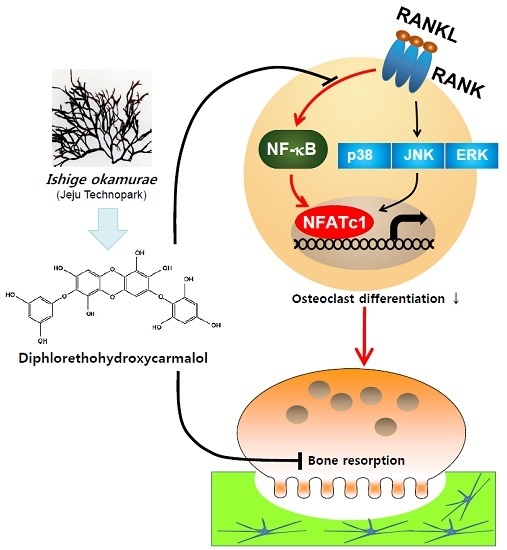
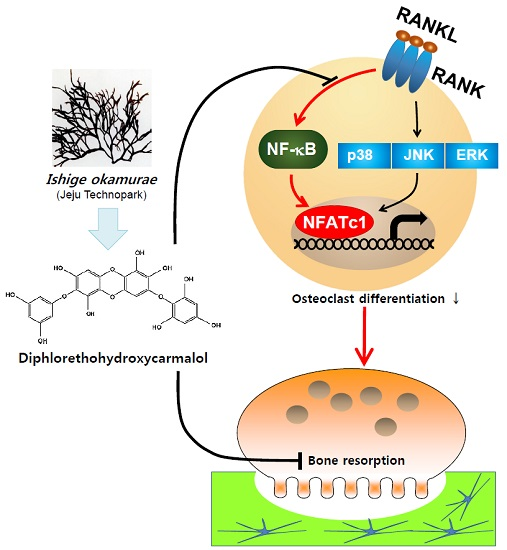
Diphlorethohydroxycarmalol from Ishige okamurae Suppresses Osteoclast Differentiation by Downregulating the NF-κB Signaling Pathway

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
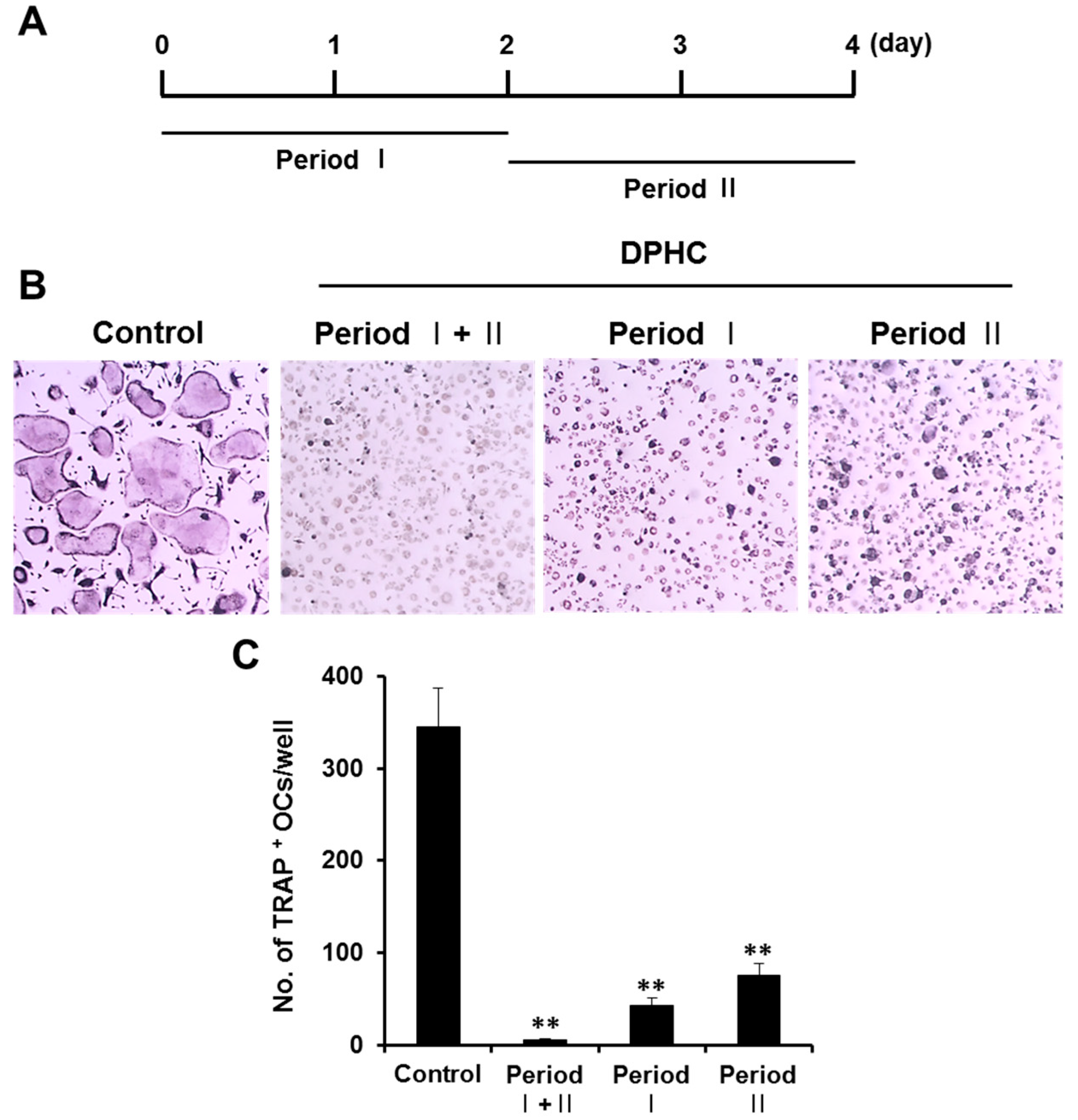
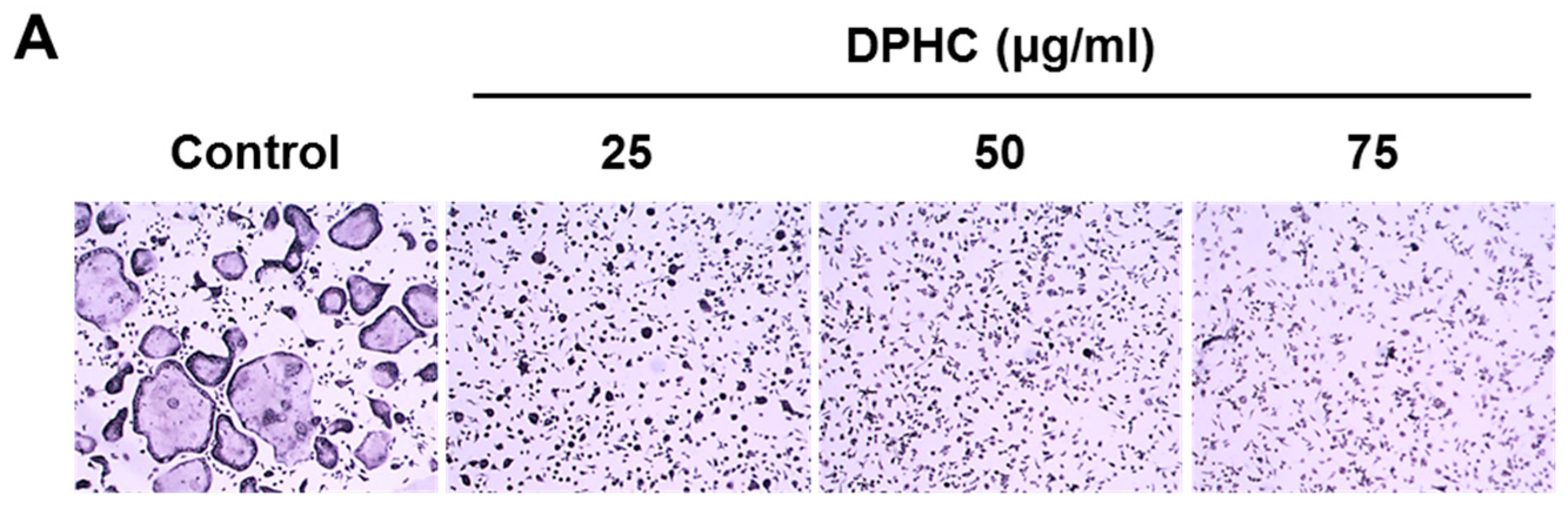
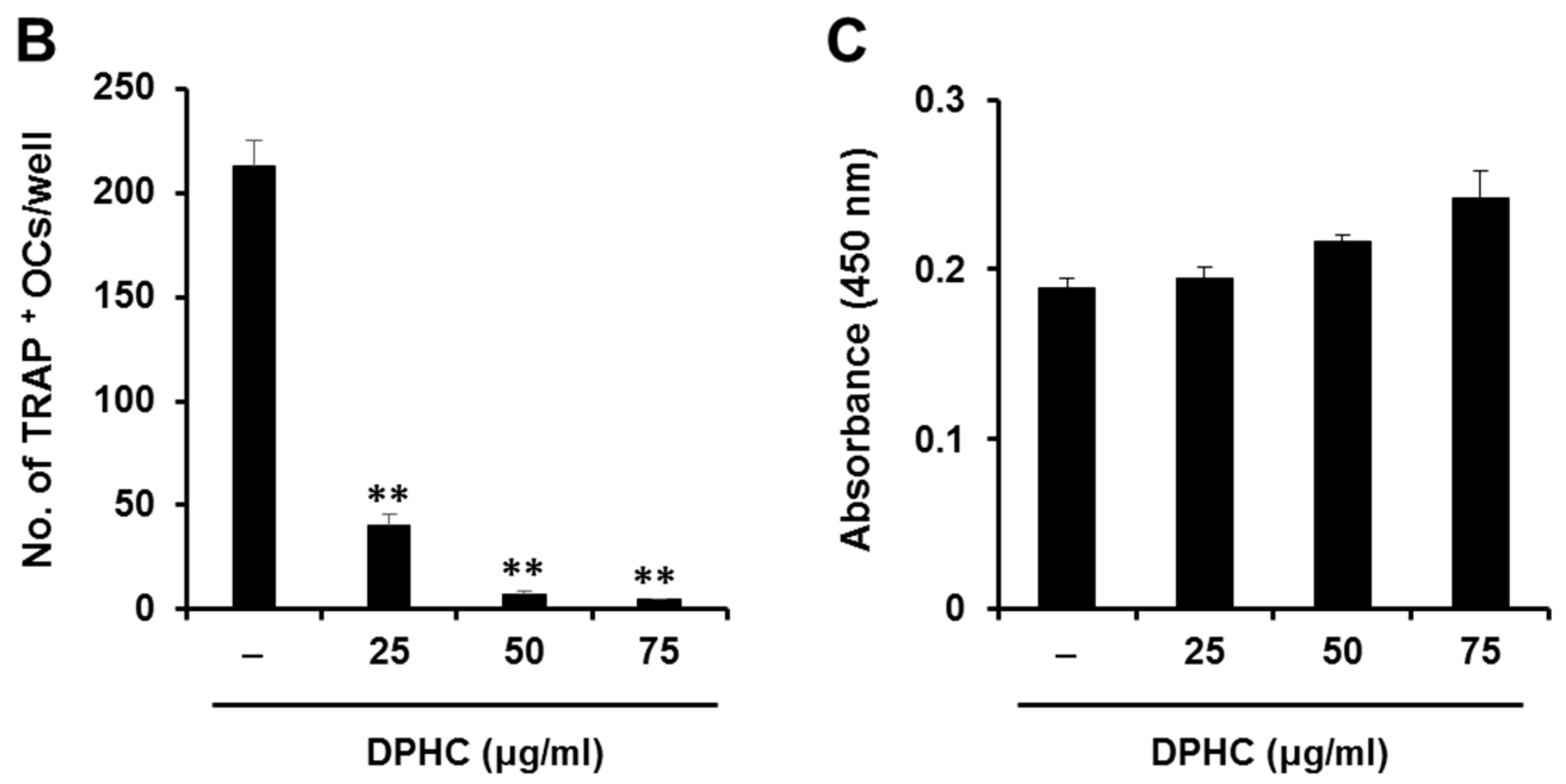
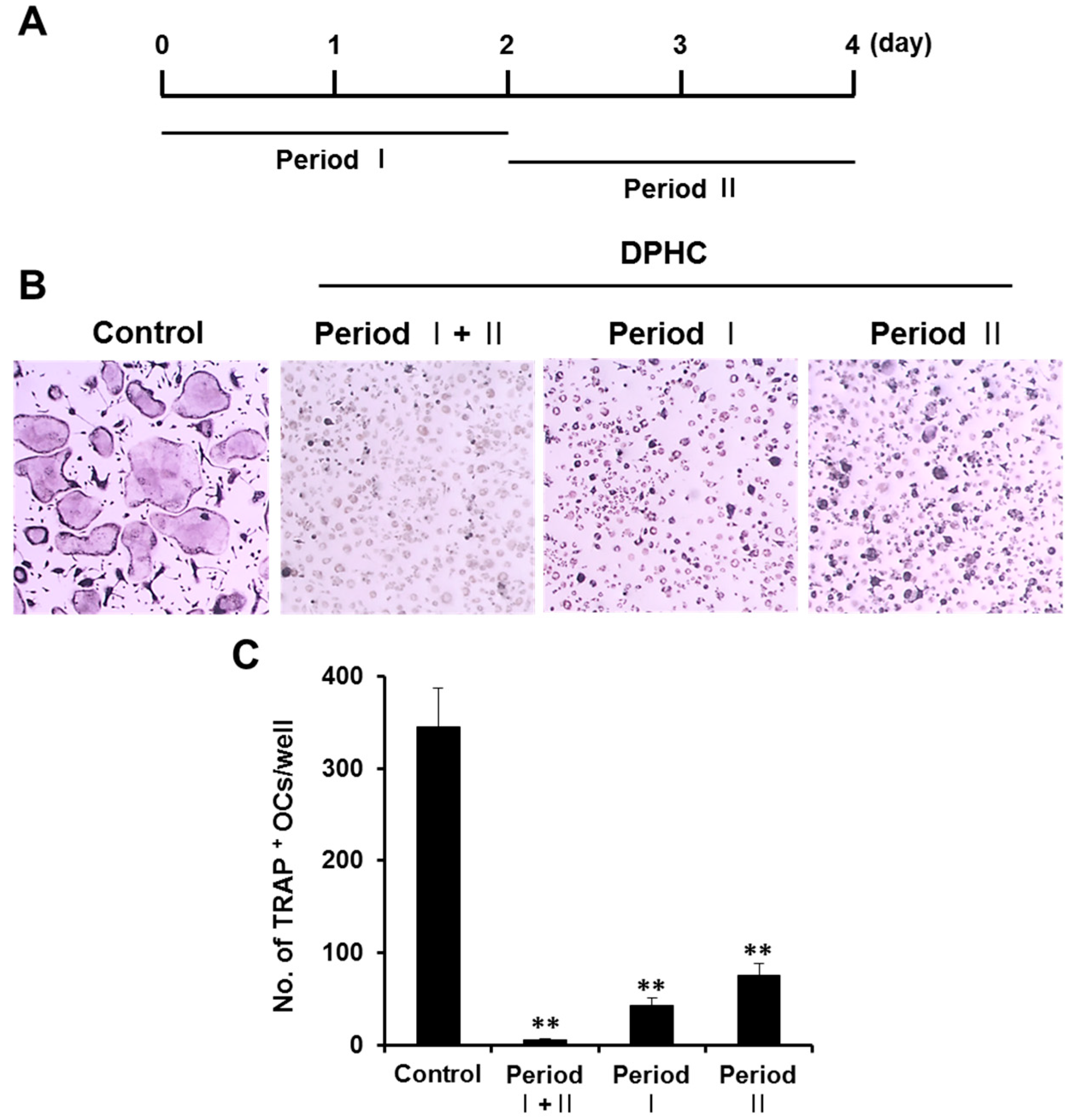
2.1. Effects of Diphlorethohydroxycarmalol (DPHC) on the Viability of Bone Marrow Macrophages (BMMs) and Osteoclast Formation
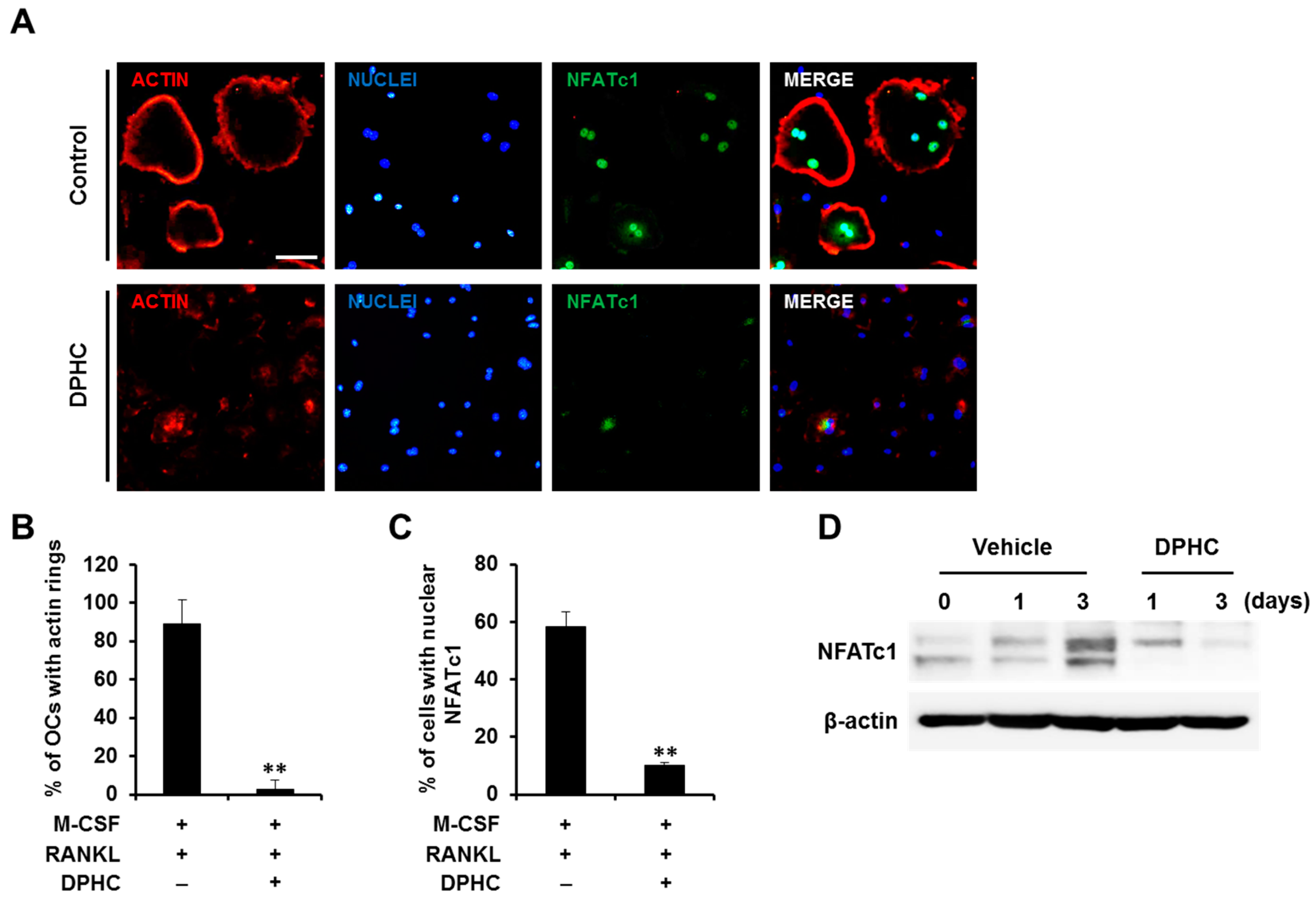
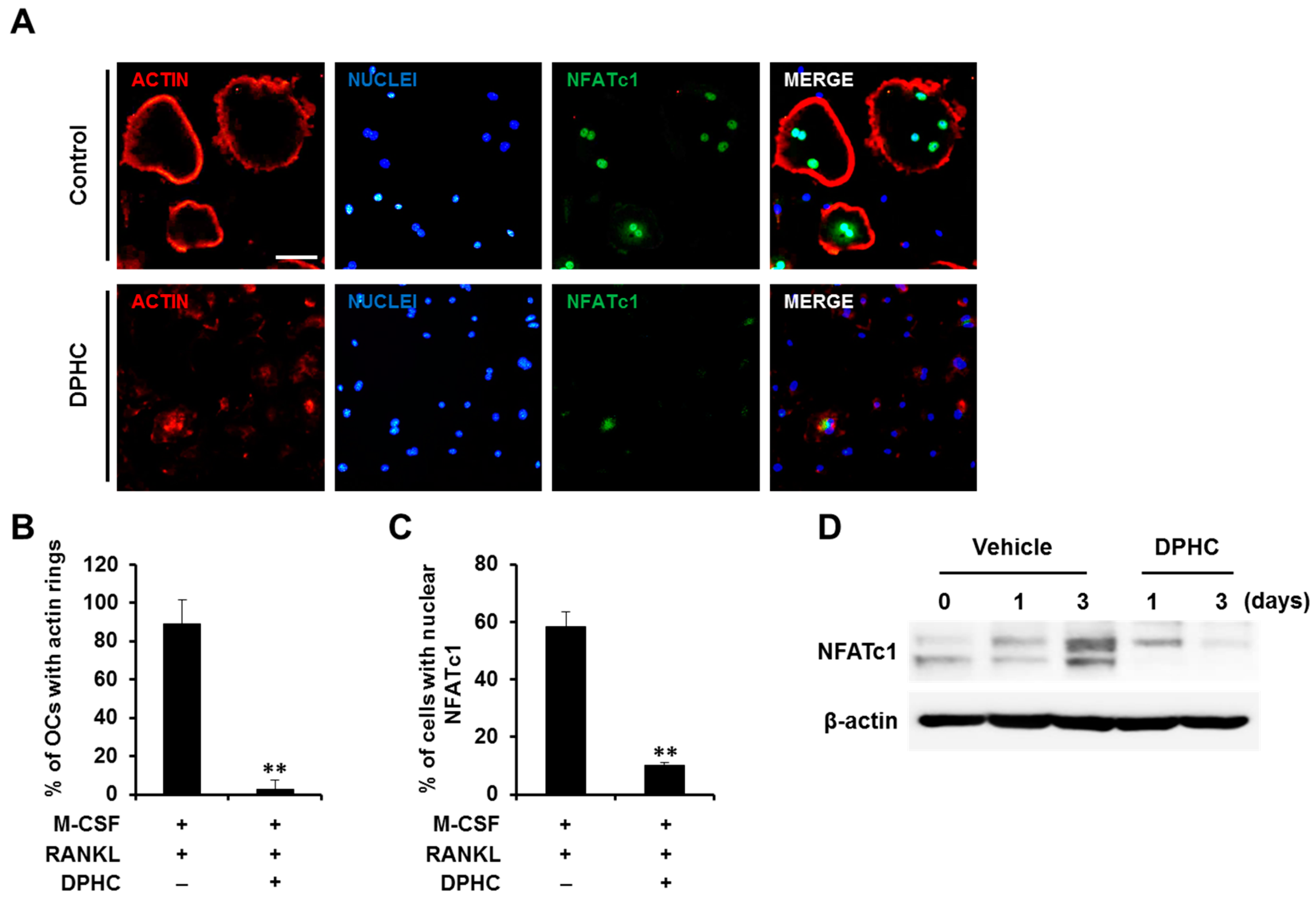
2.2. Effects of DPHC on Actin Ring Formation and NFATc1 Expression/Localization
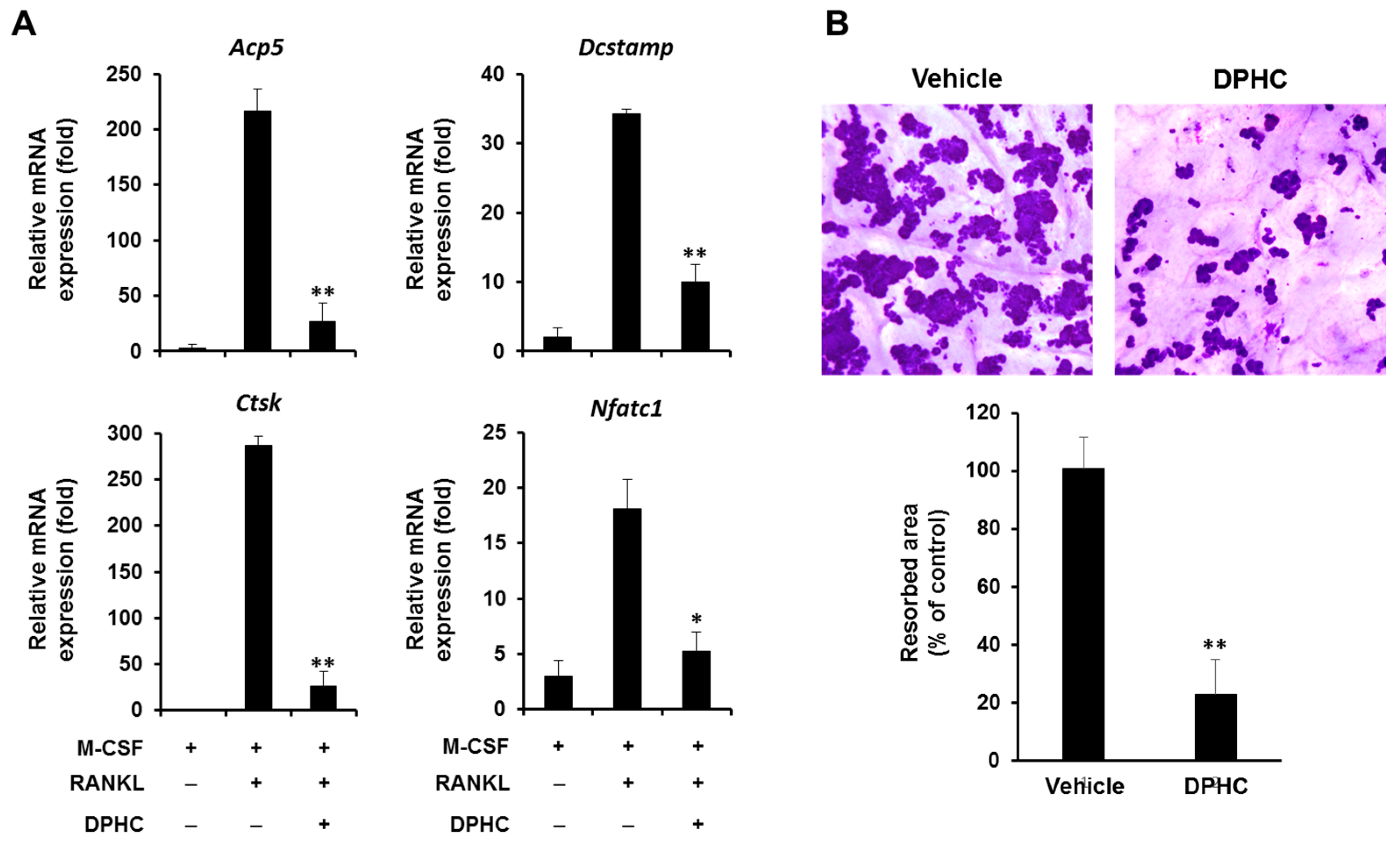
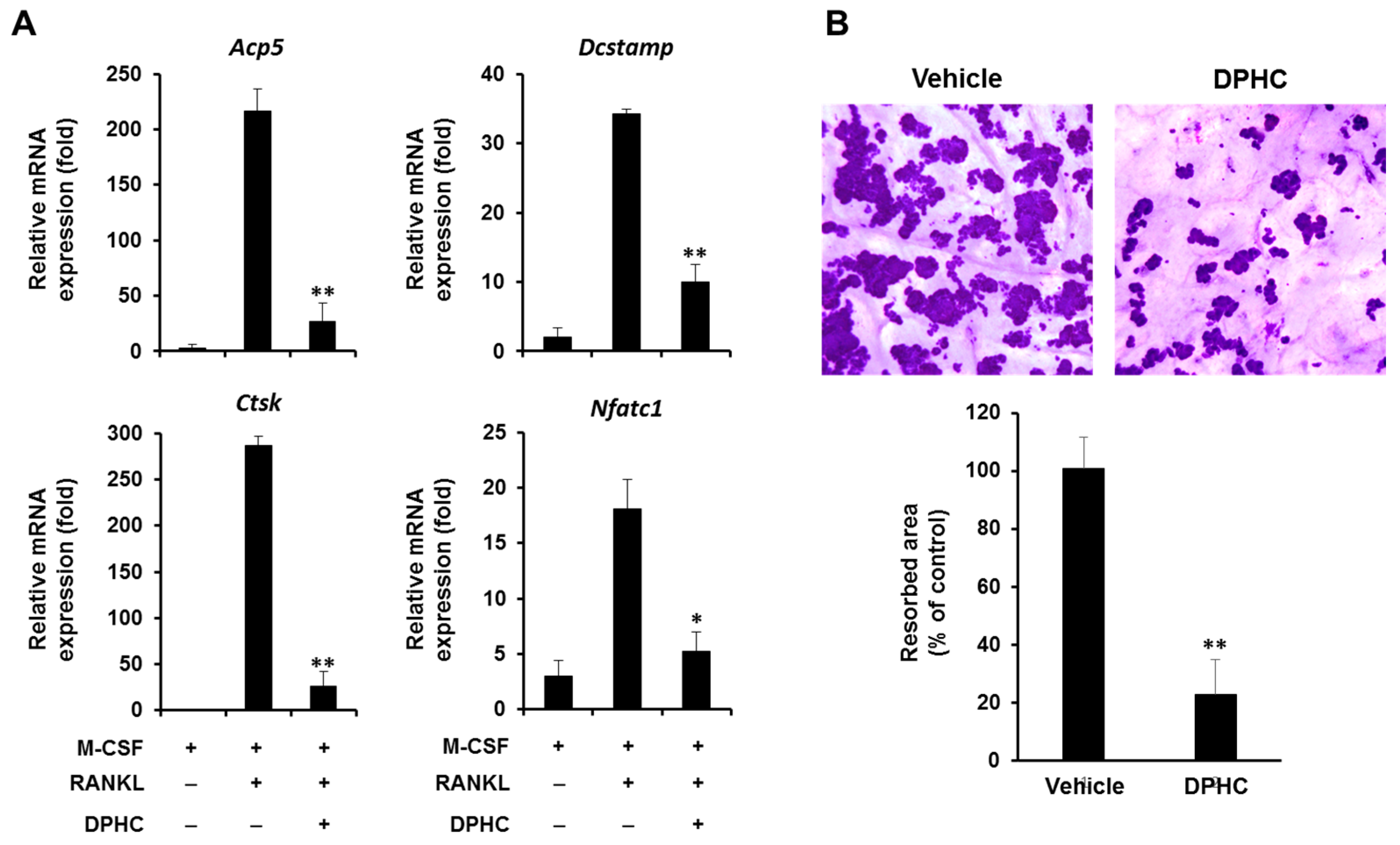
2.3. Effect of DPHC on the Expression of Osteoclastic Markers
2.4. Effect of DPHC on Bone Resorption
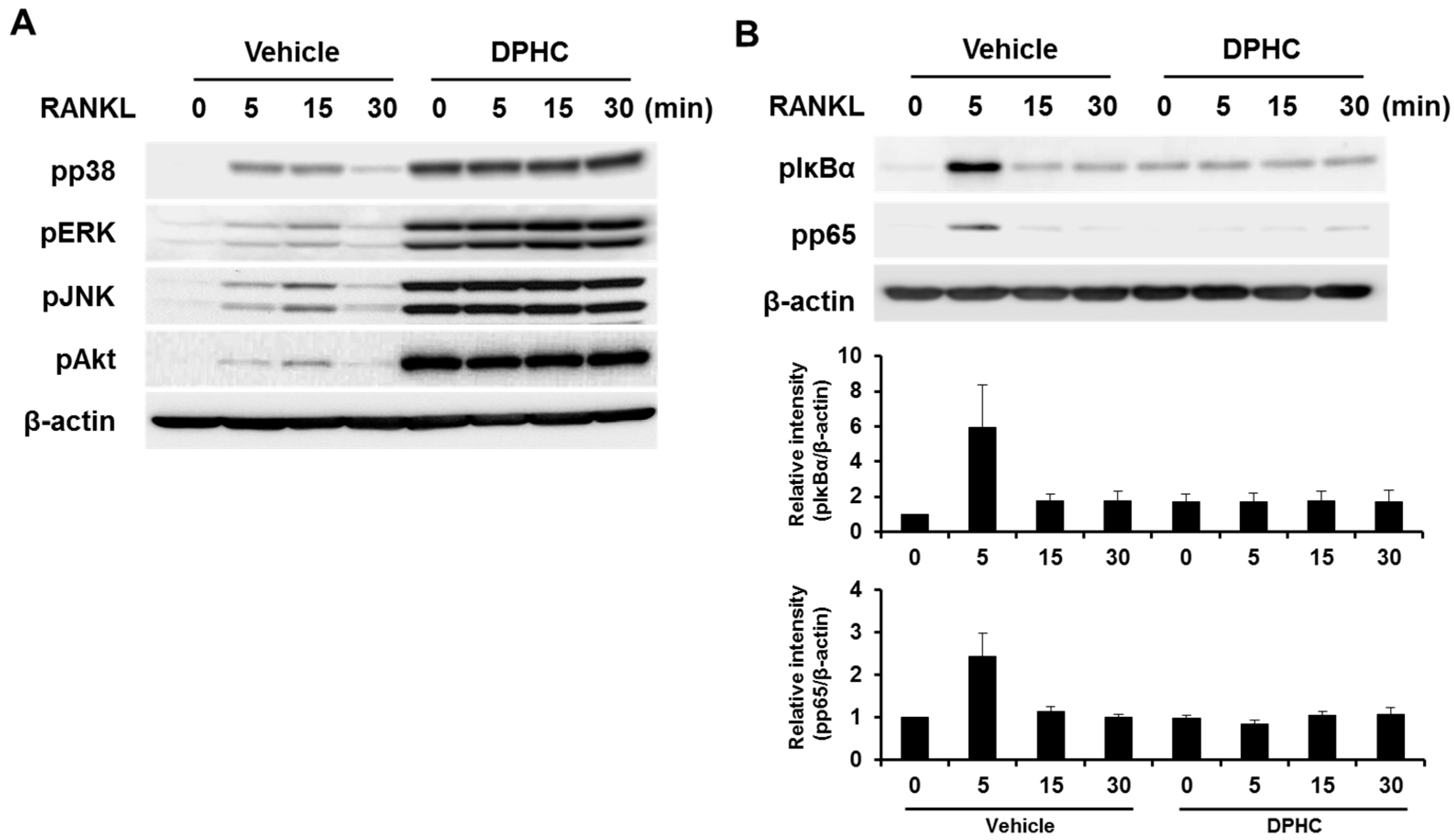
2.5. Effect of DPHC on the RANKL-Induced Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Osteoclast Differentiation
4.3. Cell Viability Assay
4.4. RNA Isolation and Real-Time PCR
4.5. Western Blotting
4.6. Immunofluorescence
4.7. Bone Resorption Pit Assay
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Manolagas, S.C.; Jilka, R.L. Bone marrow, cytokines, and bone remodeling. Emerging insights into the pathophysiology of osteoporosis. N. Engl. J. Med. 1995, 332, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Ralston, S.H.; Layfield, R. Pathogenesis of paget disease of bone. Calcif. Tissue Int. 2012, 91, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Mundy, G.R. Osteoporosis and inflammation. Nutr. Rev. 2007, 65, S147–S151. [Google Scholar] [CrossRef] [PubMed]
- Roodman, G.D. Biology of osteoclast activation in cancer. J. Clin. Oncol. 2001, 19, 3562–3571. [Google Scholar] [CrossRef] [PubMed]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef]
- Tanaka, S.; Takahashi, N.; Udagawa, N.; Tamura, T.; Akatsu, T.; Stanley, E.R.; Kurokawa, T.; Suda, T. Macrophage colony-stimulating factor is indispensable for both proliferation and differentiation of osteoclast progenitors. J. Clin. Investig. 1993, 91, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Kim, J.P.; Jung, W.K.; Lee, N.H.; Kang, H.S.; Jun, E.M.; Park, S.H.; Kang, S.M.; Lee, Y.J.; Park, P.J.; et al. Identification of chemical structure and free radical scavenging activity of diphlorethohydroxycarmalol isolated from a brown alga, ishige okamurae. J. Microbiol. Biotechnol. 2008, 18, 676–681. [Google Scholar] [PubMed]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from ishige okamurae, a brown algae, a potent alpha-glucosidase and alpha-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Han, S.C.; Kang, G.J.; Koo, D.H.; Koh, Y.S.; Hyun, J.W.; Lee, N.H.; Ko, M.H.; Kang, H.K.; Yoo, E.S. Diphlorethohydroxycarmalol inhibits interleukin-6 production by regulating nf-kappab, stat5 and socs1 in lipopolysaccharide-stimulated raw264.7 cells. Mar. Drugs 2015, 13, 2141–2157. [Google Scholar] [CrossRef] [PubMed]
- Kotake-Nara, E.; Asai, A.; Nagao, A. Neoxanthin and fucoxanthin induce apoptosis in pc-3 human prostate cancer cells. Cancer Lett. 2005, 220, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Hamann, M.T. Marine pharmacology in 2001–2002: Marine compounds with anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 265–286. [Google Scholar] [PubMed]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Lee, S.H.; Park, P.J.; Kang, D.H.; Oh, C.; Kim, D.W.; Han, J.S.; Jeon, Y.J.; et al. Protective effect of diphlorethohydroxycarmalol isolated from ishige okamurae against high glucose-induced-oxidative stress in human umbilical vein endothelial cells. Food Chem. Toxicol. 2010, 48, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Deng, L.; Chen, W.; Shao, J.; Xu, G.; Li, Y.P. Atp6v1c1 is an essential component of the osteoclast proton pump and in f-actin ring formation in osteoclasts. Biochem. J. 2009, 417, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.B.; Kim, J.Y.; Kim, K.J.; Choi, M.K.; Kim, J.J.; Kim, K.M.; Shin, Y.I.; Lee, M.S.; Kim, H.S.; Kim, J.W.; et al. Risedronate directly inhibits osteoclast differentiation and inflammatory bone loss. Biol. Pharm. Bull. 2009, 32, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Akatsu, T.; Katayama, Y.; Yasutomo, Y.; Kado, S.; Kugal, N.; Yamamoto, M.; Nagata, N. Bisphosphonates act on osteoblastic cells and inhibit osteoclast formation in mouse marrow cultures. Bone 1996, 18, 9–14. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; Sasso, G.R.; Sasso-Cerri, E.; Simoes, M.J.; Cerri, P.S. Biology of bone tissue: Structure, function, and factors that influence bone cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Martin, T.J. Coupling signals between the osteoclast and osteoblast how are messages transmitted between these temporary visitors to the bone surface? Front. Endocrinol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Sivolella, S.; Lumachi, F.; Stellini, E.; Favero, L. Denosumab and anti-angiogenetic drug-related osteonecrosis of the jaw: An uncommon but potentially severe disease. Anticancer Res. 2013, 33, 1793–1797. [Google Scholar] [PubMed]
- Che, C.T.; Wong, M.S.; Lam, C.W. Natural products from Chinese medicines with potential benefits to bone health. Molecules 2016, 21, 239. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, J.H.; Lee, J.; Jin, H.M.; Lee, S.H.; Fisher, D.E.; Kook, H.; Kim, K.K.; Choi, Y.; Kim, N. Nuclear factor of activated t cells c1 induces osteoclast-associated receptor gene expression during tumor necrosis factor-related activation-induced cytokine-mediated osteoclastogenesis. J. Biol. Chem. 2005, 280, 35209–35216. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Sato, K.; Asagiri, M.; Morita, I.; Soma, K.; Takayanagi, H. Contribution of nuclear factor of activated t cells c1 to the transcriptional control of immunoreceptor osteoclast-associated receptor but not triggering receptor expressed by myeloid cells-2 during osteoclastogenesis. J. Biol. Chem. 2005, 280, 32905–32913. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Hisatake, K.; Nogi, Y. Essential role of p38 mitogen-activated protein kinase in cathepsin k gene expression during osteoclastogenesis through association of nfatc1 and pu.1. J. Biol. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor nfatc1 (nfat2) integrate rankl signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Johnson, R.S.; Spiegelman, B.M.; Papaioannou, V. Pleiotropic effects of a null mutation in the c-fos proto-oncogene. Cell 1992, 71, 577–586. [Google Scholar] [CrossRef]
- Takatsuna, H.; Asagiri, M.; Kubota, T.; Oka, K.; Osada, T.; Sugiyama, C.; Saito, H.; Aoki, K.; Ohya, K.; Takayanagi, H.; et al. Inhibition of rankl-induced osteoclastogenesis by (−)-dhmeq, a novel nf-kappab inhibitor, through downregulation of nfatc1. J. Bone Miner. Res. 2005, 20, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Ovitt, C.; Grigoriadis, A.E.; Mohle-Steinlein, U.; Ruther, U.; Wagner, E.F. Bone and haematopoietic defects in mice lacking c-fos. Nature 1992, 360, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.J.; Lee, D.; Lee, T.; Kim, S.H.; Shin, H.I.; Bae, Y.C.; Hong, J.M.; Park, E.K. Inhibitory effects of kp-a159, a thiazolopyridine derivative, on osteoclast differentiation, function, and inflammatory bone loss via suppression of rankl-induced map kinase signaling pathway. PLoS ONE 2015, 10, e0142201. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.J.; Lee, T.; Kim, J.A.; Lee, D.; Kim, N.D.; Shin, H.I.; Bae, Y.C.; Park, E.K. Ocli-023, a novel pyrimidine compound, suppresses osteoclastogenesis in vitro and alveolar bone resorption in vivo. PLoS ONE 2017, 12, e0170159. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ihn, H.J.; Kim, J.A.; Cho, H.S.; Shin, H.-I.; Kim, G.-Y.; Choi, Y.H.; Jeon, Y.-J.; Park, E.K. Diphlorethohydroxycarmalol from Ishige okamurae Suppresses Osteoclast Differentiation by Downregulating the NF-κB Signaling Pathway. Int. J. Mol. Sci. 2017, 18, 2635. https://doi.org/10.3390/ijms18122635
Ihn HJ, Kim JA, Cho HS, Shin H-I, Kim G-Y, Choi YH, Jeon Y-J, Park EK. Diphlorethohydroxycarmalol from Ishige okamurae Suppresses Osteoclast Differentiation by Downregulating the NF-κB Signaling Pathway. International Journal of Molecular Sciences. 2017; 18(12):2635. https://doi.org/10.3390/ijms18122635
Chicago/Turabian StyleIhn, Hye Jung, Ju Ang Kim, Hye Sung Cho, Hong-In Shin, Gi-Young Kim, Yung Hyun Choi, You-Jin Jeon, and Eui Kyun Park. 2017. "Diphlorethohydroxycarmalol from Ishige okamurae Suppresses Osteoclast Differentiation by Downregulating the NF-κB Signaling Pathway" International Journal of Molecular Sciences 18, no. 12: 2635. https://doi.org/10.3390/ijms18122635