Mutational Biases and GC-Biased Gene Conversion Affect GC Content in the Plastomes of Dendrobium Genus

Abstract
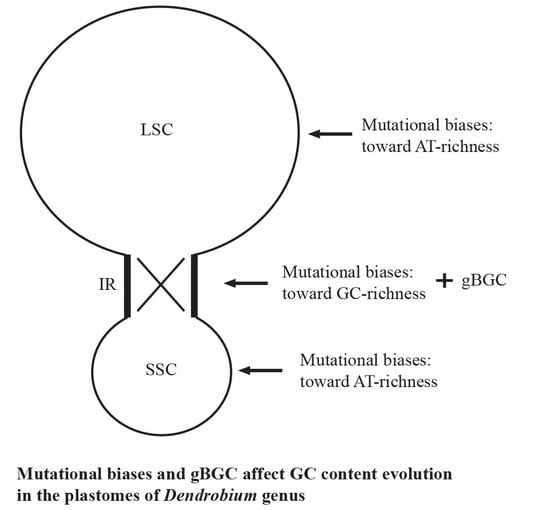
:
1. Introduction
2. Results
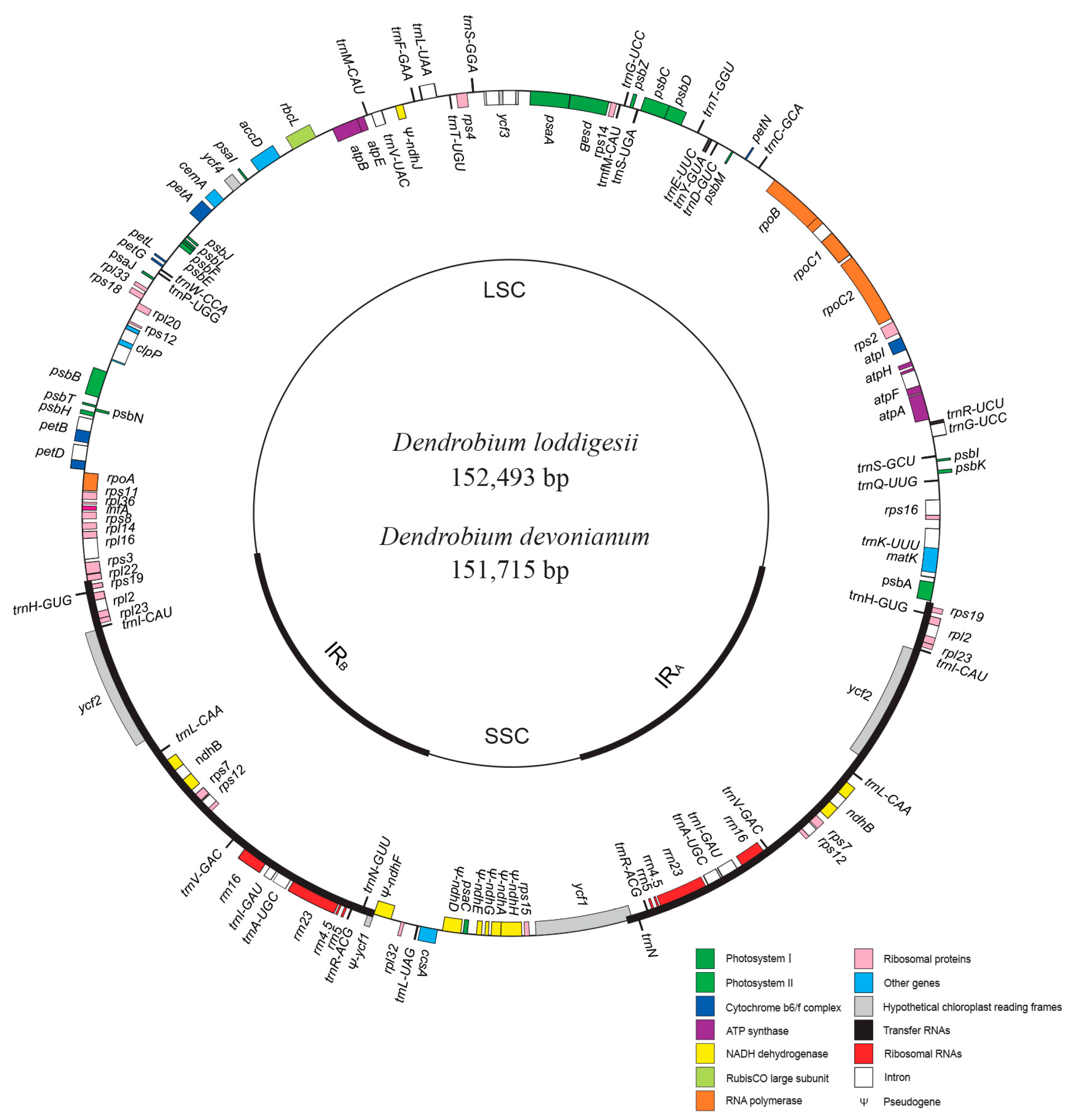
2.1. Plastome Assembly of D. loddigesii and D. devonianum
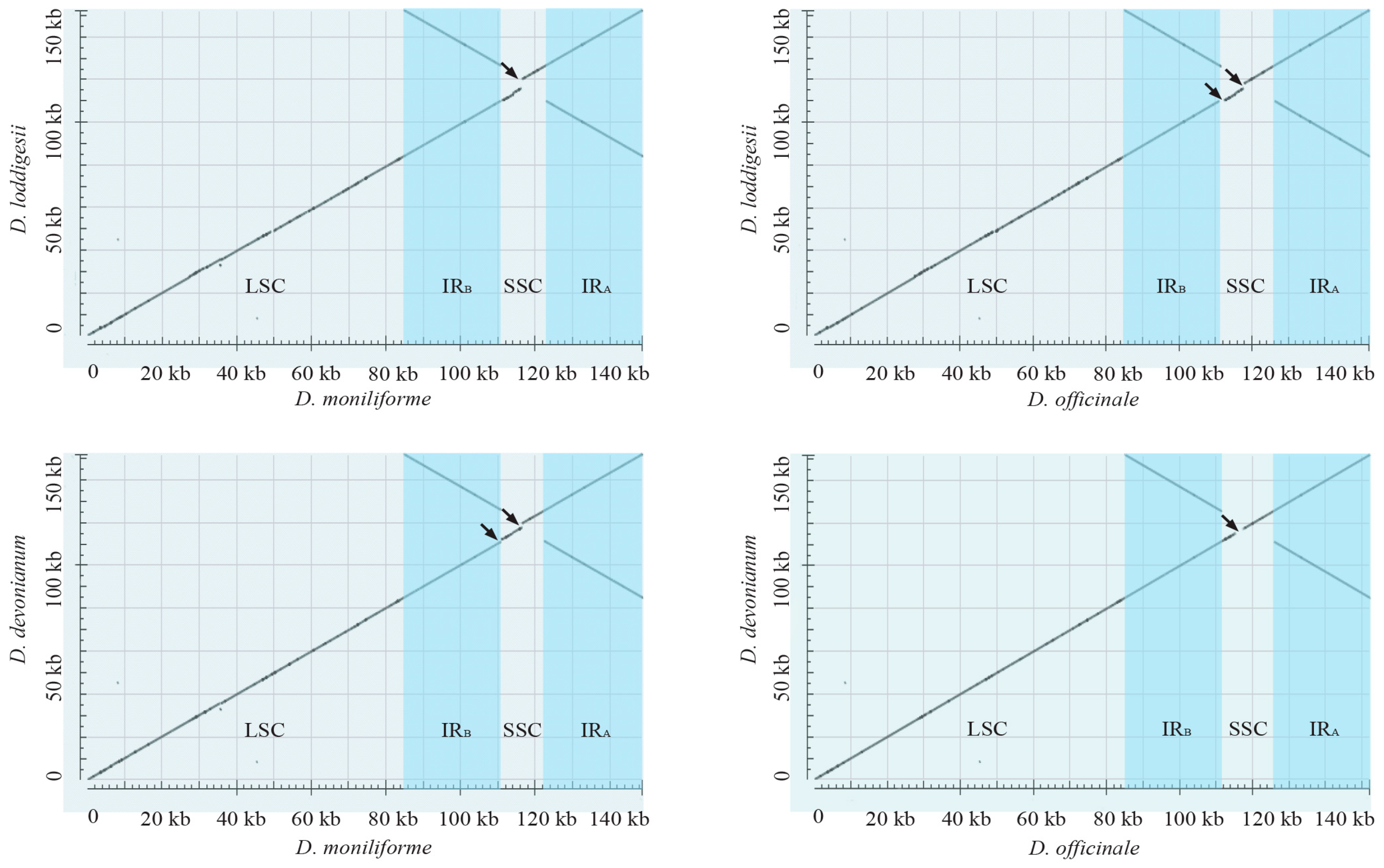
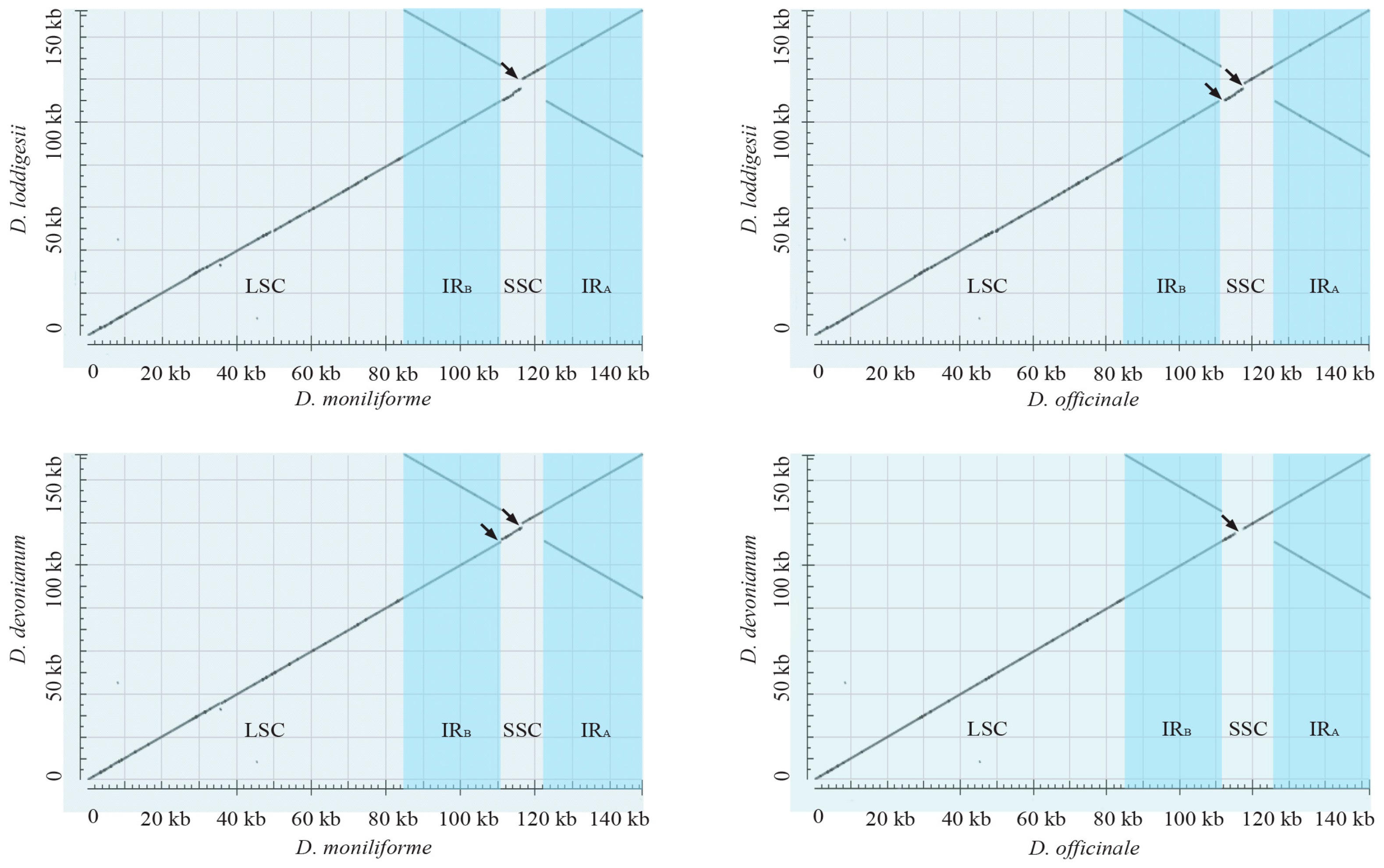
2.2. Highly Conserved Plastomic Structure and Organization in Dendrobium Genus
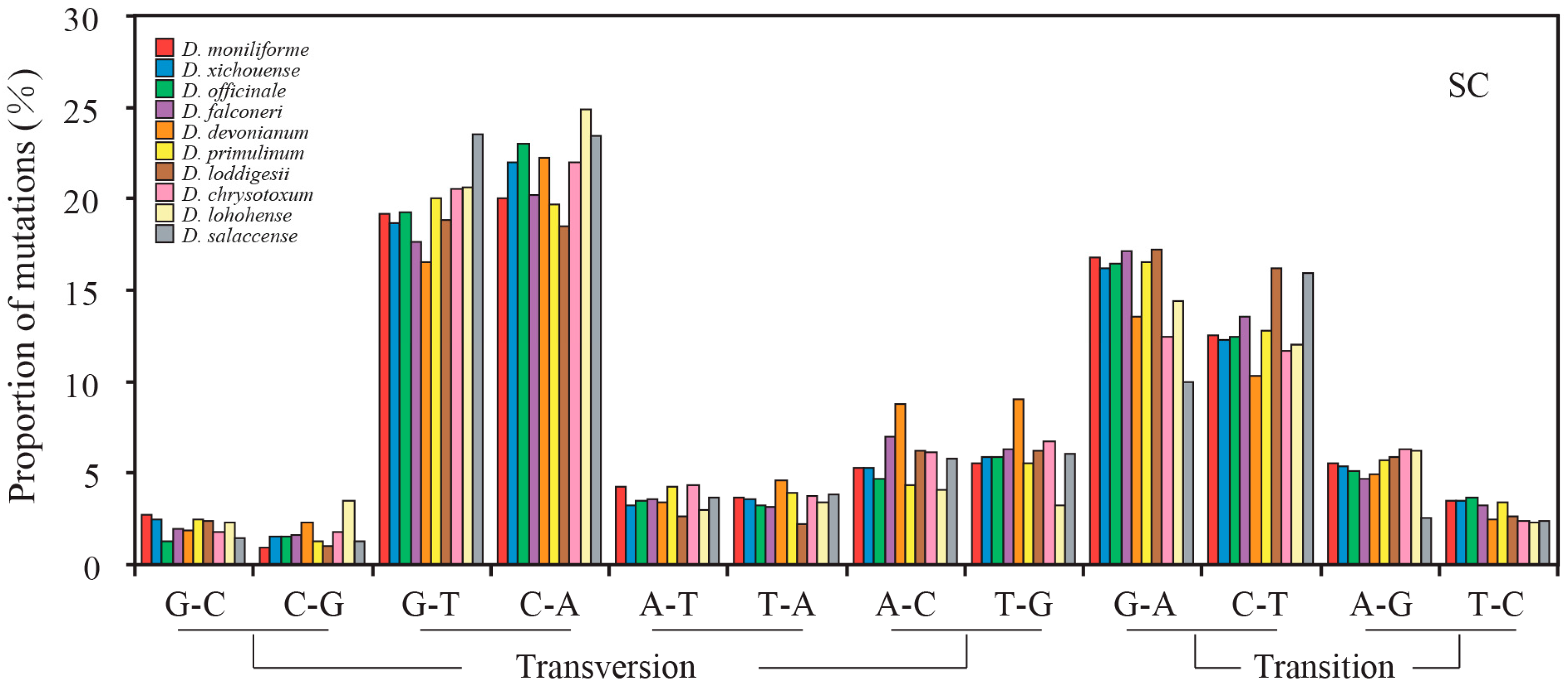
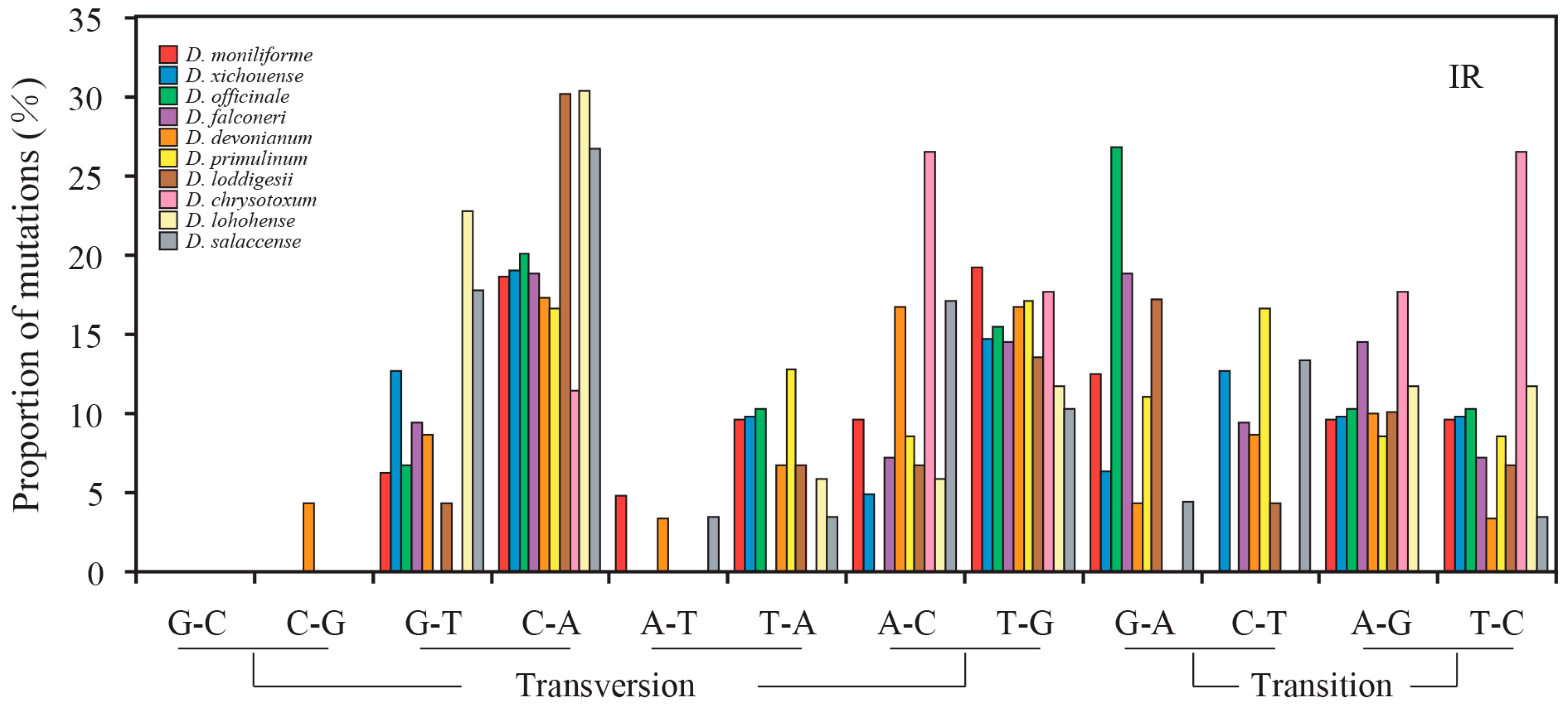
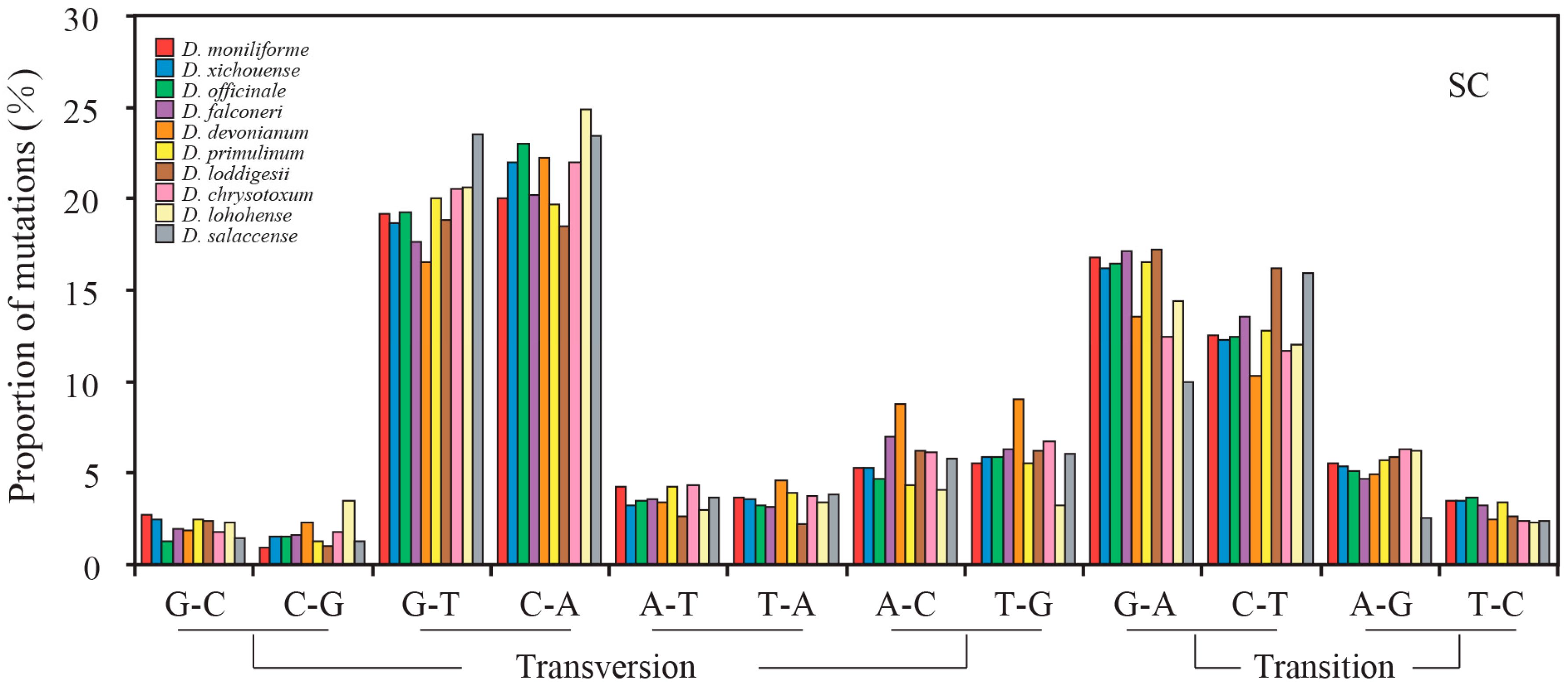
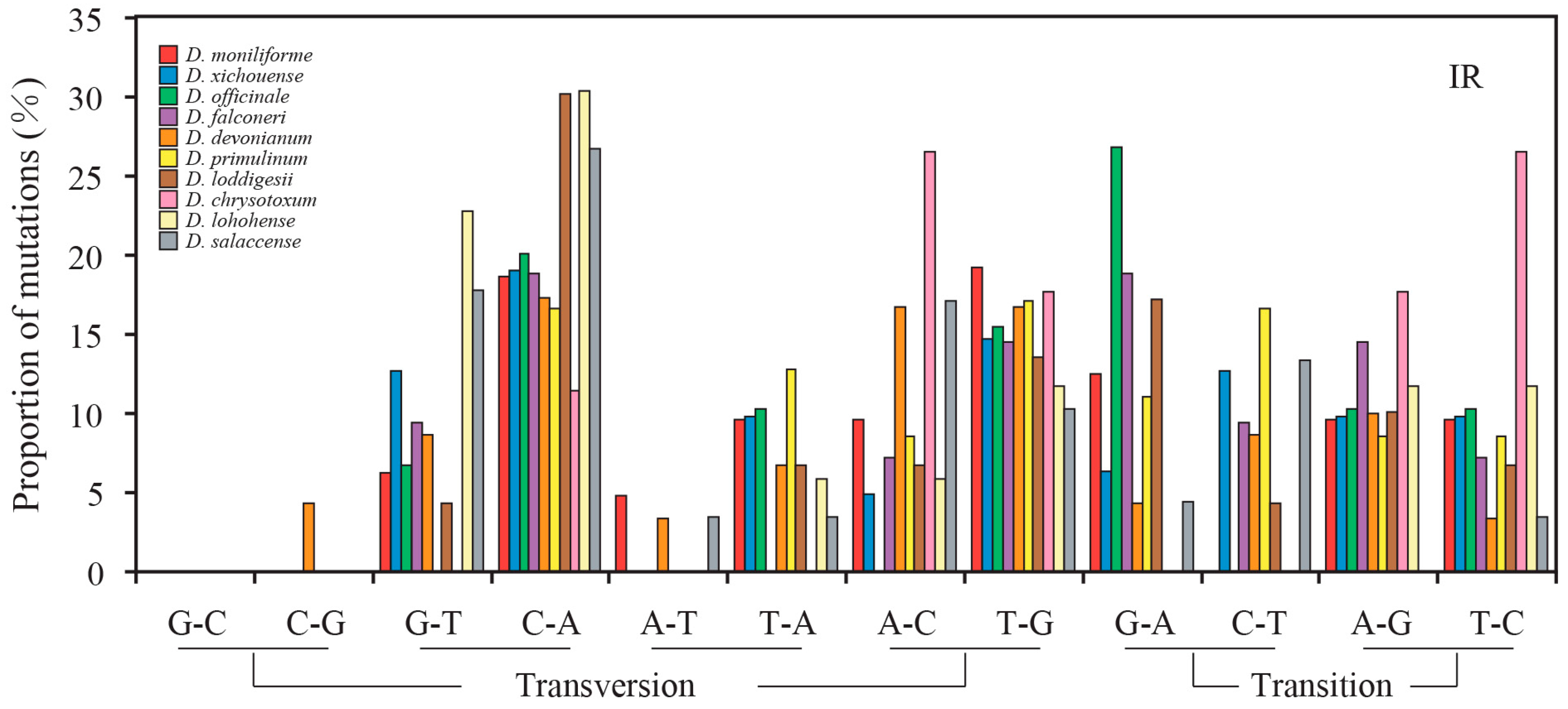
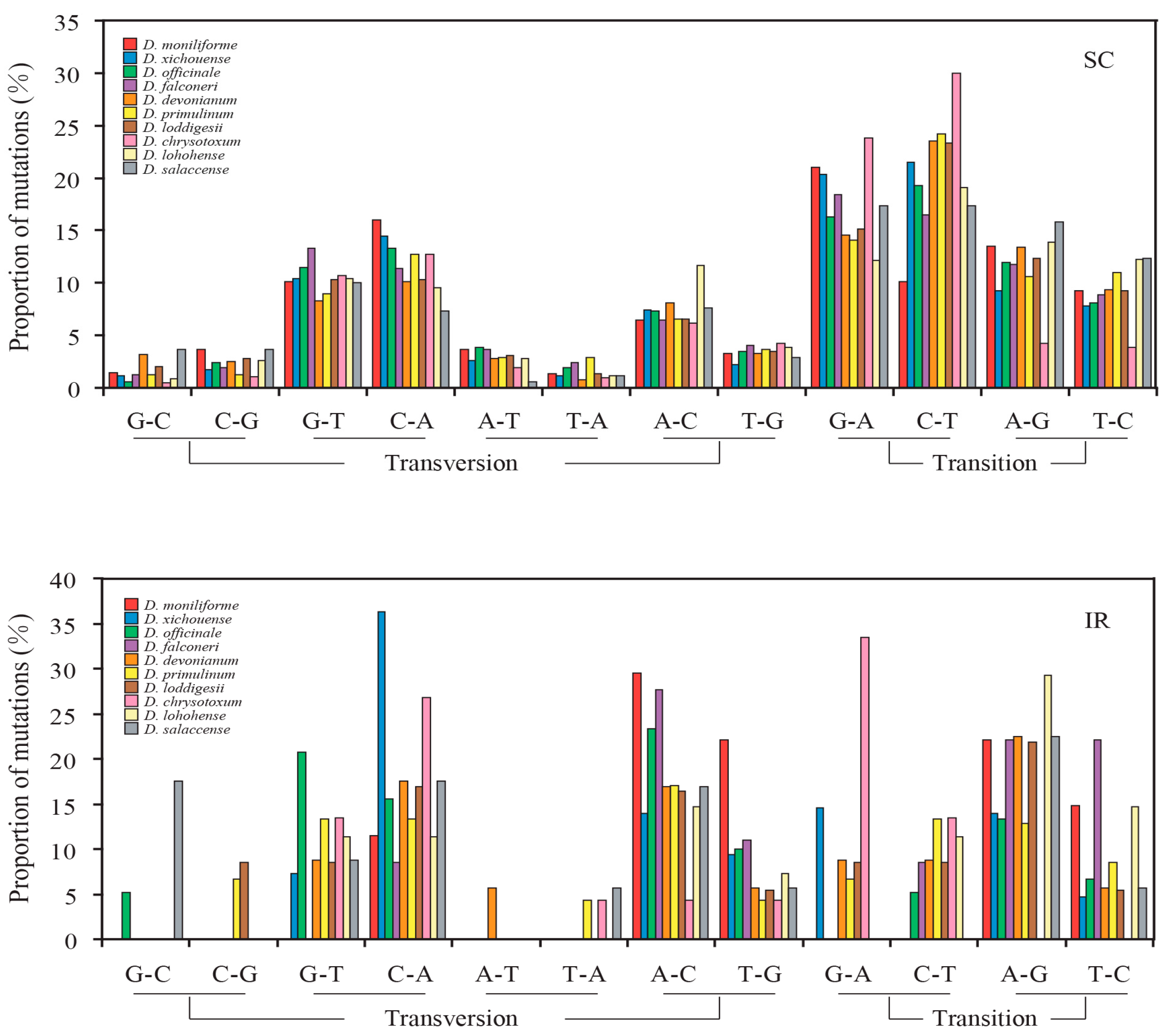
2.3. Mutational Biases of Non-Coding Loci Are Associated with the Plastomic Structure
2.4. Contrasting Mutational Biases of Protein-Coding Genes between SC and IR Regions
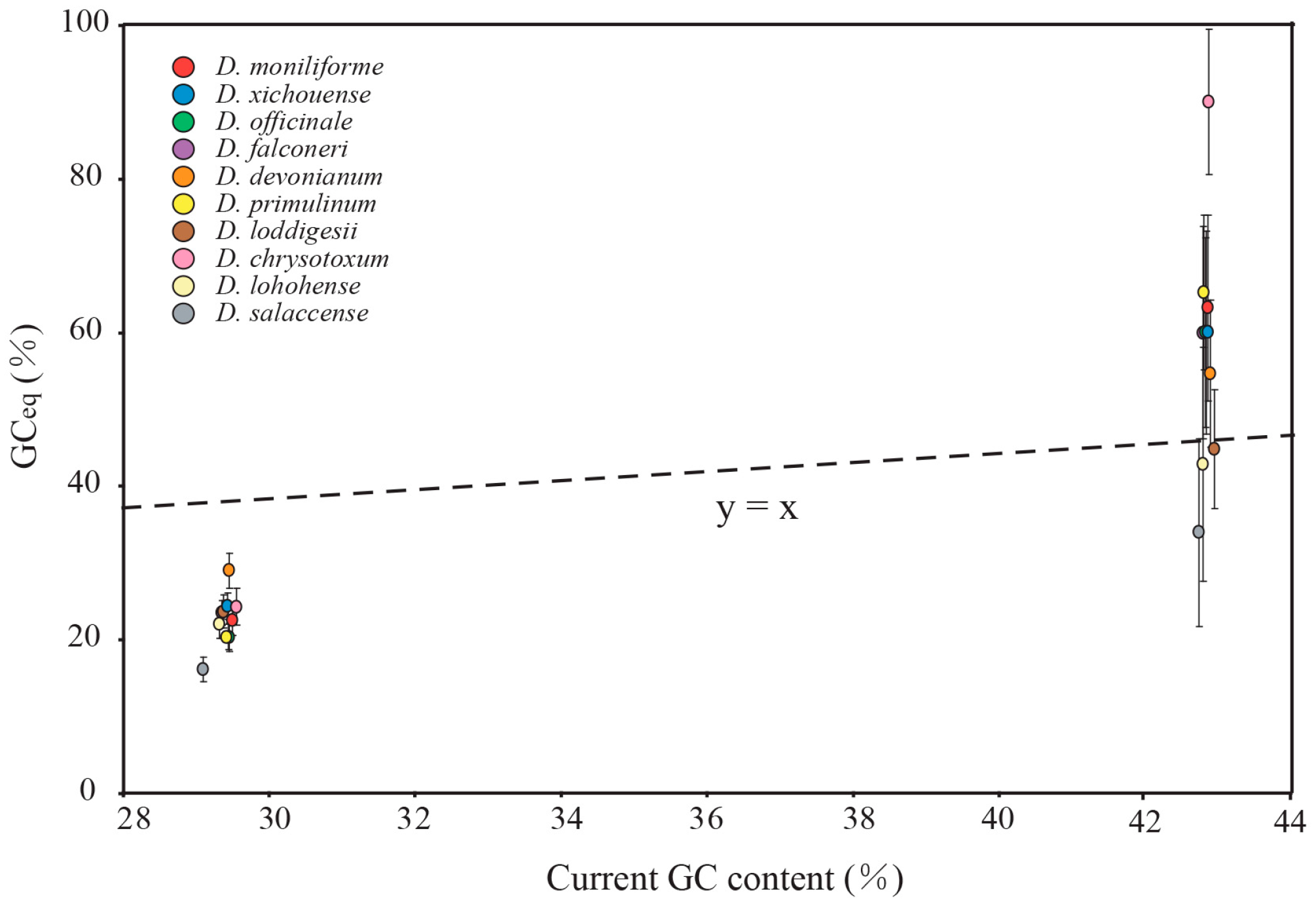
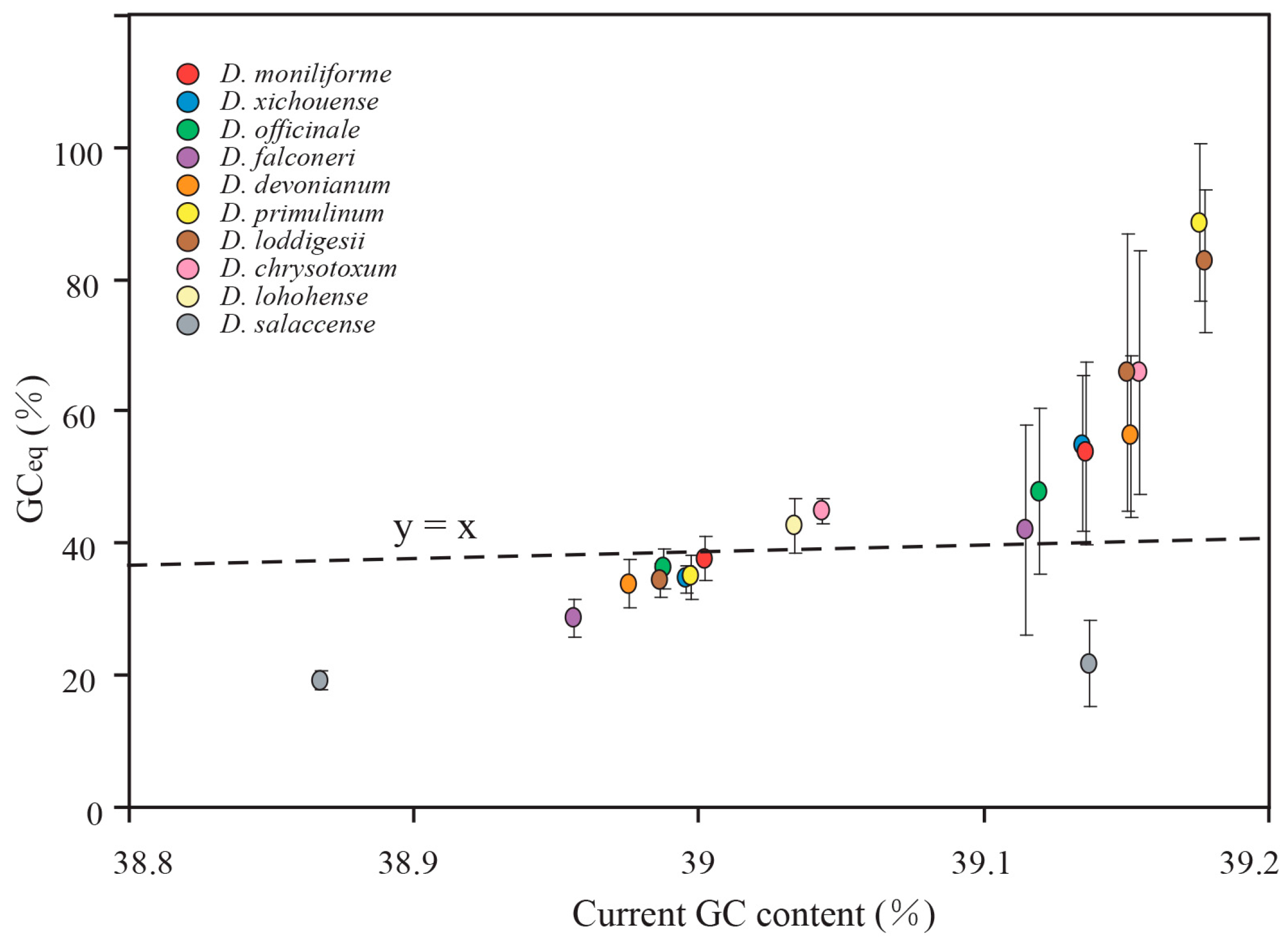
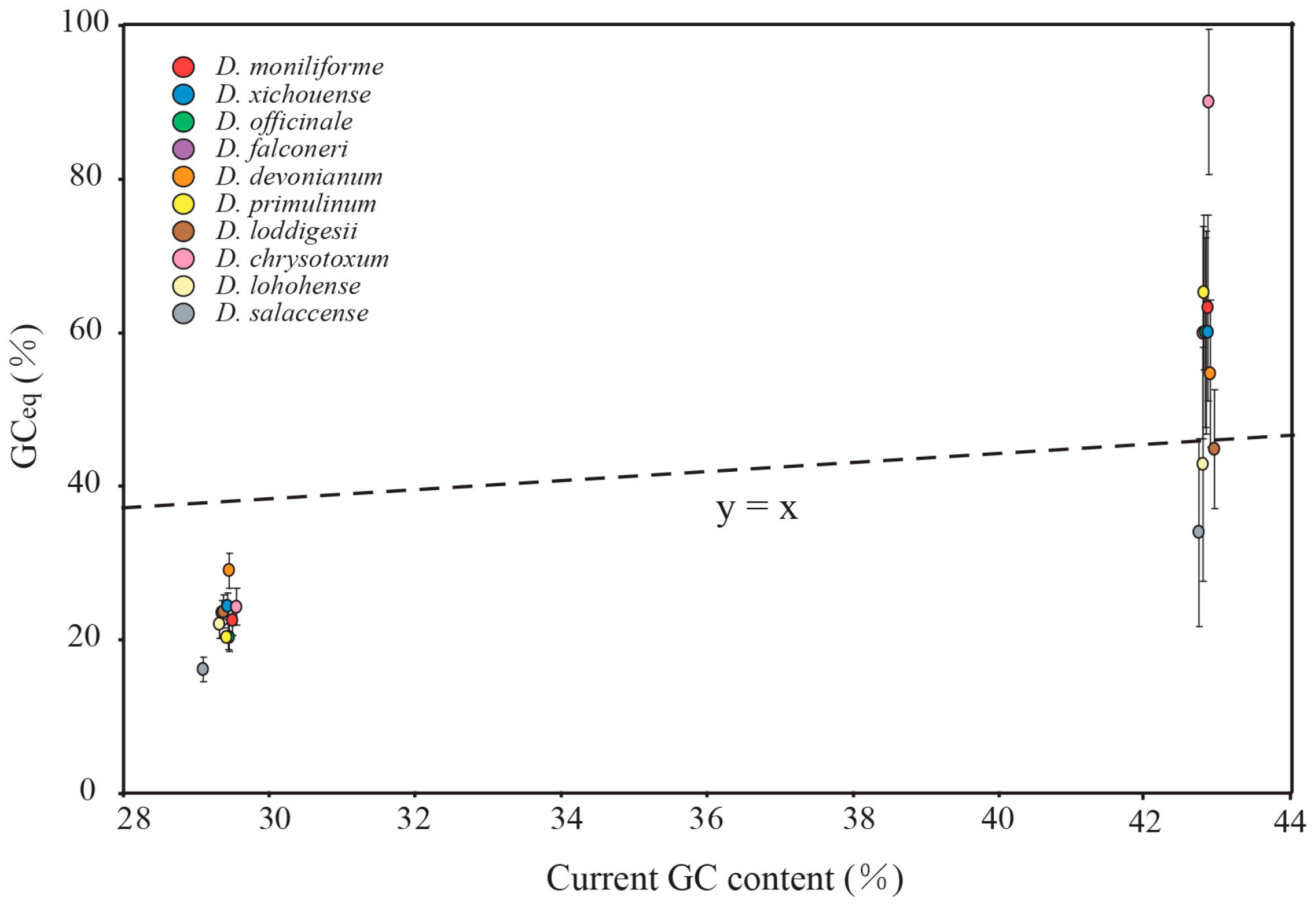
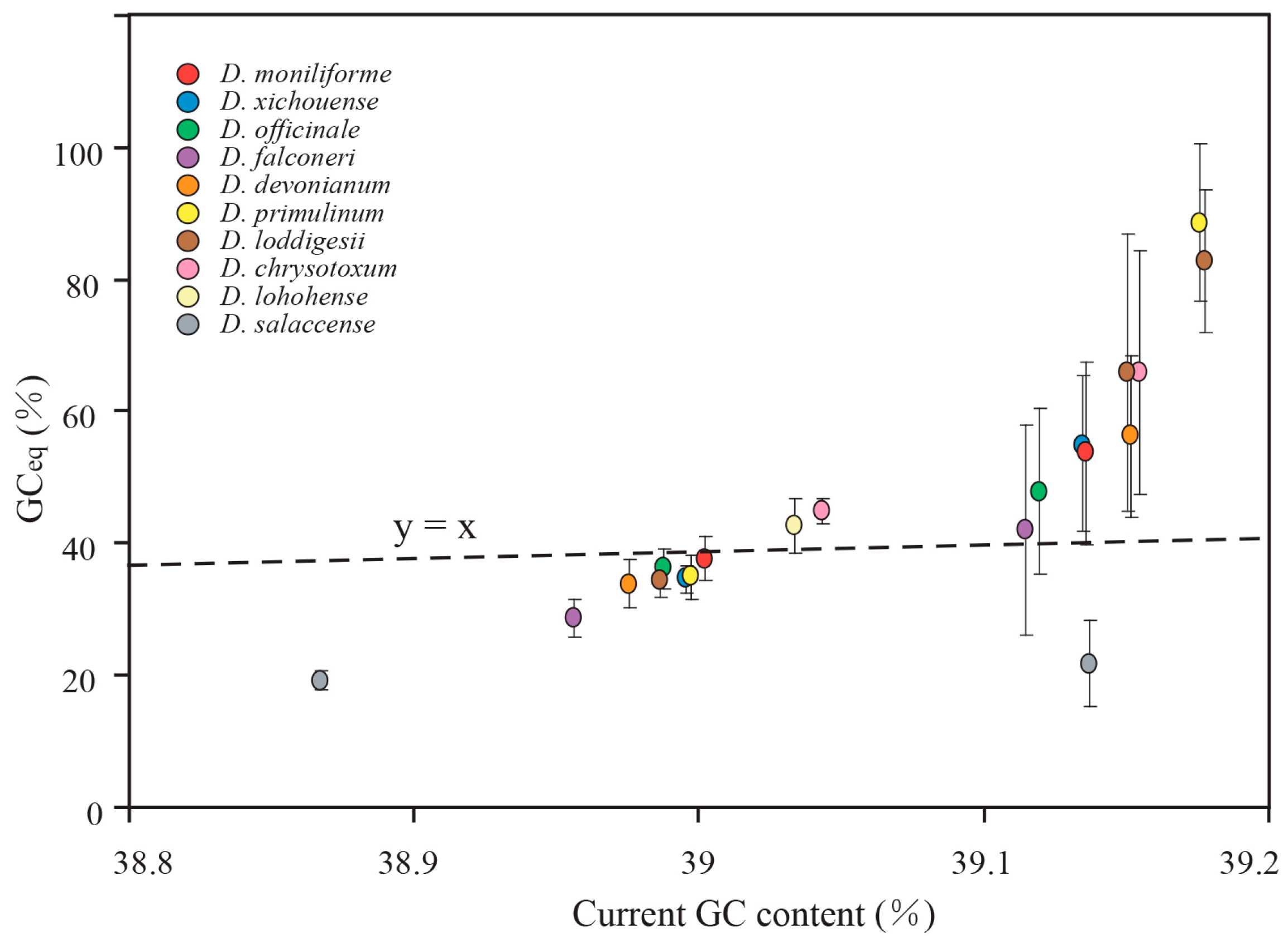
2.5. Mutational Biases Have Directly Impact on the Evolution of GC Content in Dendrobium Plastomes
2.6. gBGC Has an Effect on Maintaining Higher GC Content in IR Regions
3. Discussion
3.1. The High-Quality Complete Plastome Sequences That Could Represent for Dendrobium Species
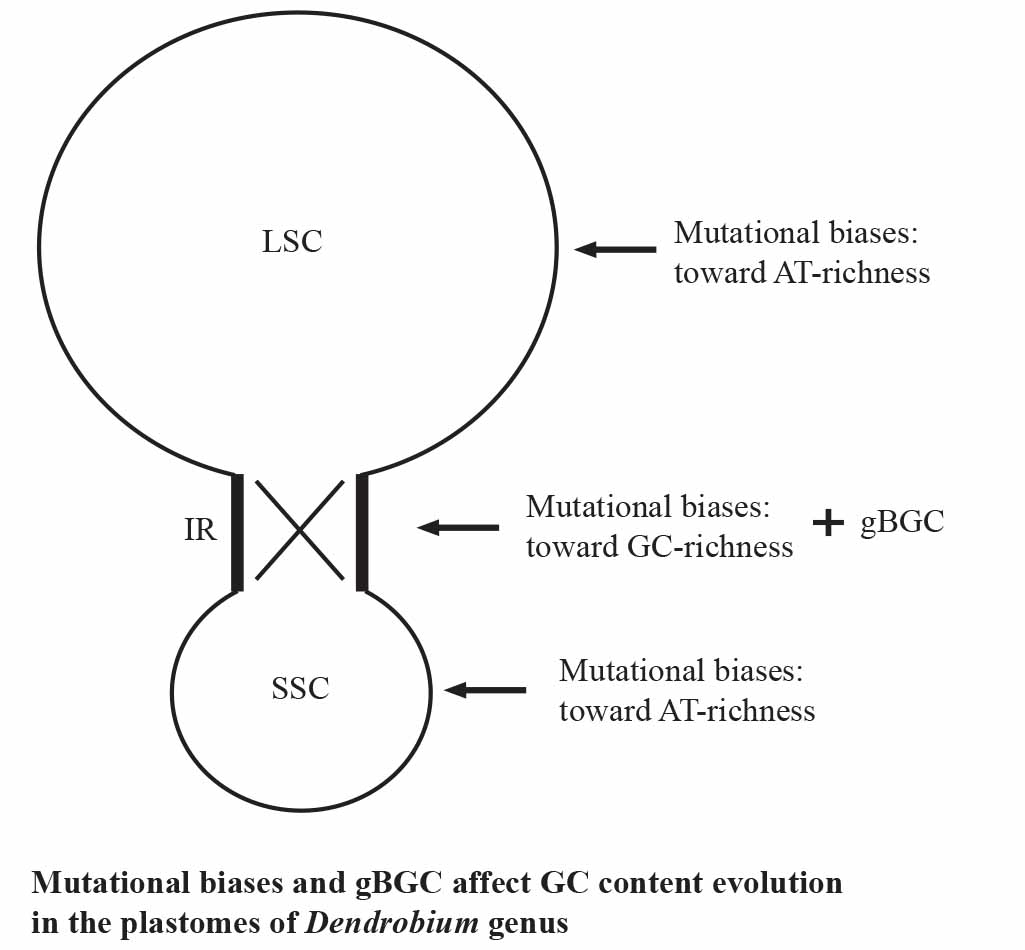
3.2. Both Mutational Biases and gBGC Affect GC Content in the Plastomes of Dendrobium Genus
3.2.1. Natural Selection
3.2.2. Mutational Bias
3.2.3. gBGC
4. Materials and Methods
4.1. DNA Extraction and Plastome Sequencing
4.2. Plastome Assembly, Annotation and Comparison
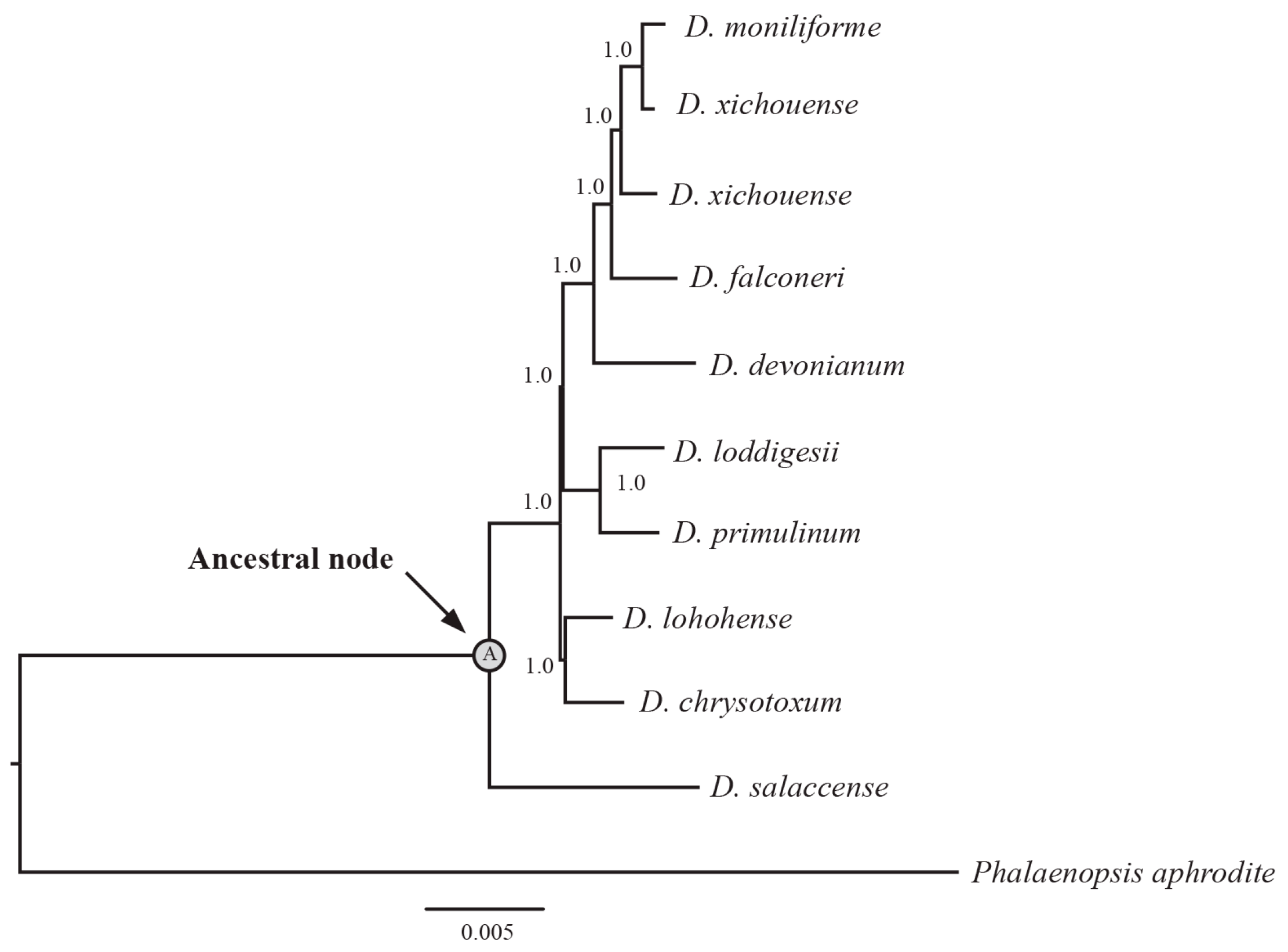
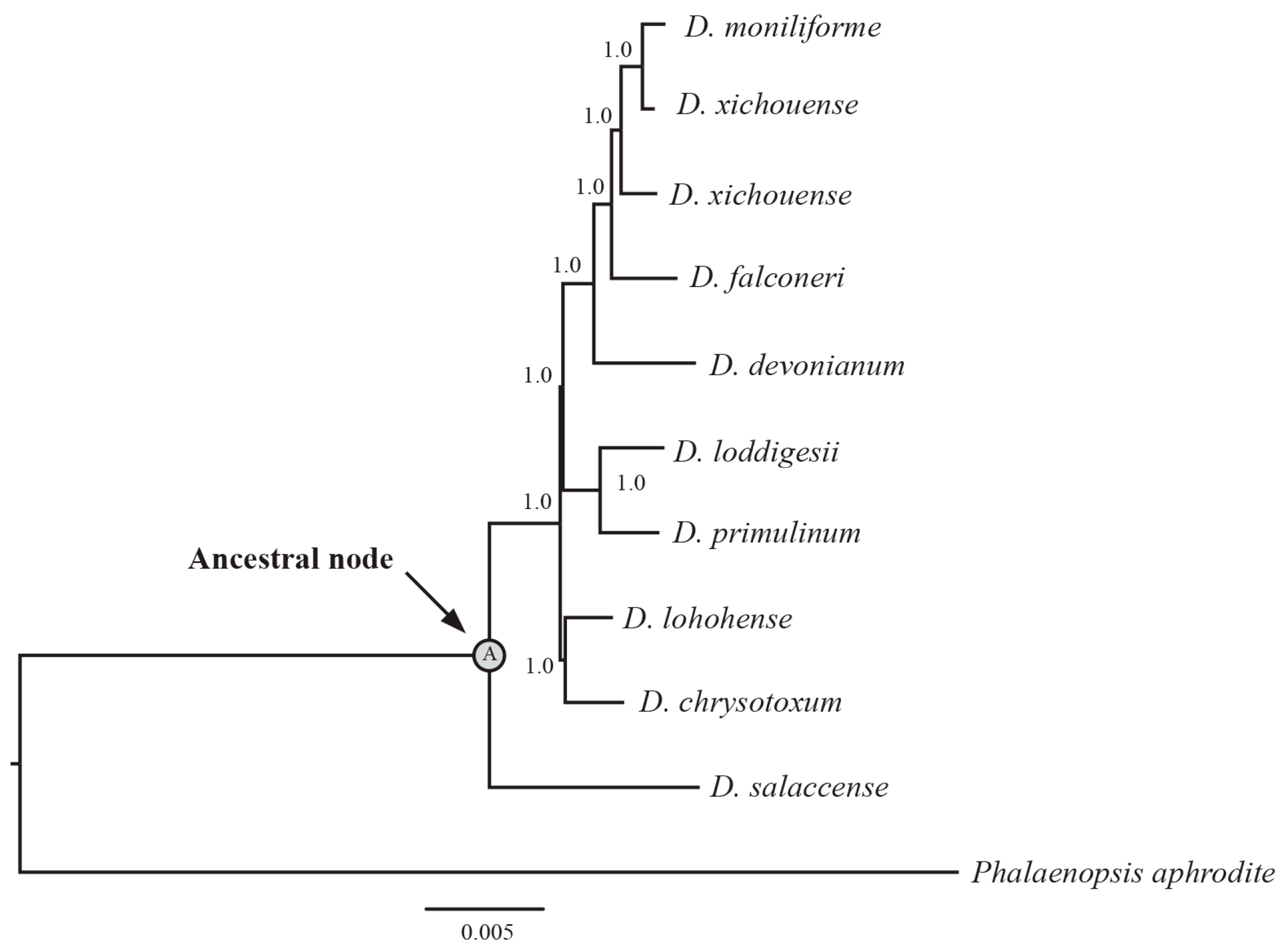
4.3. Phylogenetic Analysis
4.4. Ancestral Non-Coding Sequences Reconstruction and Calculation of the Relative Rates of the Six Nucleotide Pair Mutations
4.5. Estimation of GCeq
4.6. Natural Selection Analysis and Evolutionary Stasis of Protein-Coding Genes
4.7. Estimation of Substitution Rates
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raubeson, L.A.; Jansen, R.K. Chloroplast genomes of plants. In Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants; Henry, R.J., Ed.; CAB International: London, UK, 2005; pp. 45–68. [Google Scholar]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Lai, Y.T.; Lin, C.P.; Wang, Y.N.; Chaw, S.M. Evolution of reduced and compact chloroplast genomes (cpDNAs) in gnetophytes: Selection toward a lower-cost strategy. Mol. Phylogenet. Evol. 2009, 52, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Eyre-Walker, A.; Hurst, L.D. The evolution of isochores. Nat. Rev. Genet. 2001, 2, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Basak, S.; Ghosh, T.C. Nature of selective constraints on synonymous codon usage of rice differs in GC-poor and GC-rich genes. Gene 2007, 400, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, G. Isochores and the evolutionary genomics of vertebrates. Gene 2000, 241, 3–17. [Google Scholar] [CrossRef]
- Wang, H.C.; Singer, G.A.C.; Hickey, D.A. Mutational bias affects protein evolution in flowering plants. Mol. Biol. Evol. 2004, 21, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Günther, T.; Lampei, C.; Schmid, K.J. Mutational bias and gene conversion affect the intraspecific nitrogen stoichiometry of the Arabidopsis thaliana transcriptome. Mol. Biol. Evol. 2013, 30, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Fryxell, K.J.; Zuckerkandl, E. Cytosine deamination plays a primary role in the evolution of mammalian isochores. Mol. Biol. Evol. 2000, 17, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- Marais, G. Biased gene conversion: Implications for genome and sex evolution. Trends Genet. 2003, 19, 330–338. [Google Scholar] [CrossRef]
- Duret, L.; Galtier, N. Biased gene conversion and the evolution of mammalian genomic landscapes. Annu. Rev. Genom. Hum. Genet. 2009, 10, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Lee, R.W. Mitochondrial genome of the colorless green alga Polytomella capuana: A linear molecule with an unprecedented GC content. Mol. Biol. Evol. 2008, 25, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Pessia, E.; Popa, A.; Mousset, S.; Rezvoy, C.; Duret, L.; Marais, G.A.B. Evidence for widespread GC-biased gene conversion in eukaryotes. Genome Biol. Evol. 2012, 4, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, F.; Périan, S.; Bataillon, T.; Nesme, X.; Duret, L.; Daubin, V. GC-content evolution in bacterial genomes: The biased gene conversion hypothesis expands. PLoS Genet. 2015, 11, e1004941. [Google Scholar] [CrossRef] [PubMed]
- Lesecque, Y.; Mouchiroud, D.; Duret, L. GC-biased gene conversion in yeast is specifically associated with crossovers: Molecular mechanisms and evolutionary significance. Mol. Biol. Evol. 2013, 30, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Muyle, A.; Serres-Giardi, L.; Ressayre, A.; Escobar, J.; Glémin, S. GC-biased gene conversion and selection affect GC content in the oryza genus (rice). Mol. Biol. Evol. 2011, 28, 2695. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Chaw, S.M. Evolutionary stasis in cycad plastomes and the first case of plastome GC-biased gene conversion. Genome Biol. Evol. 2015, 7, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.D.; Clements, M.A.; Arroyo, M.T.K.; Leebens-Mack, J.; et al. Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proc. Biol. Sci. B 2015, 282, 2108–2111. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Shafer, H.L.; Leonard, O.R.; Kovach, M.J.; Schorr, M.; Morris, A.B. Chloroplast DNA sequence utility for the lowest phylogenetic and phylogeographic inferences in angiosperms: The tortoise and the hare IV. Am. J. Bot. 2014, 101, 1987–2004. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Xue, Q.; Zhu, S.; Sun, J.; Liu, W.; Ding, X. The complete plastome sequences of four orchid species: Insights into the evolution of the Orchidaceae and the utility of plastomic mutational hotspots. Front. Plant. Sci. 2017, 8, 715. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Hou, B.W.; Niu, Z.T.; Liu, W.; Xue, Q.Y.; Ding, X.Y. Comparative chloroplast genomes of photosynthetic orchids: Insights into evolution of the Orchidaceae and development of molecular markers for phylogenetic applications. PLoS ONE 2014, 9, e99016. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Zhu, S.; Pan, J.; Li, L.; Sun, J.; Ding, X. Comparative analysis of Dendrobium plastomes and utility of plastomic mutational hotspots. Sci. Rep. 2017, 7, 2073. [Google Scholar]
- Wood, H.P. The Dendrobiums; Timber Press: Portland, OR, USA, 2006. [Google Scholar]
- Feng, S.; Jiang, Y.; Wang, S.; Jiang, M.; Chen, Z.; Ying, Q.; Wang, H. Molecular identification of Dendrobium species (Orchidaceae) based on the DNA barcode ITS2 region and its application for phylogenetic study. Int. J. Mol. Sci. 2014, 16, 21975–21988. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Pan, J.; Zhu, S.; Li, L.; Xue, Q.; Liu, W.; Ding, X. Comparative analysis of the complete plastomes of Apostasia wallichii and Neuwiedia singapureana (Apostasioideae) reveals different evolutionary dynamics of IR/SSC boundary among photosynthetic orchids. Front. Plant. Sci. 2017, 8, 1713. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Chaw, S.M. Large-scale comparative analysis reveals the mechanisms driving plastomic compaction, reduction, and inversions in conifers II (cupressophytes). Genome Biol. Evol. 2016, 8, 3740–3750. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Hou, B.; Zhang, J.; Min, T.; Yuan, Y.; Niu, Z.; Ding, X. Detecting adulteration of Dendrobium officinale by real-time PCR coupled with ARMS. Int. J. Food Sci. Technol. 2012, 47, 1695–1700. [Google Scholar] [CrossRef]
- Yan, W.J.; Zhang, J.Z.; Zheng, R.; Sun, Y.L.; Ren, J.; Ding, X.Y. Combination of SYBR Green II and TaqMan Probe in the adulteration detection of Dendrobium devonianum by fluorescent quantitative PCR. Int. J. Food Sci. Technol. 2016, 50, 2572–2578. [Google Scholar] [CrossRef]
- Straub, S.C.; Fishbein, M.; Livshultz, T.; Foster, Z.; Parks, M.; Weitemier, K.; Cronn, R.C.; Liston, A. Building a model: Developing genomic resources for common milkweed (Asclepias syriaca) with low coverage genome sequencing. BMC Genom. 2011, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, W.P.; Clark, L.G.; Kelchner, S.A.; Burke, S.V.; Pires, J.C.; Edger, P.P.; Mayfield, D.R.; Triplett, J.K.; Columbus, J.T.; Ingram, A.L.; et al. A multi-step comparison of short-read full plastome sequence assembly methods in grasses. Taxon 2014, 63, 899–910. [Google Scholar]
- Kim, K.; Lee, S.C.; Lee, J.; Yu, Y.; Yang, T.J.; Choi, B.S.; Koh, H.; Waminal, N.E.; Choi, H.; Kim, N.; et al. Complete chloroplast and ribosomal sequences for 30 accessions elucidate evolution of Oryza AA genome species. Sci. Rep. 2015, 5, 15655. [Google Scholar] [CrossRef] [PubMed]
- Morton, B.R. Chloroplast DNA codon use: Evidence for selection at the psbA locus based on tRNA availability. J. Mol. Evol. 1993, 3, 273–280. [Google Scholar] [CrossRef]
- Tatarinova, T.V.; Alexandrov, N.N.; Bouck, J.B.; Feldmann, K.A. GC3 biology in corn, rice, sorghum and other grasses. BMC Genom. 2010, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Tatarinova, T.; Elhaik, E.; Pellegrini, M. Cross-species analysis of genic GC3 content and DNA methylation patterns. Genome Biol. Evol. 2013, 5, 1443–1456. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, X.; Li, Z.; Zhu, Q.; Yang, J.; Ge, S. Evidence that natural selection is the primary cause of the guanine-cytosine content variation in rice genes. J. Integr. Plant Biol. 2007, 49, 1393–1399. [Google Scholar] [CrossRef]
- McCoy, S.R.; Kuehl, J.V.; Boore, J.L.; Raubeson, L.A. The complete plastid genome sequence of Welwitschia mirabilis: An unusually compact plastome with accelerated divergence rates. BMC Evol. Biol. 2008, 8, 130. [Google Scholar] [CrossRef] [PubMed]
- Glémin, S.; Clément, Y.; David, J.; Ressayre, A. GC content evolution in coding regions of angiosperm genomes: A unifying hypothesis. Trends Genet. 2014, 30, 263. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Drouin, G.; Daoud, H.; Xia, J. Relative rates of synonymous substitutions in the mitochondrial, chloroplast, and nuclear genomes of seed plants. Mol. Phylogenet. Evol. 2008, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Duret, L.; Glémin, S.; Ranwez, V. GC-biased gene conversion promotes the fixation of deleterious amino acid changes in primates. Trends Genet. 2009, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Escobar, J.S.; Glémin, S.; Galtier, N. GC-biased gene conversion impacts ribosomal DNA evolution in vertebrates, angiosperms, and other eukaryotes. Mol. Biol. Evol. 2011, 28, 2561–2575. [Google Scholar] [CrossRef] [PubMed]
- Gotea, V.; Elnitski, L. Ascertaining regions affected by GC-biased gene conversion through weak-to-strong mutational hotspots. Genomics 2014, 103, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Goubert, C.; Modolo, L.; Vieira, C.; Valientemoro, C.; Mavingui, P.; Boulesteix, M. De novo assembly and annotation of the Asian tiger mosquito (Aedes albopictus) repeatome with dnaPipeTE from raw genomic reads and comparative analysis with the yellow fever mosquito (Aedes aegypti). Genome Biol. Evol. 2015, 7, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Glémin, S.; Arndt, P.F.; Messer, P.W.; Petrov, D.; Galtier, N.; Duret, L. Quantification of GC-biased gene conversion in the human genome. Genome Res. 2015, 25, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Chloroplast DNA exists in two orientations. Nature 1983, 301, 92–93. [Google Scholar] [CrossRef]
- Khakhlova, O.; Bock, R. Elimination of deleterious mutations in plastid genomes by gene conversion. Plant J. 2006, 46, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fan, W.; Tian, G.; Zhu, H.; He, L. The sequence and de novo assembly of the giant panda genome. Nature 2010, 463, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Hershberg, R.; Petrov, D.A. Evidence that mutation is universally biased towards AT in bacteria. PLoS Genet. 2010, 6, e1001115. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package) Version 3.6; Department of Genome Sciences, University of Washington: Seattle, DC, USA, 2005. [Google Scholar]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Plastome Length (bp) | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | GC Content (%) | GC Content of LSC (%) | GC Content of SSC (%) | GC Content of IR (%) | Accession |
|---|---|---|---|---|---|---|---|---|---|
| Dendrobium loddigesii | 152,493 | 84,089 | 16,932 | 25,736 | 37.38 | 35.63 | 30.54 | 43.57 | LC317044 |
| Dendrobium devonianum | 151,715 | 84,897 | 14,311 | 25,800 | 37.56 | 35.12 | 30.61 | 43.51 | LC317045 |
| Species | SC | IR | ||
|---|---|---|---|---|
| Numbers of GC-AT Mutations | Numbers of Normalized AT-GC Mutations | Numbers of GC-AT Mutations | Numbers of Normalized AT-GC Mutations | |
| D. moniliforme | 229 | 66.65 | 6 | 7.74 |
| D. xichouense | 226 | 65.58 | 8 | 6.18 |
| D. officinale | 229 | 61.93 | 8 | 5.39 |
| D. falconeri | 248 | 76.54 | 6 | 4.63 |
| D. devonianum | 273 | 109.66 | 9 | 10.84 |
| D. primulinum | 221 | 60.78 | 8 | 7.71 |
| D. loddigesii | 210 | 62.25 | 13 | 8.62 |
| D. chrysotoxum | 188 | 61.10 | 1 | 7.74 |
| D. lohohense | 185 | 40.77 | 7 | 5.40 |
| D. salaccense | 452 | 104.40 | 14 | 6.92 |
| Species | SC | IR | ||
|---|---|---|---|---|
| Numbers of GC-AT Mutations | Numbers of Normalized AT-GC Mutations | Numbers of GC-AT Mutations | Numbers of Normalized AT-GC Mutations | |
| D. moniliforme | 79 | 44.73 | 1 | 7.73 |
| D. xichouense | 115 | 45.95 | 8 | 5.78 |
| D. officinale | 100 | 51.09 | 8 | 10.29 |
| D. falconeri | 94 | 49.20 | 2 | 9.66 |
| D. devonianum | 89 | 53.71 | 5 | 5.79 |
| D. primulinum | 94 | 49.86 | 7 | 6.43 |
| D. loddigesii | 86 | 46.03 | 5 | 5.79 |
| D. chrysotoxum | 152 | 36.24 | 13 | 1.28 |
| D. lohohense | 59 | 48.04 | 3 | 5.79 |
| D. salaccense | 57 | 42.26 | 3 | 5.79 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Z.; Xue, Q.; Wang, H.; Xie, X.; Zhu, S.; Liu, W.; Ding, X. Mutational Biases and GC-Biased Gene Conversion Affect GC Content in the Plastomes of Dendrobium Genus. Int. J. Mol. Sci. 2017, 18, 2307. https://doi.org/10.3390/ijms18112307
Niu Z, Xue Q, Wang H, Xie X, Zhu S, Liu W, Ding X. Mutational Biases and GC-Biased Gene Conversion Affect GC Content in the Plastomes of Dendrobium Genus. International Journal of Molecular Sciences. 2017; 18(11):2307. https://doi.org/10.3390/ijms18112307
Chicago/Turabian StyleNiu, Zhitao, Qingyun Xue, Hui Wang, Xuezhu Xie, Shuying Zhu, Wei Liu, and Xiaoyu Ding. 2017. "Mutational Biases and GC-Biased Gene Conversion Affect GC Content in the Plastomes of Dendrobium Genus" International Journal of Molecular Sciences 18, no. 11: 2307. https://doi.org/10.3390/ijms18112307