
Permeation of Polymethoxyflavones into the Mouse Brain and Their Effect on MK-801-Induced Locomotive Hyperactivity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
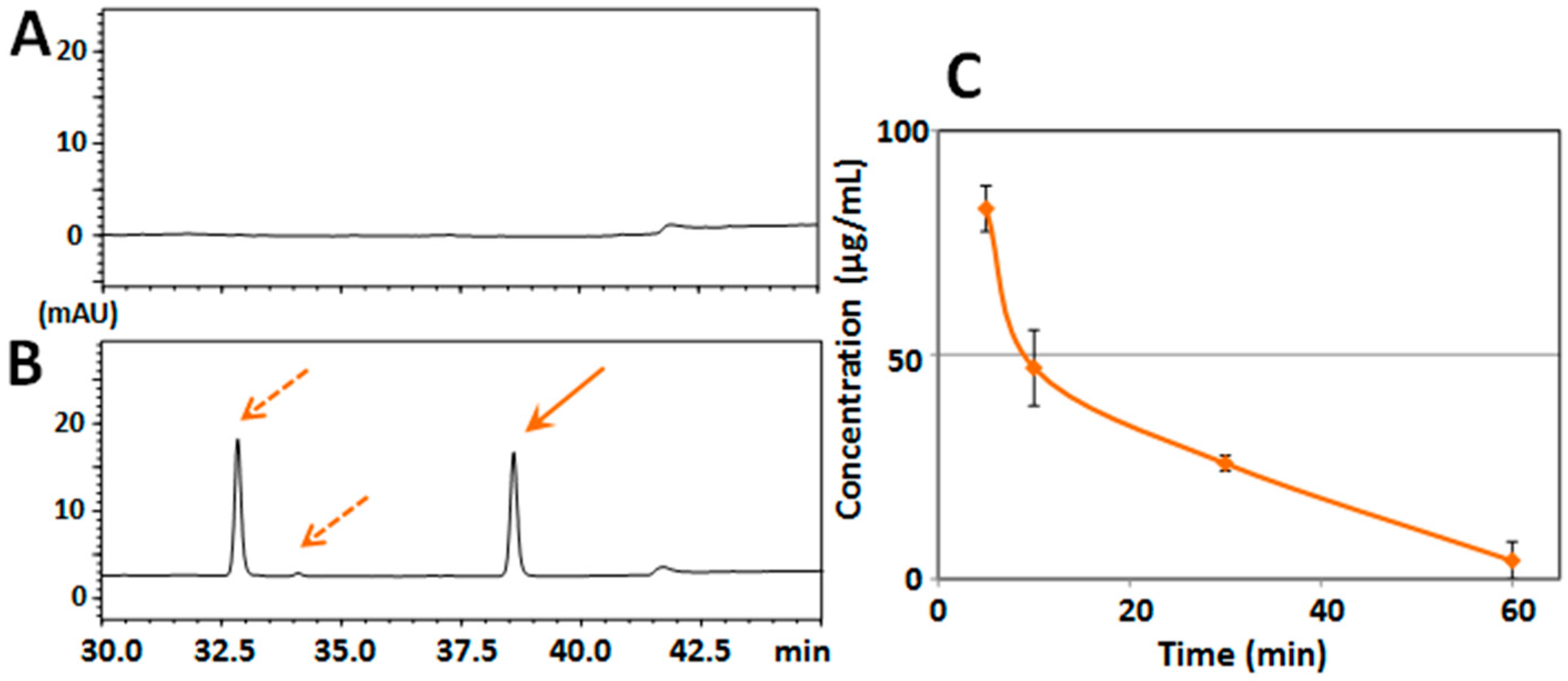
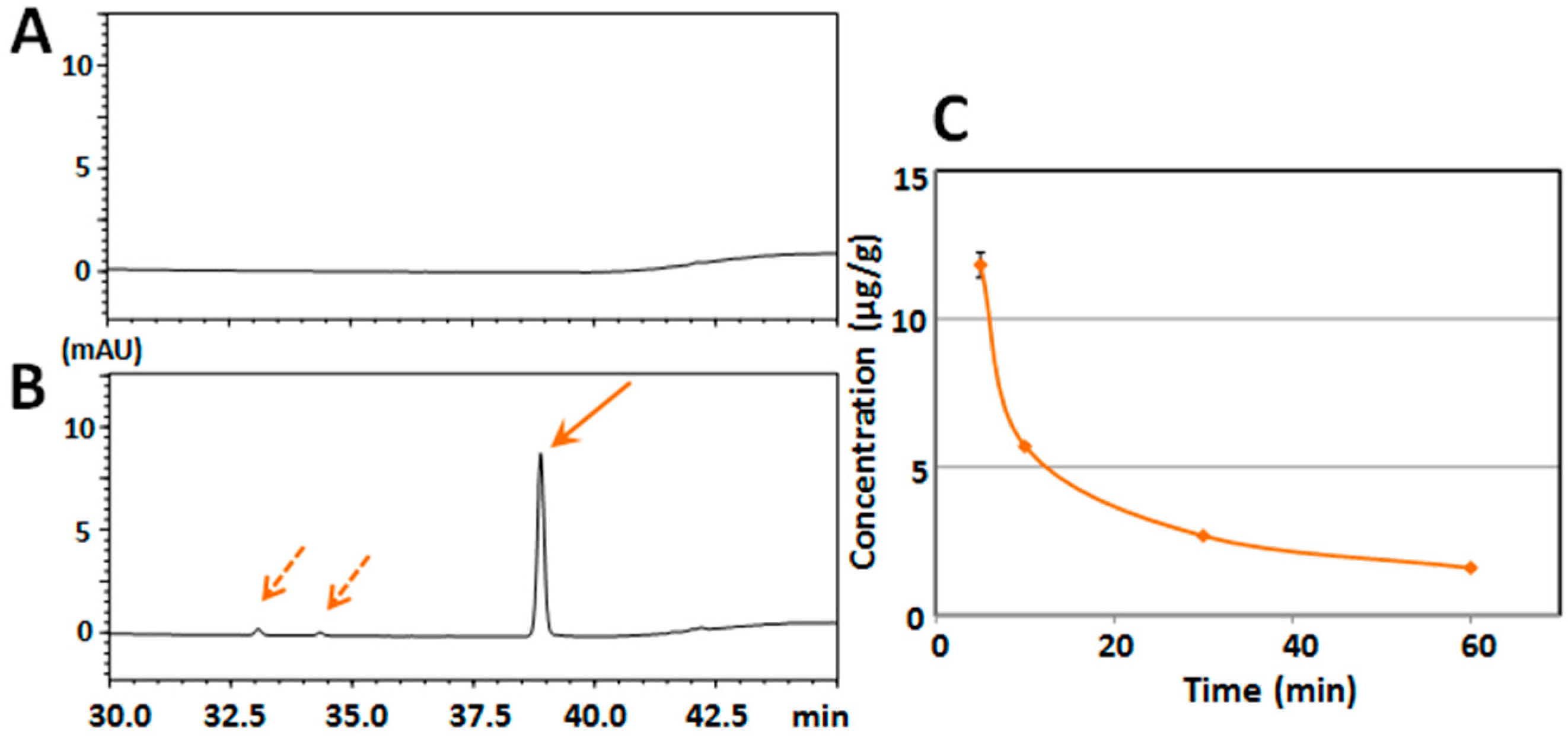
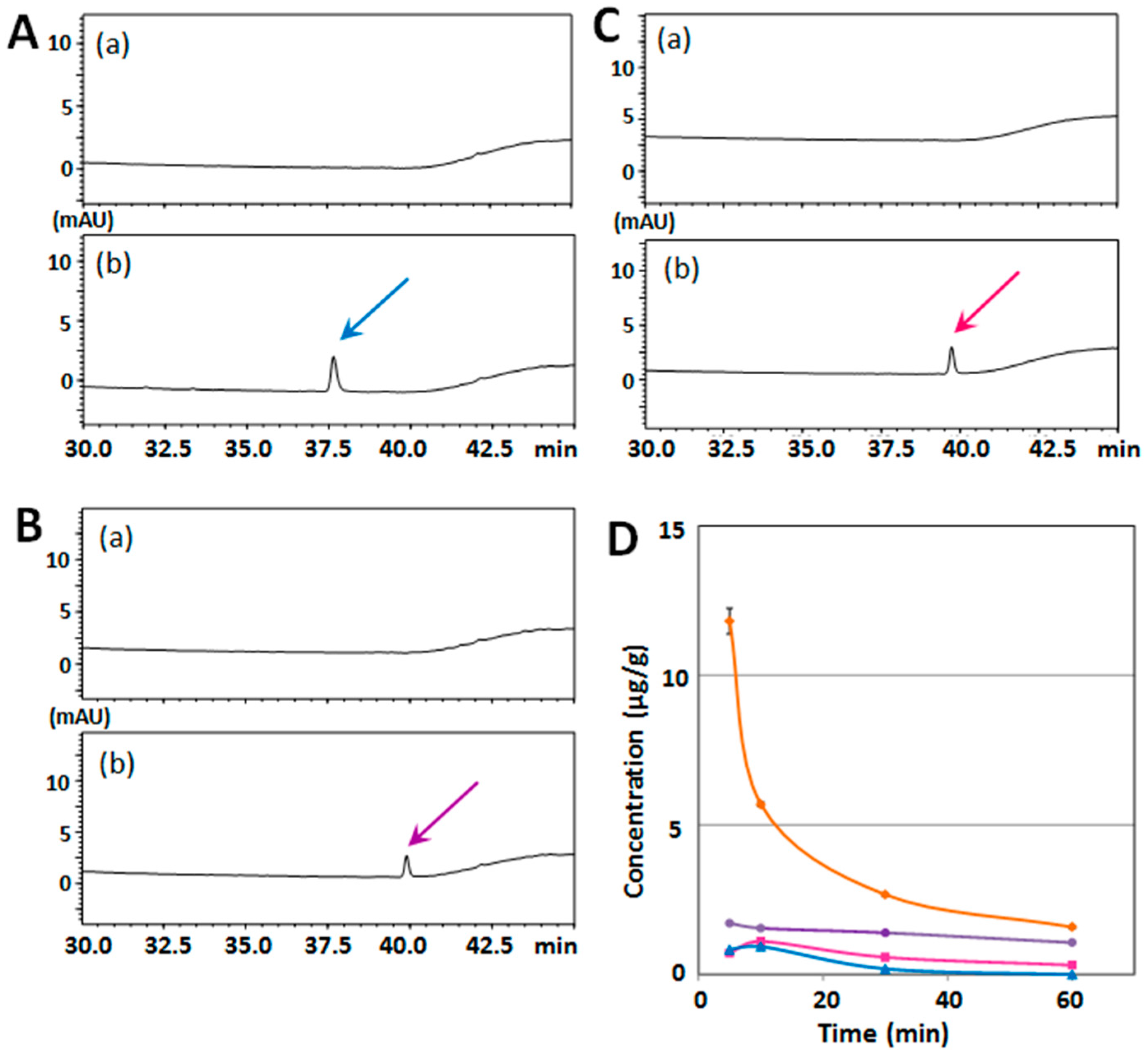
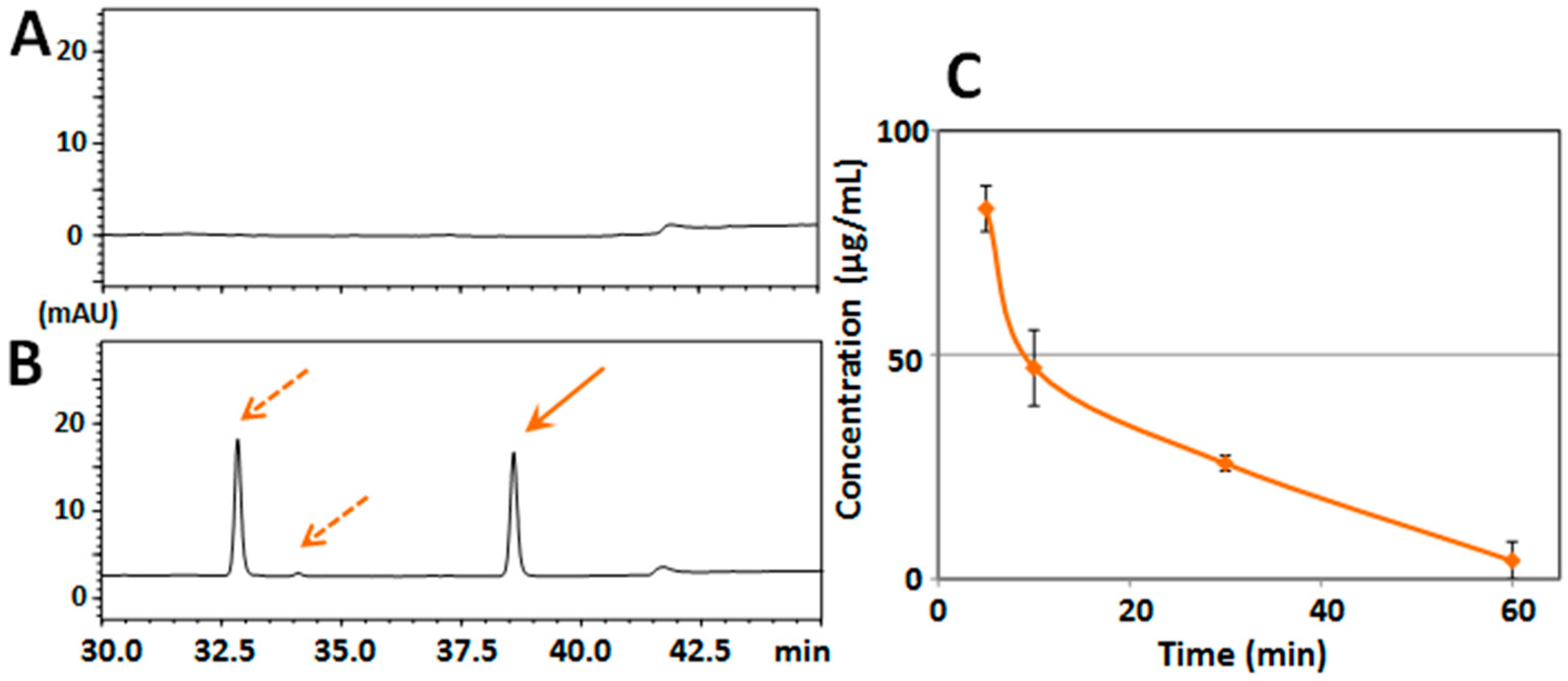
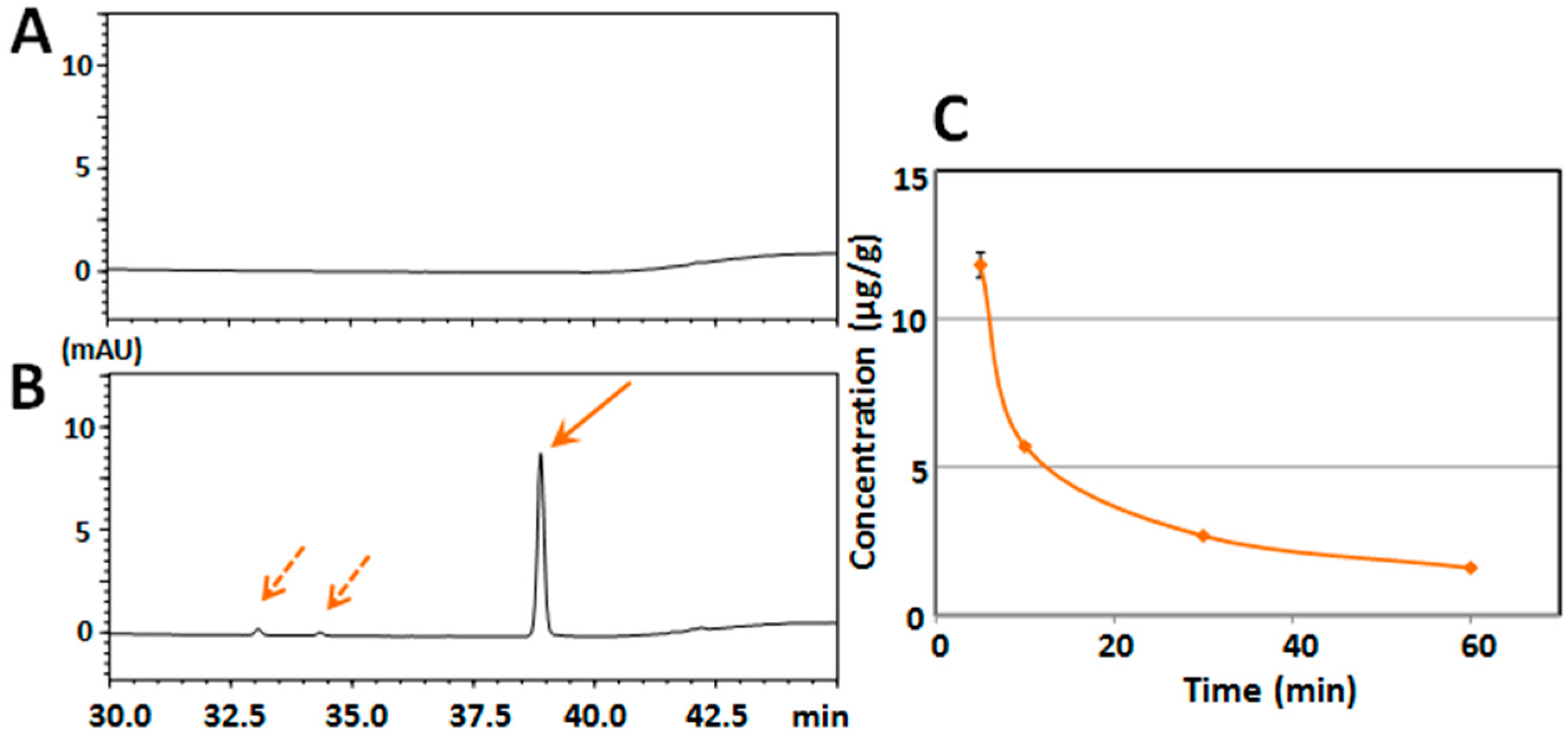
2.1. Determination of Serum/Brain Levels of 3,5,6,7,8,3′,4′-Heptamethoxyflavone after Intraperitoneal Administration
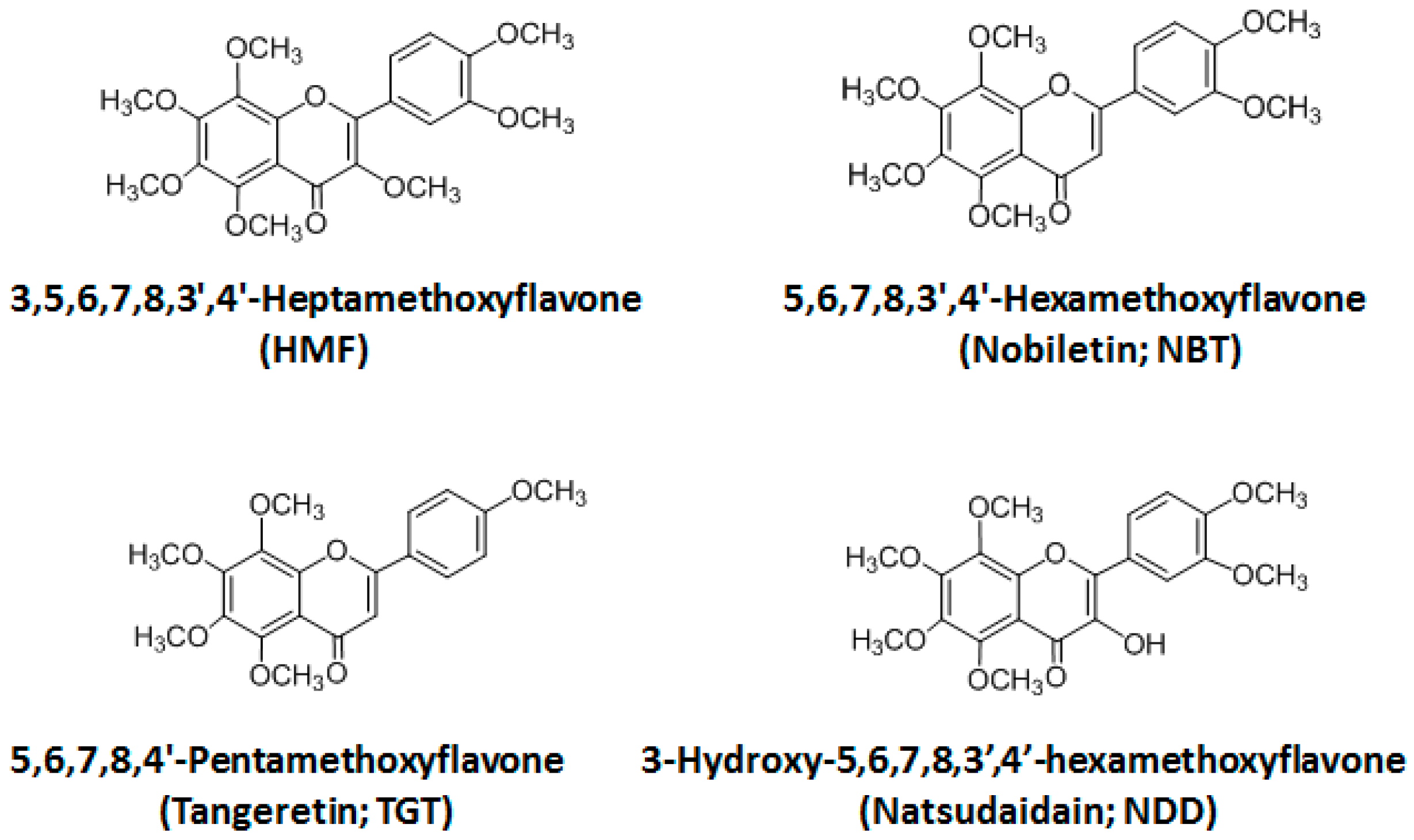
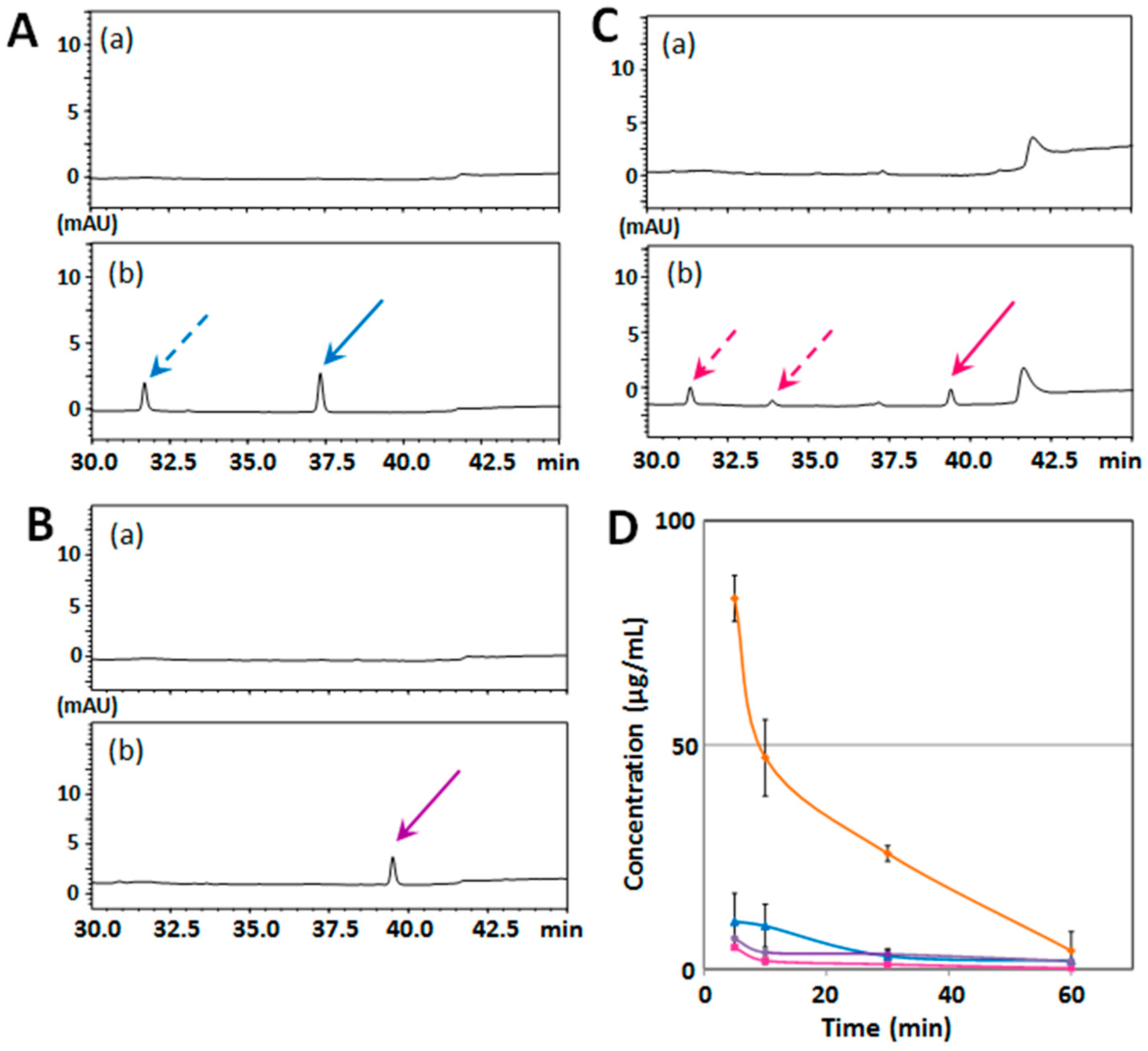
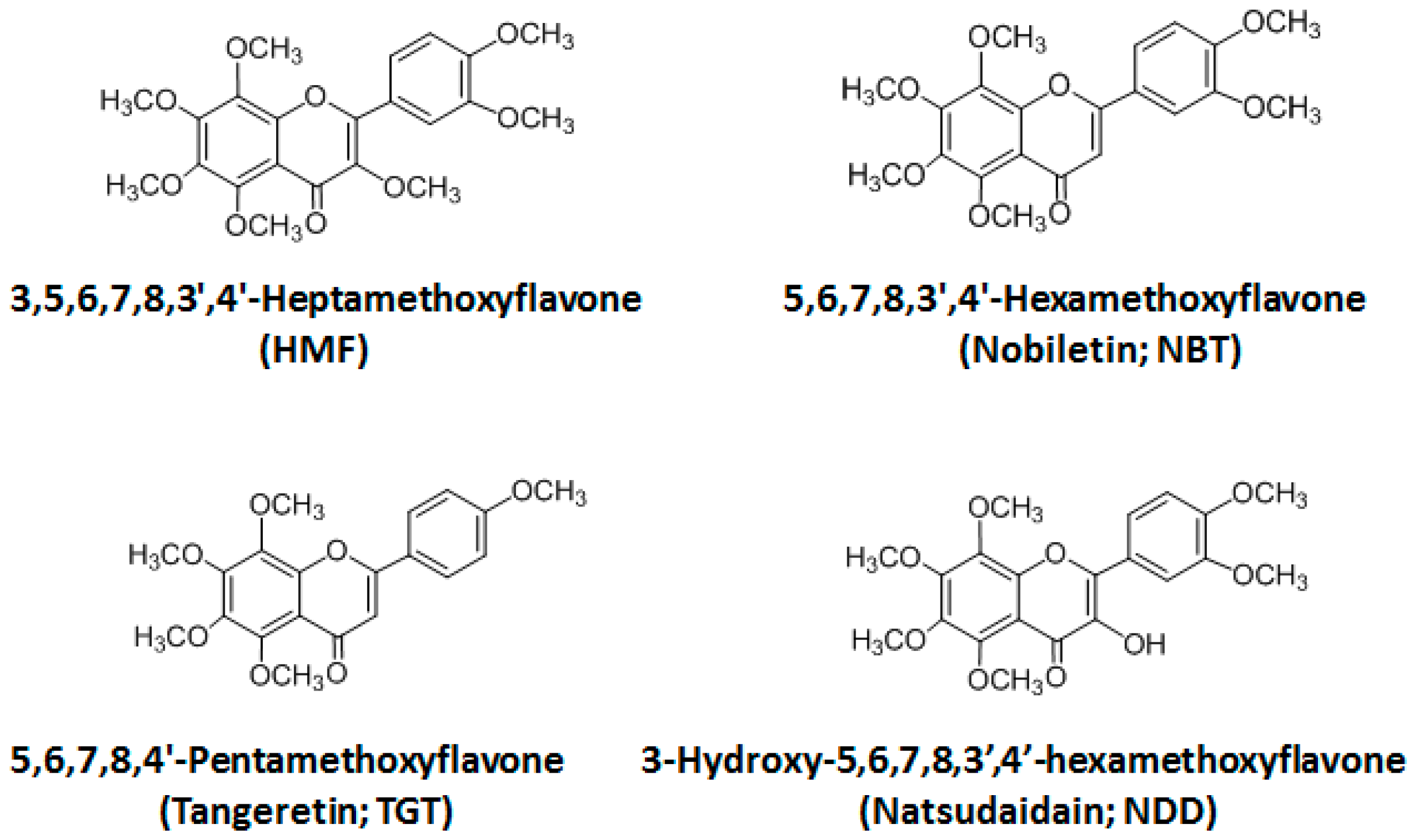
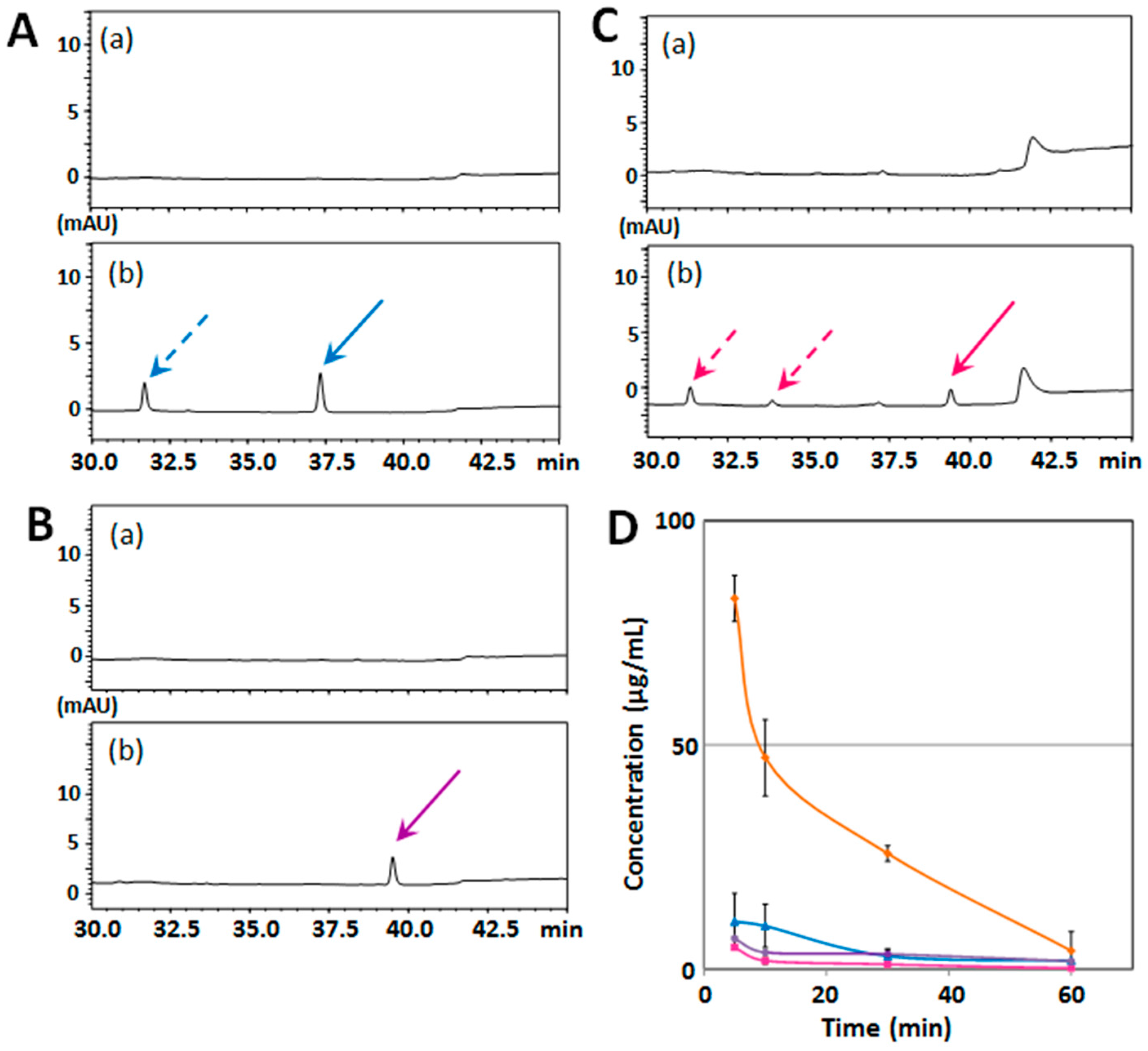
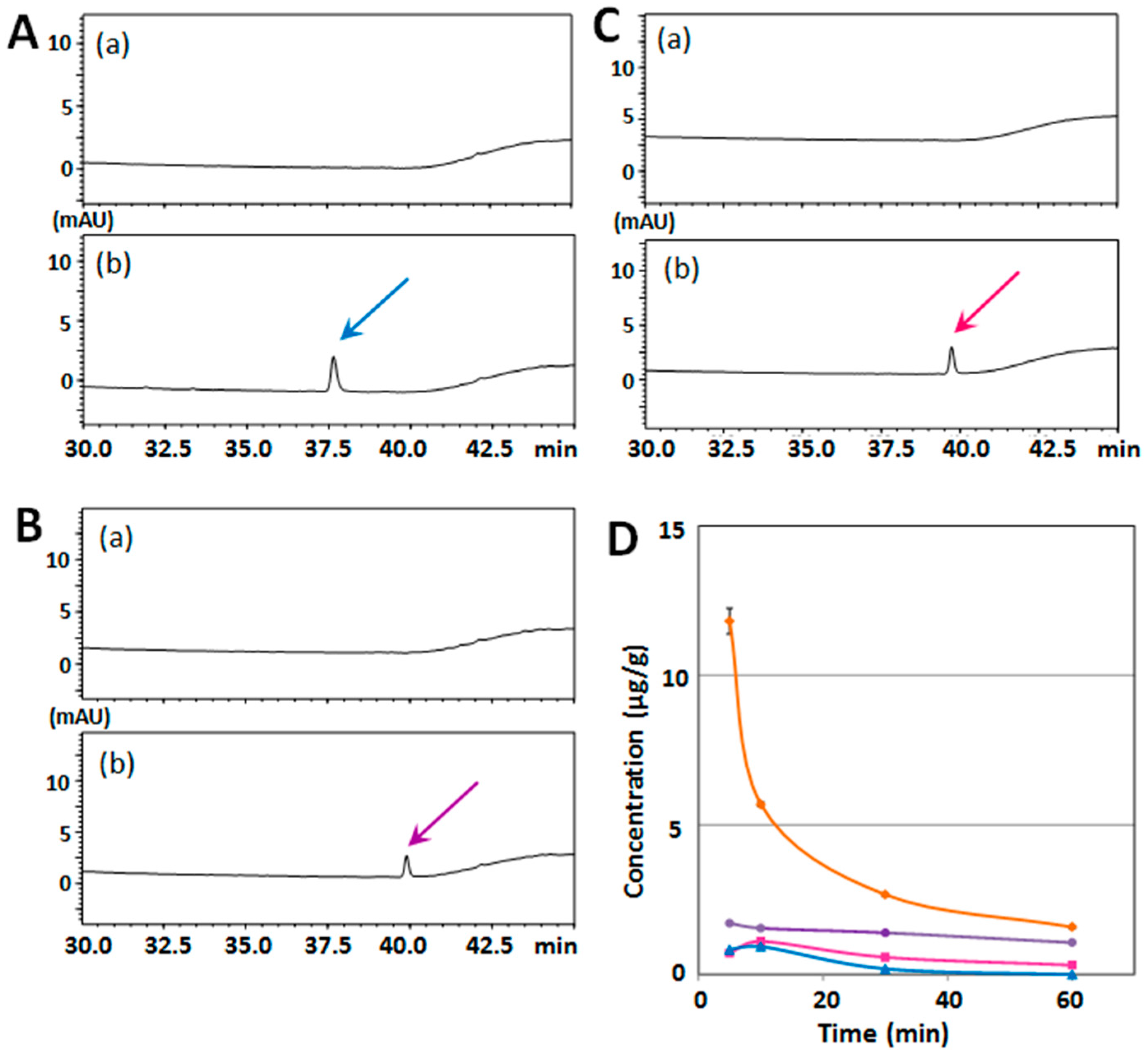
2.2. Structure–Activity Relationship of Permeability of the Blood–Brain Barrier to Polymethoxyflavones
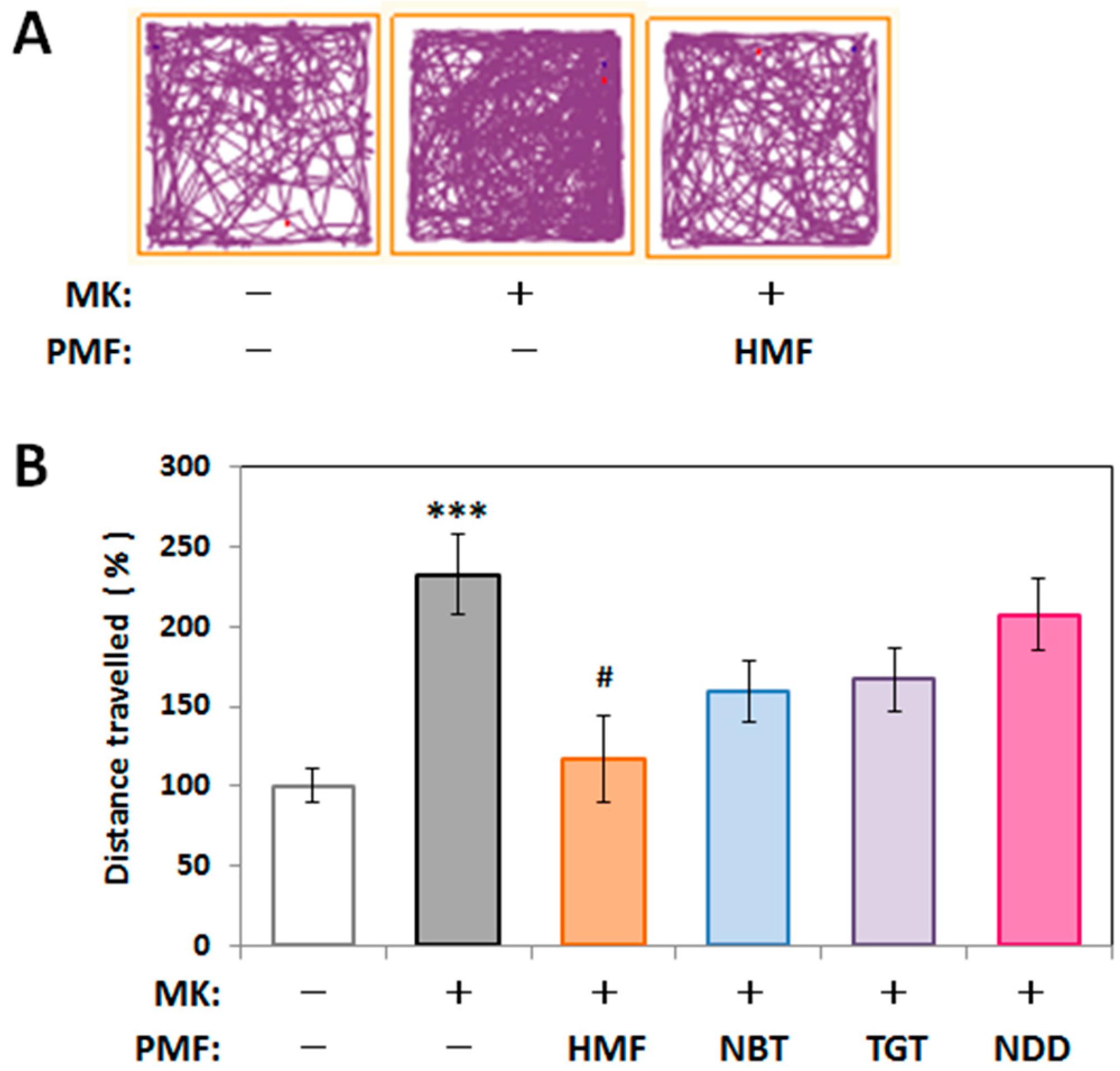
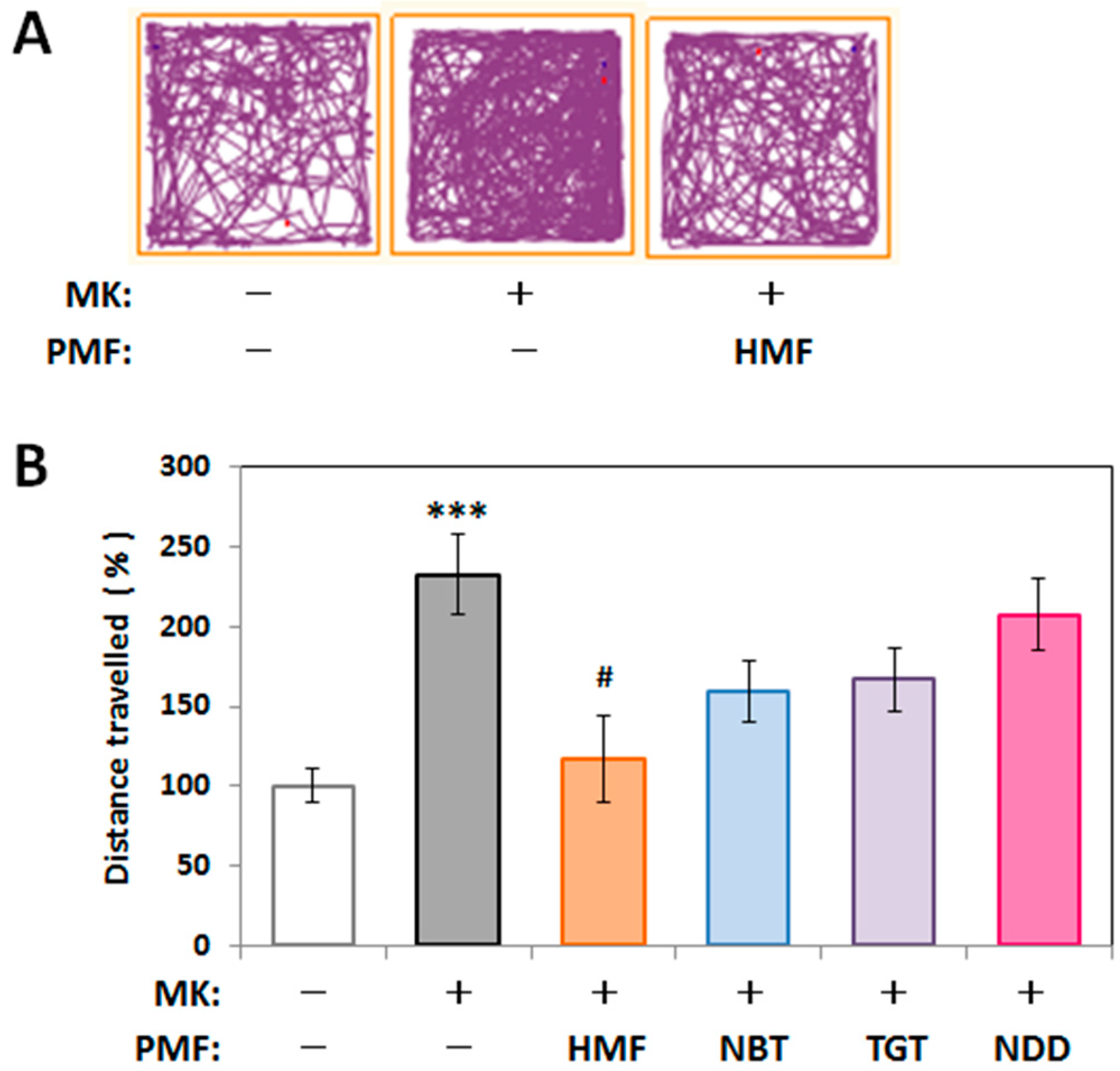
2.3. Structure–Activity Relationship of the Suppressive Effect of PMFs on MK-801-Induced Locomotive Hyperactivity
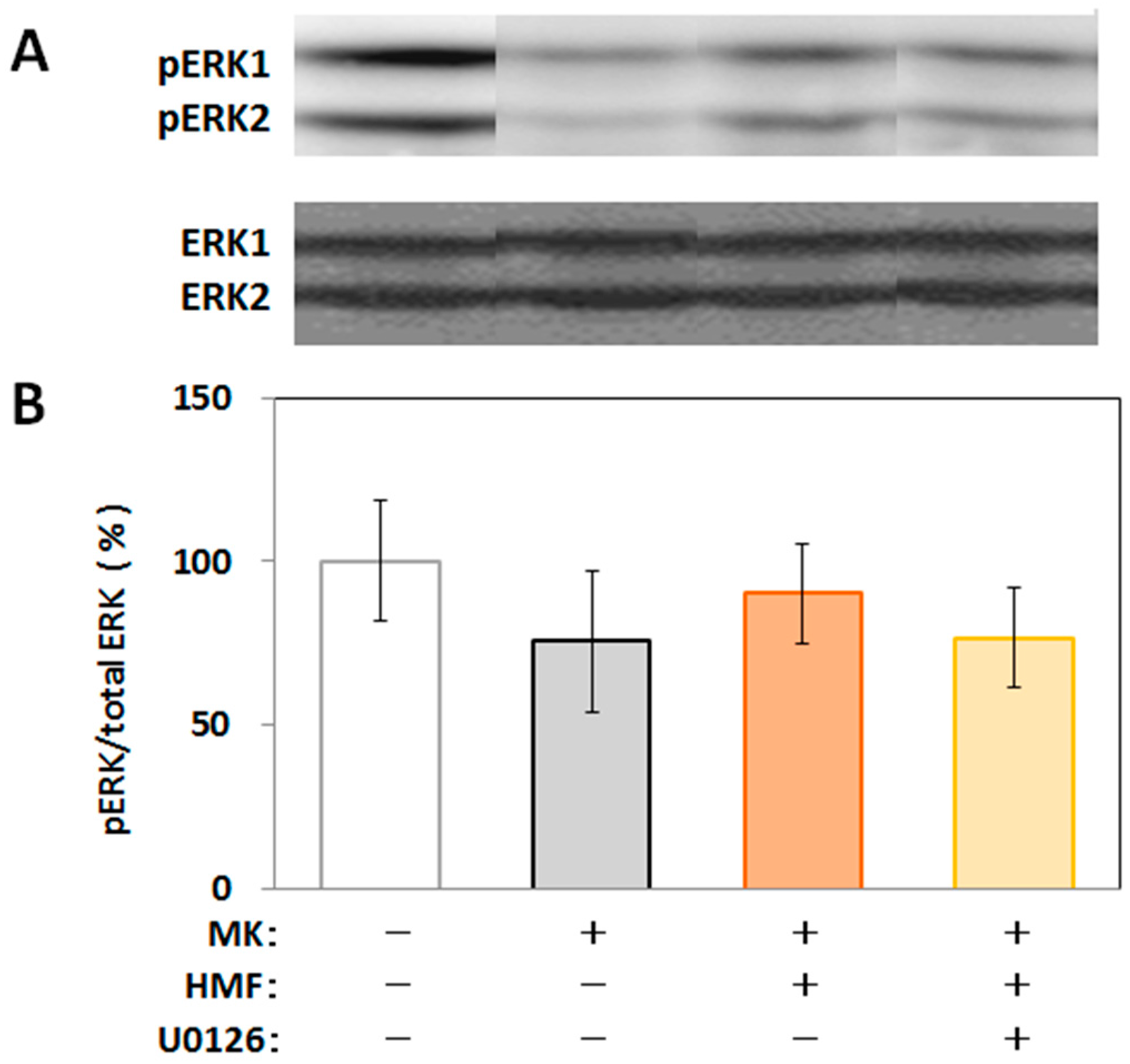
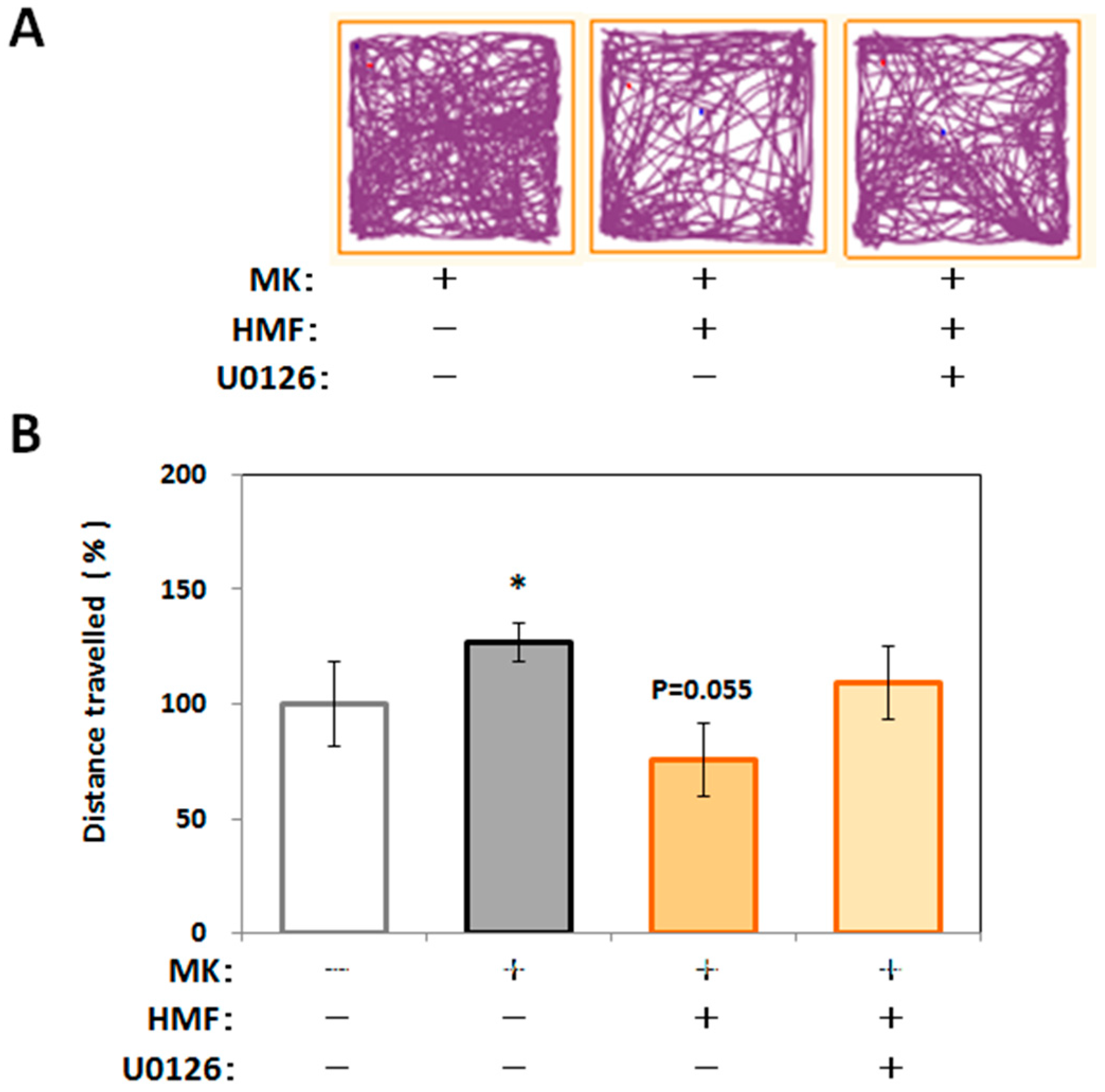
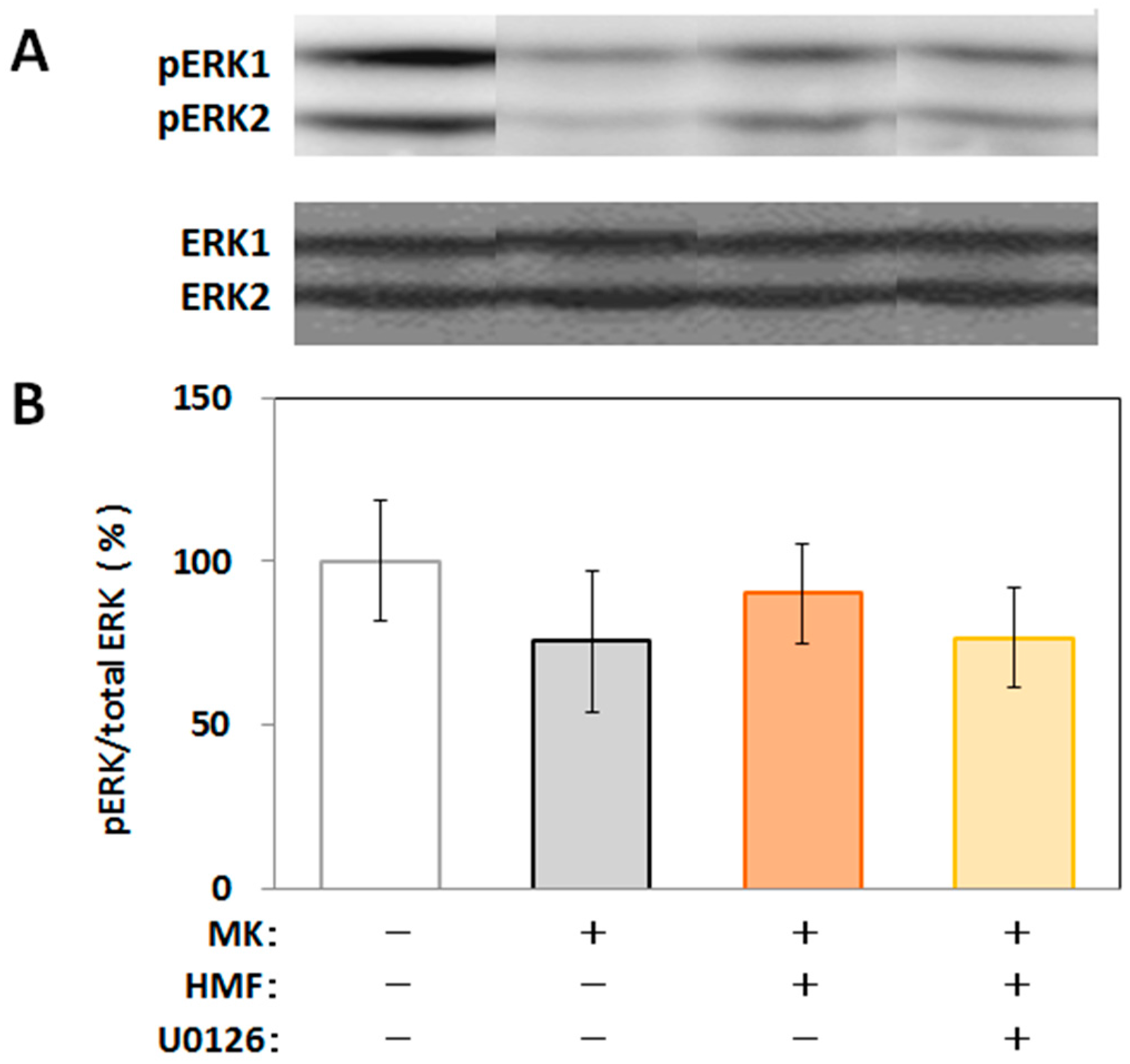
2.4. Mechanism of Inhibitory Effect of HMF on MK-801-Induced Locomotive Hyperactivity
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Determination of PMFs via HPLC/UV
3.3. Animals
3.4. Treatment with Reagents and Blood/Tissue Preparation
3.5. Sample Preparation for HPLC/UV
3.6. Open Field Test
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murakami, A.; Ohigashi, H. Anti-inflammatory and anticarcinogenesis potentials of citrus coumarins and polymethylated flavonoids. In Nutrigenomics and Proteomics in Health and Disease: Food Factors and Gene Interactions; Mine, Y., Miyashita, K., Shahidi, F., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 311–324. [Google Scholar]
- Spencer, J.P.E.; Vauzour, D.; Rendeiro, C. Flavonoids and cognition: The molecular mechanisms underlying their behavioural effects. Arch. Biochem. Biophys. 2009, 492, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Onozuka, H.; Nakajima, A.; Matsuzaki, K.; Shin, R.-W.; Ogino, K.; Saigusa, D.; Tetsu, N.; Yokosuka, A.; Sashida, Y.; Mimaki, Y.; et al. A citrus flavonoid, improves memory impairment and Aβ pathology in a transgenic mouse model of Alzheimer’s disease. J. Pharmacol. Exp. Res. 2008, 326, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Aoyama, Y.; Nguyen, T.-T.L.; Shin, E.-J.; Kim, H.-C.; Yamada, S.; Nakai, T.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; et al. Nobiletin, a citrus flavonoid, ameliorates cognitive impairment, oxidative burden, and hyperphosphorylation of tau in senescence-accelerated mouse. Behav. Brain Res. 2013, 250, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, D.; Shibuya, M.; Jinno, D.; Yamakoshi, H.; Iwabuchi, Y.; Yokosuka, A.; Mimaki, Y.; Naganuma, A.; Ohizumi, Y.; Tomioka, Y.; et al. High-performance liquid chromatography with photodiode array detection for determination of nobiletin content in the brain and serum of mice administrated the natural compound. Anal. Bioanal. Chem. 2011, 400, 635–3641. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Wahajuddin; Tewari, D.; Patel, K.; Jain, G.K. Permeability determination and pharmacokinetic study of nobiletin in rat plasma and brain by validated high-performance liquid chromatography method. Fitoter 2011, 82, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Shimada, N.; Kaji, M.; Morita, M.; Miyoshi, K.; Minami, S.; Amakura, Y.; Yoshimura, M.; Yoshida, T.; Watanabe, S.; et al. Heptamethoxyflavone a citrus flavonoid, enhances brain-derived neurotrophic factor production and neurogenesis in the hippocampus following cerebral global ischemia in mice. Neurosci. Lett. 2012, 528, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Morita, M.; Miyoshi, K.; Nishigawa, Y.; Kaji, M.; Sawamoto, A.; Terugo, T.; Toyoda, N.; Makihata, N.; Amakura, Y.; et al. 3,5,6,7,8,3′,4′-Heptamethoxyflavone, a citrus flavonoid, on protection against memory impairment and neuronal cell death in a global cerebral ischemia mouse model. Neurochem. Int. 2014, 70, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Sawamoto, A.; Okuyama, S.; Yamamoto, K.; Amakura, Y.; Yoshimura, M.; Nakajima, M.; Furukawa, Y. 3,5,6,7,8,3′,4′-Heptamethoxyflavone, a citrus flavonoid, ameliorates corticosterone-induced depression-like behavior and restores brain-derived neurotrophic factor expression, neurogenesis, and neuroplasticity in the hippocampus. Molecules 2016, 21, 541. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Miyoshi, K.; Tsumura, Y.; Amakura, Y.; Yoshimura, M.; Yoshida, T.; Nakajima, M.; Furukawa, Y. 3,5,6,7,8,3′,4′-Heptamethoxyflavone, a citrus polymethoxylated flavones, attenuates inflammation in the mouse hippocampus. Brain Sci. 2015, 5, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Fukata, T.; Nishigawa, Y.; Amakura, Y.; Yoshimura, M.; Yoshida, T.; Nakajima, M.; Furukawa, Y. Citrus flavonoid improves MK-801-induced locomotive hyperactivity: Possible relevance to schizophrenia. J. Funct. Foods 2013, 5, 2002–2006. [Google Scholar] [CrossRef]
- Inoue, Y.; Kiryu, S.; Izawa, K.; Watanabe, M.; Tojo, A.; Ohtomo, K. Comparison of subcutaneous and intraperitoneal injection of D-luciferin for in vivo bioluminescence imaging. Eur. J. Nucl. Med. Mol. Imaging 2009, 36, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Cesar, T.B.; Jackson, E.; Mertens-Talcott, S. Pharmacokinetic study of nobiletin and tangeretin in rat serum by high-performance liquid chromatography-electrospray ionization-mass spectrometry. J. Agric. Food Chem. 2011, 59, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, M.; Bevilaqua, L.R.; Ardenghi, P.; Paratcha, G.; Levi, M.; Izquierdo, I.; Medina, J.H. Learning-associated activation of nuclear MAPK, CREB and Elk-1 along with Fos production, in the rat hippocampus after a one-trial avoidance learning: Abolition by NMDA receptor blockade. Brain Res. 2000, 76, 36–46. [Google Scholar] [CrossRef]
- Furukawa, Y.; Okuyama, S.; Amakura, Y.; Watanabe, S.; Fukata, T.; Nakajima, M.; Yoshimura, M.; Yoshida, T. Isolation and characterization of activators of ERK/MAPK from citrus plants. Int. J. Mol. Sci. 2012, 13, 1832–1845. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Morita, M.; Kaji, M.; Amakura, Y.; Yoshimura, M.; Shimamoto, K.; Ookido, Y.; Nakajima, M.; Furukawa, Y. Auraptene acts as an anti-Inflammatory agent in the mouse brain. Molecules 2015, 20, 20230–20239. [Google Scholar] [CrossRef] [PubMed]
- Dyck, B.; Guest, K.; Sookram, C.; Basu, D.; Johnson, R.; Mishra, R.K. PAOPA, a potent analogue of Pro-Leu-glycinamide and allosteric modulator of the dopamine D2 receptor, prevents NMDA receptor antagonist (MK-801)-induced deficits in social interaction in the rat: Implications for the treatment of negative symptoms in schizophrenia. Schizophr. Res. 2011, 125, 88–92. [Google Scholar] [PubMed]
- Andine, P.; Widermark, N.; Axelsson, R.; Nyberg, G.; Olofsson, U.; Martensson, E.; Sandberg, M. Characterization of MK-801-induced behavior as a putative rat model of psychosis. J. Pharmacol. Exp. Ther. 1999, 290, 1393–1408. [Google Scholar] [PubMed]
- Van den Buuse, M. Modeling the positive symptoms of schizophrenia in genetically modified mice: Pharmacology and methodology aspects. Schizophr. Bull. 2010, 36, 246–270. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okuyama, S.; Miyazaki, K.; Yamada, R.; Amakura, Y.; Yoshimura, M.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Permeation of Polymethoxyflavones into the Mouse Brain and Their Effect on MK-801-Induced Locomotive Hyperactivity. Int. J. Mol. Sci. 2017, 18, 489. https://doi.org/10.3390/ijms18030489
Okuyama S, Miyazaki K, Yamada R, Amakura Y, Yoshimura M, Sawamoto A, Nakajima M, Furukawa Y. Permeation of Polymethoxyflavones into the Mouse Brain and Their Effect on MK-801-Induced Locomotive Hyperactivity. International Journal of Molecular Sciences. 2017; 18(3):489. https://doi.org/10.3390/ijms18030489
Chicago/Turabian StyleOkuyama, Satoshi, Kohei Miyazaki, Rie Yamada, Yoshiaki Amakura, Morio Yoshimura, Atsushi Sawamoto, Mitsunari Nakajima, and Yoshiko Furukawa. 2017. "Permeation of Polymethoxyflavones into the Mouse Brain and Their Effect on MK-801-Induced Locomotive Hyperactivity" International Journal of Molecular Sciences 18, no. 3: 489. https://doi.org/10.3390/ijms18030489