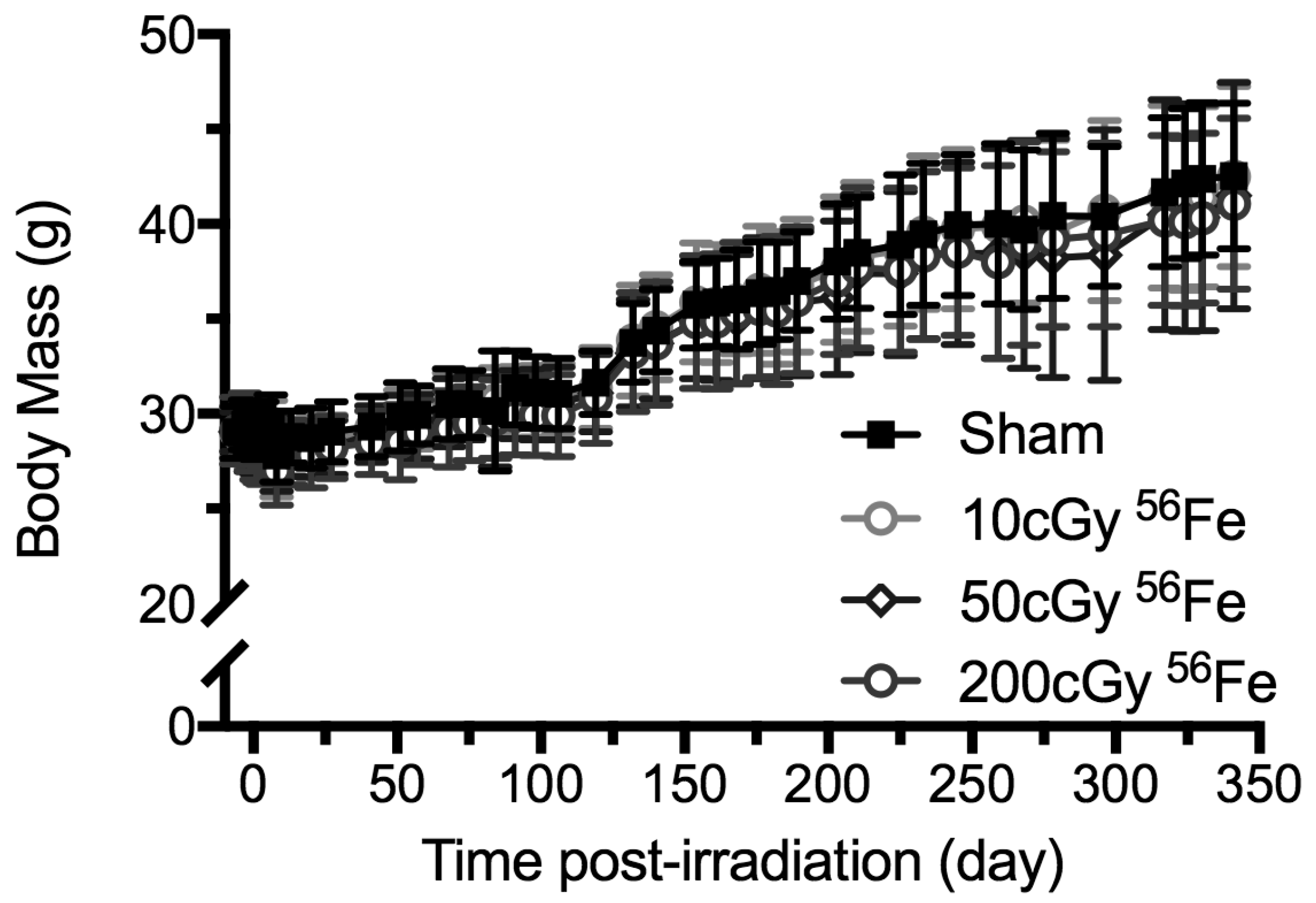
2.2. Body Mass Temporal Responses and Coat Color
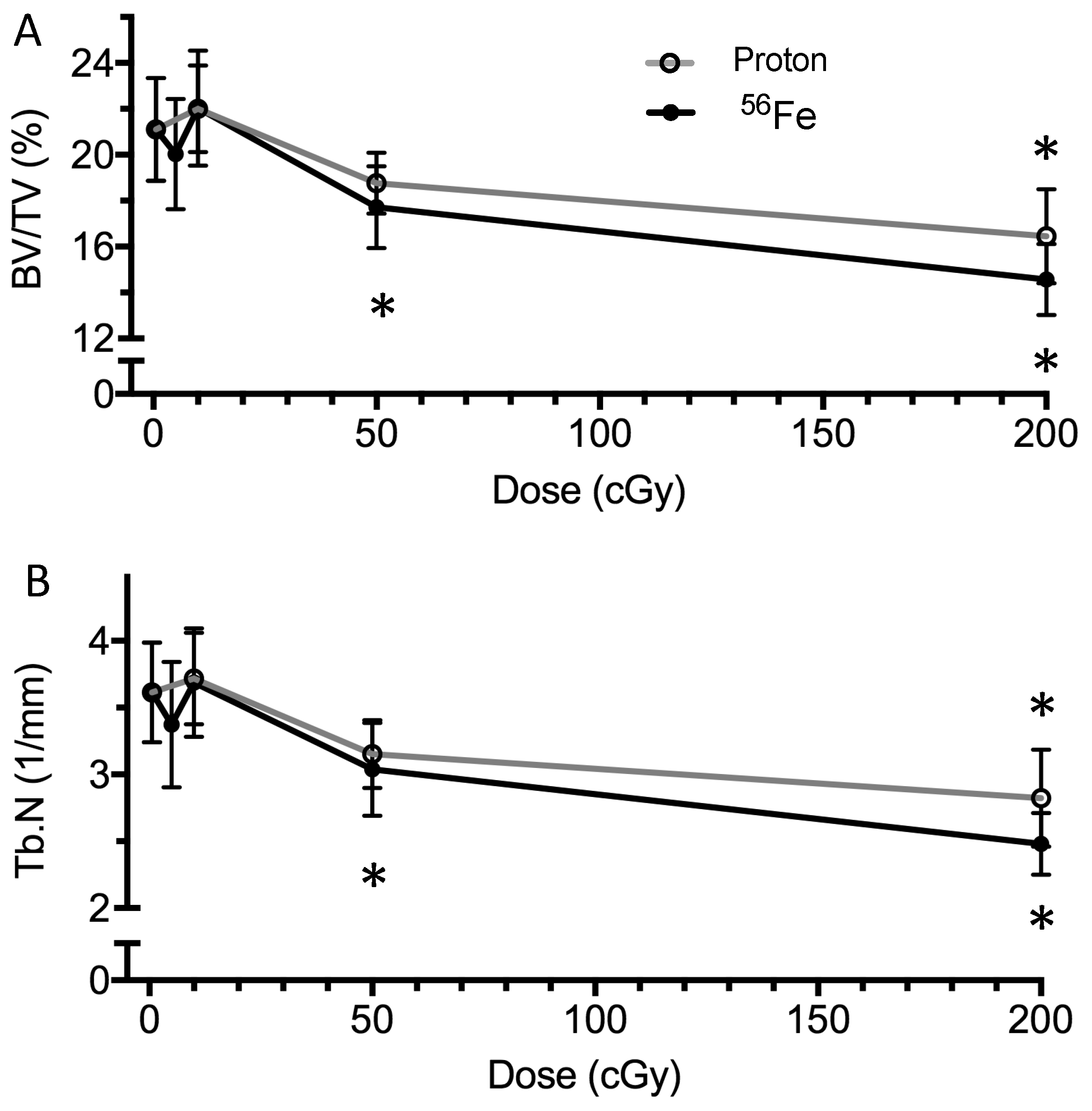
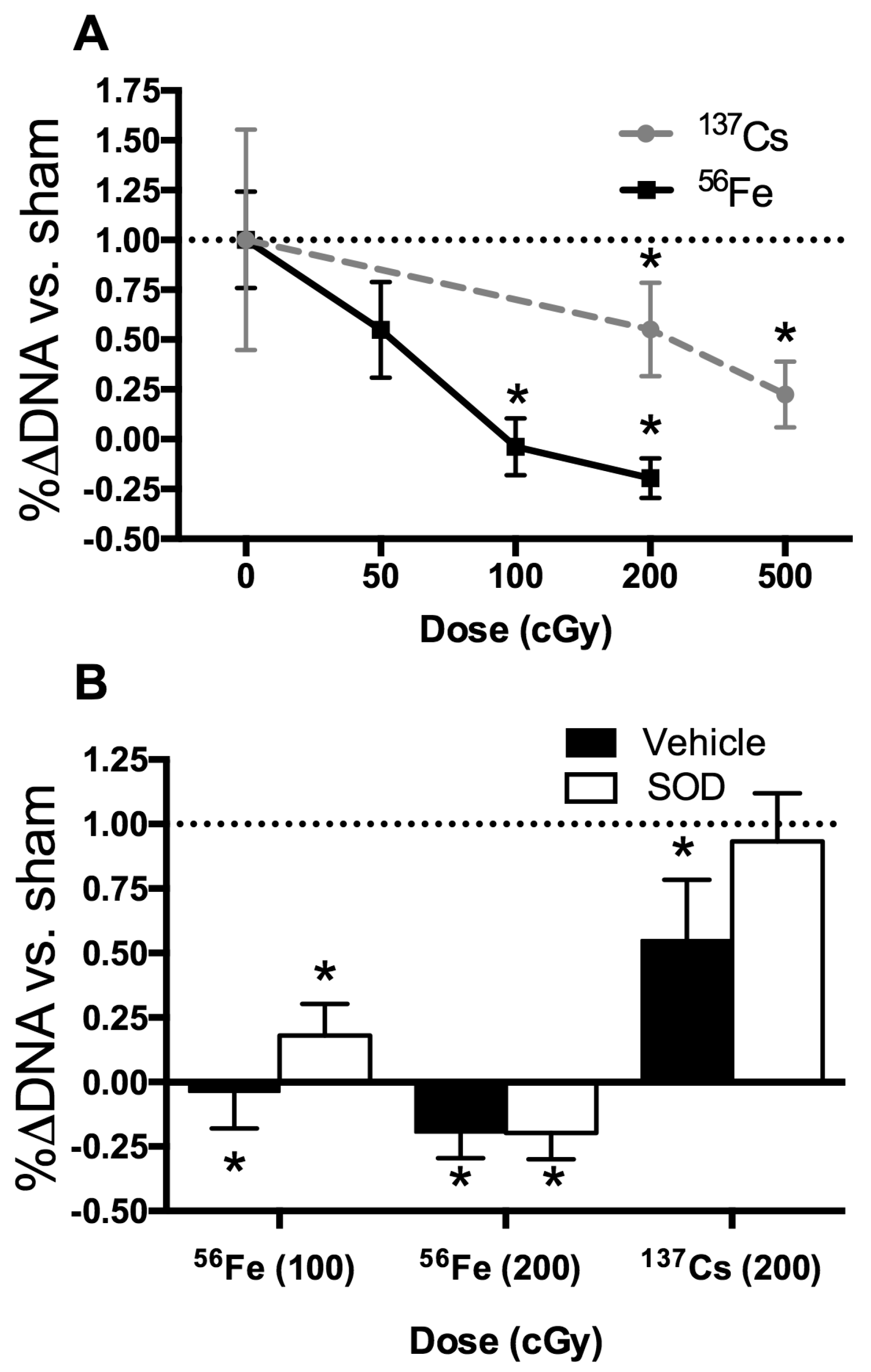
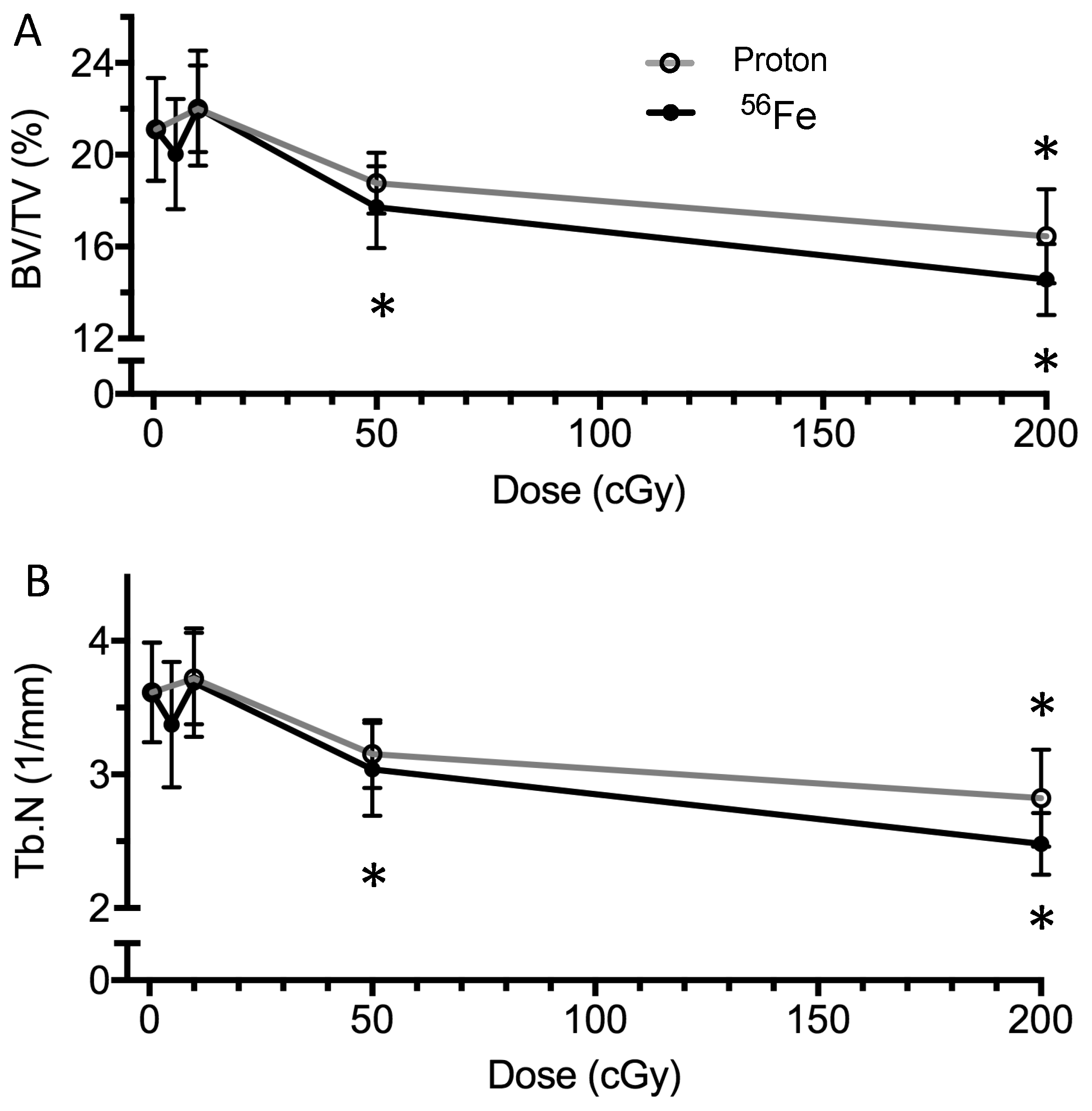
2.3. Bone Structure
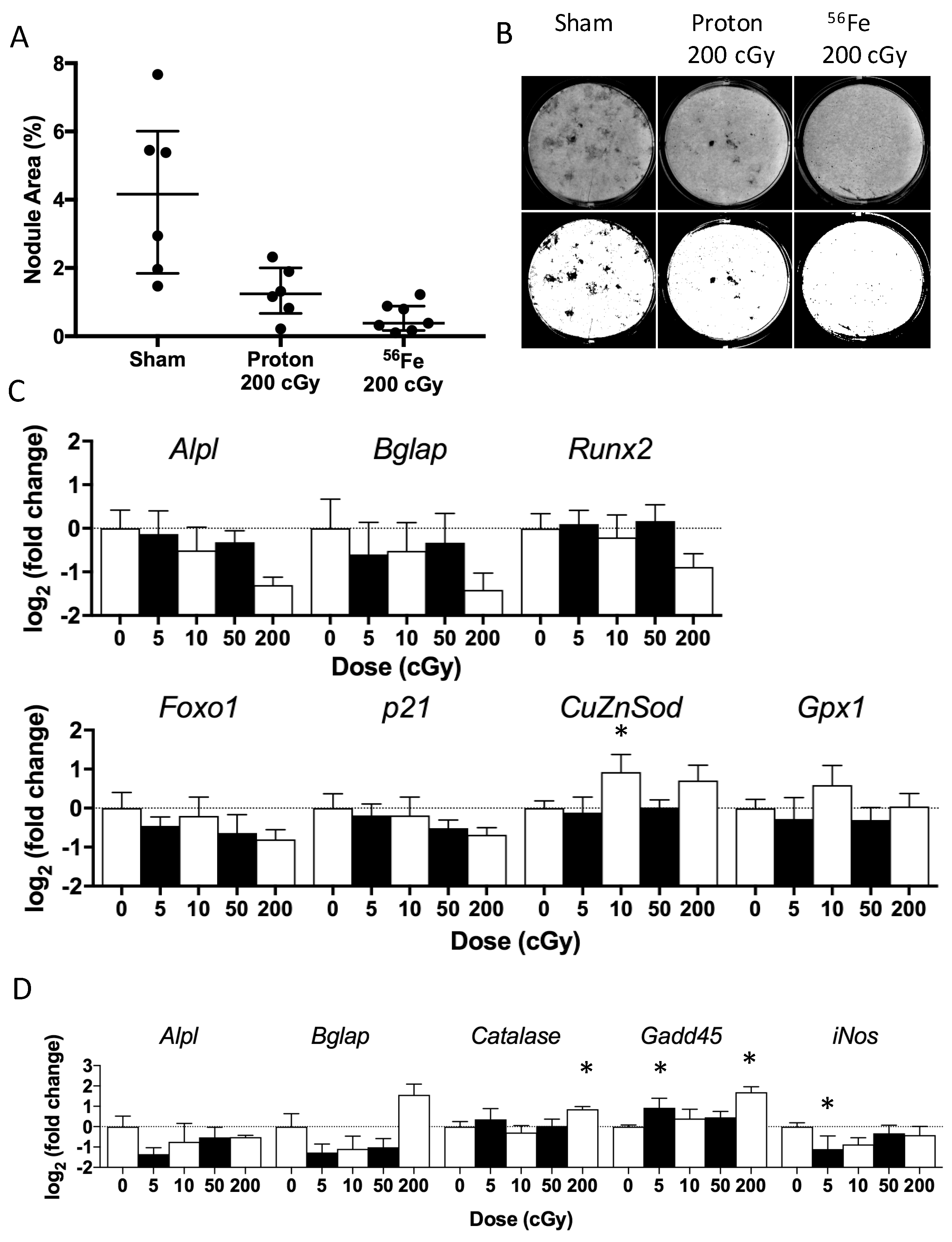
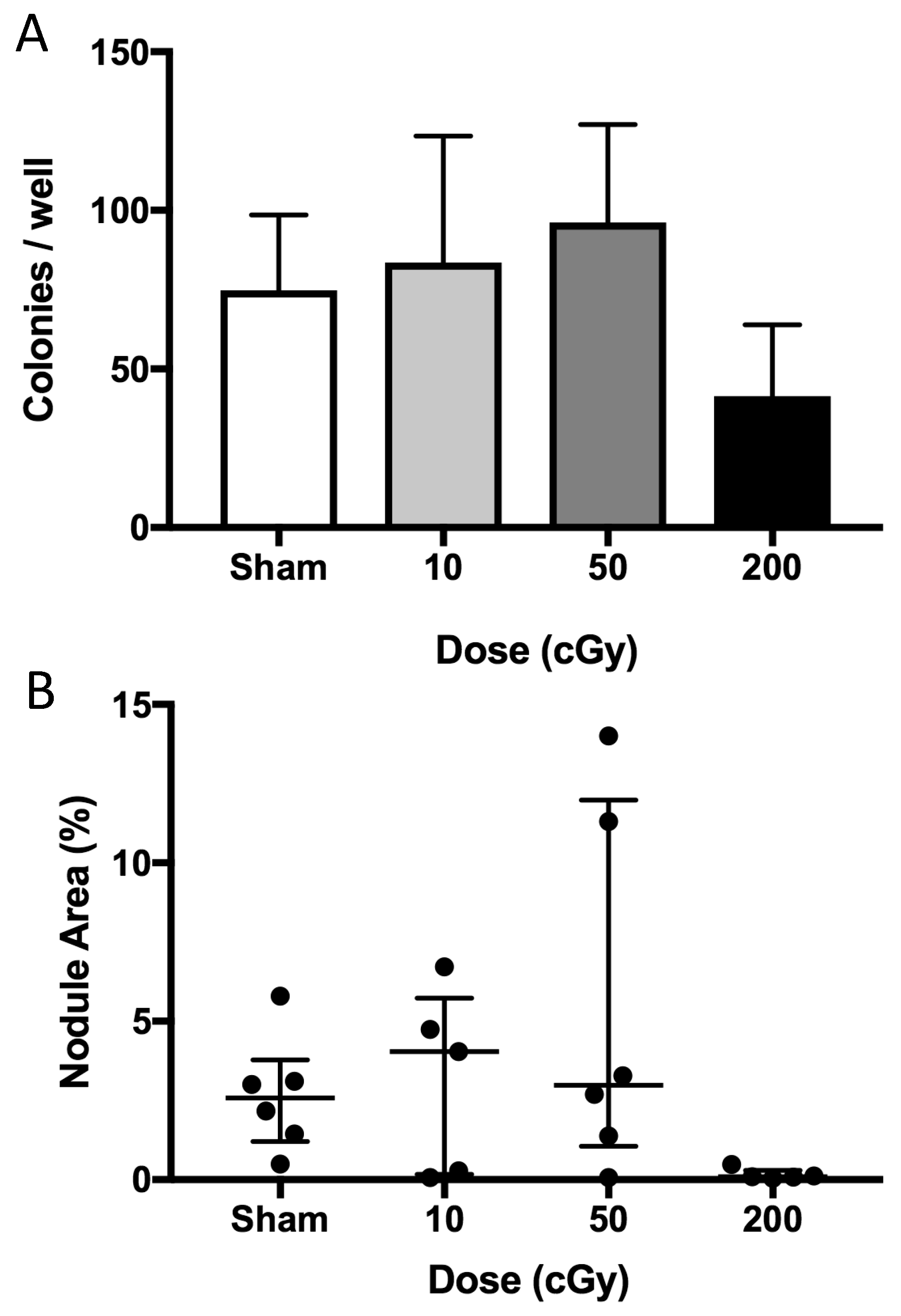
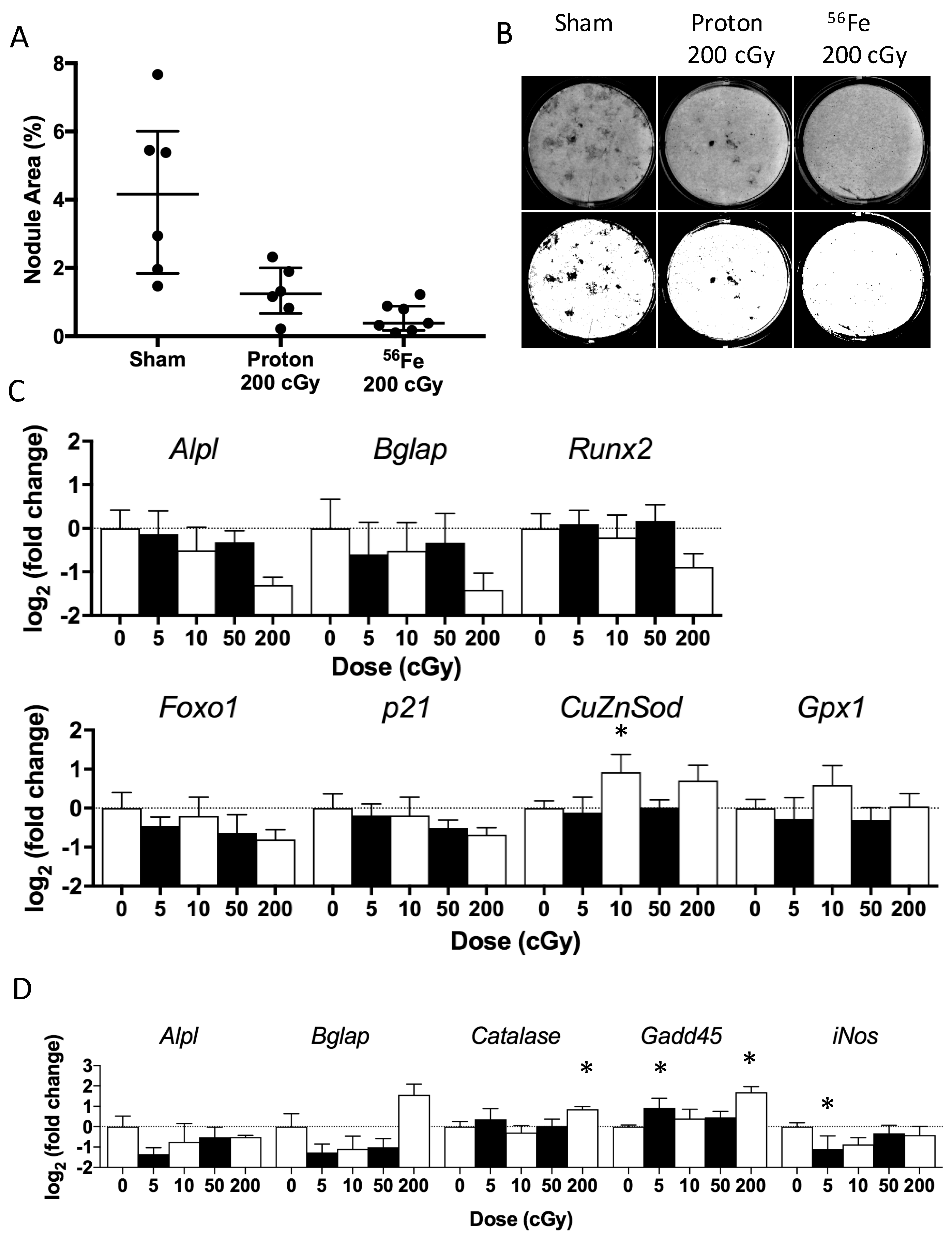
2.4. Ex Vivo Osteoblastogenesis
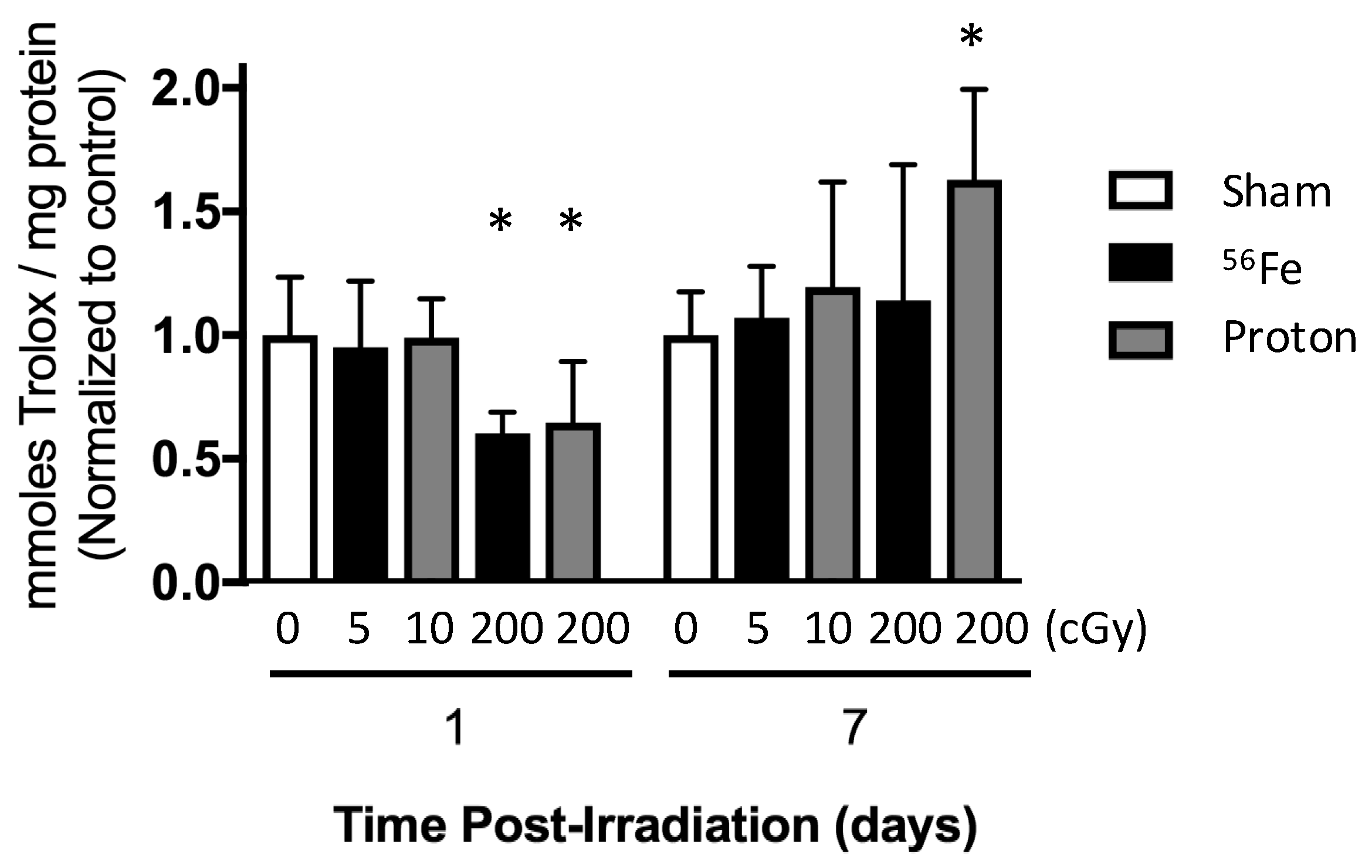
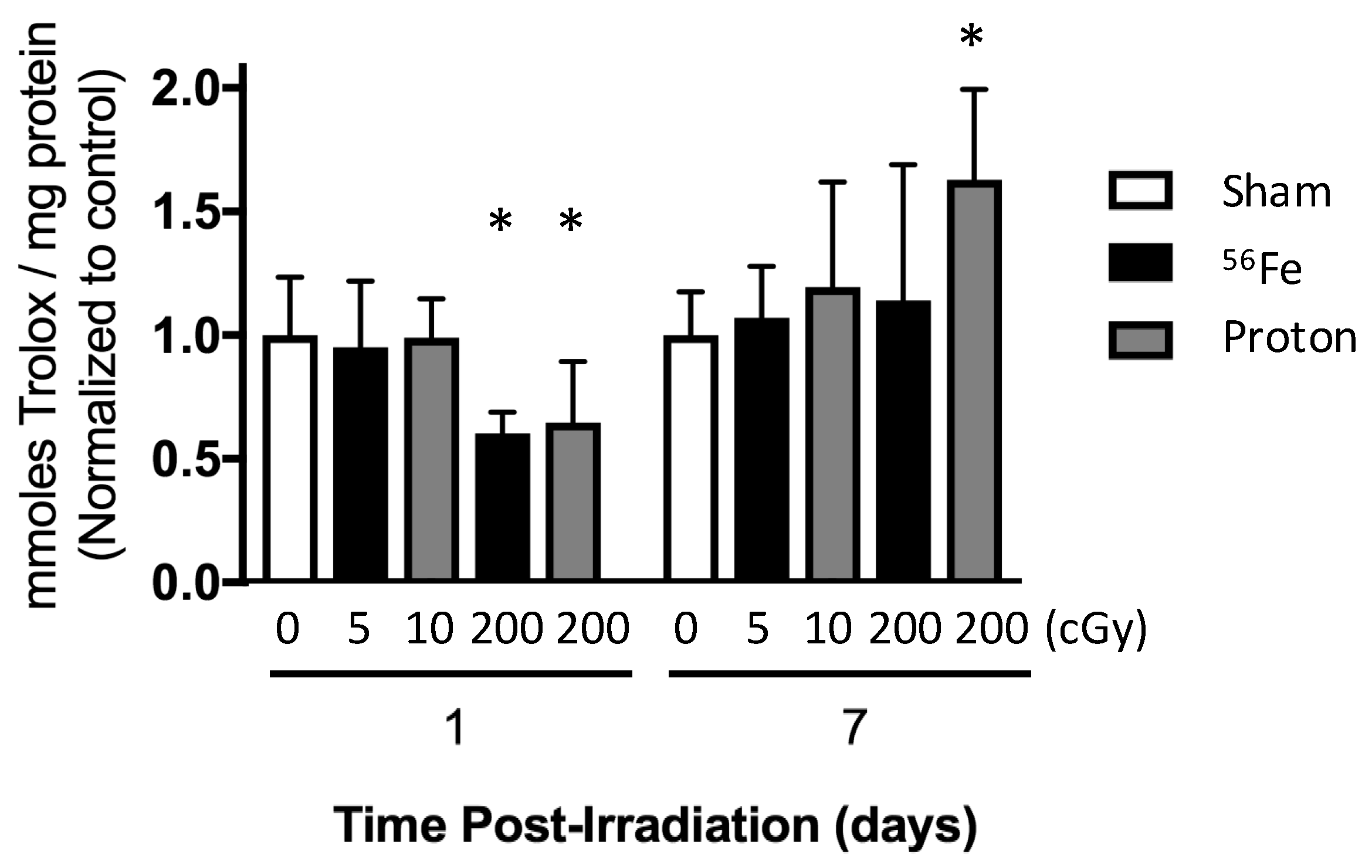
2.5. Total Antioxidant Capacity of the Marrow Extracellular Fluid
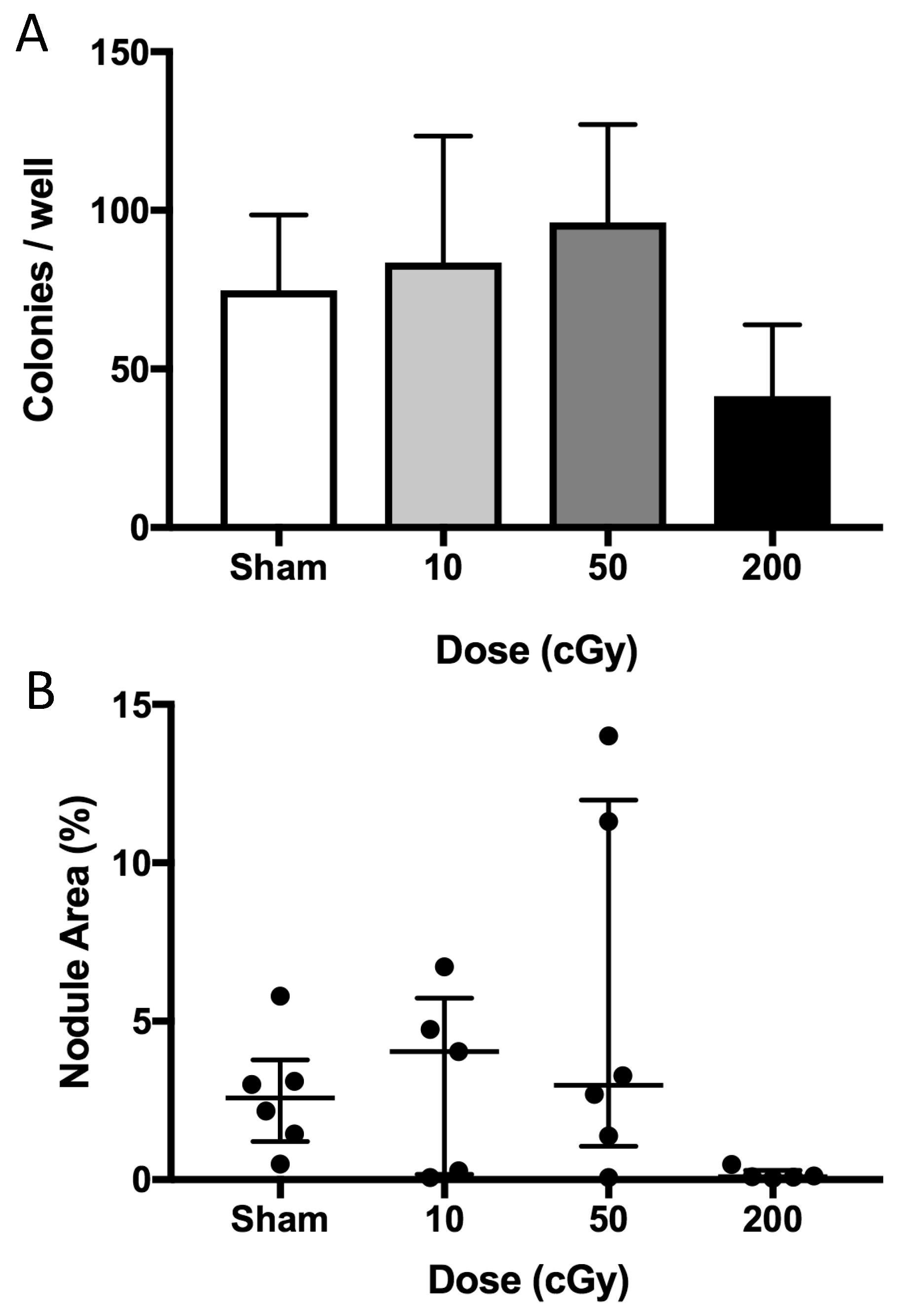
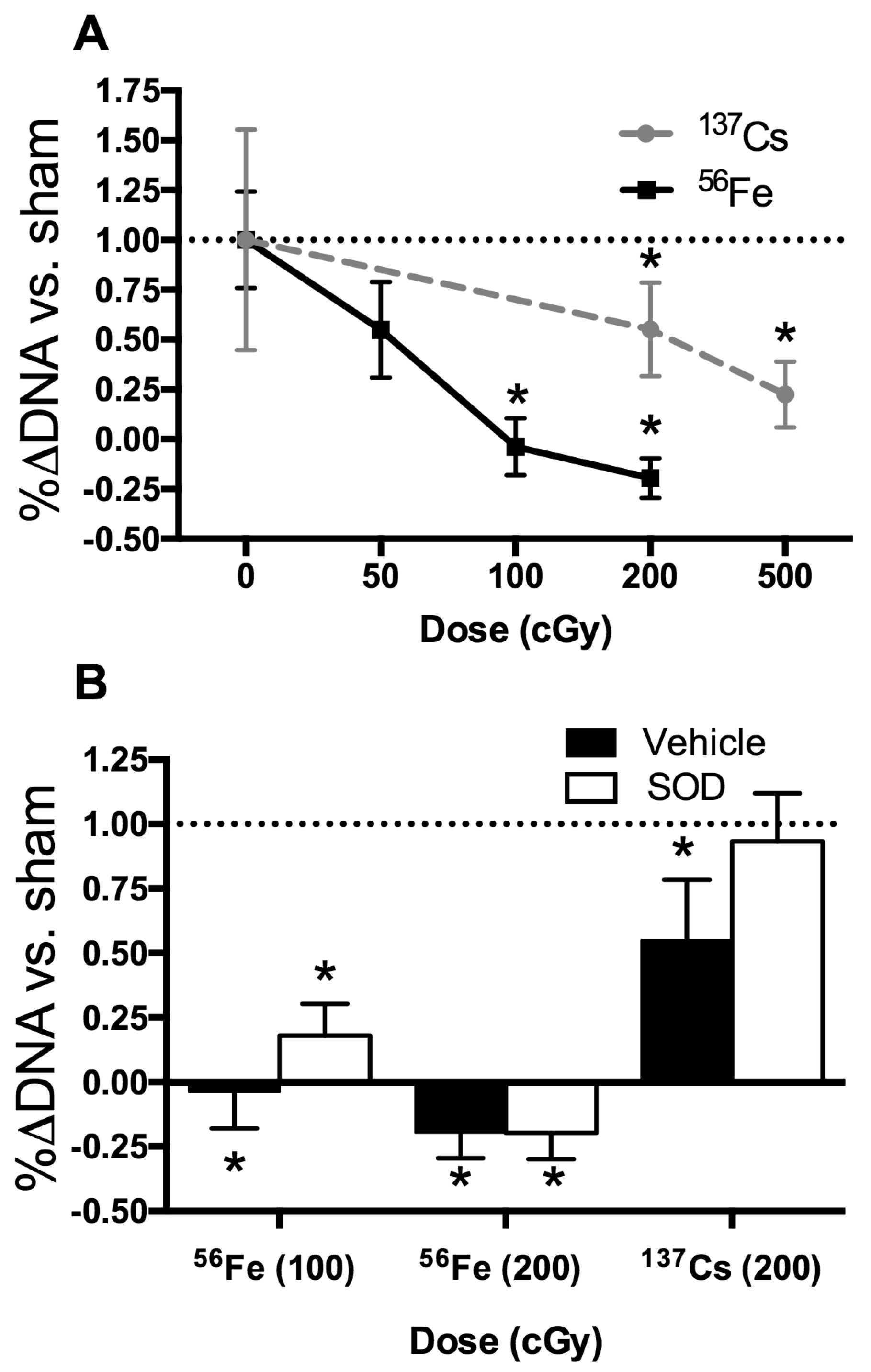
2.6. In Vitro Radiation Effects on Osteoblastogenesis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. In Vivo Experiment Design and Radiation Exposure
4.3. In Vitro Experiment Design and Radiation Exposure
4.4. Total Antioxidant Capacity
4.5. Bone Microarchitecture and Geometry
4.6. Ex Vivo Osteoblastogenic Assays and qRT-PCR Analyses
4.7. Statistics
Acknowledgments
Author Contributions
Conflict of Interest
References
- McPhee, J.C.; Charles, J.B. Human Health and Peformance Risks of Space Exploration Missions; Government Printing Office: Washington, DC, USA, 2009.
- Guo, J.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Hassler, D.M.; Ehresmann, B.; Kohler, J.; Bohm, E.; Bottcher, S.; Brinza, D.; Burmeister, S.; et al. Msl-rad radiation environment measurements. Radiat. Prot. Dosimetry 2015, 166, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Dietze, G.; Bartlett, D.T.; Cool, D.A.; Cucinotta, F.A.; Jia, X.; McAulay, I.R.; Pelliccioni, M.; Petrov, V.; Reitz, G.; Sato, T. Icrp publication 123: Assessment of radiation exposure of astronauts in space. Ann. ICRP 2013. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Cucinotta, F.A. Physical basis of radiation protection in space travel. Rev. Mod. Phys. 2011, 83, 1245–1281. [Google Scholar] [CrossRef]
- Tseng, B.P.; Giedzinski, E.; Izadi, A.; Suarez, T.; Lan, M.L.; Tran, K.K.; Acharya, M.M.; Nelson, G.A.; Raber, J.; Parihar, V.K.; et al. Functional consequences of radiation-induced oxidative stress in cultured neural stem cells and the brain exposed to charged particle irradiation. Antioxid. Redox Signal. 2014, 20, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Datta, K.; Suman, S.; Fornace, A.J., Jr. Radiation persistently promoted oxidative stress, activated mtor via pi3k/akt, and downregulated autophagy pathway in mouse intestine. Int. J. Biochem. Cell Biol. 2014, 57, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Norbury, J.W.; Slaba, T.C. Space radiation accelerator experiments – the role of neutrons and light ions. Life Sci. Space Res. 2014, 3, 90–94. [Google Scholar] [CrossRef]
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Bottcher, S.; Bohm, E.; et al. Mars’ surface radiation environment measured with the mars science laboratory’s curiosity rover. Science 2014, 343, 1244797. [Google Scholar] [CrossRef] [PubMed]
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Bottcher, S.; Bohm, E.; et al. Measurements of energetic particle radiation in transit to mars on the mars science laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- NASA. Nasa std-3001, Space Flight Human-System Standard Volume 1, Revision a, Change 1: Crew Health; NASA: Washington, DC, USA, 2015. [Google Scholar]
- Townsend, L.W.; Badhwar, G.D.; Braby, L.A.; Blakely, E.A.; Cucinotta, F.A.; Curtis, S.B.; Fry, R.J.M.; Land, C.E.; Smart, D.F. Report No. 153-Information Needed to Make Radiation Protection Recommendations for Space Missions Beyond Low-Earth Orbit; US National Council for Radiation Protection and Measurements: Bethesda, MD, USA, 2006. [Google Scholar]
- Kondo, H.; Yumoto, K.; Alwood, J.S.; Mojarrab, R.; Wang, A.; Almeida, E.A.; Searby, N.D.; Limoli, C.L.; Globus, R.K. Oxidative stress and gamma radiation-induced cancellous bone loss with musculoskeletal disuse. J. Appl. Physiol. 2010, 108, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Alwood, J.S.; Shahnazari, M.; Chicana, B.; Schreurs, A.S.; Kumar, A.; Bartolini, A.; Shirazi-Fard, Y.; Globus, R.K. Ionizing radiation stimulates expression of pro-osteoclastogenic genes in marrow and skeletal tissue. J. Interferon Cytokine Res. 2015, 35, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Ambrogini, E.; Han, L.; Manolagas, S.C.; Jilka, R.L. Increased lipid oxidation causes oxidative stress, increased peroxisome proliferator-activated receptor-gamma expression, and diminished pro-osteogenic wnt signaling in the skeleton. J. Biol. Chem. 2009, 284, 27438–27448. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C.; Almeida, M. Gone with the wnts: Beta-catenin, t-cell factor, forkhead box o, and oxidative stress in age-dependent diseases of bone, lipid, and glucose metabolism. Mol. Endocrinol. 2007, 21, 2605–2614. [Google Scholar] [CrossRef] [PubMed]
- Lean, J.M.; Jagger, C.J.; Kirstein, B.; Fuller, K.; Chambers, T.J. Hydrogen peroxide is essential for estrogen-deficiency bone loss and osteoclast formation. Endocrinology 2005, 146, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, K.; Globus, R.K.; Mojarrab, R.; Arakaki, J.; Wang, A.; Searby, N.D.; Almeida, E.A.; Limoli, C.L. Short-term effects of whole-body exposure to (56)fe ions in combination with musculoskeletal disuse on bone cells. Radiat. Res. 2010, 173, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Kurpinski, K.; Jang, D.J.; Bhattacharya, S.; Rydberg, B.; Chu, J.; So, J.; Wyrobek, A.; Li, S.; Wang, D. Differential effects of x-rays and high-energy 56fe ions on human mesenchymal stem cells. Int.J.Radiat.Oncol.Biol.Phys. 2009, 73, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.H.; Kim, K.A.; Ji, H.; Lee, D.; Lee, J.C. Irradiation inhibits the maturation and mineralization of osteoblasts via the activation of nrf2/ho-1 pathway. Mol. Cell. Biochem. 2015, 410, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.S.; Livingston, E.W.; Robbins, M.E.; Bourland, J.D.; Tirado-Lee, L.; Smith-Sielicki, H.; Bateman, T.A. Risedronate prevents early radiation-induced osteoporosis in mice at multiple skeletal locations. Bone 2010, 46, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.; Swift, J.M.; Greene, E.S.; Allen, M.R.; Cunningham, D.A.; Braby, L.A.; Bloomfield, S.A. Exposure to low-dose x-ray radiation alters bone progenitor cells and bone microarchitecture. Radiat. Res. 2017, 188, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.A.; Pecaut, M.J.; Gridley, D.S.; Travis, N.D.; Bandstra, E.R.; Willey, J.S.; Nelson, G.A.; Bateman, T.A. A murine model for bone loss from therapeutic and space-relevant sources of radiation. J. Appl. Physiol. 2006, 101, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Alwood, J.S.; Tran, L.H.; Schreurs, A.-S.; Shirazi-Fard, Y.; Kumar, A.; Hilton, D.; Tahimic, C.G.T.; Globus, R.K. Data not Shown; NASA Ames Research Center: Moffett Field, CA, USA, 2017. [Google Scholar]
- Macias, B.R.; Lima, F.; Swift, J.M.; Shirazi-Fard, Y.; Greene, E.S.; Allen, M.R.; Fluckey, J.; Hogan, H.A.; Braby, L.; Wang, S.; et al. Simulating the lunar environment: Partial weightbearing and high-let radiation-induce bone loss and increase sclerostin-positive osteocytes. Radiat. Res. 2016, 186, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Decker, H.; Hartmann, H.; Chavan, B.; Rokos, H.; Spencer, J.D.; Hasse, S.; Thornton, M.J.; Shalbaf, M.; Paus, R.; et al. Senile hair graying: H2o2-mediated oxidative stress affects human hair color by blunting methionine sulfoxide repair. FASEB J. 2009, 23, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Free radical theory of aging: An update: Increasing the functional life span. Ann. N. Y. Acad. Sci. 2006, 1067, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Searby, N.D.; Mojarrab, R.; Phillips, J.; Alwood, J.; Yumoto, K.; Almeida, E.A.C.; Limoli, C.L.; Globus, R.K. Total-body irradiation of postpubertal mice with 137cs acutely compromises the microarchitecture of cancellous bone and increases osteoclasts. Radiat. Res. 2009, 171, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Greenberger, J.S.; Epperly, M. Bone marrow-derived stem cells and radiation response. Semin. Radiat. Oncol. 2009, 19, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, A.S.; Shirazi-Fard, Y.; Shahnazari, M.; Alwood, J.S.; Truong, T.A.; Tahimic, C.G.; Limoli, C.L.; Turner, N.D.; Halloran, B.; Globus, R.K. Dried plum diet protects from bone loss caused by ionizing radiation. Sci. Rep. 2016, 6, 21343. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hellweg, C.E.; Baumstark-Khan, C.; Reitz, G.; Lau, P. Cell cycle delay in murine pre-osteoblasts is more pronounced after exposure to high-let compared to low-let radiation. Radiat. Environ. Biophys. 2014, 53, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Snyder, A.R.; Morgan, W.F. Gene expression profiling after irradiation: Clues to understanding acute and persistent responses? Cancer Metastasis Rev. 2004, 23, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.; Brunet, A.; Grenier, J.M.; Datta, S.R.; Fornace, A.J.; DiStefano, P.S.; Chiang, L.W.; Greenberg, M.E. DNA repair pathway stimulated by the forkhead transcription factor foxo3a through the gadd45 protein. Science 2002, 296, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Furukawa-Hibi, Y.; Yoshida-Araki, K.; Ohta, T.; Ikeda, K.; Motoyama, N. Foxo forkhead transcription factors induce g(2)-m checkpoint in response to oxidative stress. J. Biol. Chem. 2002, 277, 26729–26732. [Google Scholar] [CrossRef] [PubMed]
- Bartell, S.M.; Kim, H.N.; Ambrogini, E.; Han, L.; Iyer, S.; Serra Ucer, S.; Rabinovitch, P.; Jilka, R.L.; Weinstein, R.S.; Zhao, H.; et al. Foxo proteins restrain osteoclastogenesis and bone resorption by attenuating h2o2 accumulation. Nat. Commun. 2014, 5, 3773. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O.; Sanchez-Ramos, C.; Prieto-Arroyo, I.; Urbanek, P.; Steinbrenner, H.; Monsalve, M. Redox regulation of foxo transcription factors. Redox. Biol. 2015, 6, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Hoogeboom, D.; Essers, M.A.; Polderman, P.E.; Voets, E.; Smits, L.M.; Burgering, B.M. Interaction of foxo with beta-catenin inhibits beta-catenin/t cell factor activity. J. Biol. Chem. 2008, 283, 9224–9230. [Google Scholar] [CrossRef] [PubMed]
- Essers, M.A.; de Vries-Smits, L.M.; Barker, N.; Polderman, P.E.; Burgering, B.M.; Korswagen, H.C. Functional interaction between beta-catenin and foxo in oxidative stress signaling. Science 2005, 308, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C.; Hou, J. Rogue proliferation versus restorative protection: Where do we draw the line for wnt and forkhead signaling? Expert Opin. Ther. Targets 2008, 12, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.T.; Iwaniec, U.T.; Wong, C.P.; Lindenmaier, L.B.; Wagner, L.A.; Branscum, A.J.; Menn, S.A.; Taylor, J.; Zhang, Y.; Wu, H.; et al. Acute exposure to high dose gamma-radiation results in transient activation of bone lining cells. Bone 2013, 57, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Green, D.E.; Adler, B.J.; Chan, M.E.; Rubin, C.T. Devastation of adult stem cell pools by irradiation precedes collapse of trabecular bone quality and quantity. J. Bone Miner. Res. 2012, 27, 749. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wu, X.; Frassica, D.; Yu, B.; Pang, L.; Xian, L.; Wan, M.; Lei, W.; Armour, M.; Tryggestad, E.; et al. Irradiation induces bone injury by damaging bone marrow microenvironment for stem cells. Proc. Natl. Acad. Sci. USA 2011, 108, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Alwood, J.S.; Yumoto, K.; Mojarrab, R.; Limoli, C.L.; Almeida, E.A.; Searby, N.D.; Globus, R.K. Heavy ion irradiation and unloading effects on mouse lumbar vertebral microarchitecture, mechanical properties and tissue stresses. Bone 2010, 47, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.J.; Wronski, T.J.; Iwaniec, U.; Phleger, L.; Kurimoto, P.; Boudignon, B.; Halloran, B.P. Aging increases stromal/osteoblastic cell-induced osteoclastogenesis and alters the osteoclast precursor pool in the mouse. J. Bone Miner. Res. 2005, 20, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Alwood, J.S.; Kumar, A.; Tran, L.H.; Wang, A.; Limoli, C.L.; Globus, R.K. Low-dose, ionizing radiation and age-related changes in skeletal microarchitecture. J. Aging Res. 2012, 2012, 481983. [Google Scholar] [CrossRef] [PubMed]
- Oest, M.E.; Franken, V.; Kuchera, T.; Strauss, J.; Damron, T.A. Long-term loss of osteoclasts and unopposed cortical mineral apposition following limited field irradiation. J. Orthop. Res. 2015, 33, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, S.A.; Bandstra, E.R.; Travis, N.D.; Nelson, G.A.; Bourland, J.D.; Pecaut, M.J.; Gridley, D.S.; Willey, J.S.; Bateman, T.A. Spaceflight-relevant types of ionizing radiation and cortical bone: Potential let effect? Adv. Space Res. 2008, 42, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Hada, M.; Zhang, Y.; Feiveson, A.; Cucinotta, F.A.; Wu, H. Association of inter- and intrachromosomal exchanges with the distribution of low- and high-let radiation-induced breaks in chromosomes. Radiat. Res. 2011, 176, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, L.G.; Bertolotti, A.; Ranza, E.; Babini, G.; Ottolenghi, A. Investigation of the mechanisms underpinning il-6 cytokine release in bystander responses: The roles of radiation dose, radiation quality and specific ros/rns scavengers. Int. J. Radiat. Biol. 2012, 88, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.; Wang, Y.; Doetsch, P.W. A single exposure to low- or high-let radiation induces persistent genomic damage in mouse epithelial cells in vitro and in lung tissue. Radiat. Res. 2017, 188, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Kook, S.H.; Song, J.H.; Lee, J.C. A phenolic acid phenethyl urea derivative protects against irradiation-induced osteoblast damage by modulating intracellular redox state. J. Cell. Biochem. 2014, 115, 1877–1887. [Google Scholar] [PubMed]
- Mi Choi, E.; Sik Suh, K.; Jung, W.W.; Young Park, S.; Ouk Chin, S.; Youl Rhee, S.; Kim Pak, Y.; Chon, S. Actein alleviates 2,3,7,8-tetrachlorodibenzo-p-dioxin-mediated cellular dysfunction in osteoblastic mc3t3-e1 cells. Environ. Toxicol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Campbell-Beachler, M.; Gifford, P.; Haynes, K.E.; Gridley, D.S.; Pecaut, M.J. Role of nadph oxidase as a mediator of oxidative damage in low-dose irradiated and hindlimb-unloaded mice. Radiat. Res. 2017, 188, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.R.; Speacht, T.L.; Zhang, Y.; Lang, C.H.; Donahue, H.J. Simulated space radiation sensitizes bone but not muscle to the catabolic effects of mechanical unloading. PLoS ONE 2017, 12, e0182403. [Google Scholar] [CrossRef] [PubMed]
- Shahnazari, M.; Dwyer, D.; Chu, V.; Asuncion, F.; Stolina, M.; Ominsky, M.; Kostenuik, P.; Halloran, B. Bone turnover markers in peripheral blood and marrow plasma reflect trabecular bone loss but not endocortical expansion in aging mice. Bone 2012, 50, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Sham | 10 cGy | 50 cGy | 200 cGy |
|---|---|---|---|---|
| Grey/Not Grey Coat (%) | 1/12 (7.7%) | 1/12 (7.7%) | 1/11 (8.3%) | 11/0 (100%) |
| Percent Bone Volume (%) | 9.7 ± 3.1 | 8.7 ± 2.4 | 8.2 ± 2.9 | 7.2 ± 1.5 |
| Trabecular Thickness (μm) | 59.3 ± 5.3 | 58.2 ± 3.2 | 57.7 ± 2.9 | 57.1 ± 2.7 |
| Trabecular Separation (μm) | 228.8 ± 23.3 | 245.0 ± 41.8 | 240.1 ± 30.2 | 248.5 ± 18.4 |
| Trabecular Number (1/mm) | 1.62 ± 0.36 | 1.49 ± 0.35 | 1.42 ± 0.48 | 1.26 ± 0.23 |
| Proliferative Stage | Terminal Differentiation |
|---|---|
| Caspase 3 | Caspase 3 |
| Cdk2 | Catalase |
| CuZnSOD | Cdk2 |
| Foxo1 | Gadd45 |
| GPX | iNos |
| MnSOD | MnSOD |
| p21 | p53 |
| p53 | PCNA |
| PCNA | - |
| Runx2 | - |
| Gene Name | Accession No. | Official Name | Process |
|---|---|---|---|
| Runx2 | NM_009820 | Runt Related Transcription Factor 2 | Differentiation |
| Bglap | NM_007541 | Bone γ carboxyglutamate protein (osteocalcin) | Differentiation |
| Alpl | NM_007431 | Alkaline Phosphatase Ligand | Differentiation |
| Pcna | NM_011045 | Proliferating Cell Nuclear Antigen | Proliferation |
| Cdk2 | NM_016756 | Cyclin-dependent Kinase 2 | Proliferation |
| p21 | NM_007669 | Cyclin-dependent Kinase Inhibitor 1 | Proliferation |
| p53 | NM_011640 | Transformation-related protein 53 | Proliferation |
| Cat | NM_009804 | Catalase | Oxidative Metabolism |
| Gpx1 | NM_008160 | Glutathione Peroxidase | Oxidative Metabolism |
| MnSod | NM_013671 | Superoxide Dismutase 2, Mitochondrial | Oxidative Metabolism |
| CuZnSod | NM_011434 | Superoxide Dismutase | Oxidative Metabolism |
| iNos [Nos2] | NM_010927 | Nitric Oxide Synthase 2, Inducible | Oxidative Metabolism |
| Foxo1 | NM_019739 | Forkhead Box | Oxidative Metabolism |
| Gadd45a | NM_007836 | Growth Arrest and DNA Damage | Oxidative Metabolism |
| Caspase 3 | NM_009810 | Caspase 3 | Apoptosis |
| Hprt | NM_013556 | Hypoxanthine-guanine phosphoribosyltransferase | Housekeeping |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alwood, J.S.; Tran, L.H.; Schreurs, A.-S.; Shirazi-Fard, Y.; Kumar, A.; Hilton, D.; Tahimic, C.G.T.; Globus, R.K. Dose- and Ion-Dependent Effects in the Oxidative Stress Response to Space-Like Radiation Exposure in the Skeletal System. Int. J. Mol. Sci. 2017, 18, 2117. https://doi.org/10.3390/ijms18102117
Alwood JS, Tran LH, Schreurs A-S, Shirazi-Fard Y, Kumar A, Hilton D, Tahimic CGT, Globus RK. Dose- and Ion-Dependent Effects in the Oxidative Stress Response to Space-Like Radiation Exposure in the Skeletal System. International Journal of Molecular Sciences. 2017; 18(10):2117. https://doi.org/10.3390/ijms18102117
Chicago/Turabian StyleAlwood, Joshua S., Luan H. Tran, Ann-Sofie Schreurs, Yasaman Shirazi-Fard, Akhilesh Kumar, Diane Hilton, Candice G. T. Tahimic, and Ruth K. Globus. 2017. "Dose- and Ion-Dependent Effects in the Oxidative Stress Response to Space-Like Radiation Exposure in the Skeletal System" International Journal of Molecular Sciences 18, no. 10: 2117. https://doi.org/10.3390/ijms18102117