
HDAC Inhibitors and RECK Modulate Endoplasmic Reticulum Stress in Tumor Cells

Abstract
:1. Endoplasmic Reticulum Stress in Cancers
2. Histone Deacetylase (HDAC) Inhibitors Induce ER Stress and Cause Cell Death in Cancer Cells
3. Reversion-Inducing Cysteine-Rich Protein with Kazal Motifs (RECK) and Regulation of RECK Expression
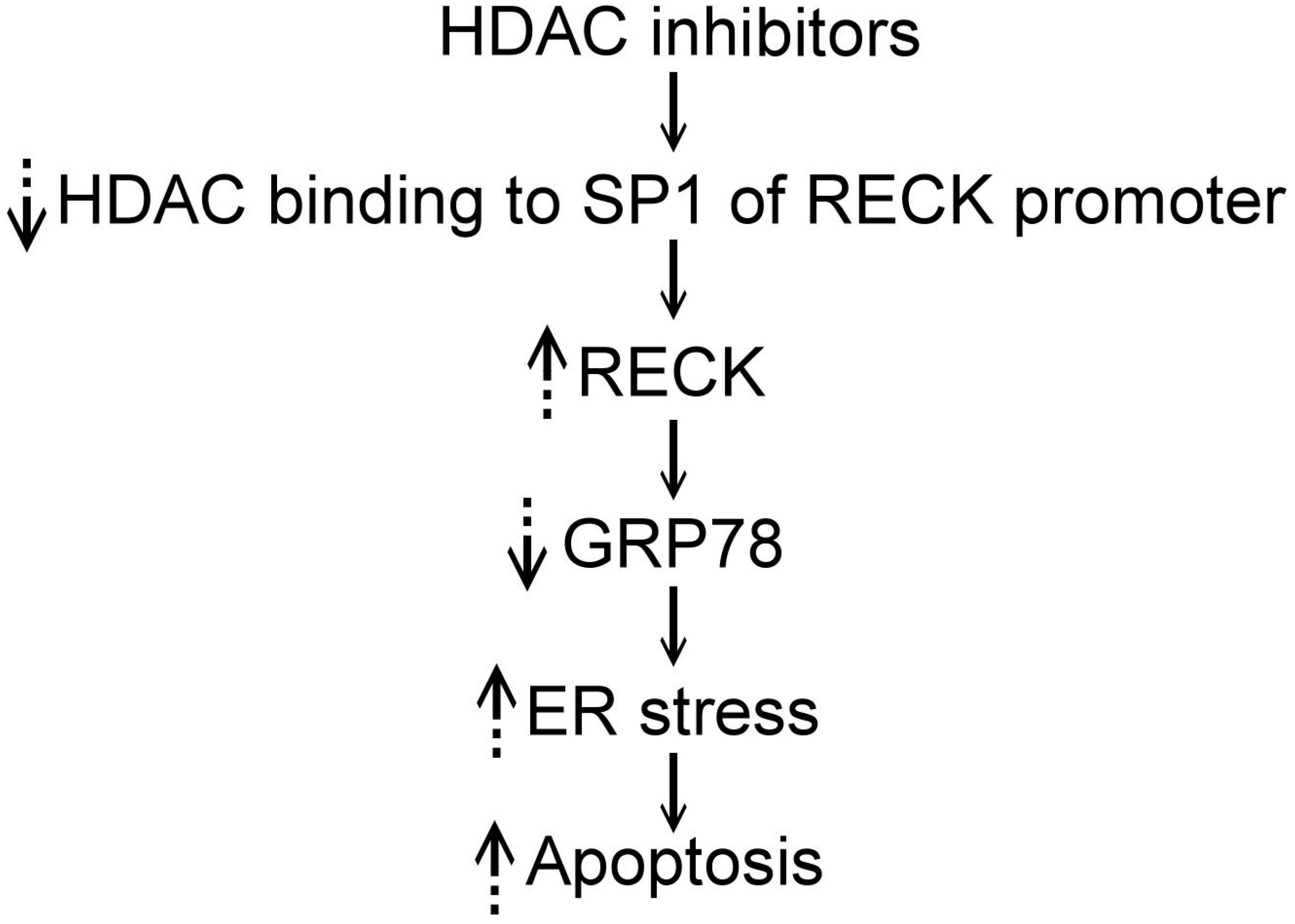
4. HDAC Inhibitors, RECK, and ER Stress
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Clarke, H.J.; Chambers, J.E.; Liniker, E.; Marciniak, S.J. Endoplasmic reticulum stress in malignancy. Cancer Cell 2014, 25, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Zhang, Y.; Lee, A.S. Beyond the endoplasmic reticulum: Atypical GRP78 in cell viability, signaling and therapeutic targeting. Biochem. J. 2011, 434, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Penaranda Fajardo, N.M.; Meijer, C.; Kruyt, F.A. The endoplasmic reticulum stress/unfolded protein response in gliomatosis, tumor progression and as a therapeutic target in glioblastoma. Biochem. Pharmacol. 2016, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. GRP78 induction in cancer: Therapeutic and prognostic implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Zhang, X. Roles of GRP78 in physiology and cancer. J. Cell Biochem. 2010, 110, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lee, A.S. Stress induction of GRP78/BiP and its role in cancer. Curr. Mol. Med. 2006, 6, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Zismanov, V.; Drucker, L.; Gottfried, M. ER homeostasis and motility of NSCLC cell lines can be therapeutically targeted with combined Hsp90 and HDAC inhibitors. Pulm. Pharmacol. Ther. 2013, 26, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Ko, B.; Baumeister, P.; Swenson, S.; Costa, F.; Markland, F.; Stiles, C.; Patterson, J.B.; Bates, S.E.; Lee, A.S. Vascular targeting and antiangiogenesis agents induce drug resistance effector GRP78 within the tumor microenvironment. Cancer Res. 2005, 65, 5785–5791. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.C.; Mao, Z.G.; Avery-Kiejda, K.A.; Wade, M.; Hersey, P.; Zhang, X.D. Glucose-regulated protein 78 antagonizes cisplatin and adriamycin in human melanoma cells. Carcinogenesis 2009, 30, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Lin, Y.; Li, J.; Harding, H.P.; Ron, D.; Jamison, S. A deregulated integrated stress response promotes interferon-γ-induced medulloblastoma. J. Neurosci. Res. 2011, 89, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hendershot, L.M. The role of unfolded protein response in tumor development: Friend or foe? Nat. Rev. Cancer 2004, 4, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, A.C.; Zhang, L.; Adam, A.P.; Aguirre-Ghiso, J.A. Functional coupling of p38-induced upregulation of BiP and activation of RNA-dependent protein kinase-like endoplasmic reticulum kinase to drug resistance of dormant carcinoma cells. Cancer Res. 2006, 66, 1702–1711. [Google Scholar] [CrossRef] [PubMed]
- Rouschop, K.M.; van den Beucken, T.; Dubois, L.; Niessen, H.; Bussink, J.; Savelkouls, K.; Keulers, T.; Mujcic, H.; Landuyt, W.; Voncken, J.W.; et al. The unfolded protein response protects human tumor cells during hypoxia through regulation of the autophagy genes MAP1LC3B and ATG5. J. Clin. Investig. 2010, 120, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ramirez, L.; Cao, H.; Nelson, D.; Hammond, E.; Lee, A.H.; Yoshida, H.; Mori, K.; Glimcher, L.H.; Denko, N.C.; Giaccia, A.J.; et al. XBP1 is essential for survival under hypoxic conditions and is required for tumor growth. Cancer Res. 2004, 64, 5943–5947. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Asano, T.; Isono, M.; Ito, K.; Asano, T. Panobinostat synergizes with bortezomib to induce endoplasmic reticulum stress and ubiquitinated protein accumulation in renal cancer cells. BMC Urol. 2014, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tsai, Y.H.; Tseng, S.H. RECK regulated endoplasmic reticulum stress response and enhanced cisplatin-induced cell death in neuroblastoma cells. Surgery 2013, 154, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Hanke, N.T.; Garland, L.L.; Baker, A.F. Carfilzomib combined with suberanilohydroxamic acid (SAHA) synergistically promotes endoplasmic reticulum stress in non-small cell lung cancer cell lines. J. Cancer Res. Clin. Oncol. 2016, 142, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, P.; Dong, D.; Fu, Y.; Lee, A.S. Transcriptional induction of GRP78/BiP by histone deacetylase inhibitors and resistance to histone deacetylase inhibitor-induced apoptosis. Mol. Cancer Ther. 2009, 8, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Fiskus, W.; Ganguly, S.; Kambhampati, S.; Bhalla, K.N. HDAC inhibitors and chaperone function. Adv. Cancer Res. 2012, 116, 239–262. [Google Scholar] [PubMed]
- Chen, Y.C.; Huang, W.J.; Hsu, J.L.; Yu, C.C.; Wang, W.T.; Guh, J.H. A novel hydroxysuberamide derivative potentiates MG132-mediated anticancer activity against human hormone refractory prostate cancers—The role of histone deacetylase and endoplasmic reticulum stress. Prostate 2013, 73, 1270–1280. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.W.; Yeh, Y.L.; Wang, Y.C.; Huang, W.J.; Chen, Y.A.; Chiou, Y.S.; Ho, S.Y.; Lin, P.; Wang, Y.J. Suberoylanilide hydroxamic acid, an inhibitor of histone deacetylase, enhances radiosensitivity and suppresses lung metastasis in breast cancer in vitro and in vivo. PLoS ONE 2013, 8, e76340. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Lee, J.Y.; Choi, A.Y.; Hwang, K.Y.; Choe, W.; Yoon, K.S.; Ha, J.; Yeo, E.J.; Kang, I. Apicidin induces endoplasmic reticulum stress- and mitochondrial dysfunction-associated apoptosis via phospholipase Cγ1- and Ca2+-dependent pathway in mouse Neuro-2a neuroblastoma cells. Apoptosis 2012, 17, 1340–1358. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.F.; Chiang, A.K. Combination of proteasome and class I HDAC inhibitors induces apoptosis of NPC cells through an HDAC6-independent ER stress-induced mechanism. Int. J. Cancer 2014, 135, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Suzuki, R.; Ohguchi, H.; Yoshida, Y.; Lu, D.; Cottini, F.; Jakubikova, J.; Bianchi, G.; Harada, T.; Gorgun, G.; et al. Class IIa HDAC inhibition enhances ER stress-mediated cell death in multiple myeloma. Leukemia 2015, 29, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Park, K.C.; Jeon, J.Y.; Kim, B.W.; Kim, H.K.; Chang, H.J.; Choi, S.H.; Park, C.S.; Chang, H.S. Potential anti-cancer effect of N-hydroxy-7-(2-naphthylthio) heptanomide (HNHA), a novel histone deacetylase inhibitor, for the treatment of thyroid cancer. BMC Cancer 2015, 15, 1003. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Edagawa, M.; Goshima, H.; Inoue, M.; Yagita, H.; Liu, Z.; Kitajima, S. Role of ATF3 in synergistic cancer cell killing by a combination of HDAC inhibitors and agonistic anti-DR5 antibody through ER stress in human colon cancer cells. Biochem. Biophys. Res. Commun. 2014, 445, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Nalluri, S.; Kolhe, R.; Yang, Y.; Fiskus, W.; Chen, J.; Ha, K.; Buckley, K.M.; Balusu, R.; Coothankandaswamy, V.; et al. Treatment with panobinostat induces glucose-regulated protein 78 acetylation and endoplasmic reticulum stress in breast cancer cells. Mol. Cancer Ther. 2010, 9, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Endo, M.; Shinohara, F.; Echigo, S.; Rikiishi, H. Enhancement of cisplatin cytotoxicity by SAHA involves endoplasmic reticulum stress-mediated apoptosis in oral squamous cell carcinoma cells. Cancer Chemother. Pharmacol. 2009, 64, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tseng, S.H. The potential of RECK inducers as antitumor agents for glioma. Anticancer Res. 2012, 32, 2991–2998. [Google Scholar] [PubMed]
- Chen, Y.; Tsai, Y.H.; Tseng, S.H. Valproic acid affected the survival and invasiveness of human glioma cells through diverse mechanisms. J. Neurooncol. 2012, 109, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.T.; Chang, H.C.; Chiang, L.C.; Hung, W.C. Histone deacetylase inhibitor up-regulates RECK to inhibit MMP-2 activation and cancer cell invasion. Cancer Res. 2003, 63, 3069–3072. [Google Scholar] [PubMed]
- Clark, J.C.; Thomas, D.M.; Choong, P.F.; Dass, C.R. RECK—A newly discovered inhibitor of metastasis with prognostic significance in multiple forms of cancer. Cancer Metastasis Rev. 2007, 26, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Meng, N.; Li, Y.; Zhang, H.; Sun, X.F. RECK, a novel matrix metalloproteinase regulator. Histol. Histopathol. 2008, 23, 1003–1010. [Google Scholar]
- Noda, M.; Takahashi, C. Recklessness as a hallmark of aggressive cancer. Cancer Sci. 2007, 98, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Silveira Correa, T.C.; Massaro, R.R.; Brohem, C.A.; Taboga, S.R.; Lamers, M.L.; Santos, M.F.; Maria-Engler, S.S. RECK-mediated inhibition of glioma migration and invasion. J. Cell Biochem. 2010, 110, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Silveira Correa, T.C.; Brohem, C.A.; Winnischofer, S.M.B.; da Silva Cardeal, L.B.; Sasahara, R.M.; Taboga, S.R.; Sogayar, M.C.; Maria-Engler, S.S. Down-regulation of the RECK-tumor- and metastasis-suppressor gene in glioma invasiveness. J. Cell Biochem. 2006, 99, 156–167. [Google Scholar]
- Namwat, N.; Puetkasichonpasutha, J.; Loilome, W.; Yongvanit, P.; Techasen, A.; Puapairoj, A.; Sripa, B.; Tassaneeyakul, W.; Khuntikeo, N.; Wongkham, S. Downregulation of reversion-inducing-cysteine-rich protein with Kazal motifs (RECK) is associated with enhanced expression of matrix metalloproteinases and cholangiocarcinoma metastases. J. Gastroenterol. 2011, 46, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Nagini, S. RECKing MMP: Relevance of reversion-inducing cysteine-rich protein with Kazal motifs as a prognostic marker and therapeutic target for cancer (a review). Anticancer Agents Med. Chem. 2012, 12, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Liu, L.T.; Hung, W.C. Involvement of histone deacetylase in RAS-induced down-regulation of the metastasis suppressor RECK. Cell Signal 2004, 16, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Cho, C.Y.; Hung, W.C. Silencing of the metastasis suppressor RECK by RAS oncogene is mediated by DNA methyltransferase 3b-induced promoter methylation. Cancer Res. 2006, 66, 8413–8420. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.C.; Chang, H.C.; Hung, W.C. HER-2/neu represses the metastasis suppressor RECK via ERK and Sp transcription factors to promote cell invasion. J. Biol. Chem. 2006, 281, 4718–4725. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Lee, K.Y.; Lee, Y.M. Downregulation of a tumor suppressor RECK by hypoxia through recruitment of HDAC1 and HIF-1α to reverse HRE site in the promoter. Biochim. Biophys. Acta 2010, 1803, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.H.; Tseng, Y.F.; Hsu, Y.C.; Lan, S.H.; Wu, S.Y.; Raghavaraju, G.; Cheng, D.E.; Lee, Y.R.; Chang, T.Y.; Chow, N.H.; et al. Ras induces experimental lung metastasis through up-regulation of RbAp46 to suppress RECK promoter activity. BMC Cancer 2015, 15, 172. [Google Scholar] [CrossRef] [PubMed]
- Somanna, N.K.; Valente, A.J.; Krenz, M.; McDonald, K.S.; Higashi, Y.; Noda, M.; Chandrasekar, B. Histone deacetyltransferase inhibitors Trichostatin A and Mocetinostat differentially regulate MMP9, IL-18 and RECK expression, and attenuate Angiotensin II-induced cardiac fibroblast migration and proliferation. Hypertens. Res. 2016, 39, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.W.; Lee, Y.M. Inhibition of histone deacetylase attenuates hypoxia-induced migration and invasion of cancer cells via the restoration of RECK expression. Mol. Cancer Ther. 2010, 9, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Berezin, V.; Walmod, P.S.; Filippov, M.; Dityatev, A. Targeting of ECM molecules and their metabolizing enzymes and receptors for the treatment of CNS diseases. Prog. Brain Res. 2014, 214, 353–388. [Google Scholar] [PubMed]
- Ahn, M.Y.; Kang, D.O.; Na, Y.J.; Yoon, S.; Choi, W.S.; Kang, K.W.; Chung, H.Y.; Jung, J.H.; Kim, H.S. Histone deacetylase inhibitor, apicidin, inhibits human ovarian cancer cell migration via class II histone deacetylase 4 silencing. Cancer Lett. 2012, 325, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, L.; Kang, S.G.; Lu, Y.; Yang, Z.; Huynh, T.; Chen, C.; Zhou, R.; Guo, M.; Zhao, Y. Gd-metallofullerenol nanomaterial suppresses pancreatic cancer metastasis by inhibiting the interaction of histone deacetylase 1 and metastasis-associated protein 1. ACS Nano 2015, 9, 6826–6836. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Dong, D.S.; Pei, L. Synergistic antitumor activity of resveratrol and miR-200c in human lung cancer. Oncol. Rep. 2014, 31, 2293–2297. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Histone Deacetylase Inhibitors |
|---|
| Apicidin |
| Gd-metallofullerenol nanomaterial |
| MS-275 |
| M344 |
| N-hydroxy-7-(2-naphthylthio) heptanomide (HNHA) |
| Panobinostat |
| Romidepsin |
| S-7-oxo-7-(4-phenylthiazol-2-ylamino)-heptyl) 2-methylpropanethioate (PTACH) |
| Sodium butyrate |
| Suberoylanilide hydroxamic acid (SAHA) |
| TMP269 |
| trans-3,4,5-trihydroxystilbene (resveratrol) |
| Trichostatin A (TSA) |
| Valproic acid (VPA) |
| WJ25591 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Tsai, Y.-H.; Tseng, S.-H. HDAC Inhibitors and RECK Modulate Endoplasmic Reticulum Stress in Tumor Cells. Int. J. Mol. Sci. 2017, 18, 258. https://doi.org/10.3390/ijms18020258
Chen Y, Tsai Y-H, Tseng S-H. HDAC Inhibitors and RECK Modulate Endoplasmic Reticulum Stress in Tumor Cells. International Journal of Molecular Sciences. 2017; 18(2):258. https://doi.org/10.3390/ijms18020258
Chicago/Turabian StyleChen, Yun, Ya-Hui Tsai, and Sheng-Hong Tseng. 2017. "HDAC Inhibitors and RECK Modulate Endoplasmic Reticulum Stress in Tumor Cells" International Journal of Molecular Sciences 18, no. 2: 258. https://doi.org/10.3390/ijms18020258