Estrogen Metabolism-Associated CYP2D6 and IL6-174G/C Polymorphisms in Schistosoma haematobium Infection

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
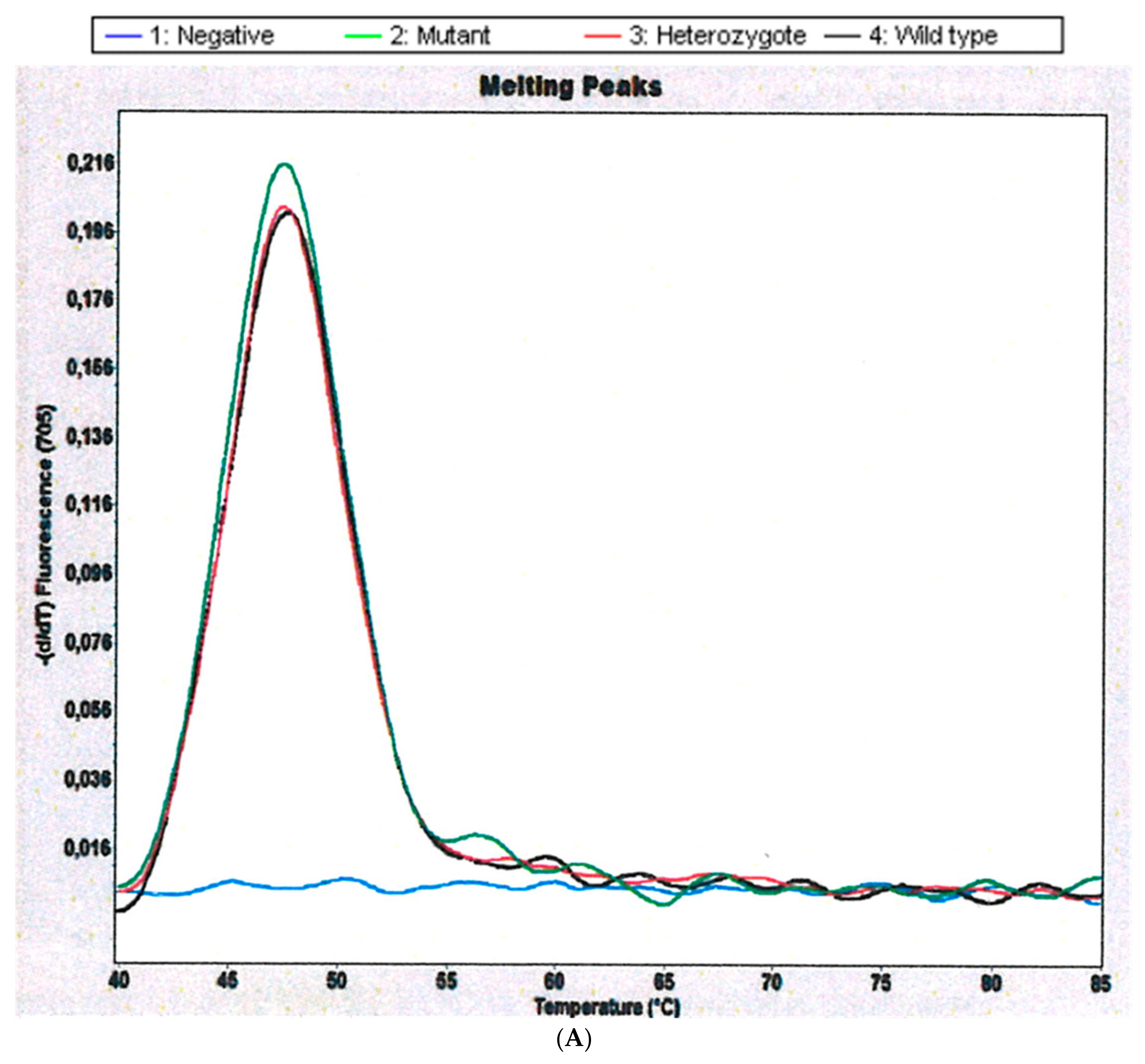
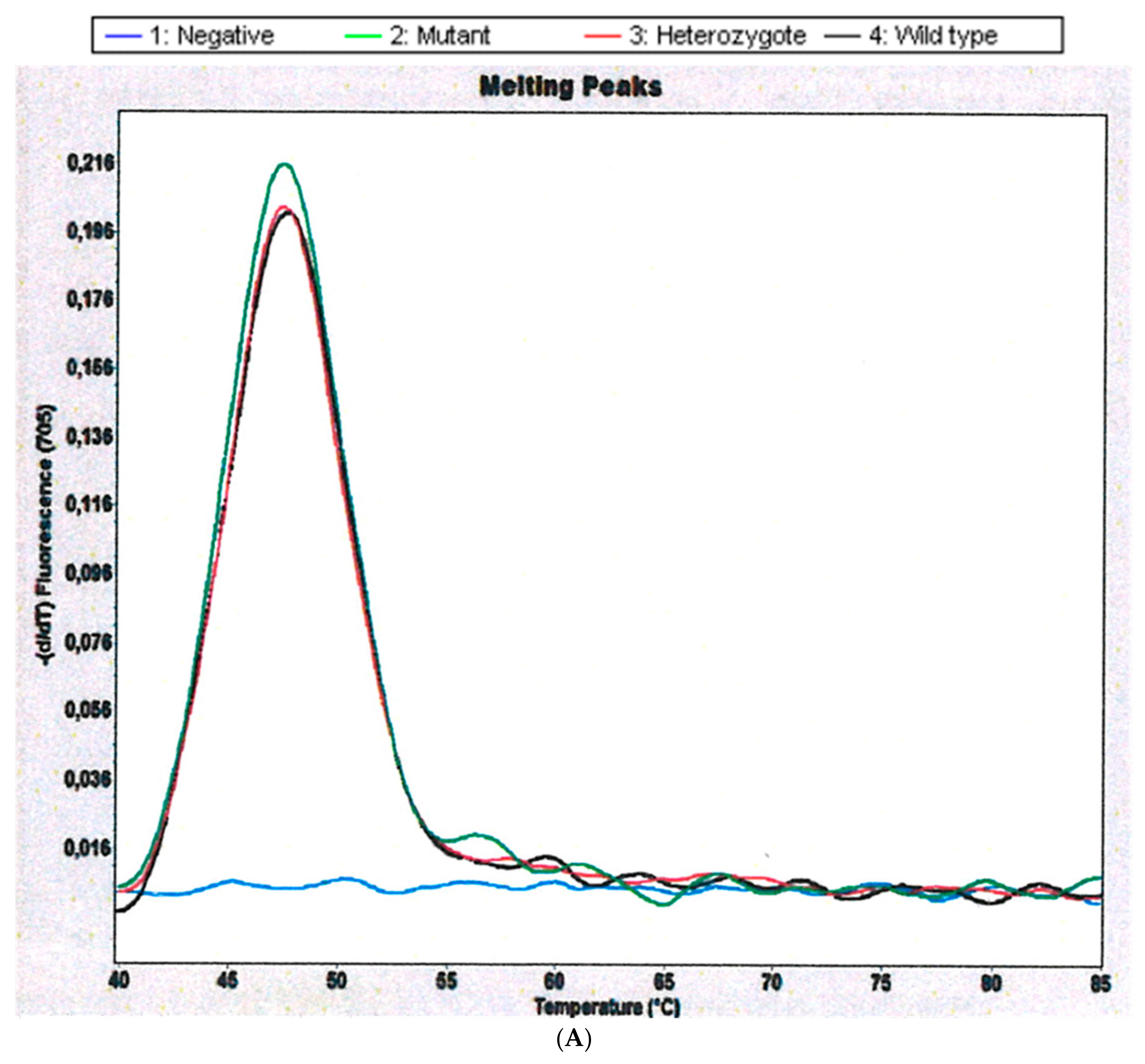
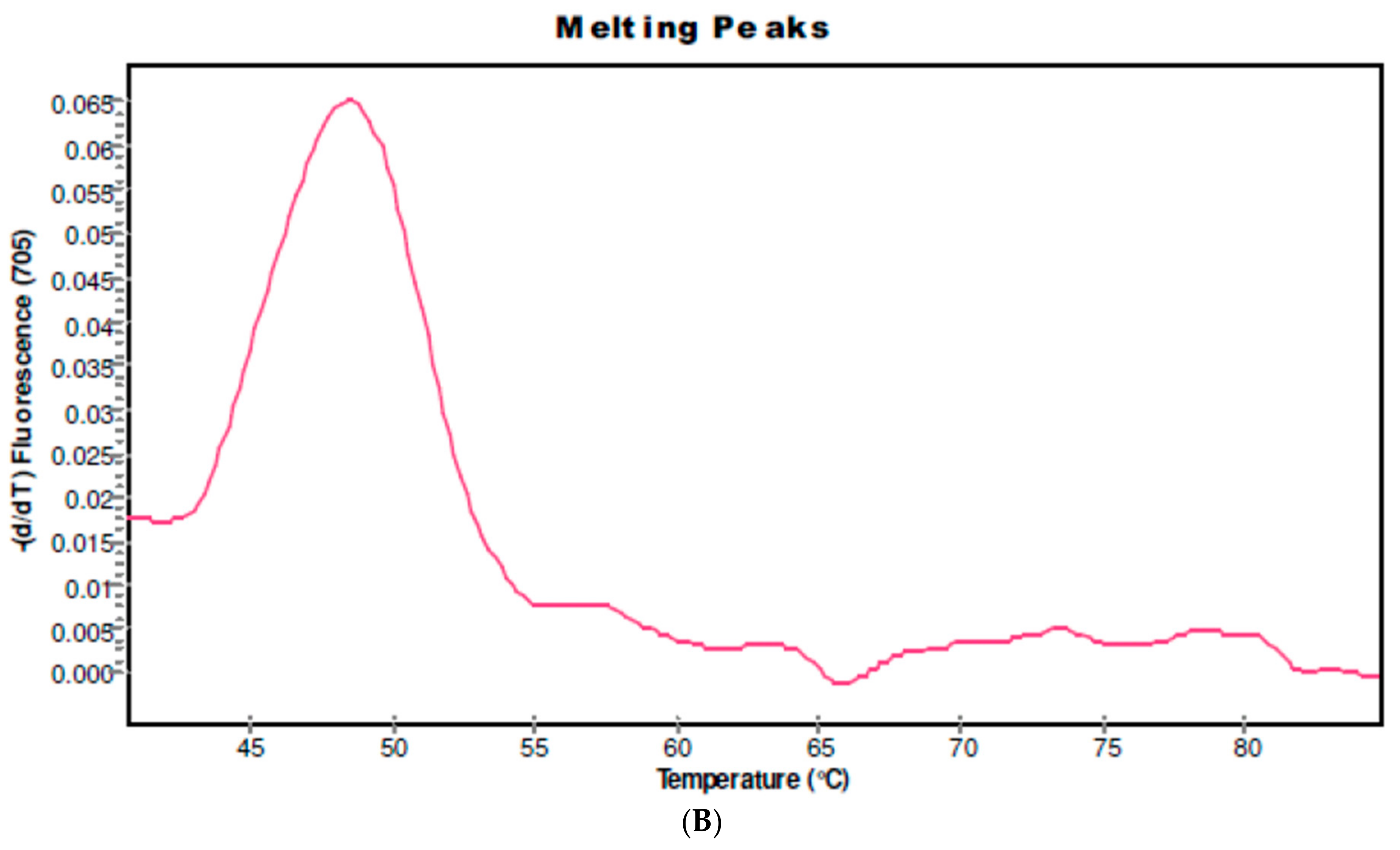
2.1. CYP 2D6 Alleles *3, *4 and *5/*5 in S. haematobium-Infected Patients
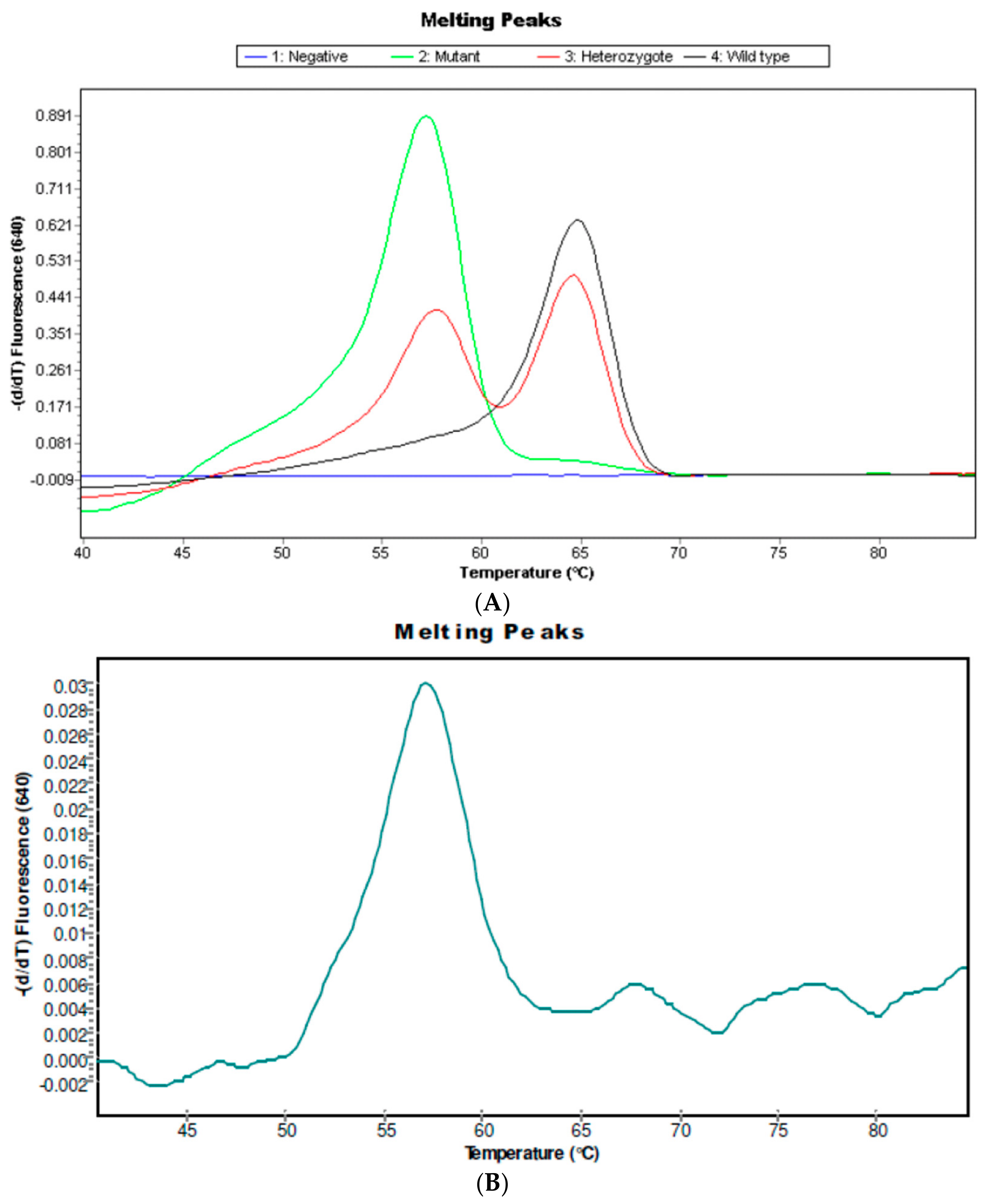
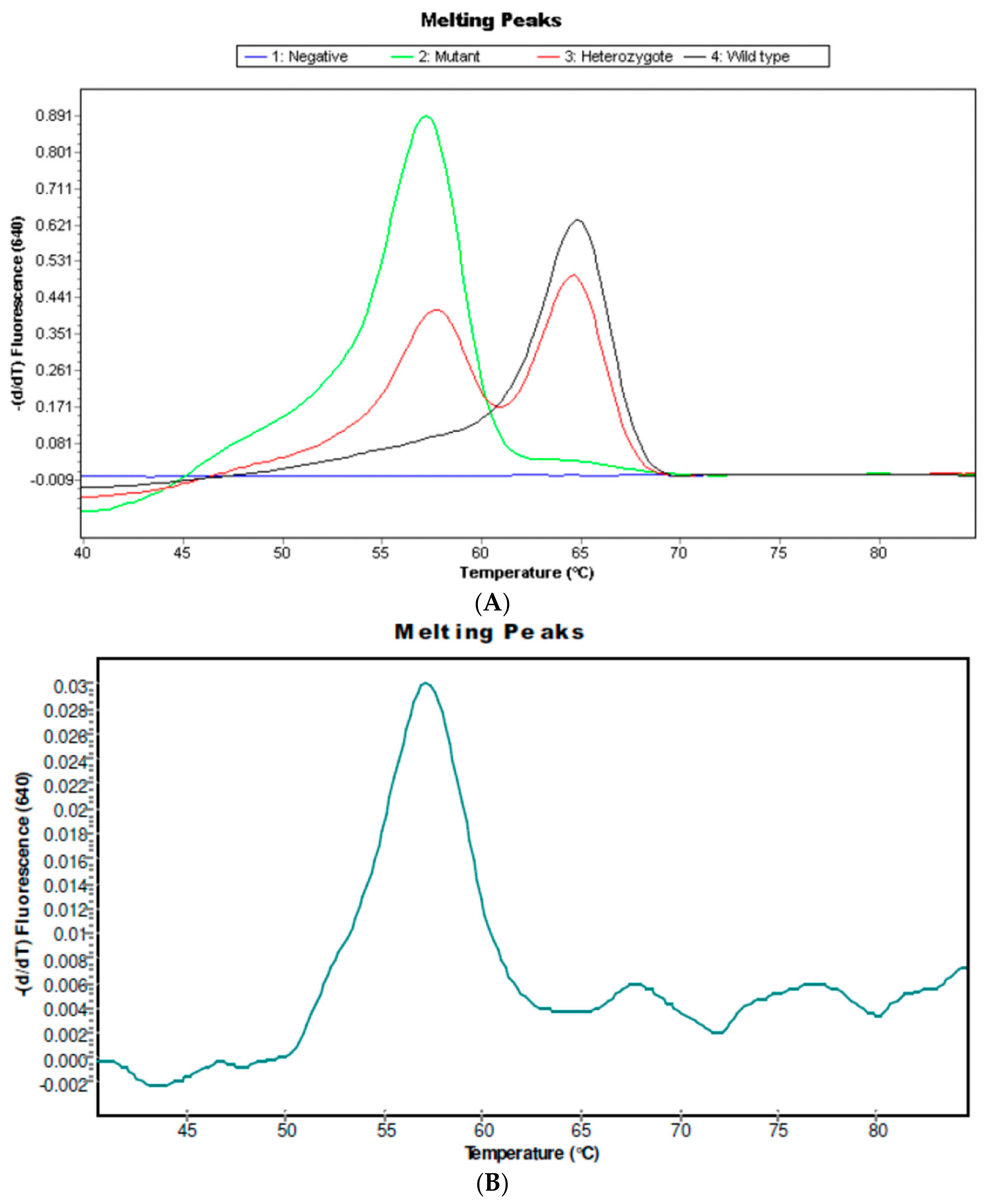
2.2. IL6-174C Variant in S. haematobium Infected Patients
3. Discussion
4. Material and Methods
4.1. Study Area, Population and Design
4.2. Urine Collection
4.3. Urine Analysis
4.4. Anthropometric Measurements
4.5. DNA Collection and Extraction
4.6. Genetic Analysis
4.6.1. Detection of CYP 2d6 Alleles *3, *4 and *5/*5
4.6.2. Detection of IL6 G-174C
4.6.3. PCR Experiment Protocol
4.7. Statistical Analysis
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
Abbreviations
| SCC | Squamous Cell Carcinoma |
| CYP | Cytochrome P450 |
| IL-6 | Interleukin 6 |
| BMI | Body Mass Index |
| WT | Wild-Type |
| PM | Poor Metabolizer |
References
- Botelho, M.C.; Alves, H.; Barros, A.; Rinaldi, G.; Brindley, P.J.; Sousa, M. The role of estrogens and estrogen receptor signaling pathways in cancer and infertility: The case of schistosomes. Trends Parasitol. 2015, 31, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Machado, J.C.; Brindley, P.J.; Correia da Costa, J.M. Targeting molecular signaling pathways of Schistosoma haemotobium infection in bladder cancer. Virulence 2011, 2, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Koslowski, N.; Sombetzki, M.; Loebermann, M.; Engelmann, R.; Grabow, N.; Österreicher, C.H.; Trauner, M.; Mueller-Hilke, B.; Reisinger, E.C. Single-sex infection with female Schistosoma mansoni cercariae mitigates hepatic fibrosis after secondary infection. PLoS Negl. Trop. Dis. 2017, 11, e0005595. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Soares, R.; Vale, N.; Ribeiro, R.; Camilo, V.; Almeida, R.; Medeiros, R.; Gomes, P.; Machado, J.C.; Correia da Costa, J.M. Schistosoma haematobium: Identification of new estrogenic molecules with estradiol antagonistic activity and ability to inactivate estrogen receptor in mammalian cells. Exp. Parasitol. 2010, 126, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Vale, N.; Gouveia, M.J.; Rinaldi, G.; Santos, J.; Santos, L.L.; Gomes, P.; Brindley, P.J.; Correia da Costa, J.M. Tumour-like phenotypes in urothelial cells after exposure to antigens from eggs of Schistosoma haematobium: An oestrogen-DNA adducts mediated pathway? Int. J. Parasitol. 2013, 43, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Gouveia, M.J.; Vale, N.; Delgado Mde, L.; Gonçalves, A.; da Silva, J.M.; Oliveira, C.; Xavier, P.; Gomes, P.; Santos, L.L.; et al. Urinary estrogen metabolites and self-reported infertility in women infected with Schistosoma haematobium. PLoS ONE 2014, 9, e96774. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Blackburn, H.L.; Ellsworth, D.L.; Shriver, C.D.; Ellsworth, R.E. Role of cytochrome P450 genes in breast cancer etiology and treatment: Effects on estrogen biosynthesis, metabolism, and response to endocrine therapy. Cancer Causes Control 2015, 26, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Terry, C.F.; Loukaci, V.; Green, F.R. Cooperative influence of genetic polymorphisms on interleukin 6 transcriptional regulation. J. Biol. Chem. 2000, 275, 18138–18144. [Google Scholar] [CrossRef] [PubMed]
- Fishman, D.; Faulds, G.; Jeffery, R.; Mohamed-Ali, V.; Yudkin, J.S.; Humphries, S.; Woo, P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J. Clin. Investig. 1998, 102, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Hulkkonen, J.; Pertovaara, M.; Antonen, J.; Pasternack, A.; Hurme, M. Elevated interleukin-6 plasma levels are regulated by the promoter region polymorphism of the IL6 gene in primary Sjogren’s syndrome and correlate with the clinical manifestations of the disease. Rheumatology 2001, 40, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Gaedigk, A.; Blum, M.; Gaedigk, R.; Eichelbaum, M.; Meyer, U.A. Deletion of the entire cytochrome P450 CYP2D6 gene as a cause of impaired drug metabolism in poor metabolizers of the debrisoquine/sparteine polymorphism. Am. J. Hum. Genet. 1991, 48, 943–950. [Google Scholar] [PubMed]
- Steen, V.M.; Molven, A.; Aarskog, N.K.; Gulbrandsen, A.K. Homologous unequal cross-over involving a 2.8 kb direct repeat as a mechanism for the generation of allelic variants of human cytochrome P450 CYP2D6 gene. Hum. Mol. Genet. 1995, 4, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Machado, A.; Carvalho, A.; Vilaça, M.; Conceição, O.; Rosa, F.; Alves, H.; Richter, J.; Bordalo, A.A. Schistosoma haematobium in Guinea-Bissau: Unacknowledged morbidity due to a particularly neglected parasite in a particularly neglected country. Parasitol. Res. 2016, 115, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Ribeiro, R.; Vale, N.; Oliveira, P.; Medeiros, R.; Lopes, C.; Machado, J.C.; Correia da Costa, J.M. Inactivation of estrogen receptor by Schistosoma haematobium total antigen in bladder urothelial cells. Oncol. Rep. 2012, 27, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Fussell, K.C.; Udasin, R.G.; Smith, P.J.; Gallo, M.A.; Laskin, J.D. Catechol metabolites of endogenous estrogens induce redox cycling and generate reactive oxygen species in breast epithelial cells. Carcinogenesis 2011, 32, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, E.L.; Rogan, E.G. Depurinating estrogen-DNA adducts, generators of cancer initiation: Their minimization leads to cancer prevention. Clin. Transl. Med. 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Alves, H.; Richter, J. Estrogen catechols detection as biomarkers in schistosomiasis induced cancer and infertility. Lett. Drug Des. Discov. 2017, 14, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Prins, J.R.; Gomez-Lopez, N.; Robertson, S.A. Interleukin-6 in pregnancy and gestational disorders. J. Reprod. Immunol. 2012, 95, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Sousa, M. New biomarkers to fight urogenital schistosomiasis: A major neglected tropical disease. Biomark. Med. 2014, 8, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Physical Status: The Use and Interpretation of Anthropometry; Technical Report Series 854; WHO Expert Committee: Geneva, Switzerland, 1995; 452p. [Google Scholar]
- Vogelstein, B.; Gillespie, D. Preparative and analytical purification of DNA from agarose. Proc. Natl. Acad. Sci. USA 1979, 76, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Sistonen, J.; Sajantila, A.; Lao, O.; Corander, J.; Barbujani, G.; Fuselli, S. CYP2D6 worldwide genetic variation shows high frequency of altered activity variants and no continental structure. Pharmacogenet. Genom. 2007, 17, 93–101. [Google Scholar]
- Sawczenko, A.; Azooz, O.; Paraszczuk, J.; Idestrom, M.; Croft, N.M.; Savage, M.O.; Ballinger, A.B.; Sanderson, I.R. Intestinal inflammation-induced growth retardation acts through IL-6 in rats and depends on the -174 IL-6 G/C polymorphism in children. Proc. Natl. Acad. Sci. USA 2005, 102, 13260–13265. [Google Scholar] [CrossRef] [PubMed]
- Weise, A.; Prause, S.; Eidens, M.; Weber, M.M.; Kann, P.H.; Forst, T.; Pfützner, A. Prevalence of CYP450 gene variations in patients with type 2 diabetes. Clin. Lab. 2010, 56, 311–318. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Population Characteristics | CYP; n = 4 (28.5%) | WT; n = 10 (71.4%) | p Value | OR | 95% CI |
|---|---|---|---|---|---|
| Age (years, median ± SD) | 10.75 | 10.8 | n.s. | ||
| Female | 1 | 4 | n.s. | 0.5242 | 0.01537, 7.015 |
| Male | 3 | 6 | n.s. | 1.908 | 0.1426, 65.05 |
| Microhaematuria (%) | 4 | 8 | n.s. | 1.599 | 0.07674, 72.45 |
| BMI (median ± SD) | 15.2 | 15.5 | n.s. |
| Population Characteristics | IL6; n = 4 (28.5%) | WT; n = 10 (71.4%) | p Value | OR | 95% CI |
|---|---|---|---|---|---|
| Age (years, median ± SD) | 6.5 ± 0.7 | 10.1 ± 3.1 | 0.005 | ||
| Female | 1 | 5 | n.s. | 1.549 | 0.03441, 69.74 |
| Male | 1 | 8 | n.s | 0.6455 | 0.01434, 29.06 |
| Microhaematuria (%) | 1 | 9 | n.s | 0.4714 | 0.01033, 21.51 |
| BMI (median ± SD) | 10.6 ± 5.9 | 14.8 ± 1.8 | 0.04 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, R.; Lacerda, P.C.; Costa, P.P.; Machado, A.; Carvalho, A.; Bordalo, A.; Fernandes, R.; Soares, R.; Richter, J.; Alves, H.; et al. Estrogen Metabolism-Associated CYP2D6 and IL6-174G/C Polymorphisms in Schistosoma haematobium Infection. Int. J. Mol. Sci. 2017, 18, 2560. https://doi.org/10.3390/ijms18122560
Cardoso R, Lacerda PC, Costa PP, Machado A, Carvalho A, Bordalo A, Fernandes R, Soares R, Richter J, Alves H, et al. Estrogen Metabolism-Associated CYP2D6 and IL6-174G/C Polymorphisms in Schistosoma haematobium Infection. International Journal of Molecular Sciences. 2017; 18(12):2560. https://doi.org/10.3390/ijms18122560
Chicago/Turabian StyleCardoso, Rita, Pedro C. Lacerda, Paulo P. Costa, Ana Machado, André Carvalho, Adriano Bordalo, Ruben Fernandes, Raquel Soares, Joachim Richter, Helena Alves, and et al. 2017. "Estrogen Metabolism-Associated CYP2D6 and IL6-174G/C Polymorphisms in Schistosoma haematobium Infection" International Journal of Molecular Sciences 18, no. 12: 2560. https://doi.org/10.3390/ijms18122560