
Ixeris dentata (Thunb. Ex Thunb.) Nakai Extract Inhibits Proliferation and Induces Apoptosis in Breast Cancer Cells through Akt/NF-κB Pathways


Abstract
:1. Introduction
2. Results
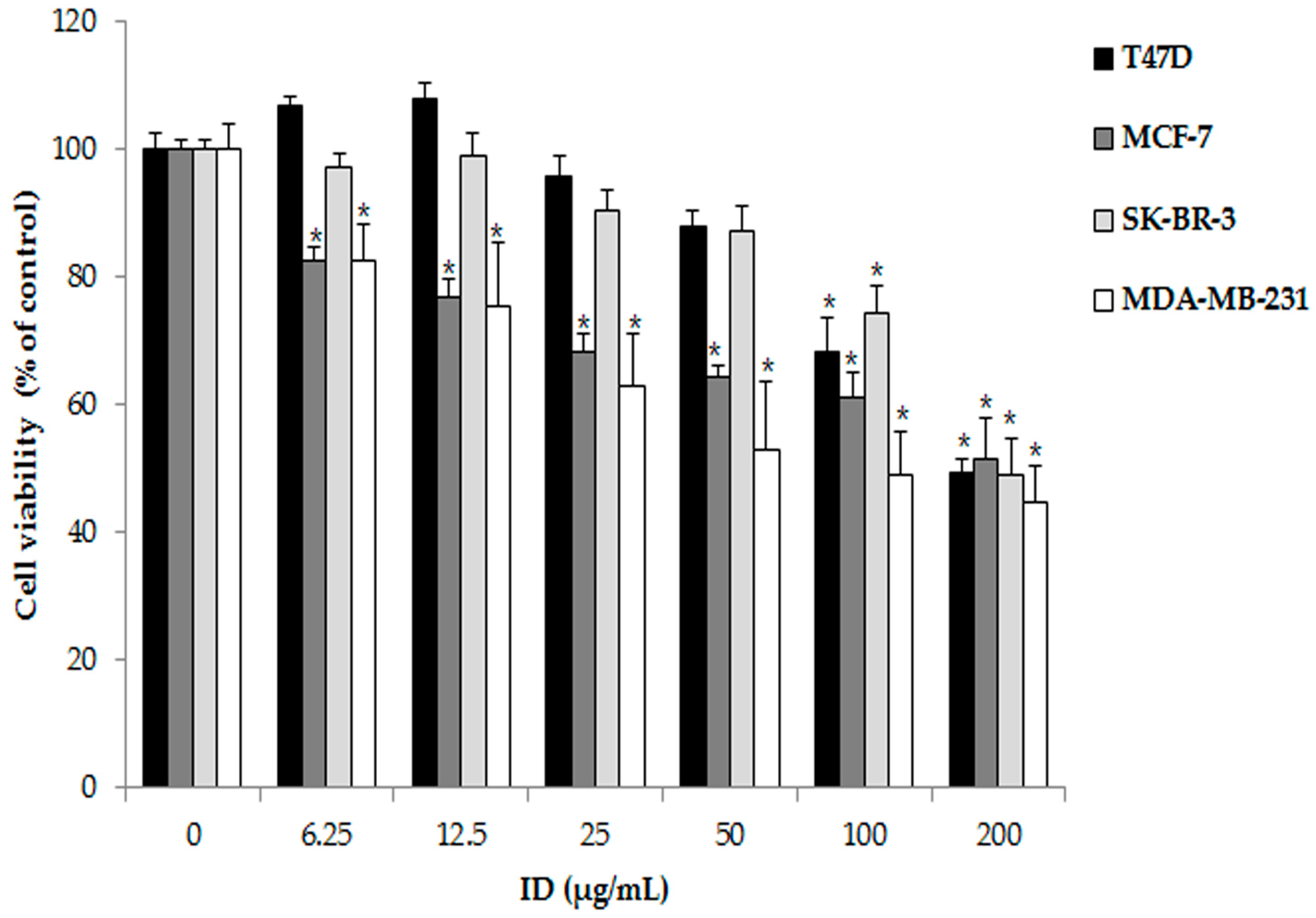
2.1. Effects of Ixeris dentata (ID) Extract on Survival Rate Inhibition in T47D, MCF-7, SK-BR-3, and MDA-MB-231 Cells
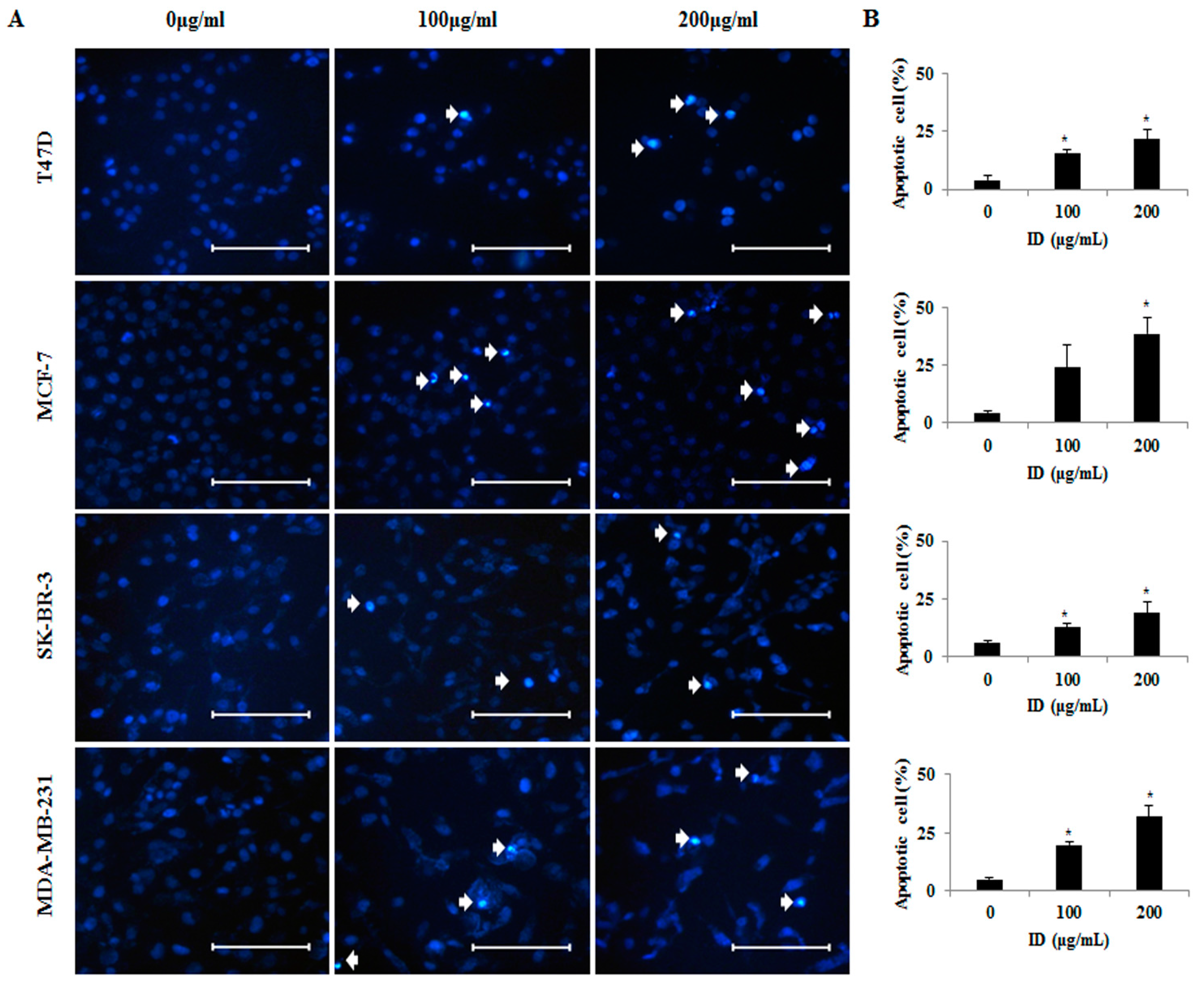
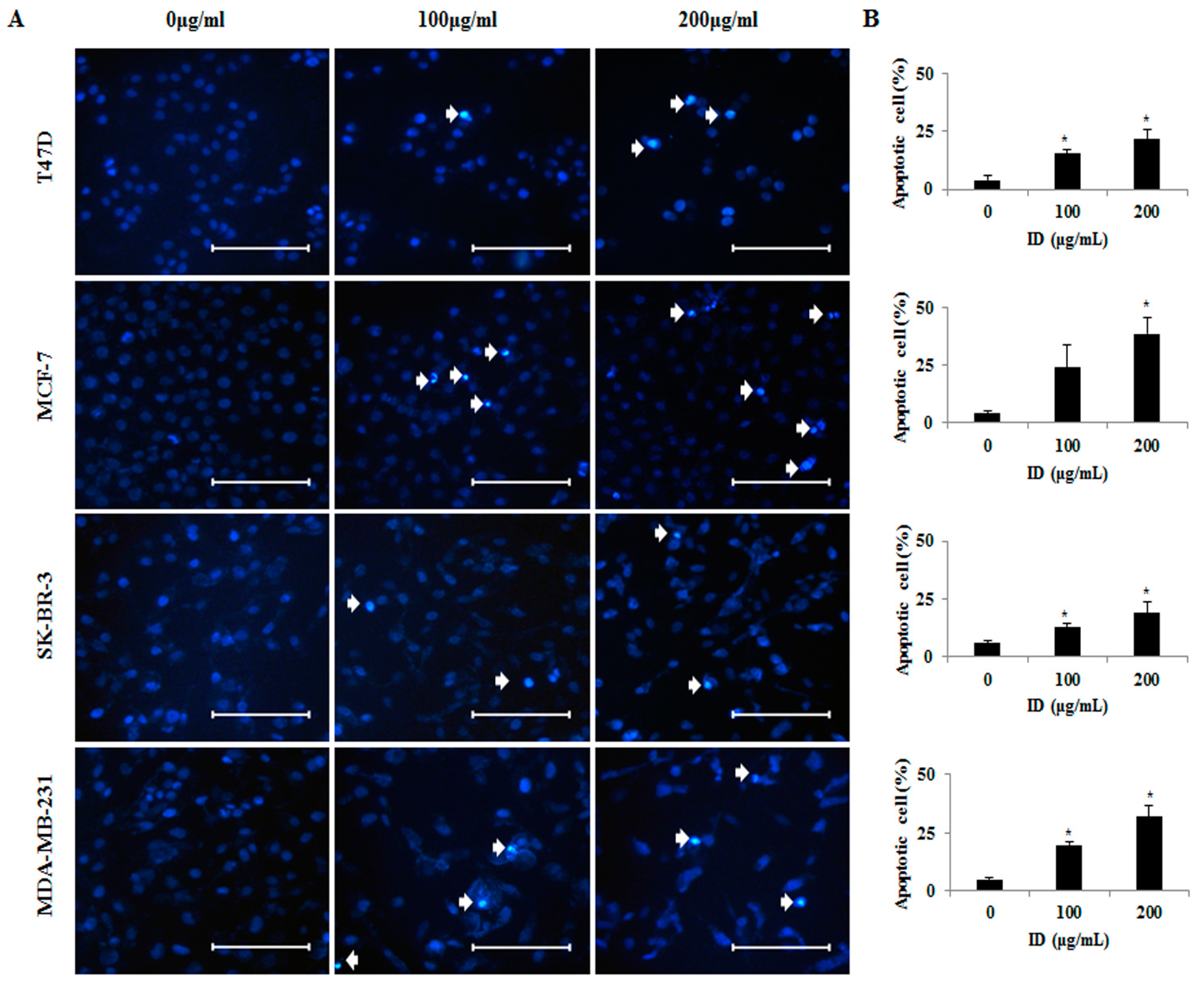
2.2. Effects of ID Extract on Morphological Changes in T47D, MCF-7, SK-BR-3, and MDA-MB-231 Cells
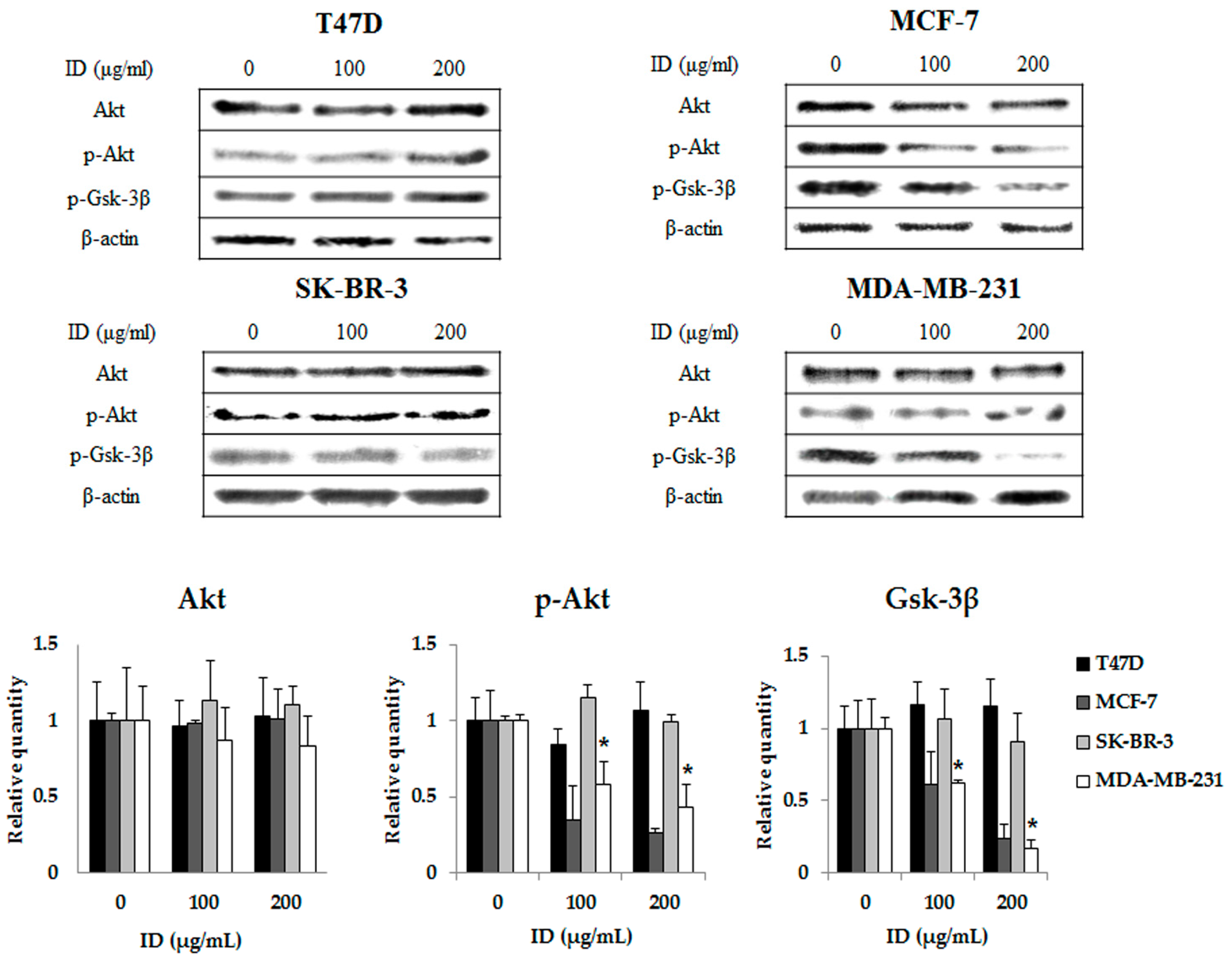
2.3. Inhibitory Effects of ID Extract on Akt Kinase Activation
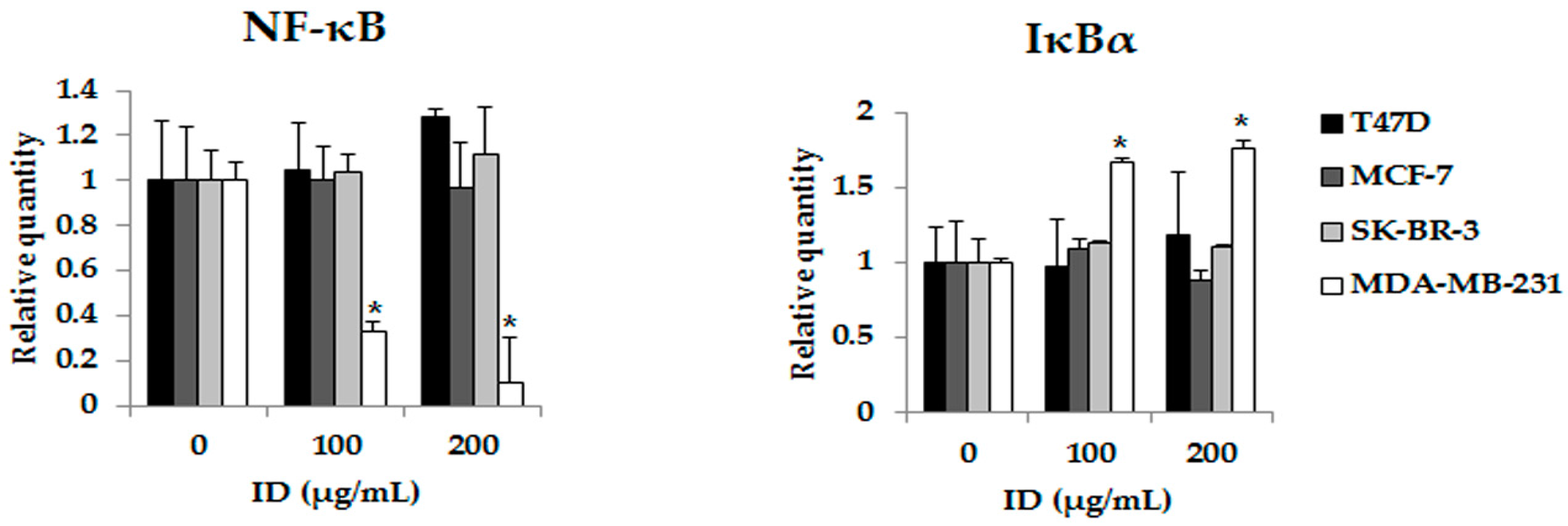
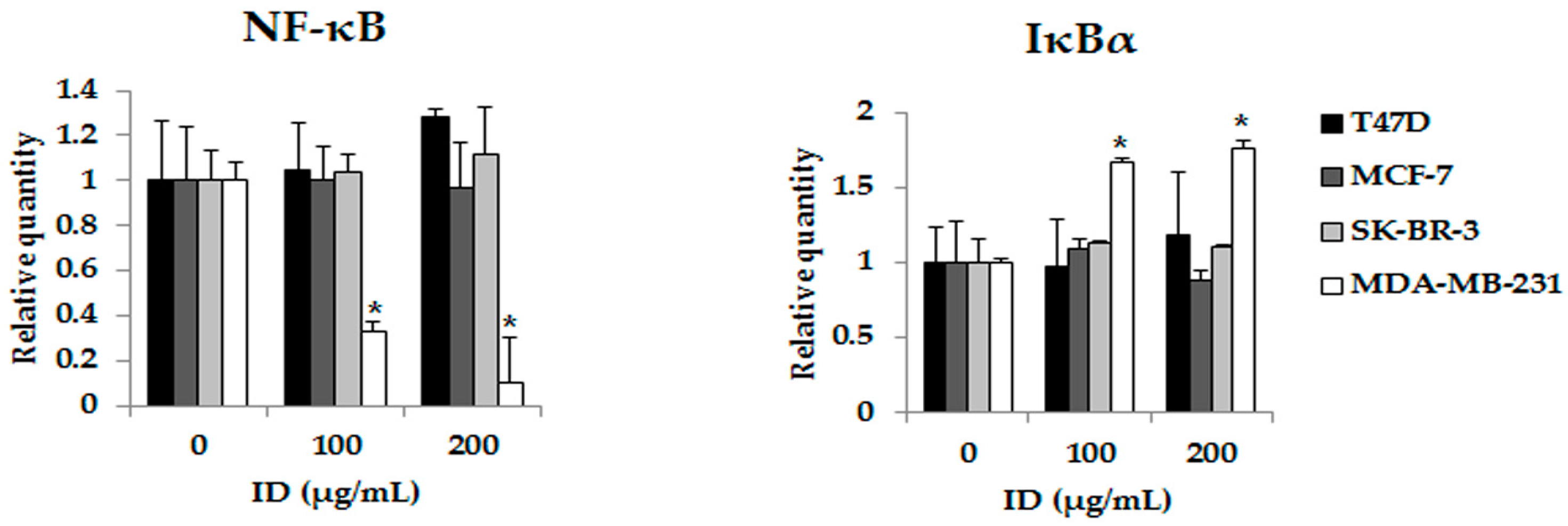
2.4. Inhibitory Effects of ID Extract on NF-κB Kinase Activation
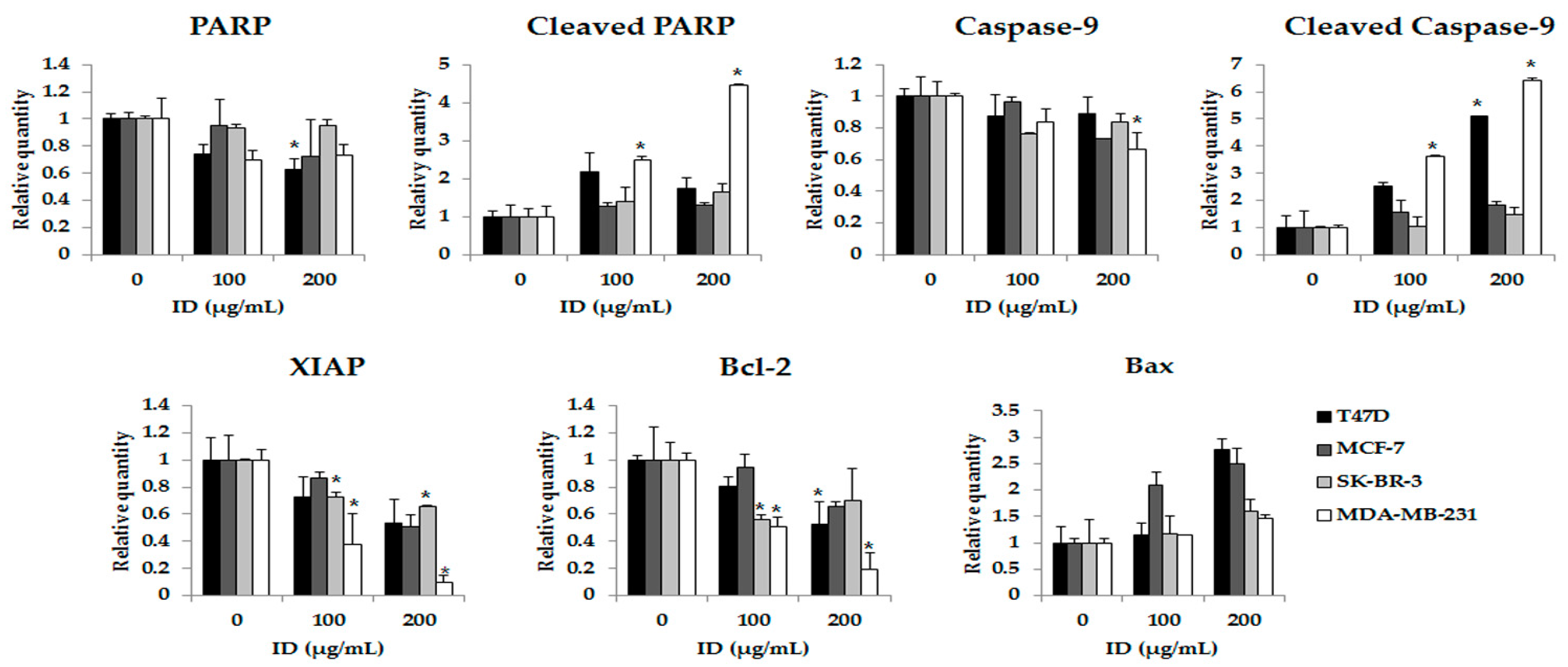
2.5. Effects of ID Extract on the Expression of Apoptosis Pathway Proteins
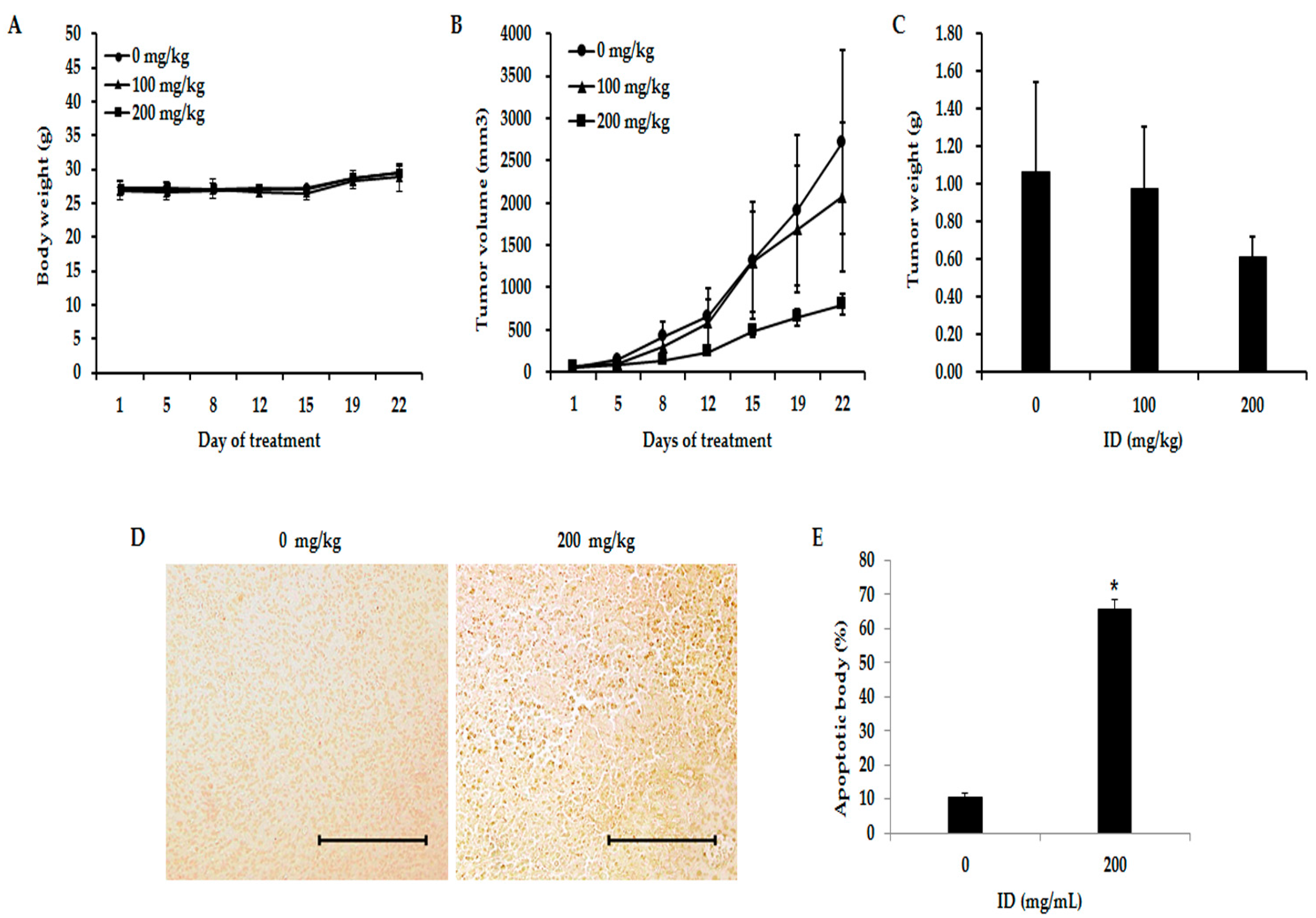
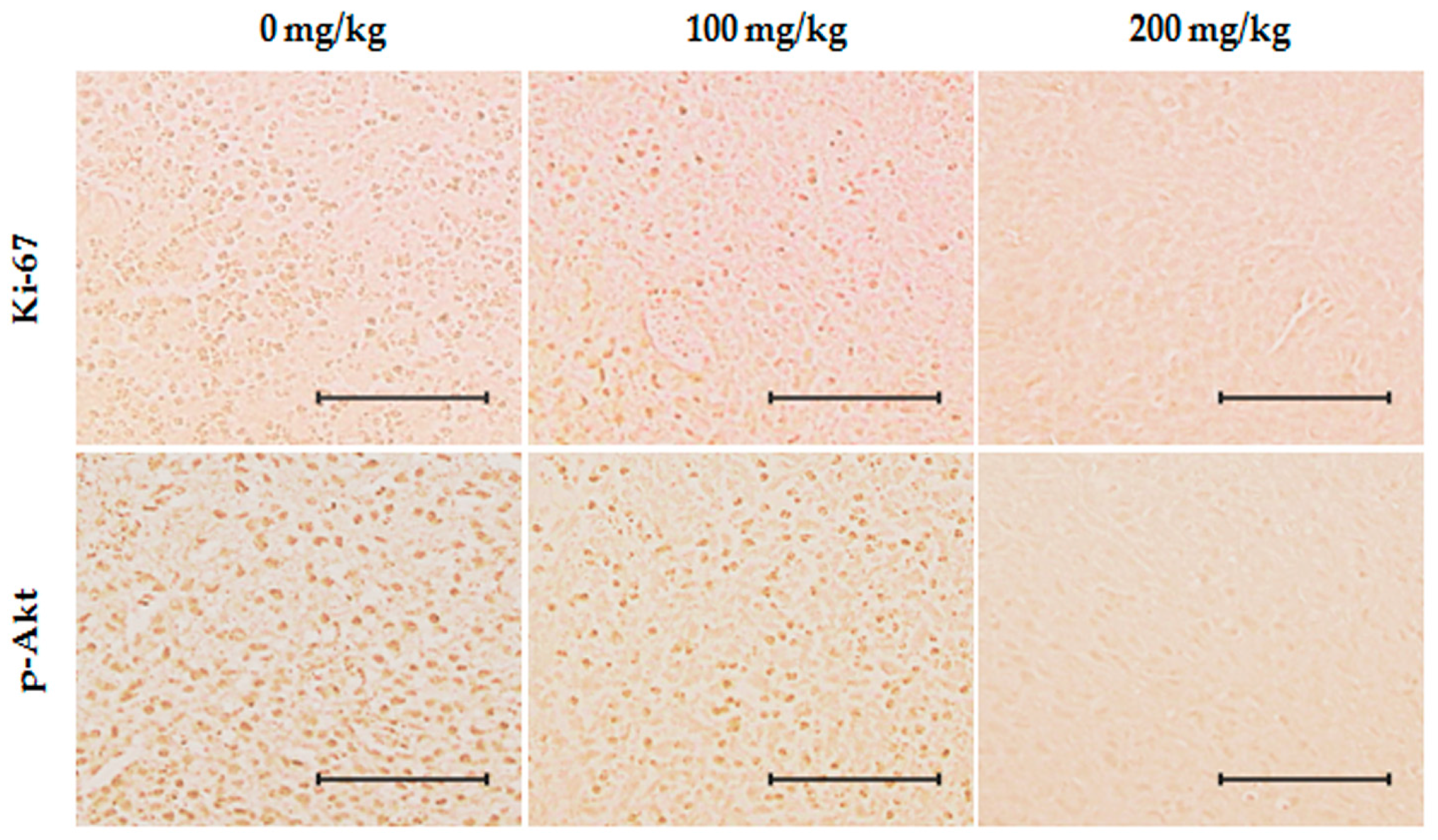
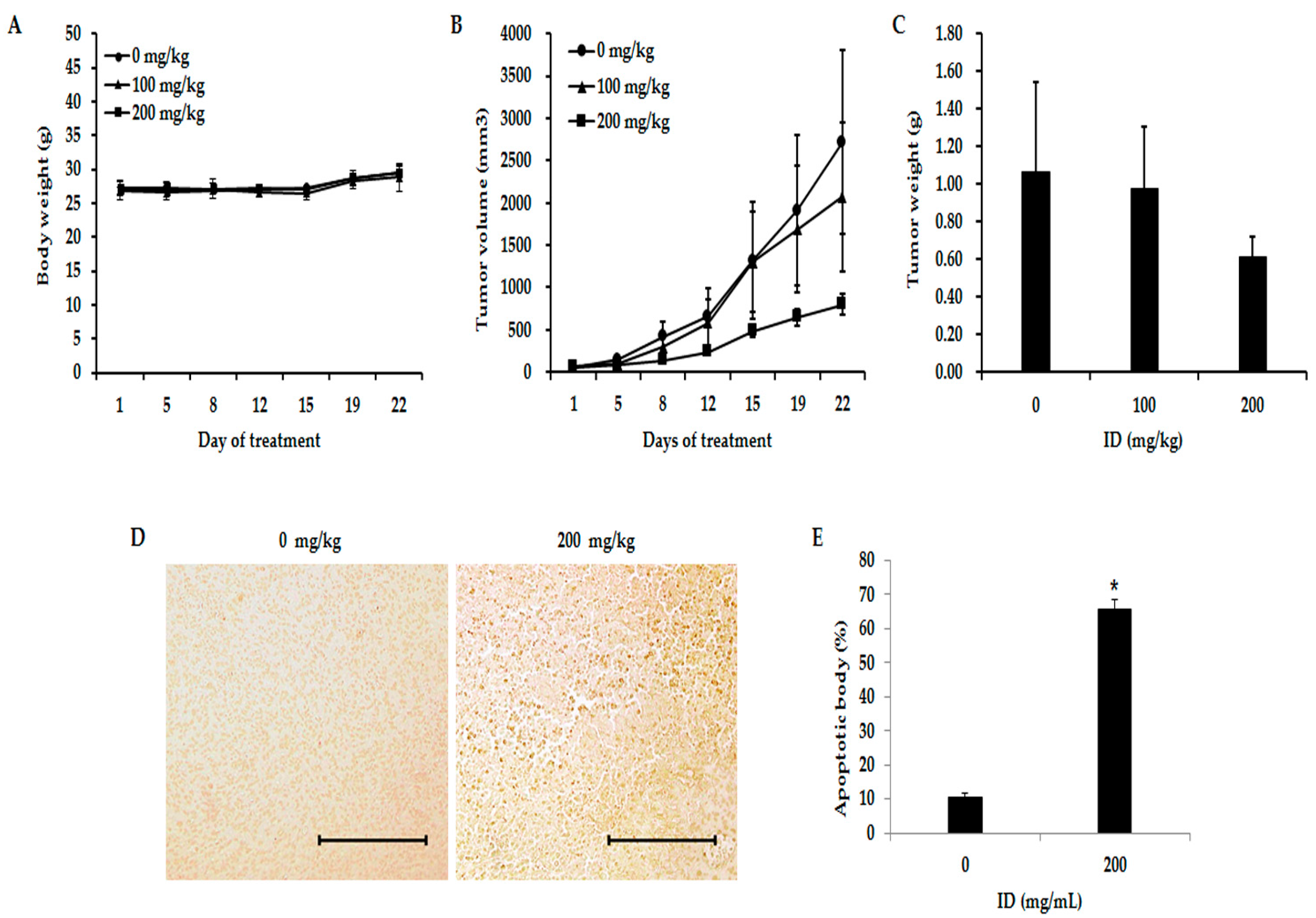
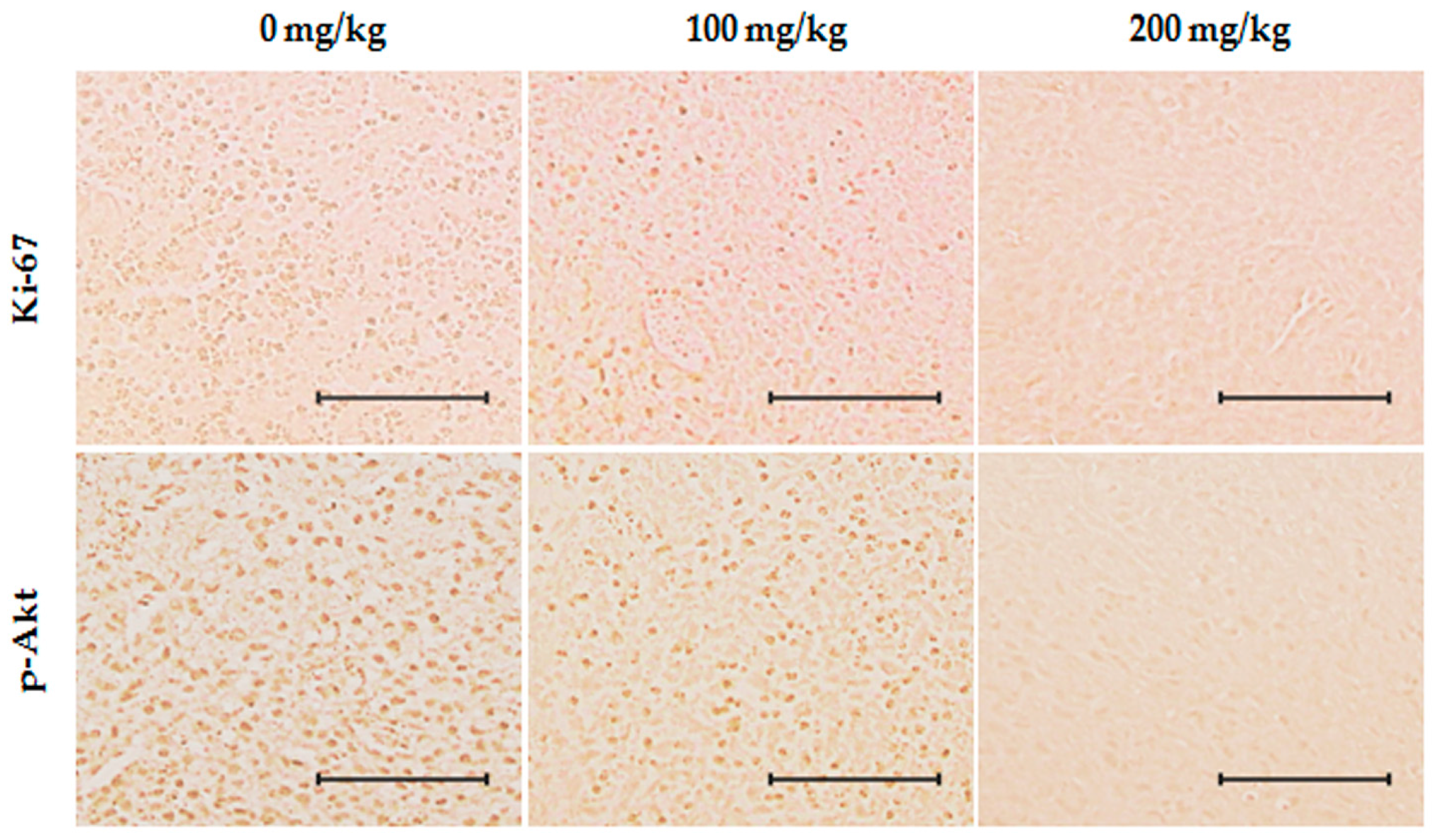
2.6. Inhibition of Effects of ID Extract on Tumor Growth in Nude Mice
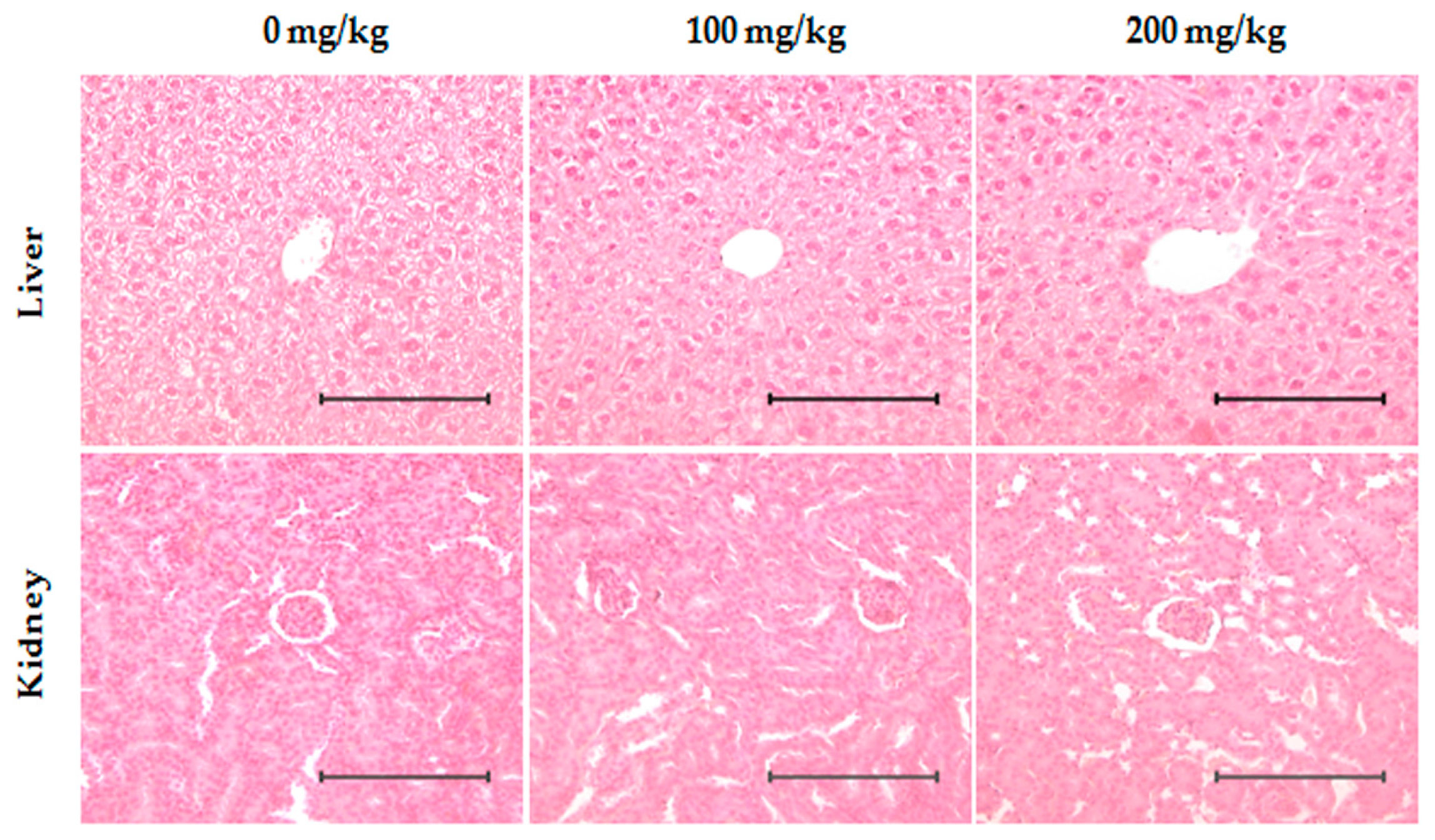
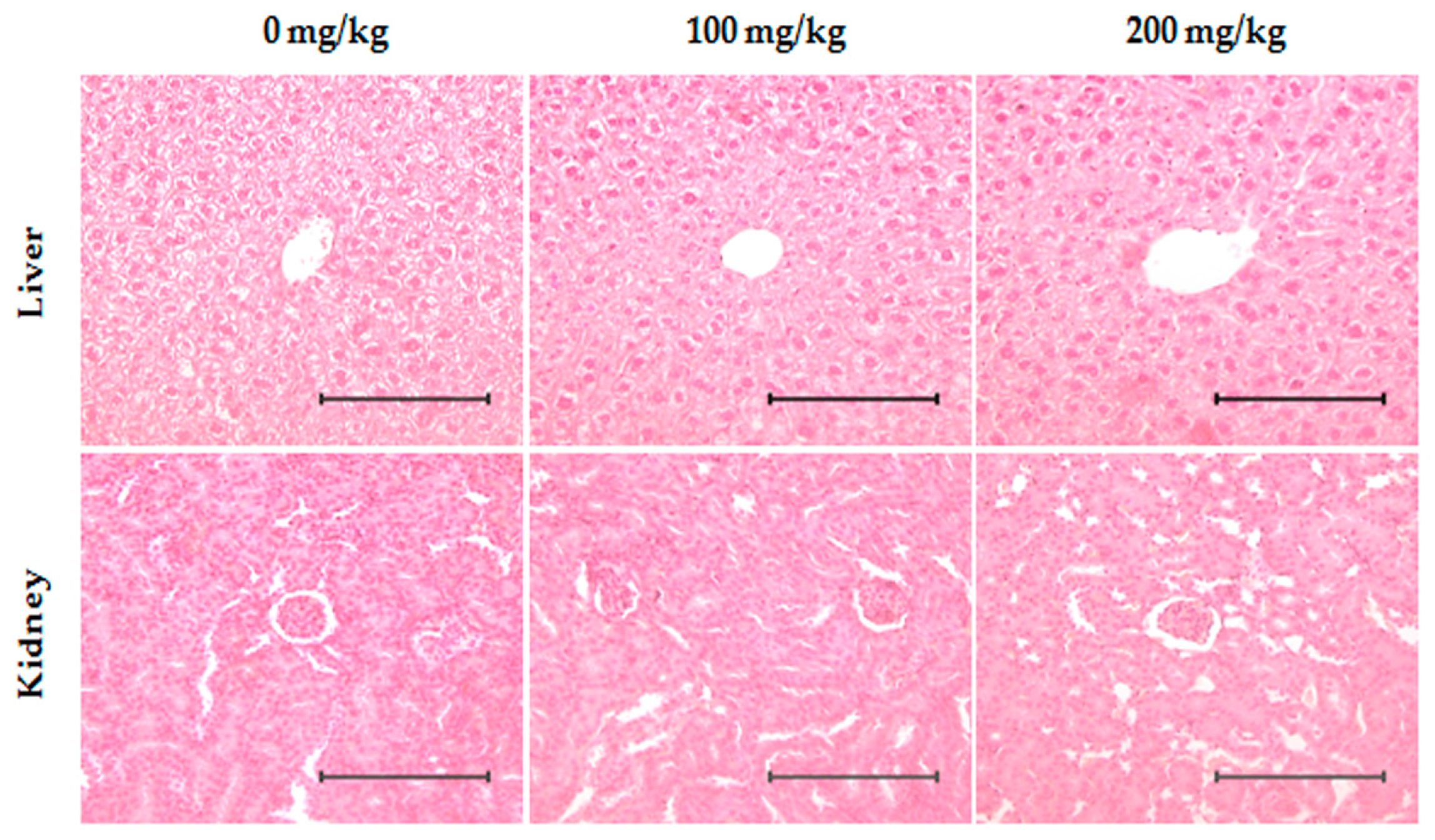
2.7. Toxicity Evaluation of ID Extract in Liver and Kidney Tissues
3. Discussion
4. Materials and Methods
4.1. Chemicals, Drugs, and Antibodies
4.2. Preparation of Ixeris dentata (Thunb. Ex Thunb.) Nakai (ID) Extract
4.3. Cell Line and Culture
4.4. Cell Viability Assay
4.5. Nuclear Morphology
4.6. Western Blot Analysis
4.7. Animal and Xenograft
4.8. TUNEL Assay
4.9. Immunohistochemistry
4.10. Histological Examination
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| DAB | 3′-Diaminobenzidine tetrahydrochloride |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| ECL | Enhanced chemiluminescence |
| ER | Estrogen receptor |
| Gsk-3β | Glycogen synthase kinase-3β |
| HER2 | Human epidermal growth factor receptor 2 |
| HRP | Horseradish peroxidase |
| ID | Ixeris dentata (Thunb. Ex Thunb.) Nakai |
| IKK | IκB kinase |
| IκBs | Inhibitors of NF-κB |
| NF-κB | Nuclear factor-κB |
| PARP | Poly(ADP-ribose)polymerase |
| PR | Progesterone receptor |
| TBS | Tris-buffered saline |
| XIAP | X chromosome-linked inhibitor of apoptosis |
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, J.L.; Berkowitz, G.S. Breast cancer epidemiology. Cancer Res. 1988, 48, 5615–5623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, X.; Huang, Z.; Xu, W.; Liu, P. An ER-associated miRNA signature predicts prognosis in ER-positive breast cancer. J. Exp. Clin. Cancer Res. 2014, 33, 94. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Hung, M.H.; Wang, D.S.; Chu, P.Y.; Su, J.C.; Teng, T.H.; Huang, C.T.; Chao, T.T.; Wang, C.T.; Shiau, C.W.; et al. Tamoxifen induces apoptosis through cancerous inhibitor of protein phosphatase 2A–dependent phospho-Akt inactivation in estrogen receptor–negative human breast cancer cells. Breast Cancer Res. 2014, 16, 431. [Google Scholar] [CrossRef] [PubMed]
- Kennecke, H.; Yerushalmi, R.; Woods, R.; Cheang, M.C.U.; Voduc, D.; Speers, C.H.; Nielsen, T.O.; Gelmon, K. Metastatic behavior of breast cancer subtypes. J. Clin. Oncol. 2010, 28, 3271–3277. [Google Scholar] [CrossRef] [PubMed]
- Park, B.Y.; Choi, K.S.; Lee, Y.Y.; Jun, J.K.; Seo, H.G. Cancer screening status in Korea, 2011: Results from the Korean National Cancer Screening Survey. Asian Pac. J. Cancer Prev. 2012, 13, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.M.; Hong, S.H.; Lee, H.J.; Won, J.H.; Kim, J.M.; Jeong, D.M.; Baek, S.H.; Lim, J.P.; Kim, H.M. Ixeris dentata green sap inhibits both compound 48/80-induced aanaphylaxis-like response and IgE-mediated anaphylactic response in murine model. Biol. Pharm. Bull. 2002, 25, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Kusumoto, Y.; Nagao, M.; Shiojima, K.; Ageta, H. Composite constituents: Aliphatics and triterpenoids isolated from the whole plants of Ixeris debilis and I. dentata. Yakugaku Zasshi J. Pharm. Soc. Jpn. 1983, 103, 356–359. [Google Scholar]
- Seto, M.; Miyase, T.; Fukushima, S. Sesquiterpene lactones from Ixeris dentata Nakai. Chem. Pharm. Bull. 1986, 34, 4170–4176. [Google Scholar] [CrossRef]
- Choi, J.S.; Young, H.S.; Kim, B.W. Hypoglycemic and hypolipemic effects of Ixeris dentata in diabetic rats. Arch. Pharm. Res. 1990, 13, 269–273. [Google Scholar] [CrossRef]
- Oh, S.H.; Sung, T.H.; Kim, M.R. Ixeris dentata extract maintains glutathione concentrations in mouse brain tissue under oxidative stress induced by kainic acid. J. Med. Food 2003, 6, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H. Inhibitory effects of Ixeris dentata on the mutagenicity of aflatoxin B1, N-methyl-N′-nitro-N-nitrosoguanidine and the growth of MG-63 human osteosarcoma cells. J. Korean Soc. Food Nutr. 1995, 24, 305–312. [Google Scholar]
- Lim, S.S.; Lee, J.H. Effect of Aster scaber and Ixeris dentata on contractility and vasodilation of cardiovascula and endothelial cell in hyperlipidemic rat. J. Korean Soc. Food Sci. Nutr. 1997, 26, 300–307. [Google Scholar]
- Lee, E. Effects of Ixeris dentata extract on the production of pro-inflammatory cytokines in the LPS stimulated rat and Raw 264.7 cells. Korean J. Plant Resour. 2011, 24, 604–612. [Google Scholar] [CrossRef]
- Park, E.K.; Sung, J.H.; Trinh, H.T.; Bea, E.A.; Yun, H.K.; Hong, S.S.; Kim, D.H. Lactic acid bacterial fermentation increases the antiallergic effects of Ixeris dentata. J. Microbiol. Biotechnol. 2008, 18, 308–313. [Google Scholar] [PubMed]
- Kim, M.J.; Kim, J.S.; Kang, W.H.; Jeong, D.M. Effect on antimutagenic and cancer cell growth inhibition of Ixeris dentata Nakai. Korean J. Med. Crop Sci. 2002, 10, 139–143. [Google Scholar]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Brazil, D.P.; Hemmings, B.A. Ten years of protein kinase B signalling: A hard Akt to follow. Trends Biochem. Sci. 2001, 26, 657–664. [Google Scholar] [CrossRef]
- Sun, L.; Li, W.; Li, W.; Xiong, L.; Li, G.; Ma, R. Astragaloside IV prevents damage to human mesangial cells through the inhibition of the NADPH oxidase/ROS/Akt/NF-κB pathway under high glucose conditions. Int. J. Mol. Med. 2014, 34, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Harhaj, E.W.; Sun, S.C. NF-κB-inducing kinase regulates the processing of NF-κB2 p100. Mol. Cell 2001, 7, 401–409. [Google Scholar] [CrossRef]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-κB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-κB and Rel proteins: Evolutionarily conserved mediators of immune responses. Annu. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Hu, X.Y.; Lu, T.; Cheng, Y.N.; Young, C.Y.F.; Yuan, H.Q.; Lou, H.X. Retigeric acid B exhibits antitumor activity through suppression of nuclear factor-κB signaling in prostate cancer cells in vitro and in vivo. PLoS ONE 2012, 7, e38000. [Google Scholar] [CrossRef] [PubMed]
- Han, S.S.; Yun, H.K.; Son, D.J.; Tompkins, V.S.; Peng, L.; Chung, S.T.; Kim, S.K.; Park, E.S.; Janz, S. NF-κB/STAT3/PI3K signaling crosstalk in iMycEμ B lymphoma. Mol. Cancer 2010, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Pecorino, L. Molecular Biology of Cancer: Mechanisms, Targets, and Therapeutics; Oxford University Press: Oxford, UK, 2012; pp. 124–149. [Google Scholar]
- Nizamutdinova, I.T.; Lee, G.W.; Son, K.H.; Jeon, S.J.; Kang, S.S.; Kim, Y.S.; Lee, J.H.; Seo, H.G.; Chang, K.C.; Kim, H.J. Tanshinone I effectively induces apoptosis in estrogen receptor-positive (MCF-7) and estrogen receptor-negative (MDA-MB-231) breast cancer cells. Int. J. Oncol. 2008, 33, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Wang, H.S.; Liu, N.; Ge, L.C. Bisphenol A stimulates the epithelial mesenchymal transition of estrogen negative breast cancer cells via FOXA1 signals. Arch. Biochem. Biophys. 2015, 585, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Heo, B.G.; Chon, S.U.; Park, Y.J.; Bae, J.H.; Park, S.M.; Park, Y.S.; Jang, H.G.; Gorinstein, S. Antiproliferative activity of Korean wild vegetables on different human tumor cell lines. Plant Foods Hum. Nutr. 2009, 64, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.W.; Liu, C.H.; Du, C.M.; Zhang, J.; Wu, W.Q.; Gu, Z.L. Induction of apoptosis in human hepatocarcinoma SMMC-7721 cells in vitro by flavonoids from Astragalus complanatus. J. Ethnopharmacol. 2009, 123, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, J.S.; Jeong, D.M.; Ham, S.S.; Yu, C.Y. Effect of antioxidant, antimutagenicity and anticancer of root extract from Ixeris dentata Nakai. Korean J. Med. Crop Sci. 2002, 10, 222–229. [Google Scholar]
- Kim, H.J.; Kang, K.J. Effects of Ixeris dentata Extract on Radical Oxygen Species and Bcl-2 Family in Human Breast Cancer Cells. J. East Asian Soc. Diet. Life 2014, 24, 739–747. [Google Scholar] [CrossRef]
- Vara, J.Á.F.; Casado, E.; de Castro, E.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.R.; Ahmed, S.O.; Ahmed, M.; Kham, O.S.; Al AbdulMohsen, S.; Platanias, L.C.; Al-Kuraya, K.S.; Uddin, S. Cross-talk between NF-κB and the PI3-kinase/AKT pathway can be targeted in primary effusion lymphoma (PEL) cell lines for efficient apoptosis. PLoS ONE 2012, 7, e39945. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; He, X.; Xia, W.; Hsu, J.M.; Chen, C.T.; Li, L.Y.; Lee, D.F.; Yang, J.Y.; Xie, X.; Liu, J.C.; et al. Myeloid cell leukemia-1 inversely correlates with glycogen synthase kinase-3β activity and associates with poor prognosis in human breast cancer. Cancer Res. 2007, 67, 4564–4571. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Guo, W.; Liang, X.; Rao, Y. Both the establishment and the maintenance of neuronal polarity require active mechanisms: Critical roles of GSK-3β and its upstream regulators. Cell 2005, 120, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Debatin, K.M. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immunother. 2004, 53, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Lin, A. NF-κB at the crossroads of life and death. Nat. Immunol. 2002, 3, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, J.; Lemke, H.; Baisch, H.; Wacker, H.H.; Schwab, U.; Stein, H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J. Immunol. 1984, 133, 1710–1715. [Google Scholar] [PubMed]
- Inwald, E.C.; Klinkhammer-Schalke, M.; Hofstädter, F.; Zeman, F.; Koller, M.; Gerstenhauer, M.; Ortamann, O. Ki-67 is a prognostic parameter in breast cancer patients: Results of a large population-based cohort of a cancer registry. Breast Cancer Res. Treat. 2013, 139, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Gong, F.Y.; Wu, X.X.; Sun, Y.; Li, Y.H.; Chen, T.; Xu, Q. Anti-inflammatory and immunosuppressive effect of flavones isolated from Artemisia vestita. J. Ethnopharmacol. 2008, 120, 1–6. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Group | Pre-Experiment | Post-Experiment | Inhibition Rate b (%) |
|---|---|---|---|---|
| Size (mm3) | Size (mm3) | |||
| MDA-MB-231 | 0 mg/kg a | 53 | 2719 | |
| 100 mg/kg | 61 | 2076 | 24 | |
| 200 mg/kg | 65 | 801 | 71 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-A.; Lee, H.-N.; Choo, G.-S.; Kim, H.-J.; Che, J.-H.; Jung, J.-Y. Ixeris dentata (Thunb. Ex Thunb.) Nakai Extract Inhibits Proliferation and Induces Apoptosis in Breast Cancer Cells through Akt/NF-κB Pathways. Int. J. Mol. Sci. 2017, 18, 275. https://doi.org/10.3390/ijms18020275
Shin S-A, Lee H-N, Choo G-S, Kim H-J, Che J-H, Jung J-Y. Ixeris dentata (Thunb. Ex Thunb.) Nakai Extract Inhibits Proliferation and Induces Apoptosis in Breast Cancer Cells through Akt/NF-κB Pathways. International Journal of Molecular Sciences. 2017; 18(2):275. https://doi.org/10.3390/ijms18020275
Chicago/Turabian StyleShin, Seong-Ah, Hae-Nim Lee, Gang-Sik Choo, Hyeong-Jin Kim, Jeong-Hwan Che, and Ji-Youn Jung. 2017. "Ixeris dentata (Thunb. Ex Thunb.) Nakai Extract Inhibits Proliferation and Induces Apoptosis in Breast Cancer Cells through Akt/NF-κB Pathways" International Journal of Molecular Sciences 18, no. 2: 275. https://doi.org/10.3390/ijms18020275