The E3 Ubiquitin Ligase RNF7 Negatively Regulates CARD14/CARMA2sh Signaling

 ,
,
Abstract
:1. Introduction
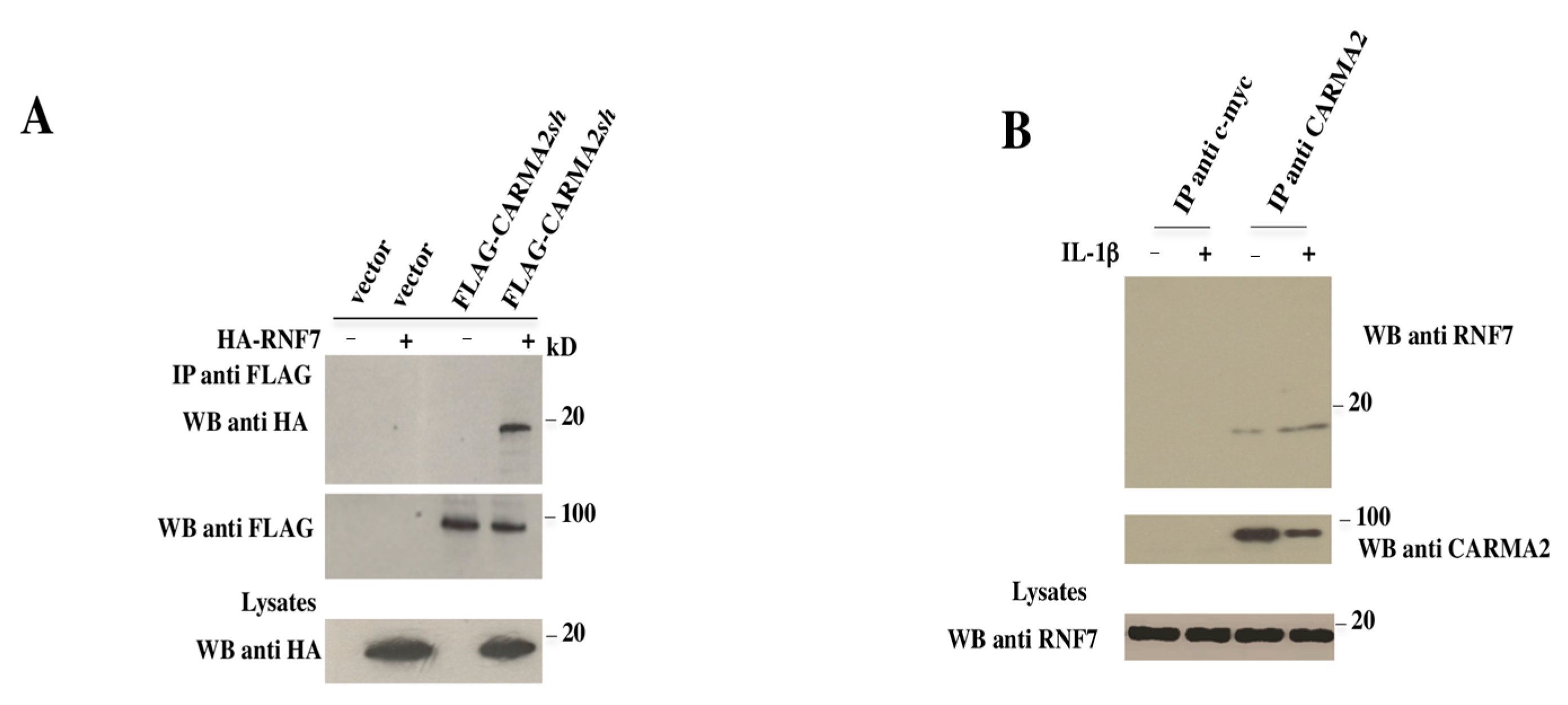
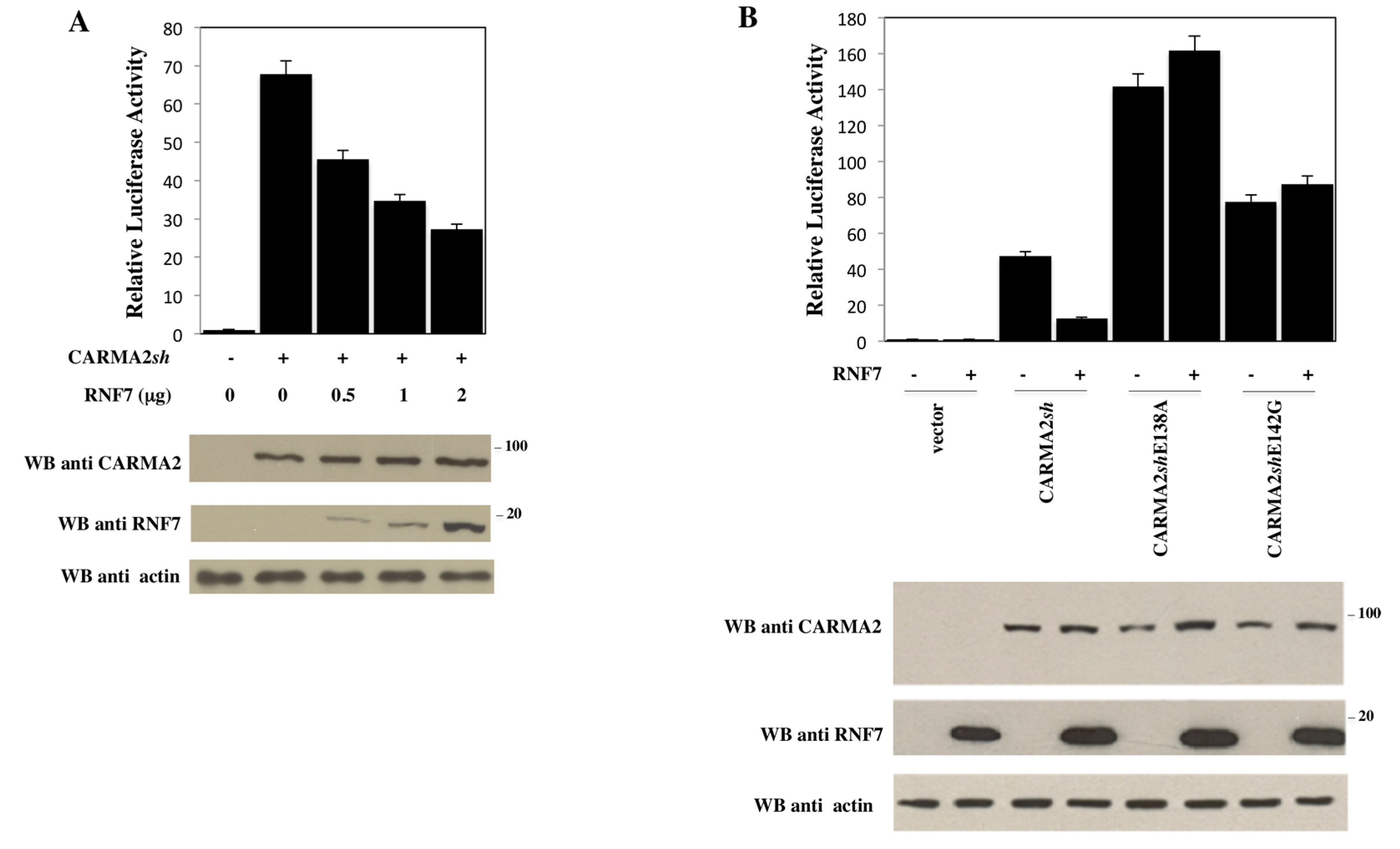
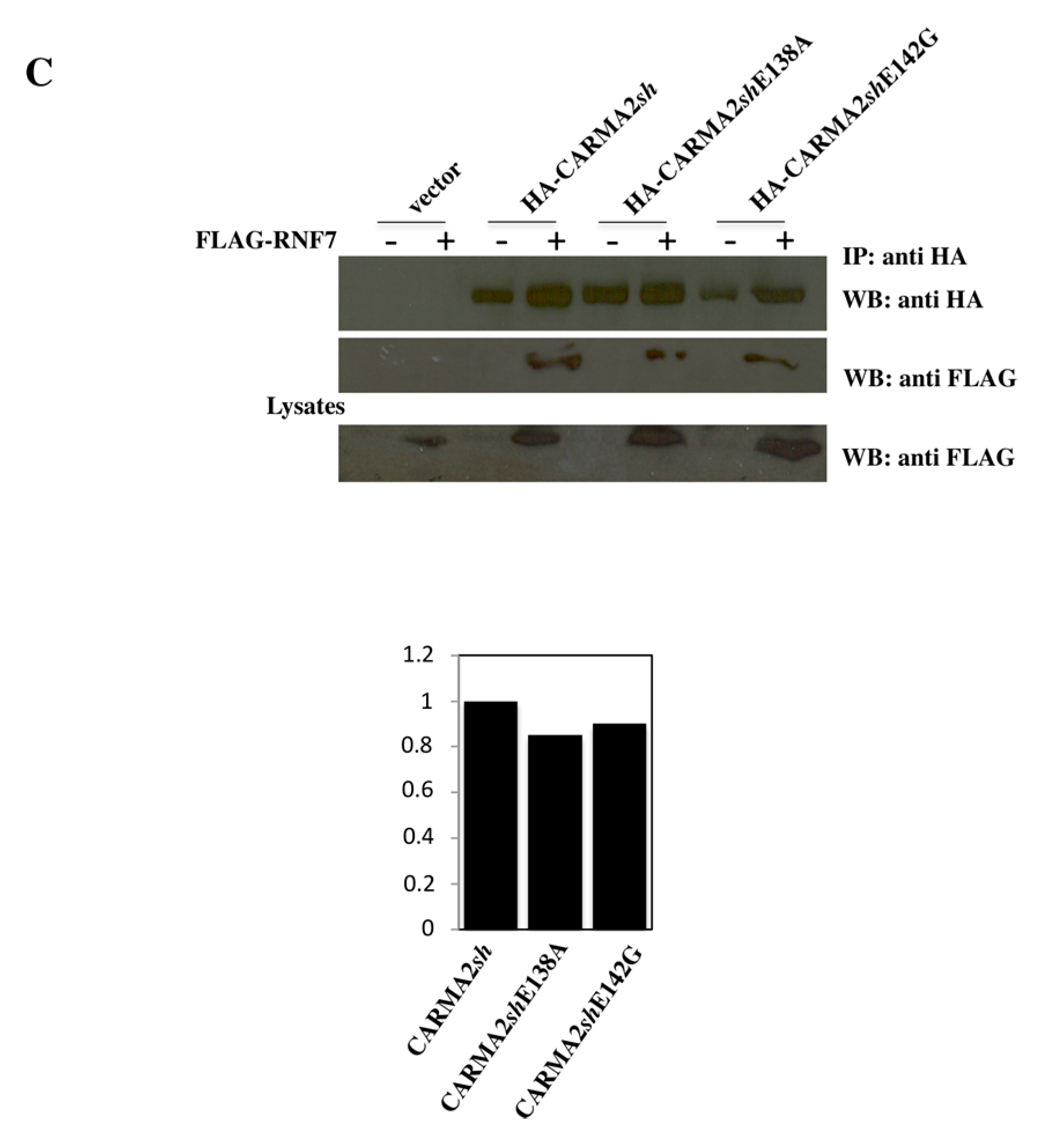
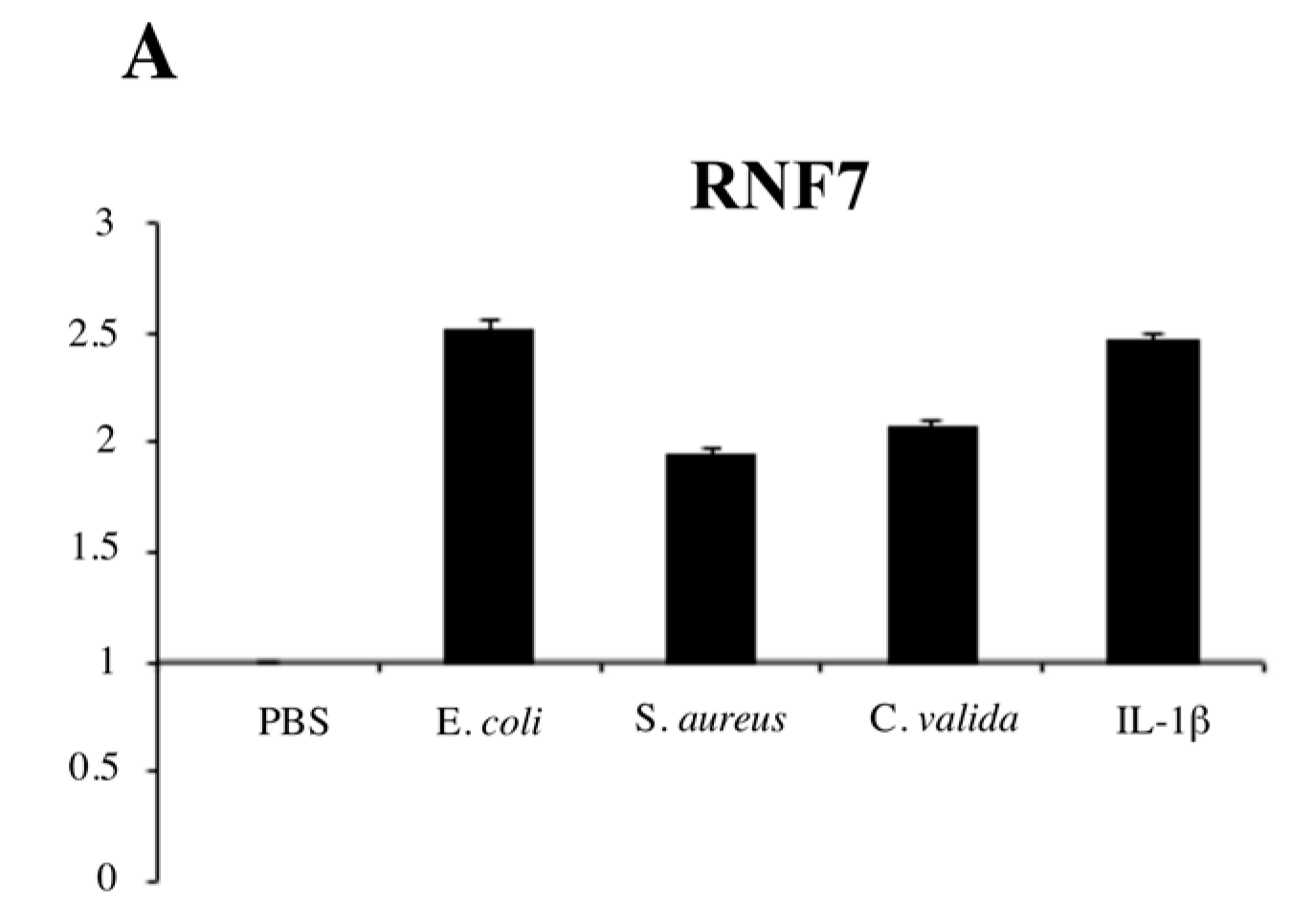
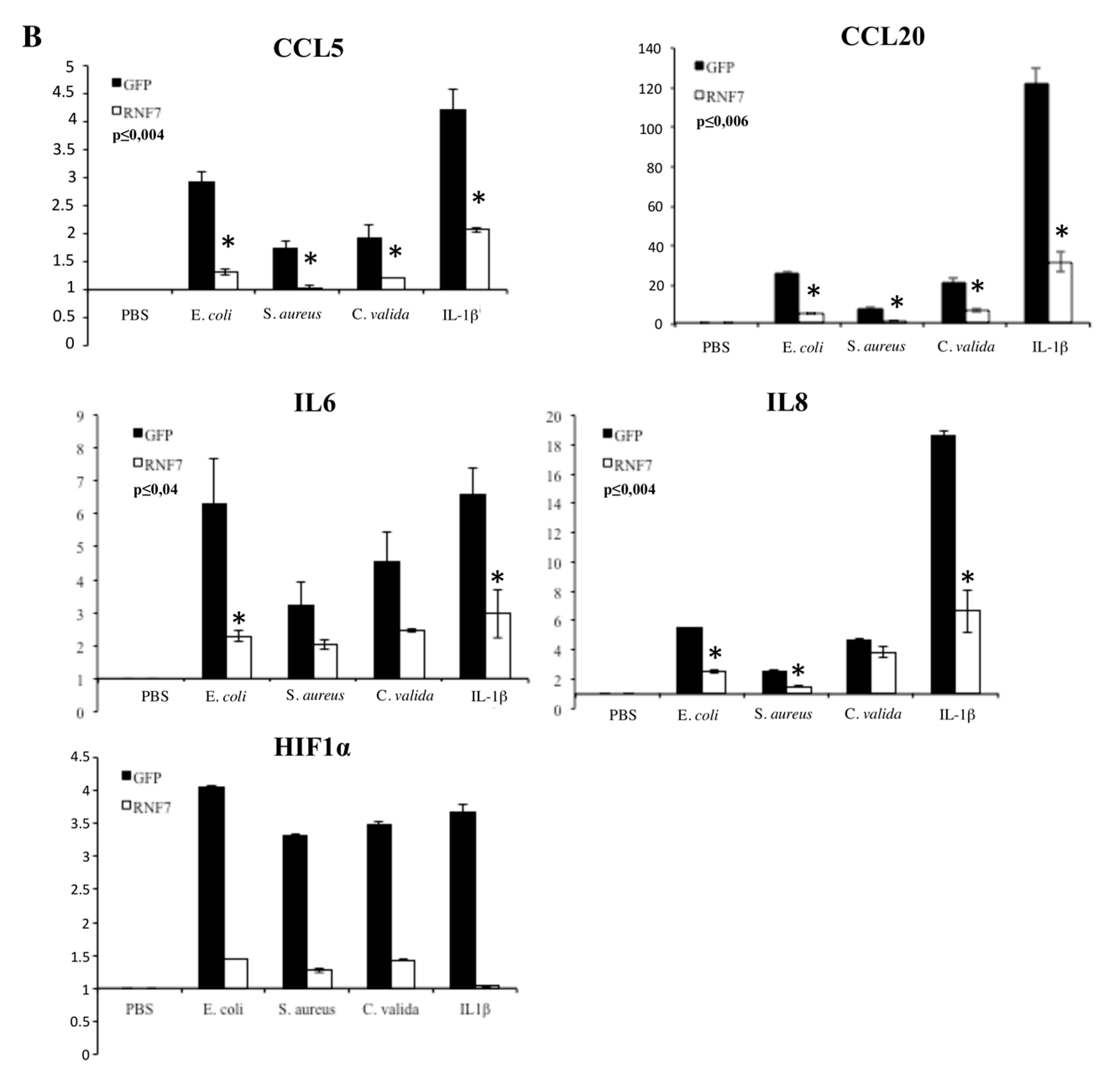
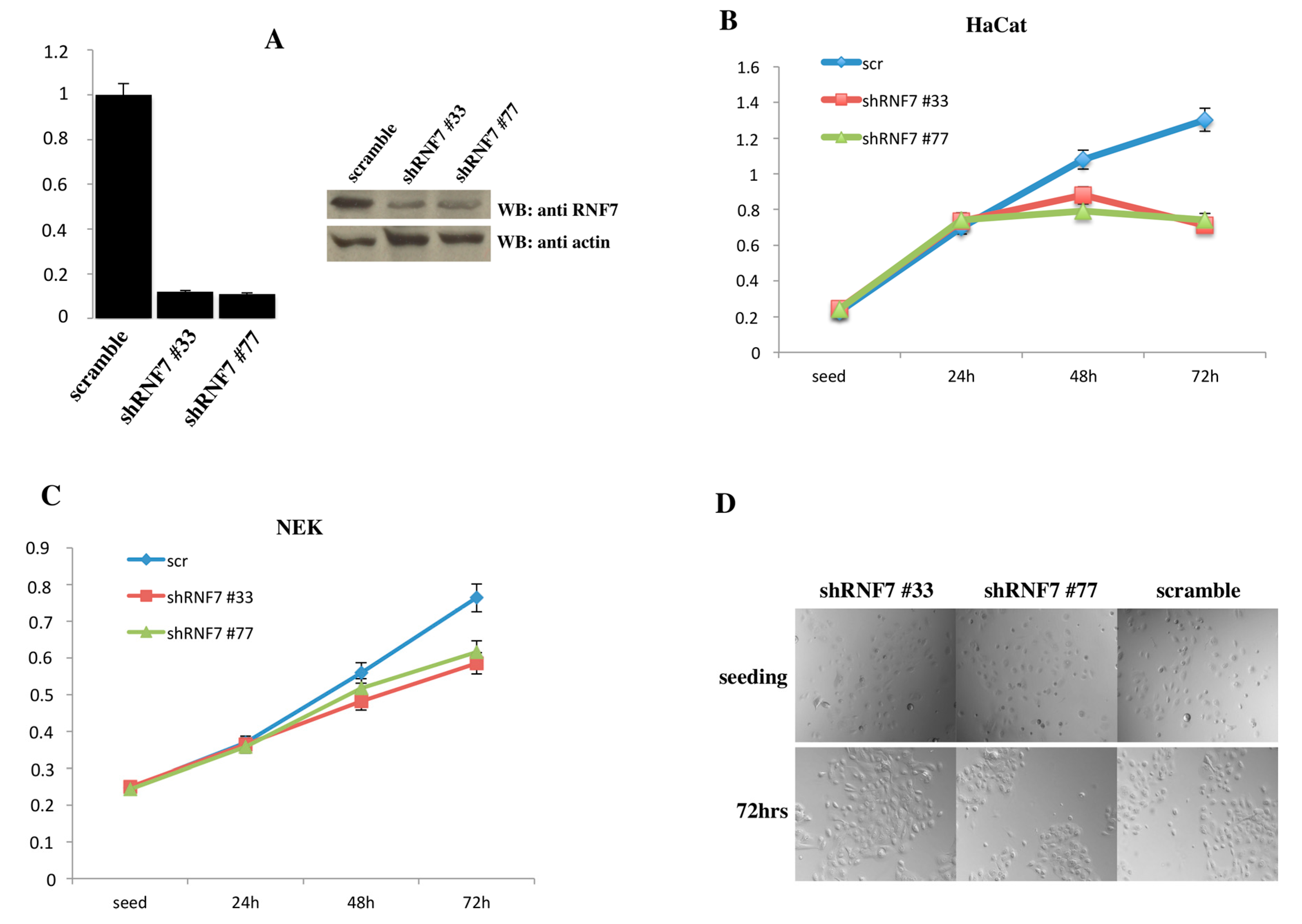
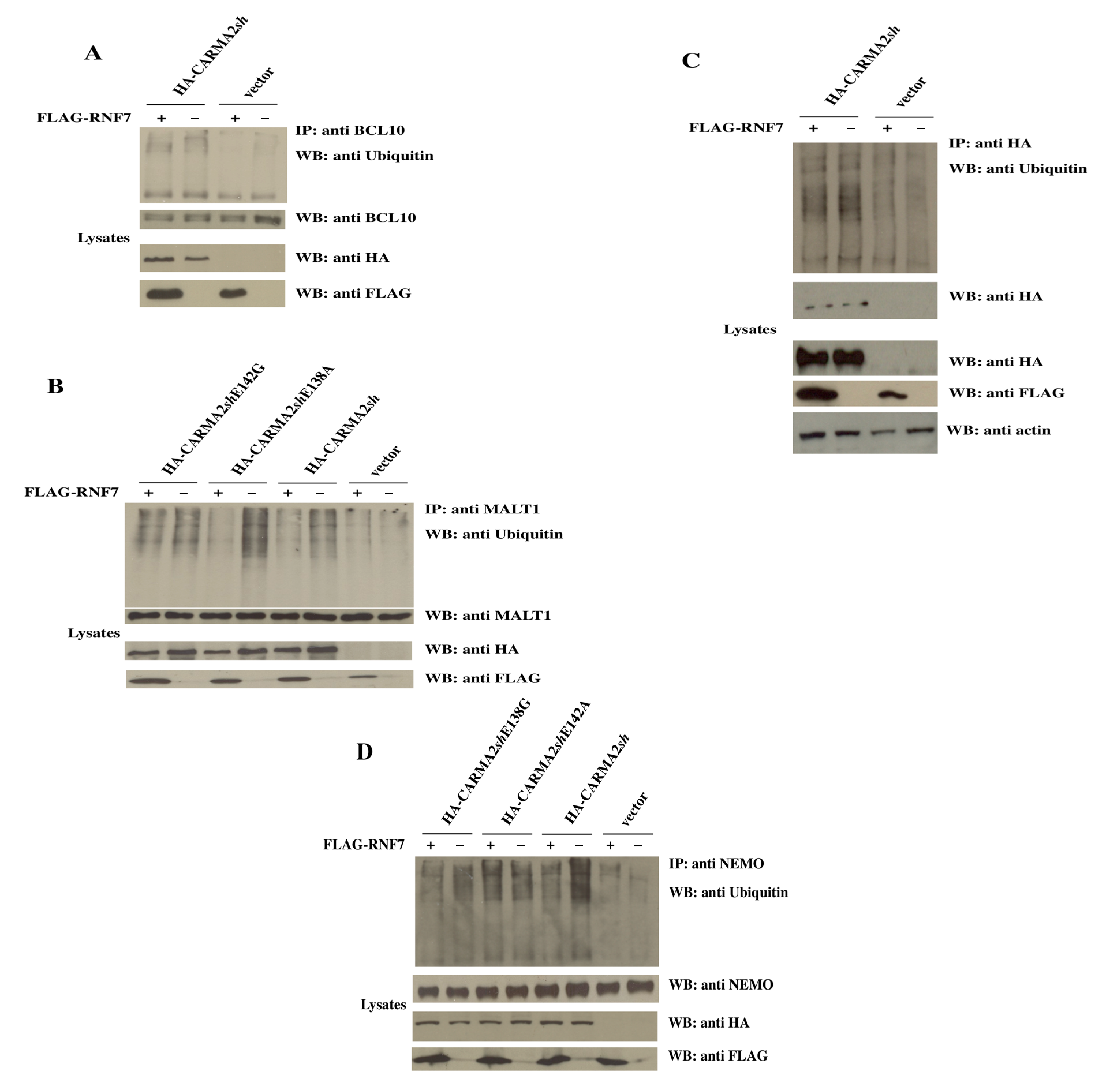
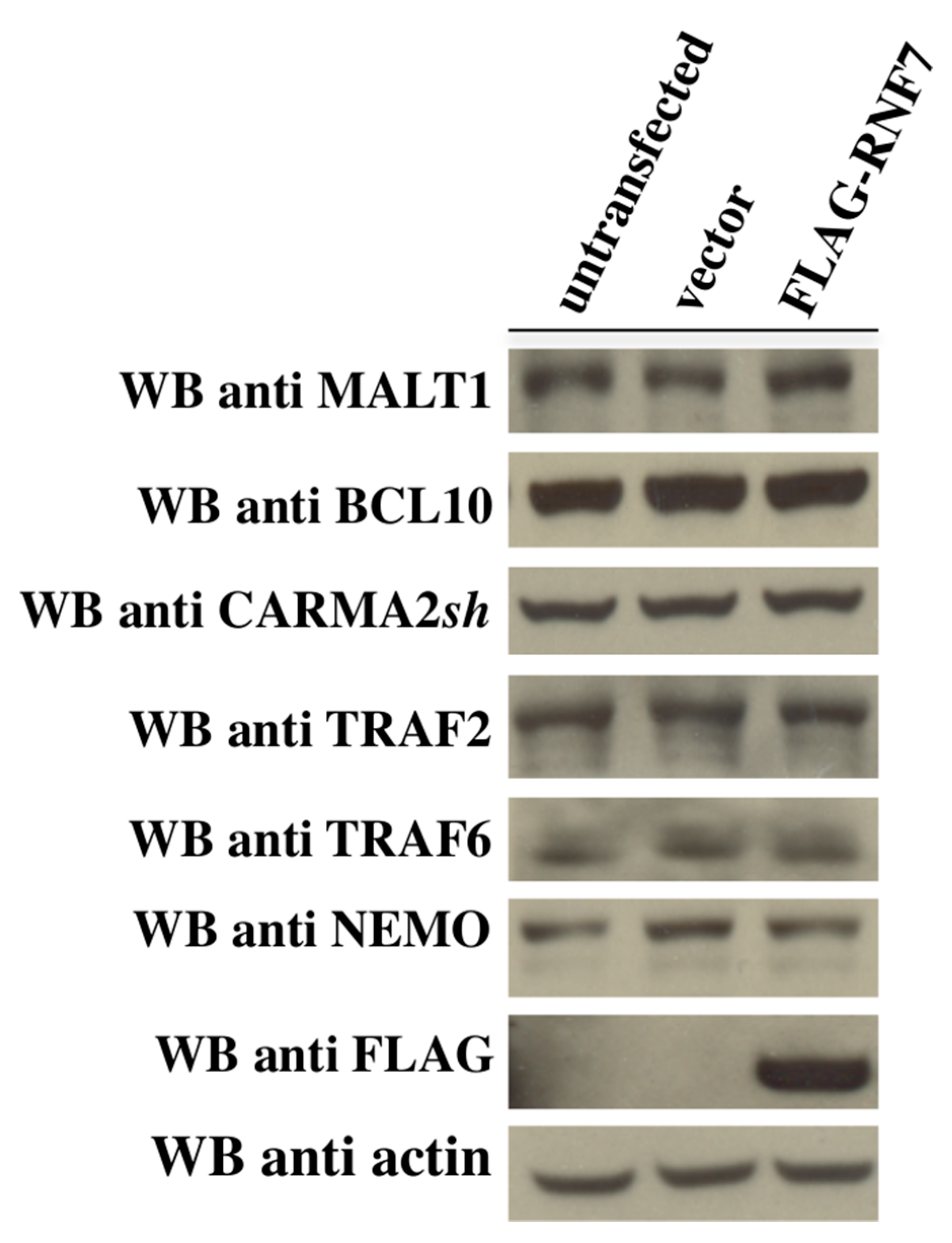
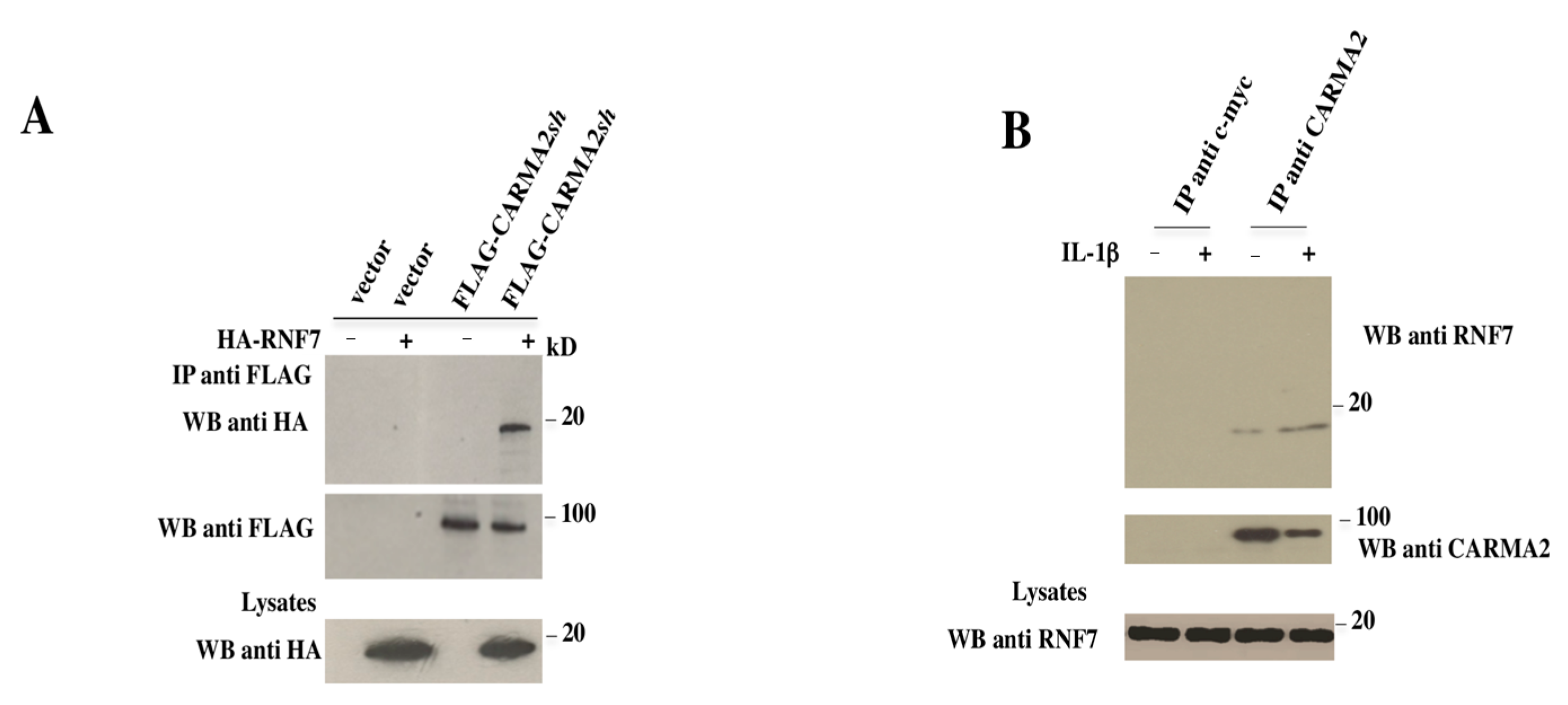
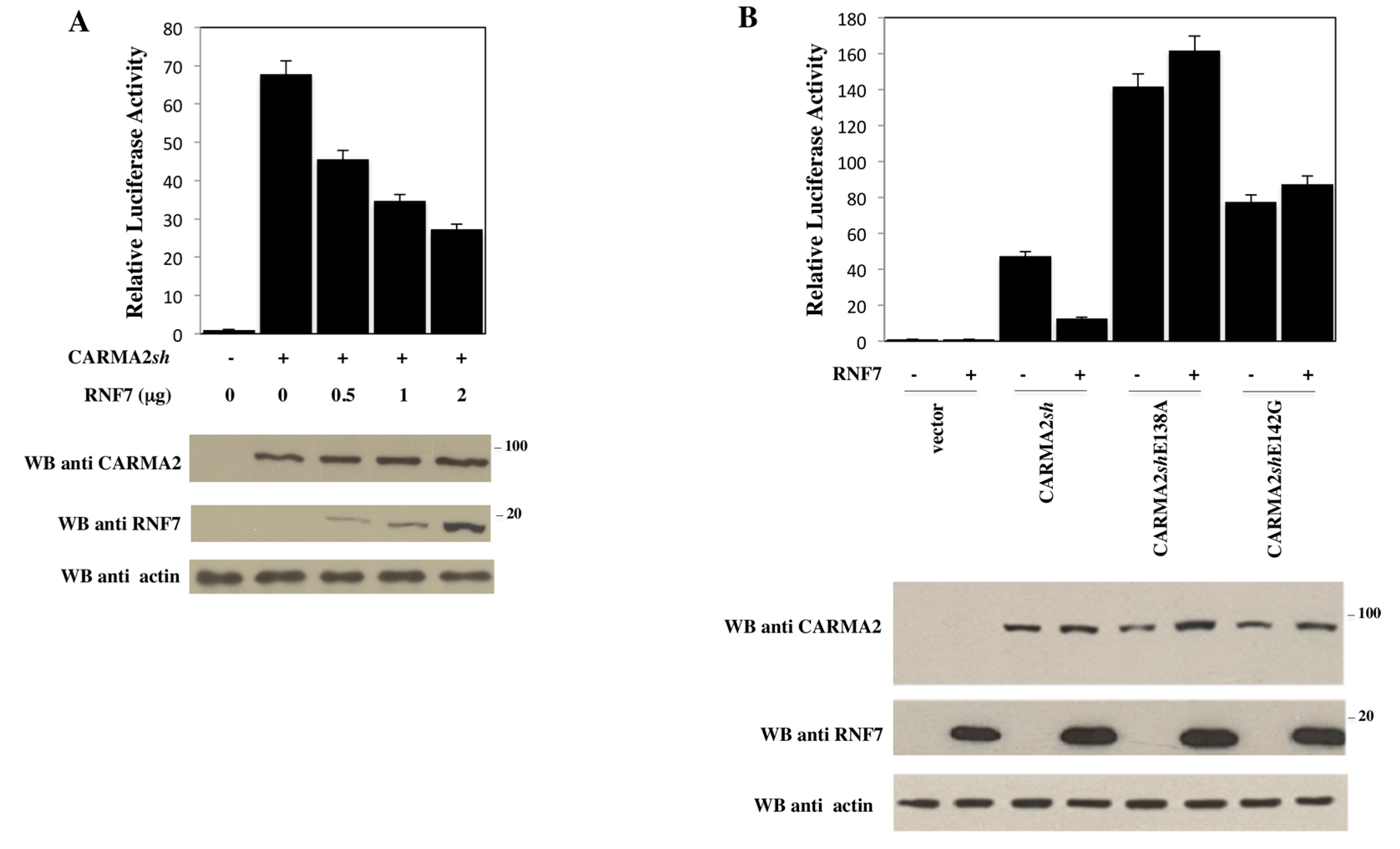
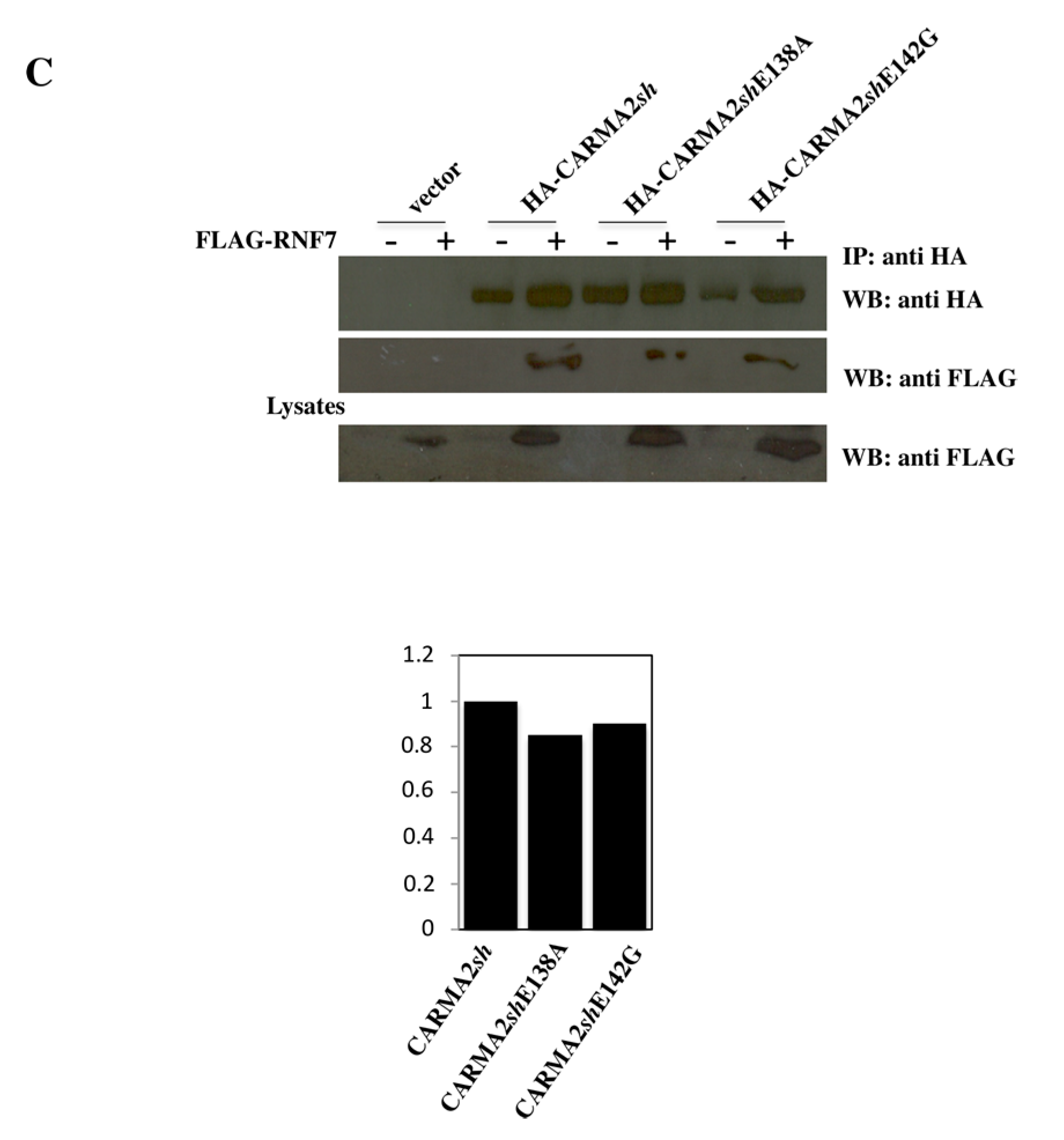
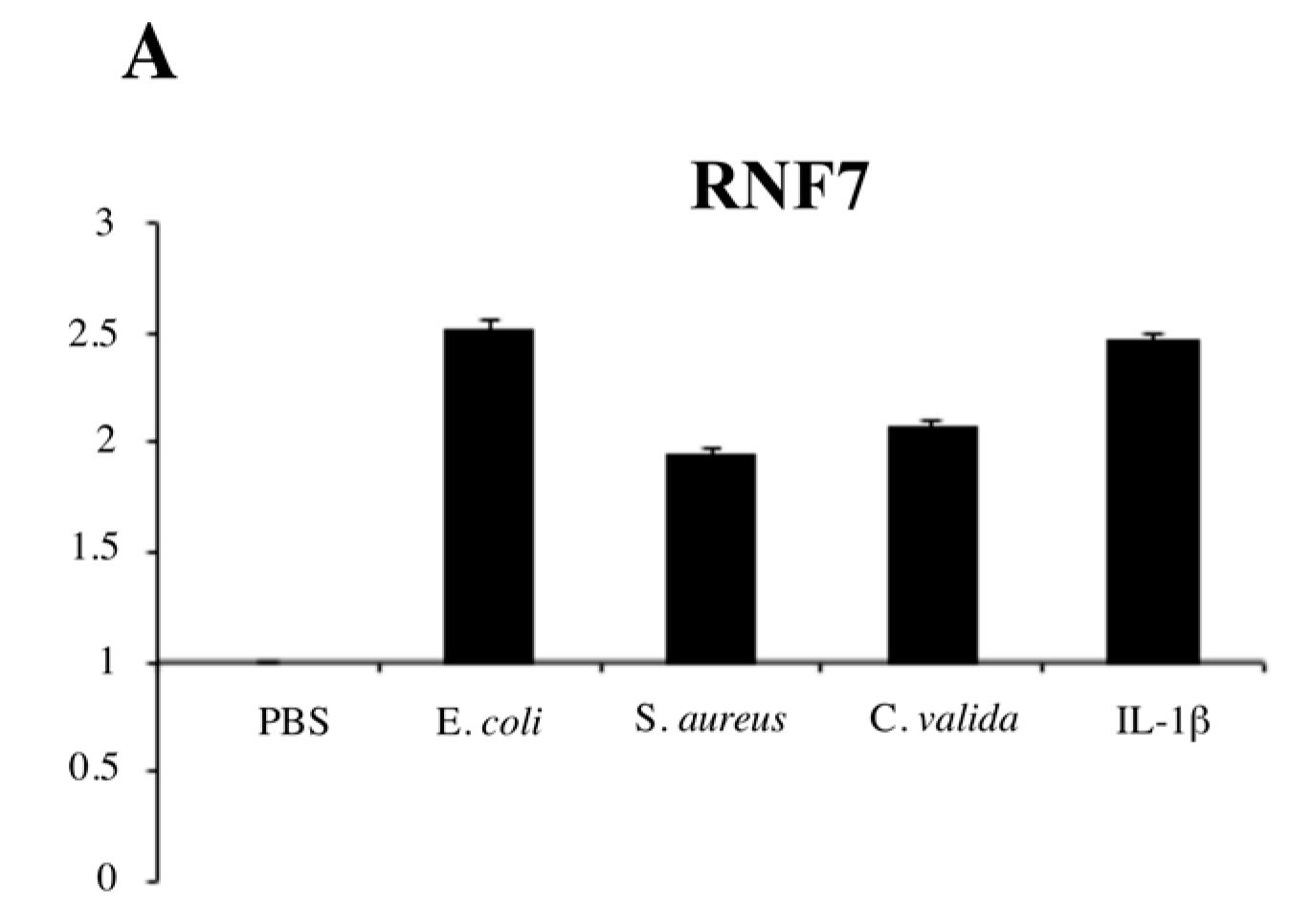
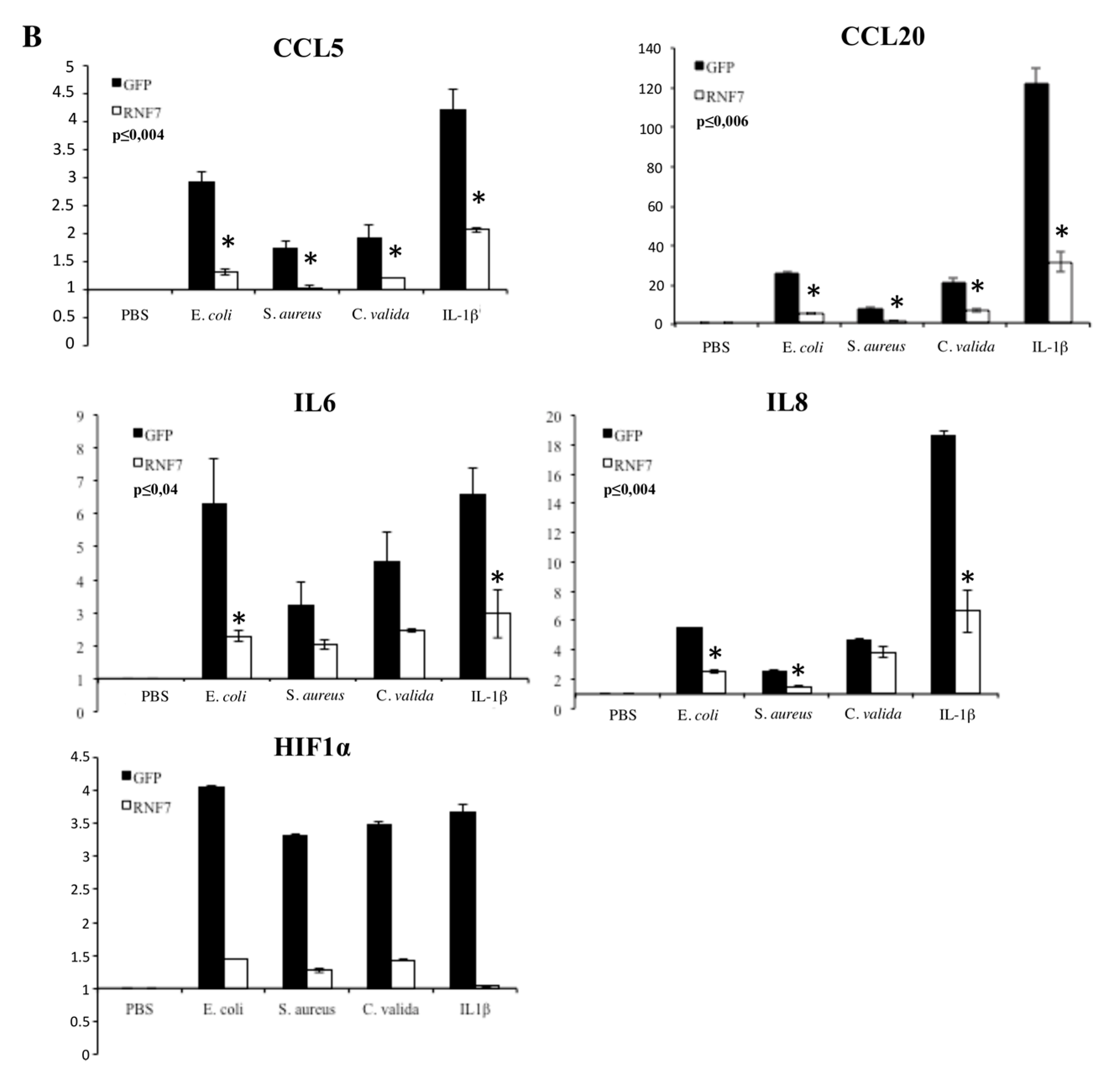
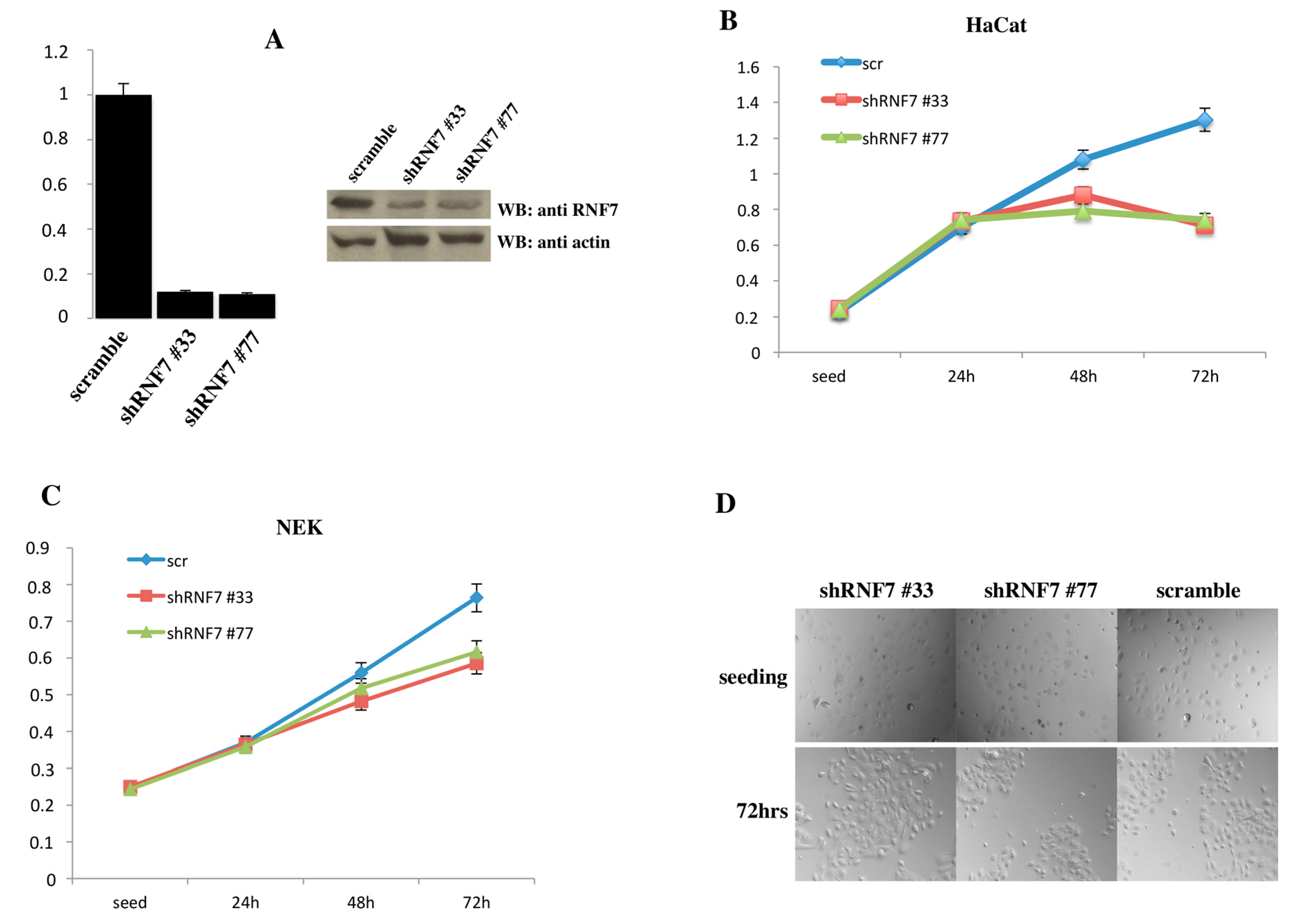
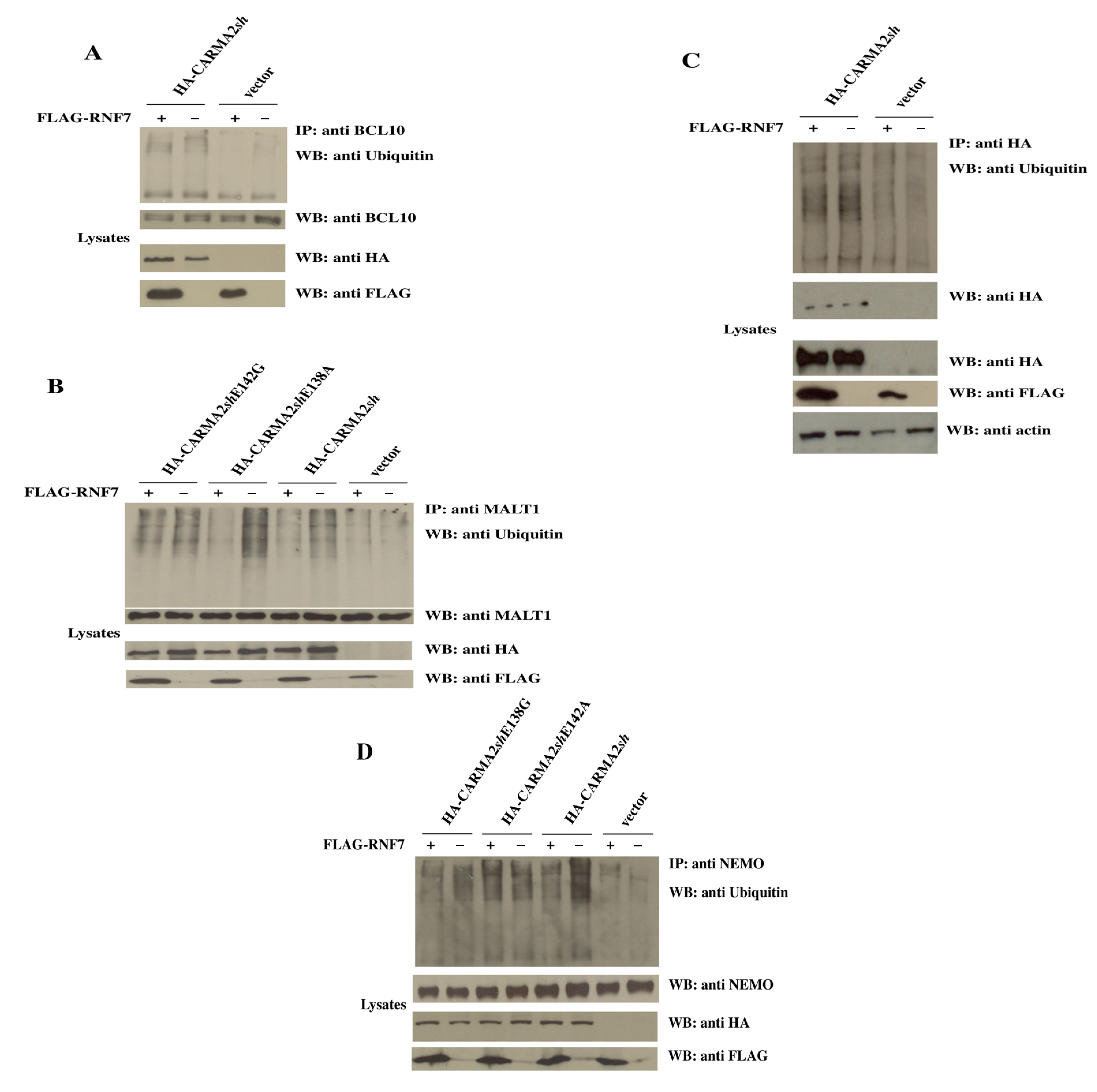
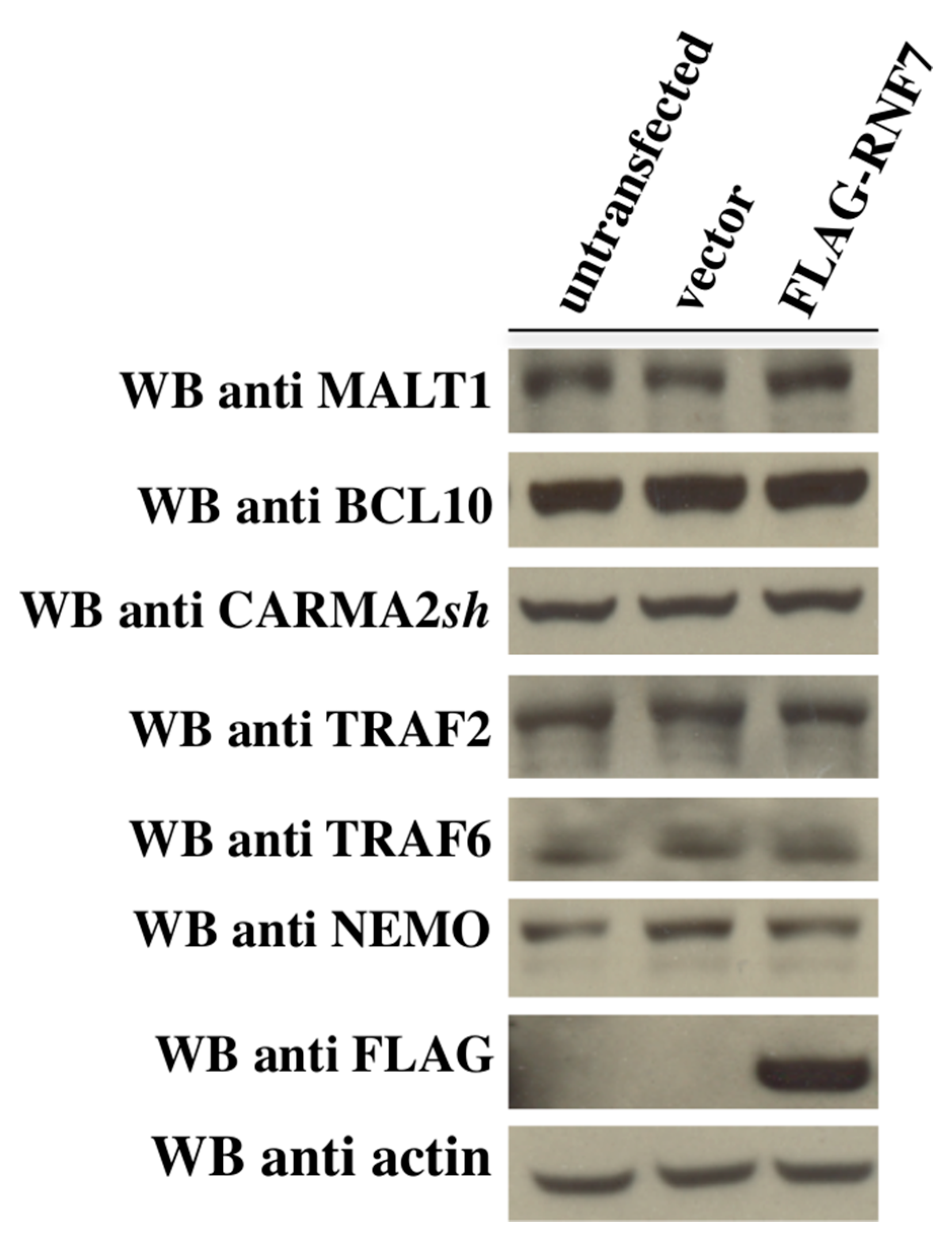
2. Results
3. Discussion
4. Materials and Methods
4.1. The Two-Hybrid Screening
4.2. Cell Culture and Tranfection
4.3. Immunoblot Analysis and Coprecipitation
4.4. Luciferase and β-Galactosidase Assays
4.5. Real-Time RT-PCR
4.6. MTT Assay
- TRCN0000298777 (#77):CCGGCCTGTGGGTGAAACAGAACAACTCGAGTTGTTCTGTTTCACCCACAGGTTTTTG
- TRCN0000295933 (#33):CCGGGTAATCCAGTGCCCTACAAAGCTCGAGCTTTGTAGGGCACTGGATTACTTTTTG.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blonska, M.; Lin, X. NF-κB signaling pathways regulated by CARMA family of scaffold proteins. Cell Res. 2011, 21, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, I.; Vito, P.; Stilo, R. The three CARMA sisters: So different, so similar: A portrait of the three CARMA proteins and their involvement in human disorders. J. Cell. Physiol. 2014, 229, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Stilo, R.; Liguoro, D.; Di Jeso, B.; Formisano, S.; Consiglio, E.; Leonardi, A.; Vito, P. Physical and functional interaction of CARMA1 and CARMA3 with Ikappa kinase gamma-NFkappaB essential modulator. J. Biol. Chem. 2004, 279, 34323–34331. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, I.; Mazzone, P.; D’Andrea, L.E.; Ferravante, A.; Zotti, T.; Telesio, G.; De Rubis, G.; Reale, C.; Pizzulo, M.; Muralitharan, S.; et al. CARMA2sh and ULK2 control pathogen-associated molecular patterns recognition in human keratinocytes: Psoriasis-linked CARMA2sh mutants escape ULK2 censorship. Cell Death Dis. 2017, 8, e2627. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Pierson, K.C.; Yang, C.F.; Joyce, C.E.; Ryan, C.; Duan, S.; Helms, C.A.; Liu, Y.; et al. PSORS2 is due to mutations in CARD14. Am. J. Hum. Genet. 2012, 90, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and common variants in CARD14, encoding an epidermal regulator of NF-kappaB, in psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Fuchs-Telem, D.; Sarig, O.; van Steensel, M.A.; Isakov, O.; Israeli, S.; Nousbeck, J.; Richard, K.; Winnepenninckx, V.; Vernooij, M.; Shomron, N.; et al. Familial pityriasis rubra pilaris is caused by mutations in CARD14. Am. J. Hum. Genet. 2012, 91, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.; Jordan, C.T.; Cao, L.; Lim, E.; Bouchlaka Souissi, C.; Jrad, A.; Omrane, I.; Kouidhi, S.; Zaraa, I.; Anbunathan, H.; et al. CARD14 alterations in Tunisian patients with psoriasis and further characterization in European cohorts. Br. J. Dermatol. 2016, 174, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.; Bouchlaka-Souissi, C.; Helms, C.A.; Zaraa, I.; Jordan, C.T.; Anbunathan, H.; Bouhaha, R.; Kouidhi, S.; Doss, N.; Dhaoui, R.; et al. Genome-wide linkage scan for psoriasis susceptibility loci in multiplex Tunisian families. Br. J. Dermatol. 2013, 168, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Eskin-Schwartz, M.; Basel-Vanagaite, L.; David, M.; Lagovsky, I.; Ben-Amitai, D.; Smirin-Yosef, P.; Atzmony, L.; Hodak, E. Intra-familial variation in clinical phenotype of CARD14-related Psoriasis. Acta Derm. Venereol. 2016, 96, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, T.; Li, S.J.; Fan, Y.M.; Shi, G.; Zhu, K.J. CARD14 gene polymorphism c.C2458T (p.Arg820Trp) is associated with clinical features of psoriasis vulgaris in a Chinese cohort. J. Dermatol. 2016, 43, 294–297. [Google Scholar] [CrossRef] [PubMed]
- González-Lara, L.; Coto-Segura, P.; Penedo, A.; Eiris, N.; Díaz, M.; Santos-Juanes, J.; Queiro, R.; Coto, E. SNP rs11652075 in the CARD14 Gene as a risk factor for Psoriasis (PSORS2) in a Spanish Cohort. DNA Cell Biol. 2013, 32, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Dainichi, T.; Fujisawa, A.; Nakano, H.; Sawamura, D.; Kabashima, K. CARD14 Glu138 mutation in familial pityriasis rubra pilaris does not warrant differentiation from familial psoriasis. J. Dermatol. 2016, 43, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Mössner, R.; Frambach, Y.; Wilsmann-Theis, D.; Löhr, S.; Jacobi, A.; Weyergraf, A.; Müller, M.; Philipp, S.; Renner, R.; Traupe, H.; et al. Palmoplantar Pustular Psoriasis Is Associated with Missense Variants in CARD14, but Not with Loss-of-Function Mutations in IL36RN in European Patients. J. Investig. Dermatol. 2015, 135, 2538–2541. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Zhang, Q.; Chen, M.; Fu, X.; Wang, C.; Wang, Z.; Yu, G.; Yu, Y.; Li, X.; Sun, Y.; et al. Variant analysis of CARD14 in a Chinese Han population with psoriasis vulgaris and generalized pustular psoriasis. J. Investig. Dermatol. 2014, 134, 2994–2996. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, K.; Muto, M.; Akiyama, M. CARD14 c.526G>C (p.Asp176His) is a significant risk factor for generalized pustular psoriasis with psoriasis vulgaris in the Japanese cohort. J. Investig. Dermatol. 2014, 134, 1755–1757. [Google Scholar] [CrossRef] [PubMed]
- Berki, D.M.; Liu, L.; Choon, S.E.; David Burden, A.; Griffiths, C.E.M.; Navarini, A.A.; Tan, E.S.; Irvine, A.D.; Ranki, A.; Ogo, T.; et al. Activating CARD14 Mutations Are Associated with Generalized Pustular Psoriasis but Rarely Account for Familial Recurrence in Psoriasis Vulgaris. J. Investig. Dermatol. 2015, 135, 2964–2970. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Shi, G.; Liu, H.; Zhu, C.; Fan, Y. Variants of CARD14 gene and psoriasis vulgaris in southern Chinese cohort. An. Bras. Dermatol. 2016, 91, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Van Nuffel, E.; Schmitt, A.; Afonina, I.S.; Schulze-Osthoff, K.; Beyaert, R.; Hailfinger, S. CARD14-Mediated Activation of Paracaspase MALT1 in Keratinocytes: Implications for Psoriasis. J. Investig. Dermatol. 2017, 137, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, I.; Zotti, T.; Ferravante, A.; Vessichelli, M.; Vito, P.; Stilo, R. Alternative splicing of CARMA2/CARD14 transcripts generates protein variants with differential effect on NF-κB activation and endoplasmic reticulum stress-induced cell death. J. Cell. Physiol. 2011, 226, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.S.; van Nuffel, E.; Baudelet, G.; Driege, Y.; Kreike, M.; Staal, J.; Beyaert, R. The paracaspase MALT1 mediates CARD14-induced signaling in keratinocytes. EMBO Rep. 2016, 17, 914–927. [Google Scholar] [CrossRef] [PubMed]
- Howes, A.; O’Sullivan, P.A.; Breyer, F.; Ghose, A.; Cao, L.; Krappmann, D.; Bowcock, A.M.; Ley, S.C. Psoriasis mutations disrupt CARD14 autoinhibition promoting BCL10-MALT1-dependent NF-κB activation. Biochem. J. 2016, 473, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Harden, J.L.; Lewis, S.M.; Pierson, K.C.; Suárez-Fariñas, M.; Lentini, T.; Ortenzio, F.S.; Zaba, L.C.; Goldbach-Mansky, R.; Bowcock, A.M.; Lowes, M.A. CARD14 expression in dermal endothelial cells in psoriasis. PLoS ONE 2014, 9, e111255. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Wang, Y.; Aviram, M.; Swaroop, M.; Loo, J.A.; Bian, J.; Tian, Y.; Mueller, T.; Bisgaier, C.L.; Sun, Y. SAG, a novel zinc RING finger protein that protects cells from apoptosis induced by redox agents. Mol. Cell. Biol. 1999, 19, 3145–3155. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, H. Functional characterization of SAG/RBX2/ROC2/RNF7, an antioxidant protein and an E3 ubiquitin ligase. Protein Cell 2013, 4, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Swaroop, M.; Wang, Y.; Miller, P.; Duan, H.; Jatkoe, T.; Madore, S.; Sun, Y. Yeast homolog of human SAG/ROC2/Rbx2 Rbx2/ Hrt2 is essential for cell growth, but not for germination: Chip profiling implicates its role in cell cycle regulation. Oncogene 2000, 19, 2855–2866. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S. Toll-like receptors in skin. Adv. Dermatol. 2008, 24, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Moreno-García, M.E.; Sommer, K.; Shinohara, H.; Bandaranayake, A.D.; Kurosaki, T.; Rawlings, D.J. MAGUK-controlled ubiquitination of CARMA1 modulates lymphocyte NF-kappaB activity. Mol. Cell. Biol. 2010, 30, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.M.; Chan, W.; Jattani, R.P.; Mackie, D.S.; Pomerantz, J.L. Negative Regulation of CARD11 Signaling and Lymphoma Cell Survival by the E3 Ubiquitin Ligase RNF181. Mol. Cell. Biol. 2015, 36, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Wegener, E.; Welteke, V.; Ferch, U.; Arslan, S.C.; Ruland, J.; Scheidereit, C.; Krappmann, D. Malt1 ubiquitination triggers NF-kappaB signaling upon T-cell activation. EMBO J. 2007, 26, 4634–4645. [Google Scholar] [CrossRef] [PubMed]
- Aksentijevich, I.; Zhou, Q. NF-κB Pathway in Autoinflammatory Diseases: Dysregulation of Protein Modifications by Ubiquitin Defines a New Category of Autoinflammatory Diseases. Front. Immunol. 2017, 8, 399. [Google Scholar] [CrossRef] [PubMed]
- Sebban, H.; Yamaoka, S.; Courtois, G. Posttranslational modifications of NEMO and its partners in NF-kappaB signaling. Trends Cell Biol. 2006, 16, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.K.; Yang, C.; Chan, W.; Wang, Z.; Deibel, K.E.; Pomerantz, J.L. Molecular Determinants of Scaffold-induced Linear Ubiquitinylation of B Cell Lymphoma/Leukemia 10 (Bcl10) during T Cell Receptor and Oncogenic Caspase Recruitment Domain-containing Protein 11 (CARD11) Signaling. J. Biol. Chem. 2016, 291, 25921–25936. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, E.L.; Ferravante, A.; Scudiero, I.; Zotti, T.; Reale, C.; Pizzulo, M.; De La Motte, L.R.; De Maio, C.; Mazzone, P.; Telesio, G.; et al. The Dishevelled, EGL-10 and pleckstrin (DEP) domain-containing protein DEPDC7 binds to CARMA2 and CARMA3 proteins, and regulates NF-κB activation. PLoS ONE 2014, 9, e116062. [Google Scholar] [CrossRef]
- Guiet, C.; Silvestri, E.; de Smaele, E.; Franzoso, G.; Vito, P. c-FLIP efficiently rescues TRAF-2−/− cells from TNF-induced apoptosis. Cell Death Differ. 2002, 9, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, I.; Zotti, T.; Ferravante, A.; Vessichelli, M.; Reale, C.; Masone, M.C.; Leonardi, A.; Vito, P.; Stilo, R. Tumor necrosis factor (TNF) receptor-associated factor 7 is required for TNFα-induced Jun NH2-terminal kinase activation and promotes cell death by regulating polyubiquitination and lysosomal degradation of c-FLIP protein. J. Biol. Chem. 2012, 287, 6053–6061. [Google Scholar] [CrossRef] [PubMed]
- Guiet, C.; Vito, P. Caspase recruitment domain (CARD)-dependent cytoplasmic filaments mediate bcl10-induced NF-kappaB activation. J. Cell Biol. 2000, 148, 1131–1140. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Fused to GAL4 Domain | Yeast Growth on Selective Media | |
|---|---|---|
| DNA-Binding | Activating | |
| - | RNF7 | − |
| Vector | RNF7 | − |
| Fas-associated protein with death domain (FADD) | RNF7 | − |
| CARMA2sh | RNF7 | + |
| CARMA2sh | - | − |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telesio, G.; Scudiero, I.; Pizzulo, M.; Mazzone, P.; Zotti, T.; Voccola, S.; Polvere, I.; Vito, P.; Stilo, R. The E3 Ubiquitin Ligase RNF7 Negatively Regulates CARD14/CARMA2sh Signaling. Int. J. Mol. Sci. 2017, 18, 2581. https://doi.org/10.3390/ijms18122581
Telesio G, Scudiero I, Pizzulo M, Mazzone P, Zotti T, Voccola S, Polvere I, Vito P, Stilo R. The E3 Ubiquitin Ligase RNF7 Negatively Regulates CARD14/CARMA2sh Signaling. International Journal of Molecular Sciences. 2017; 18(12):2581. https://doi.org/10.3390/ijms18122581
Chicago/Turabian StyleTelesio, Gianluca, Ivan Scudiero, Maddalena Pizzulo, Pellegrino Mazzone, Tiziana Zotti, Serena Voccola, Immacolata Polvere, Pasquale Vito, and Romania Stilo. 2017. "The E3 Ubiquitin Ligase RNF7 Negatively Regulates CARD14/CARMA2sh Signaling" International Journal of Molecular Sciences 18, no. 12: 2581. https://doi.org/10.3390/ijms18122581