Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation

 and
and
Abstract
:1. Introduction
2. Results
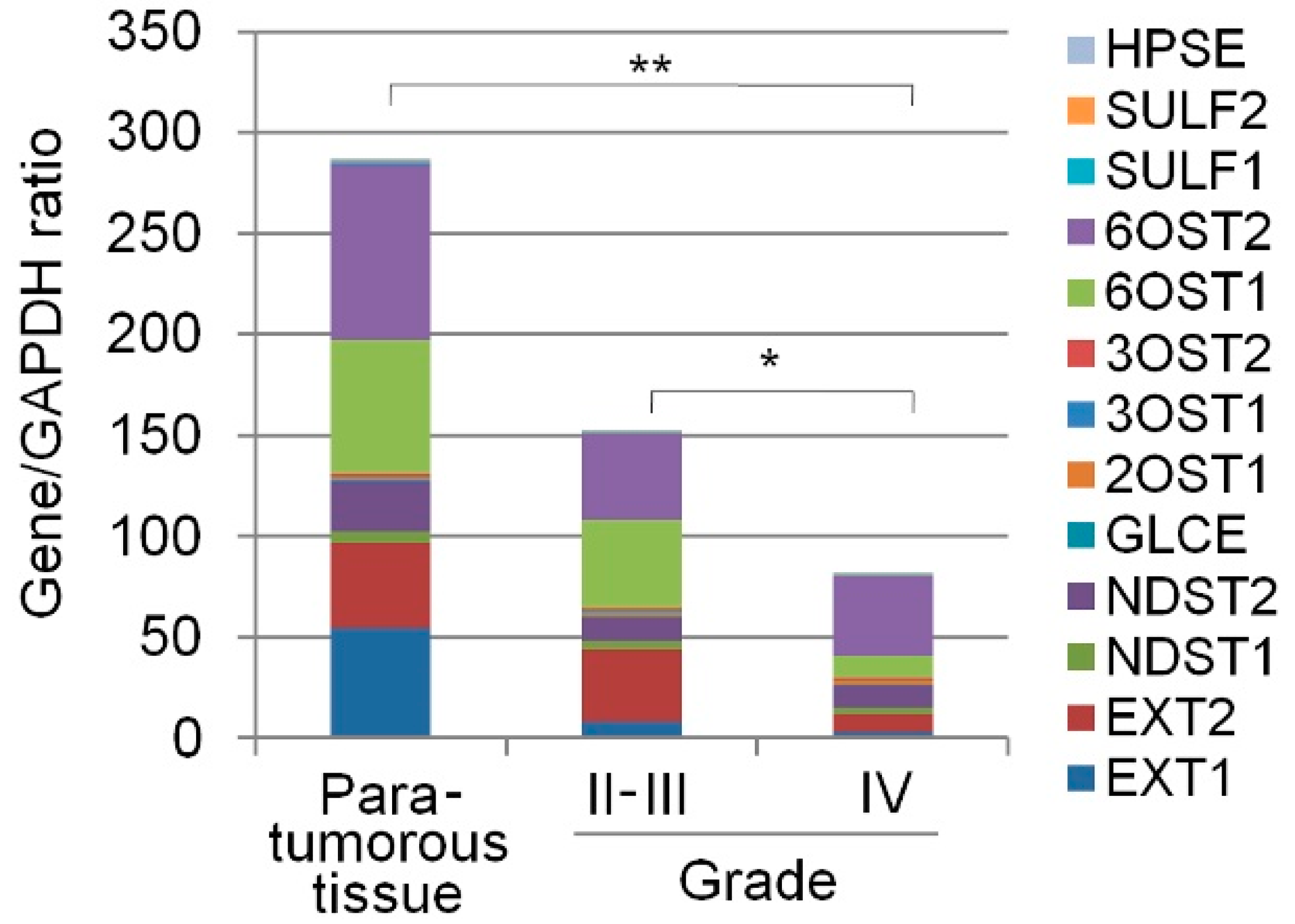
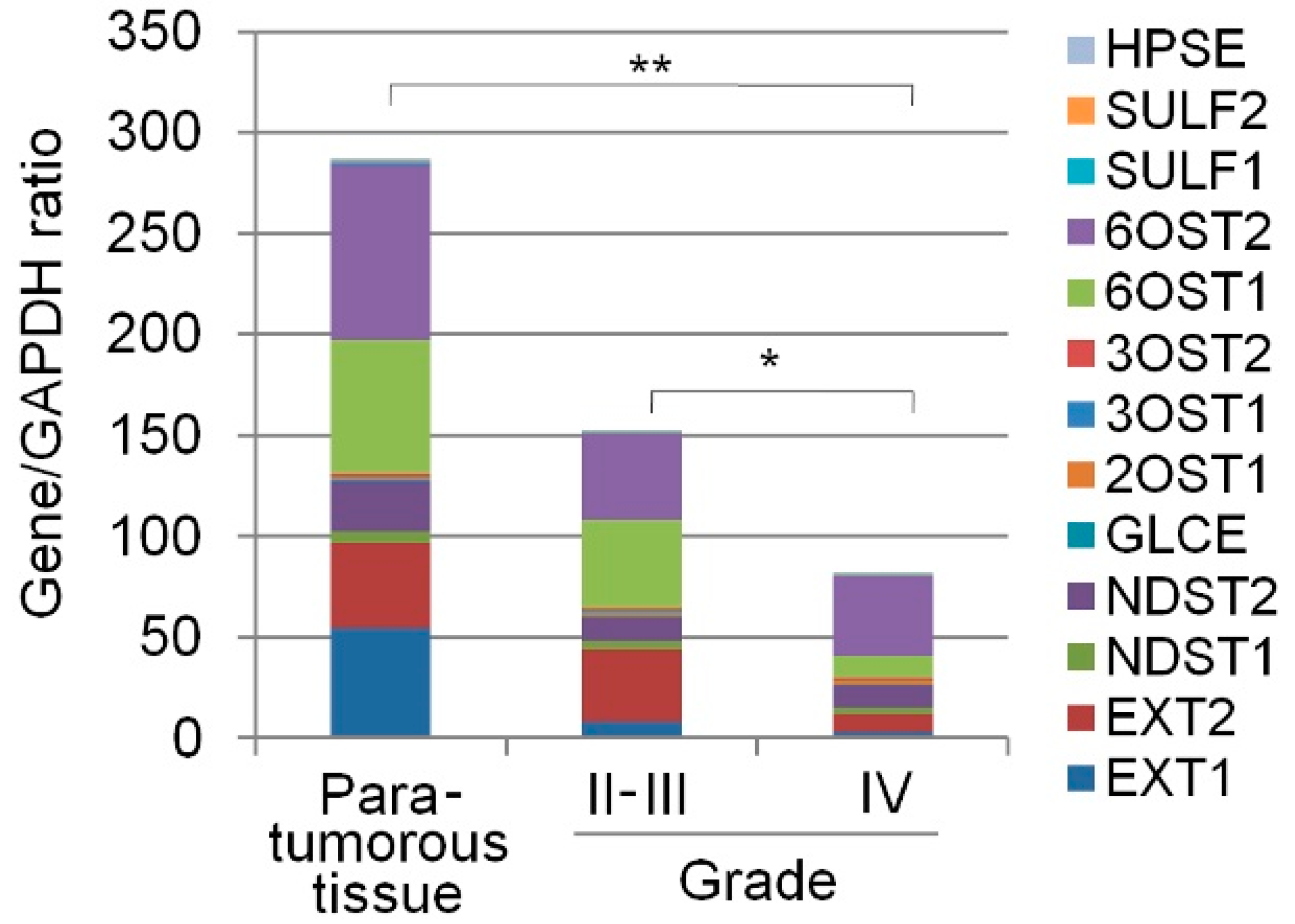
2.1. Transcriptional Activity of HS(Heparan sulfate)Biosynthetic System Is Inhibited in Glioma
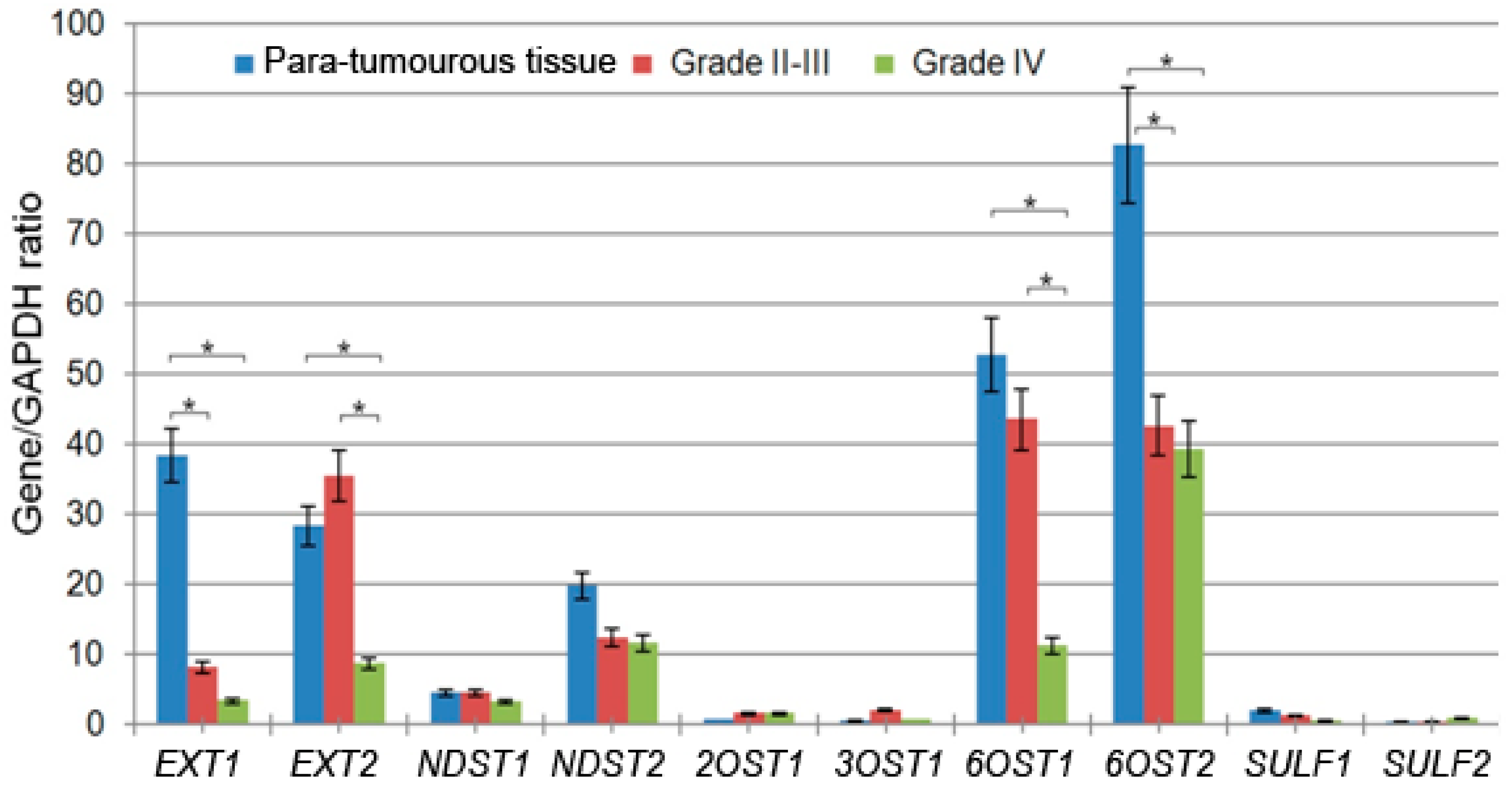
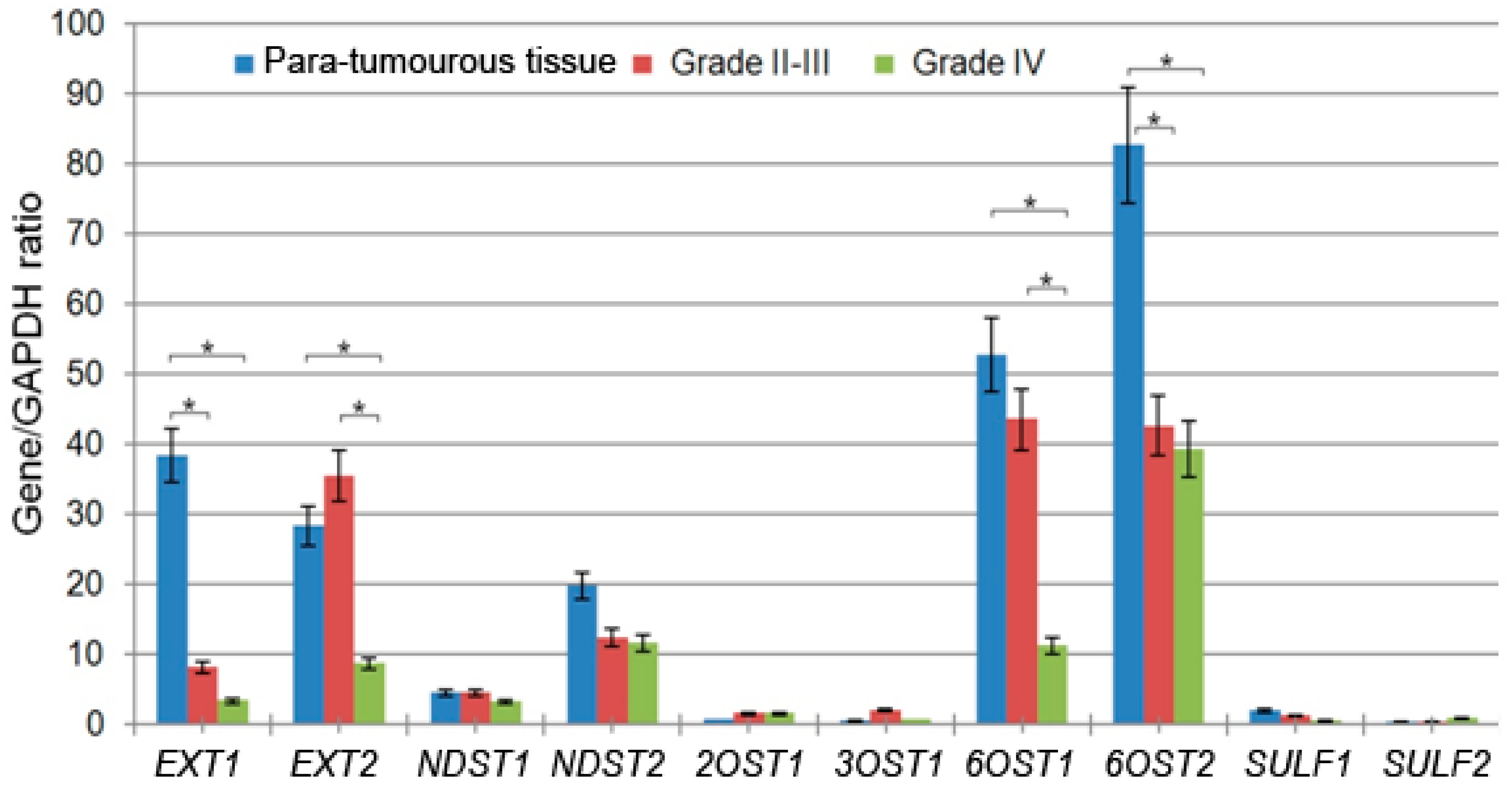
2.2. Elongation and 6-O-Sulfation Steps Are Main Contributors to HS Biosynthetic System Inhibition
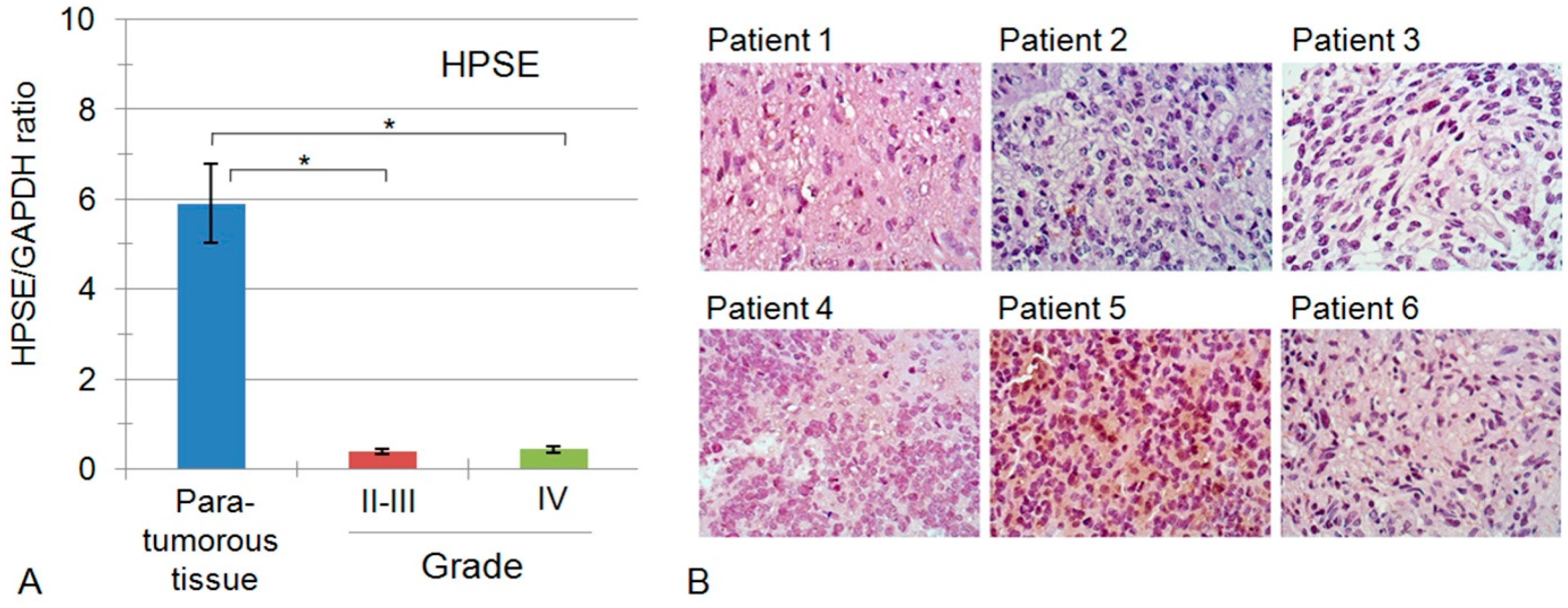
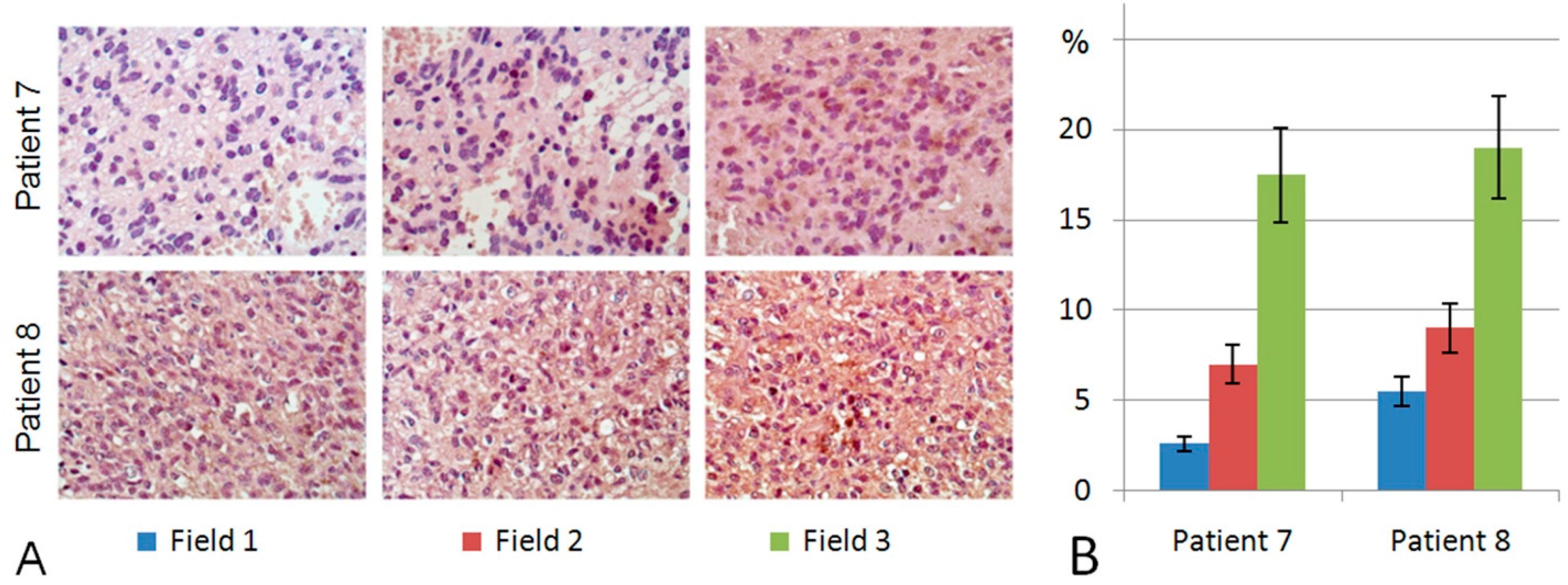
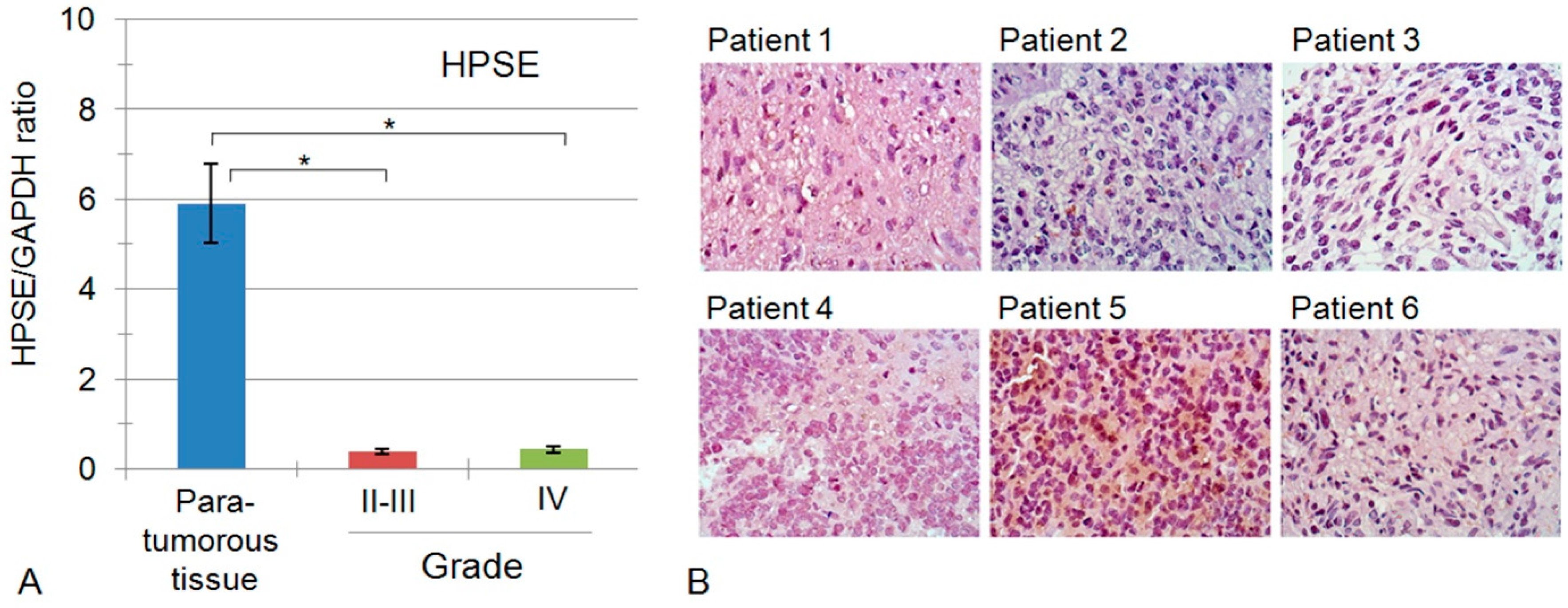
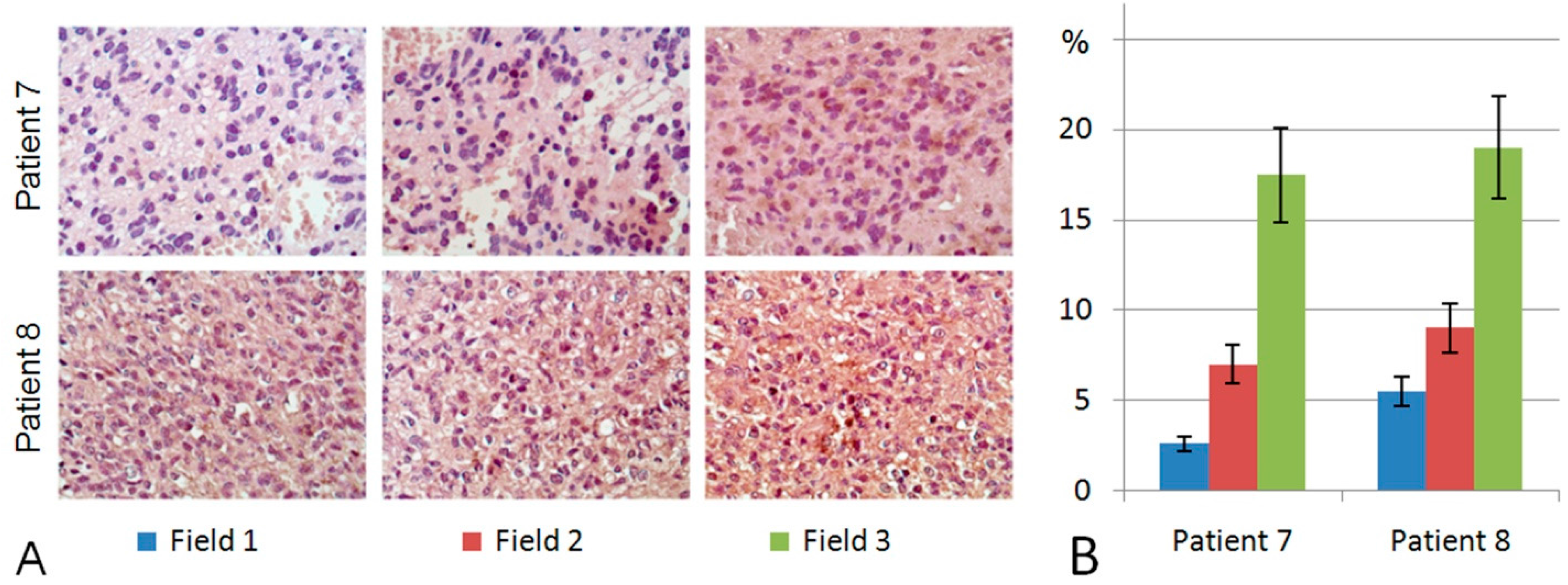
2.3. Heparanase Expression in Glioblastoma Possesses Both Inter- and Intratumour Heterogeneity
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. RT-PCR Analysis
4.3. Immunohistochemical Analysis
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HS | Heparan Sulfate |
| HSPG | Heparan Sulfate Proteoglycan |
| GBM | Glioblastoma Multiforme |
| ECM | Extracellular Matrix |
| TME | Tumour Microenvironment |
References
- Xiao, W.; Sohrabi, A.; Seidlits, S.K. Integrating the glioblastoma microenvironment into engineered experimental models. Future Sci. OA 2017, 3, FSO189. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, C.; Hrabětová, S. Brain extracellular space: The final frontier of neuroscience. Biophys. J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.D.; Coulson-Thomas, V.J.; Foscarin, S.; Kwok, J.C.; Fawcett, J.W. GAG-ing with the neuron: The role of glycosaminoglycan patterning in the central nervous system. Exp. Neurol. 2015, 274, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S.; Kitagawa, H. Formation and remodeling of the brainextracellular matrix in neural plasticity: Roles of chondroitin sulfate and hyaluronan. Biochim. Biophys. Acta 2017, 1861, 2420–2434. [Google Scholar] [CrossRef] [PubMed]
- Swarup, V.P.; Mencio, C.P.; Hlady, V.; Kuberan, B. Sugar glues for broken neurons. Biomol. Concepts 2013, 4, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Malavaki, C.J.; Theocharis, A.D.; Lamari, F.N.; Kanakis, I.; Tsegenidis, T.; Tzanakakis, G.N.; Karamanos, N.K. Heparan sulfate: Biological significance, tools for biochemical analysis and structural characterization. Biomed. Chromatogr. 2011, 25, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.J.; Esko, J.D.; Tor, Y. Targeting heparin and heparan sulfate protein interactions. Org. Biomol. Chem. 2017, 15, 5656–5668. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparansulphate proteoglycans fine-tune mammalian physiology. Nature 2007, 26, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E.I.; Johnson, Z.; Bonvin, P.; Handel, T.M. Glycosaminoglycan interactions with chemokines add complexity to a complex system. Pharm. Basel. 2017, 10, E70. [Google Scholar] [CrossRef] [PubMed]
- Monneau, Y.; Arenzana-Seisdedos, F.; Lortat-Jacob, H. The sweet spot: How GAGs help chemokines guide migrating cells. J. Leukoc. Biol. 2016, 99, 935–953. [Google Scholar] [CrossRef] [PubMed]
- Coulson-Thomas, V.J. The role of heparansulphate in development: The ectodermal story. Int. J. Exp. Pathol. 2016, 97, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Xiong, A.; Kundu, S.; Forsberg-Nilsson, K. Heparan sulfate in the regulation of neural differentiation and glioma development. FEBS J. 2014, 281, 4993–5008. [Google Scholar] [CrossRef] [PubMed]
- Steck, P.A.; Moser, R.P.; Bruner, J.M.; Liang, L.; Freidman, A.N.; Hwang, T.L.; Yung, W.K. Altered expression and distribution of heparan sulfate proteoglycans in human gliomas. Cancer Res. 1989, 49, 2096–2103. [Google Scholar] [PubMed]
- Tran, V.M.; Wade, A.; McKinney, A.; Chen, K.; Lindberg, O.R.; Engler, J.R.; Persson, A.I.; Phillips, J.J. Heparan sulfate glycosaminoglycans in glioblastoma promote tumour invasion. Mol. Cancer Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.; Gloster, T. The enzymatic degradation of heparan sulfate. Protein Pept. Lett. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kreuger, J.; Kjellén, L. Heparan sulfate biosynthesis: Regulation and variability. J. Histochem. Cytochem. 2012, 60, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, F.; Sheng, J. “Coding” and “Decoding”: Hypothesis for the regulatory mechanism involved in heparan sulfate biosynthesis. Carbohydr. Res. 2016, 428, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Suhovskih, A.V.; Domanitskaya, N.V.; Tsidulko, A.Y.; Prudnikova, T.Y.; Kashuba, V.I.; Grigorieva, E.V. Tissue-specificity of heparan sulfate biosynthetic machinery in cancer. Cell Adhes. Migr. 2015, 9, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Hull, E.E.; Montgomery, M.R.; Leyva, K.J. Epigenetic regulation of the biosynthesis & enzymatic modification of heparan sulfate proteoglycans: Implications for tumorigenesis and cancer biomarkers. Int. J. Mol. Sci. 2017, 18, E1361. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas (TCGA) Research Network. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar]
- Wade, A.; Robinson, A.E.; Engler, J.R.; Petritsch, C.; James, C.D.; Phillips, J.J. Proteoglycans and their roles in brain cancer. FEBS J. 2013, 280, 2399–2417. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.J.; Huillard, E.; Robinson, A.E.; Ward, A.; Lum, D.H.; Polley, M.Y.; Rosen, S.D.; Rowitch, D.H.; Werb, Z. Heparan sulfatesulfatase SULF2 regulates PDGFRα signaling and growth in human and mouse malignant glioma. J. Clin. Investig. 2012, 122, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Bret, C.; Moreaux, J.; Schved, J.F.; Hose, D.; Klein, B. SULFs in human neoplasia: Implication as progression and prognosis factors. J. Transl. Med. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, R.D.; Elkin, M.; Rapraeger, A.C.; Ilan, N.; Vlodavsky, I. Heparanase regulation of cancer, autophagy and inflammation: New mechanisms and targets for therapy. FEBS J. 2017, 284, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Zhou, S. The Functions of Heparanase in Human Diseases. Mini Rev. Med. Chem. 2017, 17, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Rivara, S.; Milazzo, F.M.; Giannini, G. Heparanase: A rainbow pharmacological target associated to multiple pathologies including rare diseases. Future Med. Chem. 2016, 8, 647–680. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Jiang, F.; Kalkanis, S.N.; Zhang, Z.G.; Zhang, X.; Zheng, X.; Jiang, H.; Mikkelsen, T.; Chopp, M. Increased chemotactic migration and growth inheparanase-overexpressing human U251ngliomacells. Cancer Res. 2008, 10, 23–27. [Google Scholar] [CrossRef]
- Zetser, A.; Bashenko, Y.; Miao, H.Q.; Vlodavsky, I.; Ilan, N. Heparanase affects adhesive and tumorigenic potential of human glioma cells. Cancer Res. 2003, 15, 63–85. [Google Scholar]
- Ueno, Y.; Yamamoto, M.; Vlodavsky, I.; Pecker, I.; Ohshima, K.; Fukushima, T. Decreased expression of heparanase in glioblastomamultiforme. J. Neurosurg. 2005, 102, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Xiong, A.; Spyrou, A.; Wicher, G.; Marinescu, V.D.; Edqvist, P.D.; Zhang, L.; Essand, M.; Dimberg, A.; Smits, A.; et al. Heparanase promotes glioma progression and is inversely correlated with patient survival. Mol. Cancer Res. 2016, 14, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Nelson, K.K.; de Carvalho, A.C.; Kalkanis, S.N. Heparanaseexpression ofgliomain human and animal models. J. Neurosurg. 2010, 113, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Suhovskih, A.V.; Tsidulko, A.Y.; Kutsenko, O.S.; Kovner, A.V.; Aidagulova, S.V.; Ernberg, I.; Grigorieva, E.V. Transcriptional activity of heparan sulfate biosynthetic machinery is specifically impaired in benign prostate hyperplasia and prostate cancer. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Tsidulko, A.Y.; Matskova, L.; Astakhova, L.A.; Ernberg, I.; Grigorieva, E.V. Proteoglycan expression correlates with the phenotype of malignant and non-malignant EBV-positive B-cell lines. Oncotarget 2015, 6, 43529–43539. [Google Scholar] [CrossRef] [PubMed]
- Suhovskih, A.V.; Aidagulova, S.V.; Kashuba, V.I.; Grigorieva, E.V. Proteoglycans as potential microenvironmental biomarkers for colon cancer. Cell Tissue Res. 2015, 361, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Soares da Costa, D.; Reis, R.L.; Pashkuleva, I. Sulfation of glycosaminoglycans and its implications in human health and disorders. Annu. Rev. Biomed. Eng. 2017, 19, 1–26. [Google Scholar] [CrossRef] [PubMed]
- El Masri, R.; Seffouh, A.; Lortat-Jacob, H.; Vivès, R.R. The “in and out” of glucosamine 6-O-sulfation: The 6th sense of heparan sulfate. Glycoconj. J. 2017, 34, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Kerever, A.; Yoshimura, Y.; Suzuki, Y.; Nonaka, R.; Higashi, K.; Toida, T.; Mercier, F.; Arikawa-Hirasawa, E. Heparan sulfate alterations in extracellular matrix structures and fibroblast growth factor-2 signaling impairment in the aged neurogenic niche. J. Neurochem. 2017, 142, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.; Khurana, A.; Shridhar, V.; Dredge, K. The role of heparanase and sulfatases in the modification of heparan sulfate proteoglycans within the tumour microenvironment and opportunities for novel cancer therapeutics. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Primary Source | Gene Name | Description | Para-Tumourous Tissue | Grade II-III | Grade IV |
|---|---|---|---|---|---|---|
| 2131 | HGNC:3512 | EXT1 | Exostosin Glycosyltransferase 1 | 54.9 ± 58 | 8.2 ± 8.38 | 3.4 ± 2.1 |
| 2132 | HGNC:3513 | EXT2 | Exostosin Glycosyltransferase 2 | 41.5 ± 54 | 35.5 ± 54 | 8.7 ± 5.7 |
| 3340 | HGNC:7680 | NDST1 | N-deacetylase and N-sulfotransferase 1 | 6.2 ± 2 | 4.5 ± 7 | 3.2 ± 6 |
| 8509 | HGNC:7681 | NDST2 | N-deacetylase and N-sulfotransferase 2 | 25.5 ± 4 | 12.4 ± 8 | 11.6 ± 14 |
| 26035 | HGNC:17855 | GLCE | Glucuronic Acid Epimerase | 0.001 ± 0.001 | 0.005 ± 0.008 | 0.002 ± 0.001 |
| 9653 | HGNC:5193 | 2OST1/HS2ST1 | Heparan Sulfate 2-O-Sulfotransferase 1 | 1.12 ± 1.4 | 1.6 ± 1.33 | 1.58 ± 1.1 |
| 9957 | HGNC:5194 | 3OST1/HS3ST1 | Heparan Sulfate-Glucosamine 3-O-Sulfotransferase 1 | 0.8 ± 1.0 | 2.05 ± 1.3 | 0.81 ± 1 |
| 9956 | HGNC:5195 | 3OST2/HS3ST2 | Heparan Sulfate-Glucosamine 3-O-Sulfotransferase 2 | 0.84 ± 0.35 | 0.22 ± 0.31 | 0.24 ± 0.39 |
| 9394 | HGNC:5201 | 6OST1/HS6ST1 | Heparan Sulfate 6-O-Sulfotransferase 1 | 66.6 ± 13.9 | 43.6 ± 49 | 11.2 ± 11 |
| 90161 | HGNC:19133 | 6OST2/HS6ST2 | Heparan Sulfate 6-O-Sulfotransferase 2 | 86.8 ± 19 | 42.6 ± 43 | 39.3 ± 77 |
| 23213 | HGNC:20391 | SULF1 | Sulfatase 1 | 2.01 ± 2.1 | 1.2 ± 1.3 | 0.56 ± 0.88 |
| 55959 | HGNC:20392 | SULF2 | Sulfatase 2 | 0.37 ± 0.44 | 0.37 ± 0.43 | 0.85 ± 0.81 |
| 10855 | HGNC:5164 | HPSE | Heparanase | 0.51 ± 0.007 | 0.39 ± 0.058 | 0.44 ± 0.35 |
| Factors | n (Total = 42) | % |
|---|---|---|
| Age | ||
| Median (range) | 48.5 ± 12.1 | |
| >48.5 | 22 | 54.8 |
| ≤48.5 | 20 | 45.2 |
| Sex | ||
| Male | 19 | 45.2 |
| Female | 23 | 54.8 |
| Tumour Grade | ||
| II | 9 | 23.8 |
| III | 5 | 11.6 |
| IV | 28 | 66.6 |
| Relapse | ||
| Yes | 8 | 19.0 |
| No | 34 | 81.0 |
| Tumour development | ||
| Primary | 27 | 64.3 |
| Secondary | 15 | 35.7 |
| Death | ||
| Yes | 31 | 73.9 |
| No | 11 | 26.1 |
| Gene | Sequence |
|---|---|
| EXT1 | F 5′-AGCACAAGGATTCTCGCTGT-3′ |
| R 5′-GGAACCAGACAGAAAGTGGC-3′ | |
| EXT2 | F 5′-GTATTTTGCAGAGCATCCCC-3′ |
| R 5′-GGGCAATGGCTTTAATTGAC-3′ | |
| NDST1 | F 5′-GTATGTCAACCTGGACGCCT-3′ |
| R 5′-CACTCAGCAGGCTGTTCTCA-3′ | |
| NDST2 | F 5′-TGGTCCAAGGAGAAAACCTG-3′ |
| R 5′-GTGCAGGCTCAGGAAGAAGT-3′ | |
| GLCE | F 5′-CACCCCTTCTCTGCTGTCTC-3′ |
| R 5′-GAATCAAAACTTCAGTTTCTTGTCA-3′ | |
| 2OST1/HS2ST1 | F 5′-CCAGATCCAGAAACTGGAGG-3′ |
| R 5′-TCCATTGTATGTCGCTGCTC-3′ | |
| 3OST1/HS3ST1 | F 5′-CAGGAGCCTATTTAGGGCG-3′ |
| R 5′-AGCAGGGAAGCCTCCTAGTC-3′ | |
| 3OST2/HS3ST2 | F 5′-ACCCCACTTCTTTGACAGGA-3′ |
| R 5′-CAAAGTAGCTGGGCGTCTTC-3′ | |
| 6OST1/HS6ST1 | F 5′-CGACTGGACCGAGCTCAC-3′ |
| R 5′-GGTCTCGTAGCAGGGTGATG-3′ | |
| 6OST2/HS6ST2 | F 5′-TCACCAGCTGTGTGCCC-3′ |
| R 5′-GTGTCGGAGGATGGTGATGT-3′ | |
| SULF1 | F 5′-ATGCAGGTTCTTCAAGGCAG-3′ |
| R 5′-ATCCTGGTTGAATAATCAATCTCT-3′ | |
| SULF2 | F 5′-CAATAGCACTGGCTACCGGA-3′ |
| R 5′-TTTTTAAGGAGTCCGACCCA-3′ | |
| Heparanase-1/HPSE | F 5′-TTCCTGTCCGTCACCATTG-3′ |
| R 5′-TACGCAGGAGACAAGCCTCT-3′ | |
| GAPDH | F 5′-AAGGTGAAGGTCGGAGTCAA-3′ |
| R 5′-AATGAAGGGGTCATTGATGG-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ushakov, V.S.; Tsidulko, A.Y.; De La Bourdonnaye, G.; Kazanskaya, G.M.; Volkov, A.M.; Kiselev, R.S.; Kobozev, V.V.; Kostromskaya, D.V.; Gaytan, A.S.; Krivoshapkin, A.L.; et al. Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation. Int. J. Mol. Sci. 2017, 18, 2301. https://doi.org/10.3390/ijms18112301
Ushakov VS, Tsidulko AY, De La Bourdonnaye G, Kazanskaya GM, Volkov AM, Kiselev RS, Kobozev VV, Kostromskaya DV, Gaytan AS, Krivoshapkin AL, et al. Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation. International Journal of Molecular Sciences. 2017; 18(11):2301. https://doi.org/10.3390/ijms18112301
Chicago/Turabian StyleUshakov, Victor S., Alexandra Y. Tsidulko, Gabin De La Bourdonnaye, Galina M. Kazanskaya, Alexander M. Volkov, Roman S. Kiselev, Vyacheslav V. Kobozev, Diana V. Kostromskaya, Alexey S. Gaytan, Alexei L. Krivoshapkin, and et al. 2017. "Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation" International Journal of Molecular Sciences 18, no. 11: 2301. https://doi.org/10.3390/ijms18112301