
Exploring the Functions of 9-Lipoxygenase (DkLOX3) in Ultrastructural Changes and Hormonal Stress Response during Persimmon Fruit Storage

Abstract
:1. Introduction
2. Results
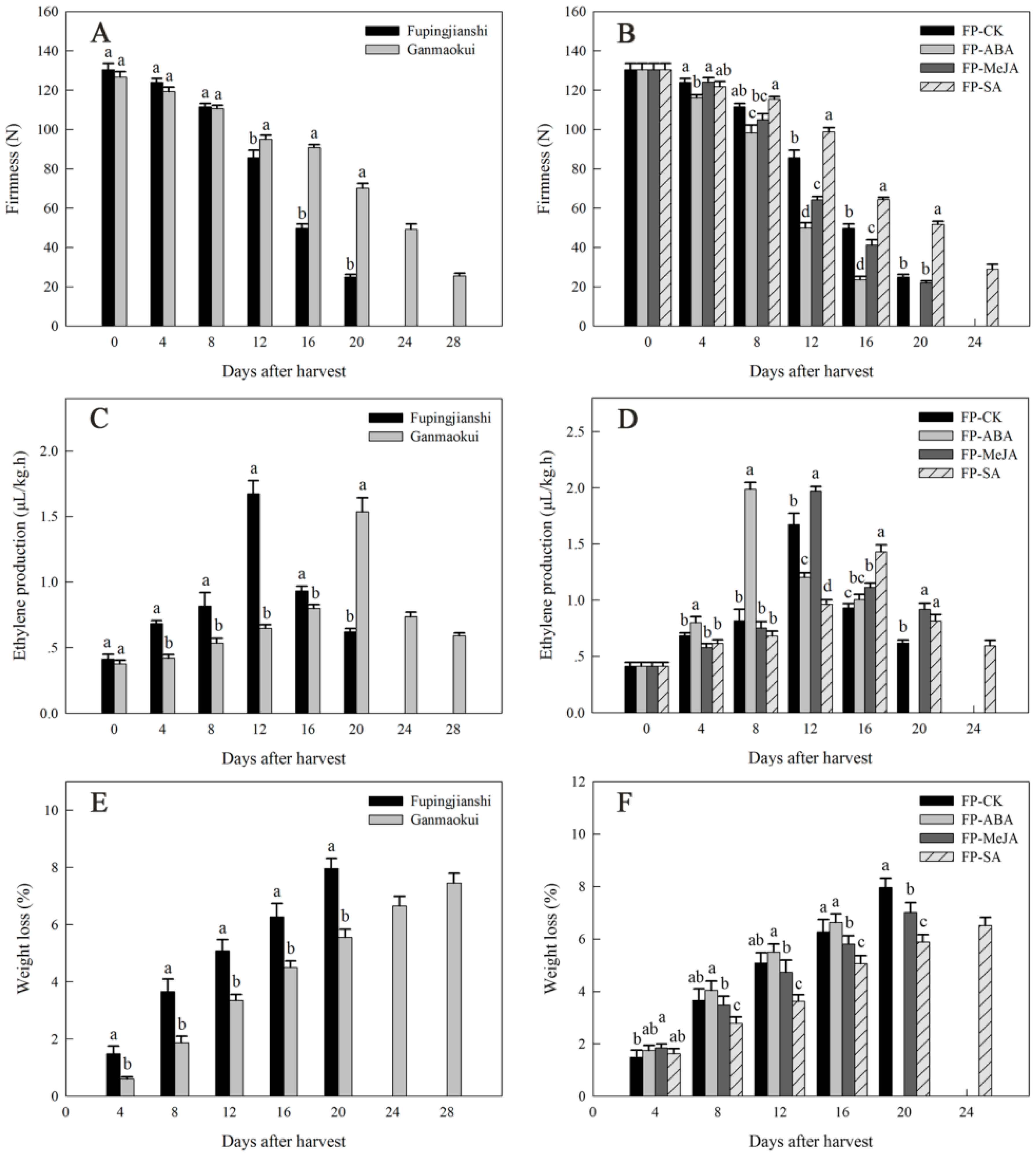
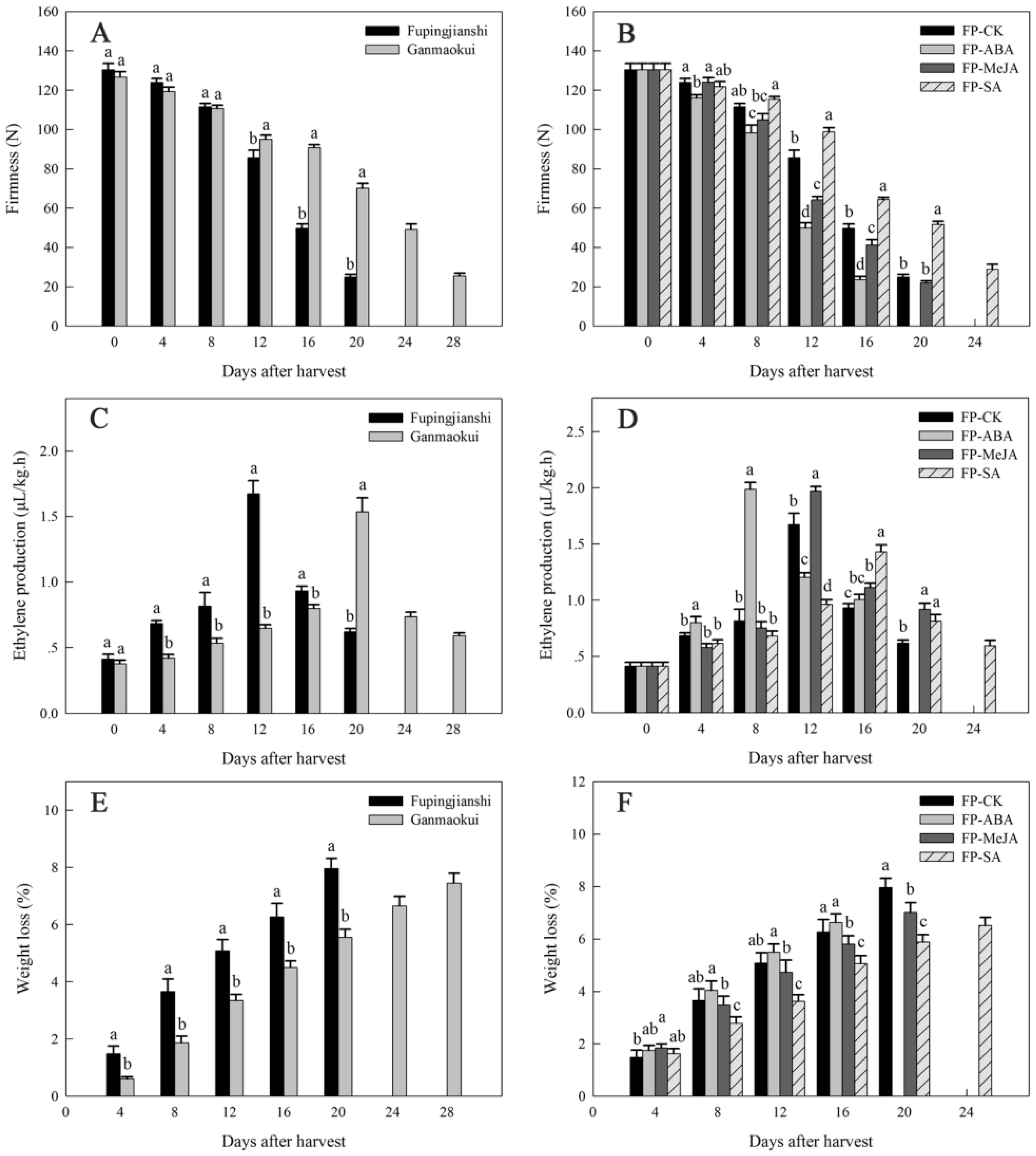
2.1. Physiological Characterization during Persimmon Fruit Storage
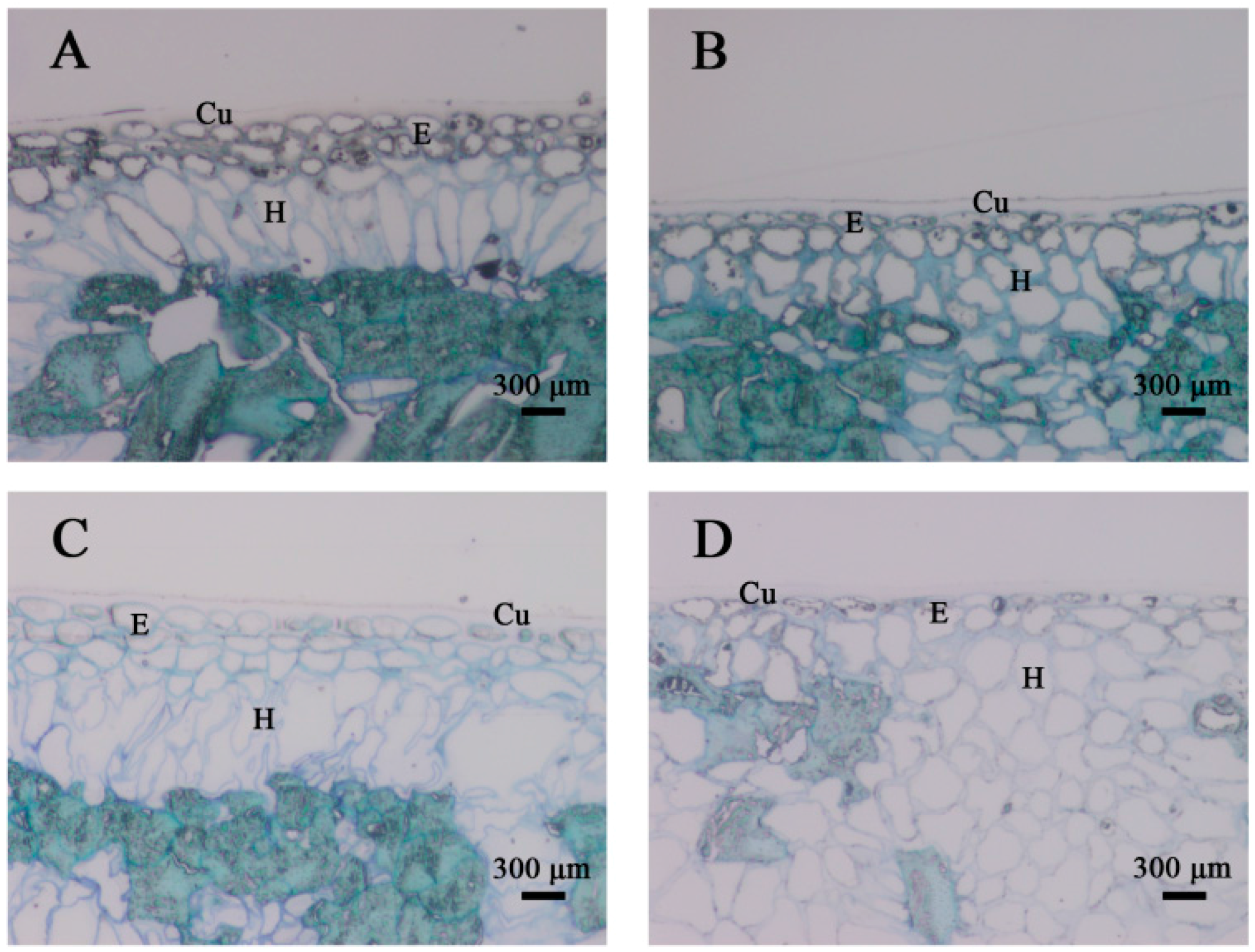
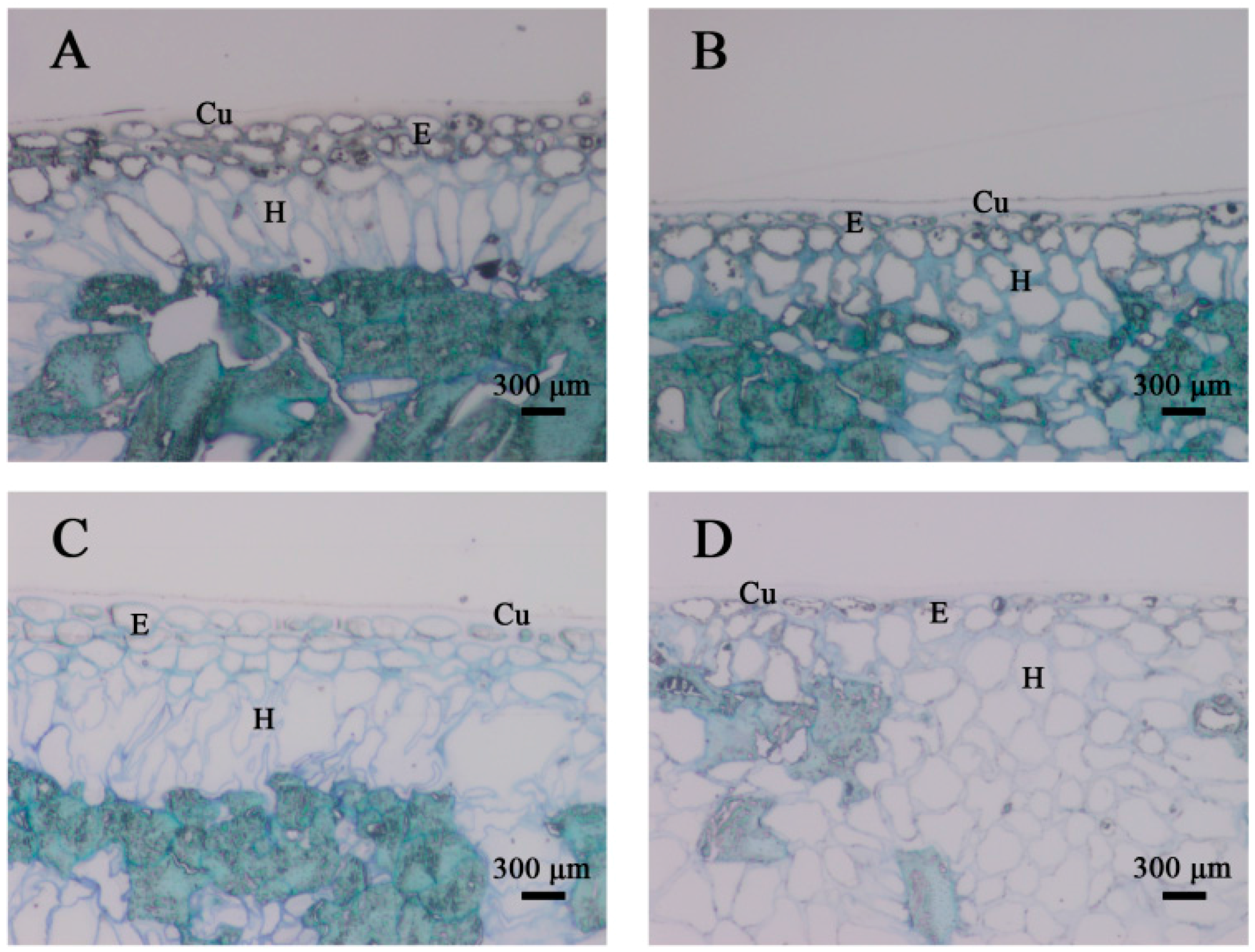
2.2. Structural Analyses of the Persimmon Fruit Peel during Storage
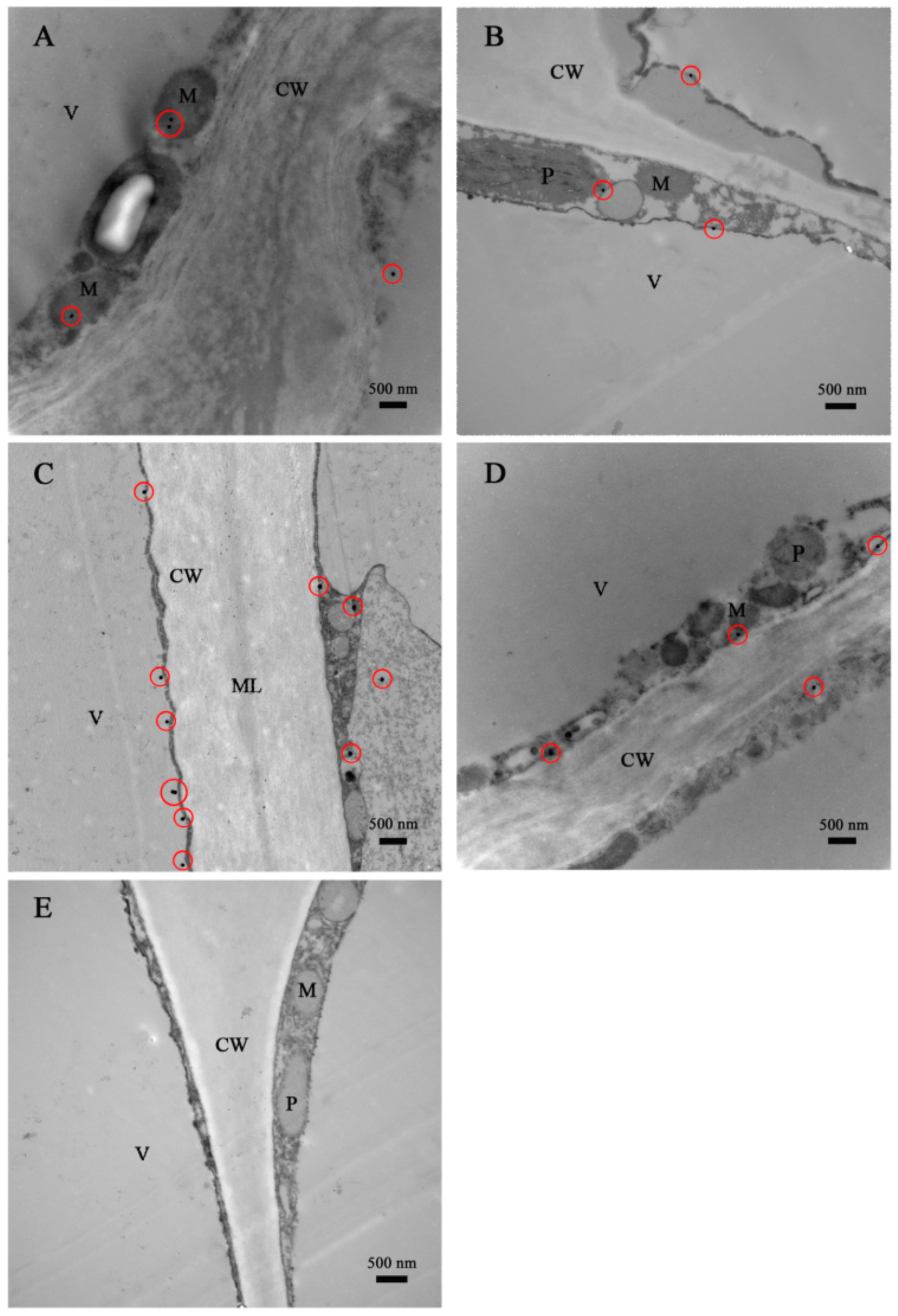
2.3. Immunocytolocalization of DkLOX3 in Persimmon Fruit during Storage
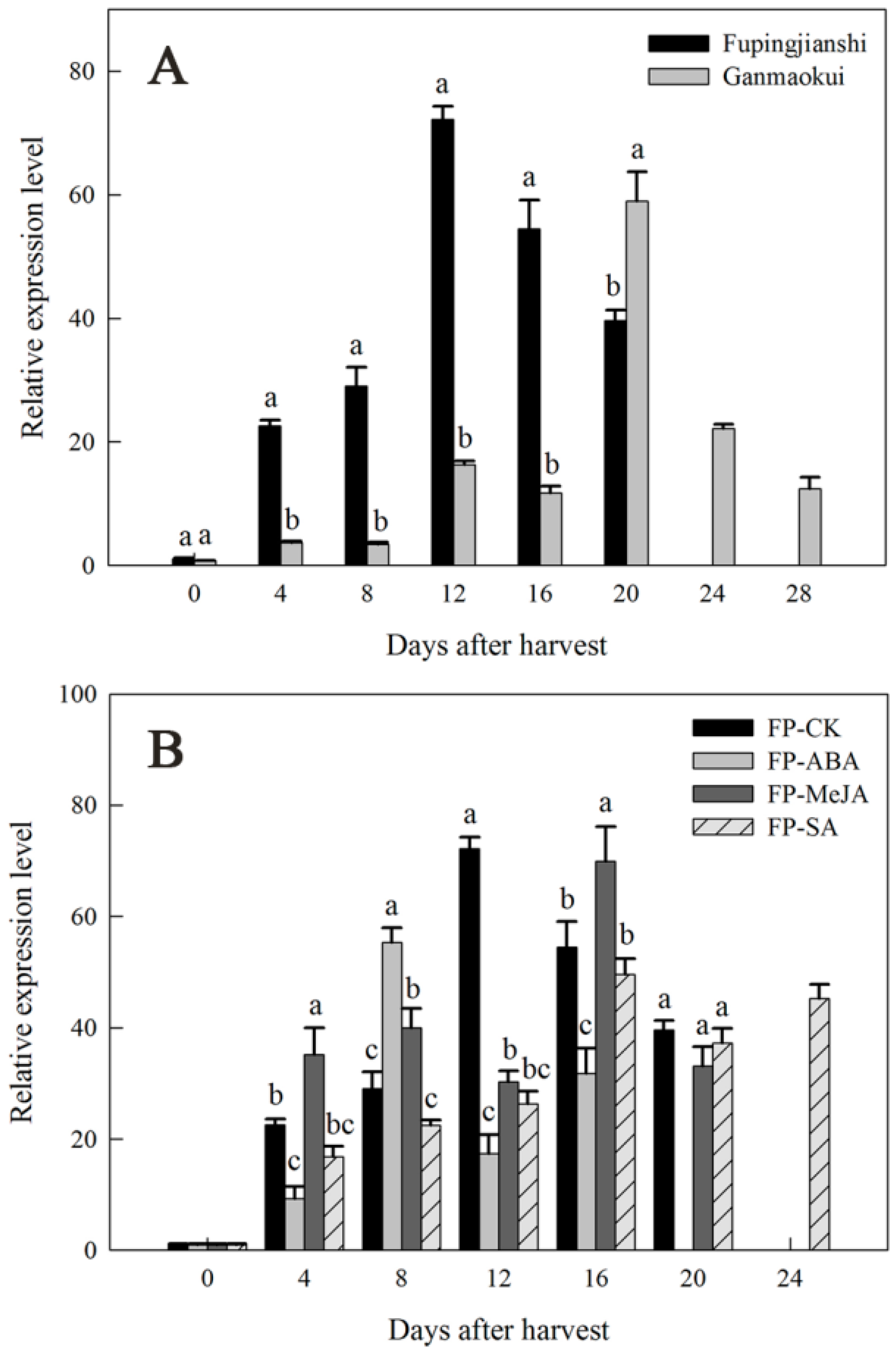
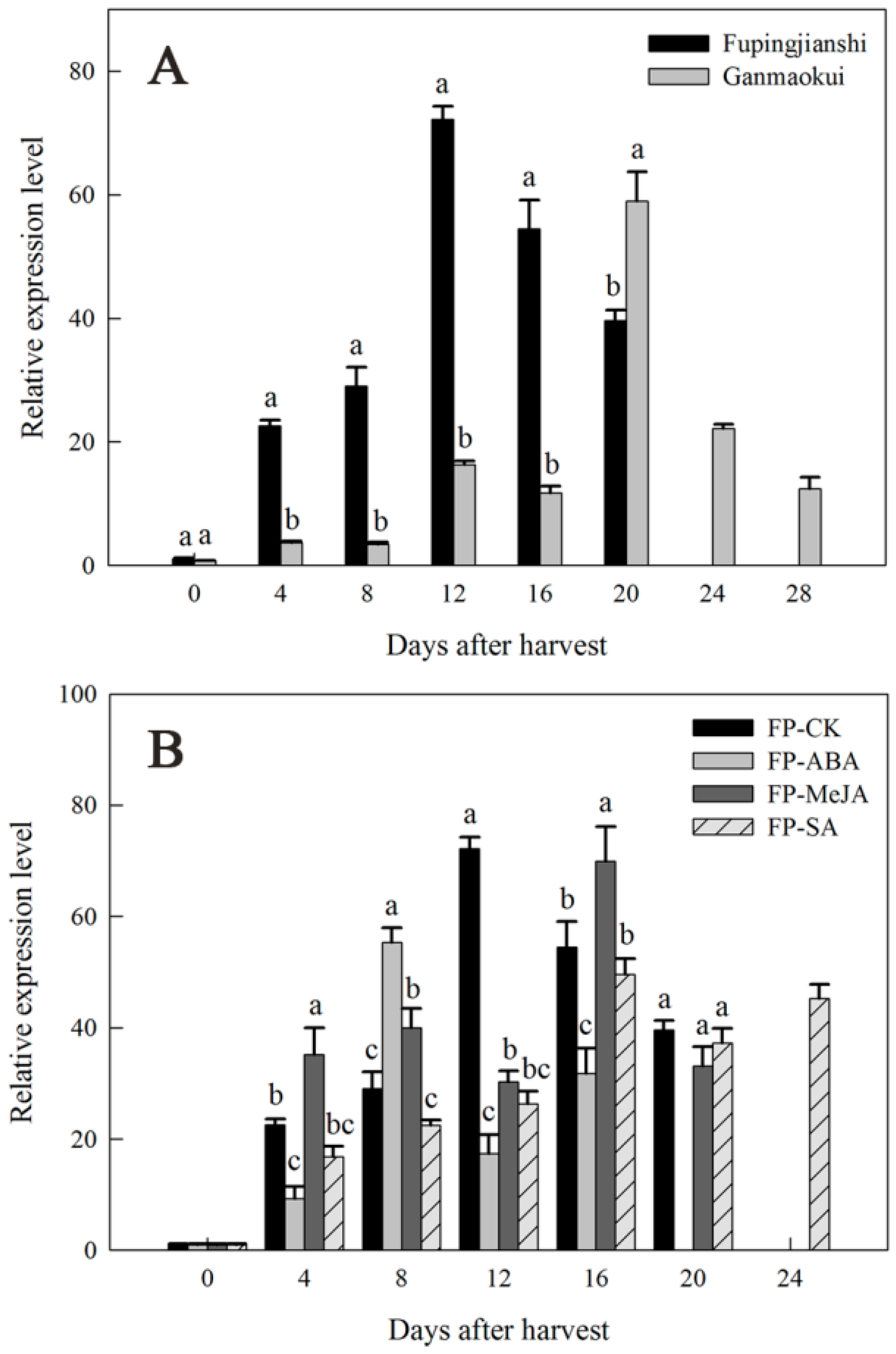
2.4. Expression of DkLOX3 during Persimmon Fruit Storage
2.5. Isolation and Sequence Analysis of the DkLOX3 Promoter
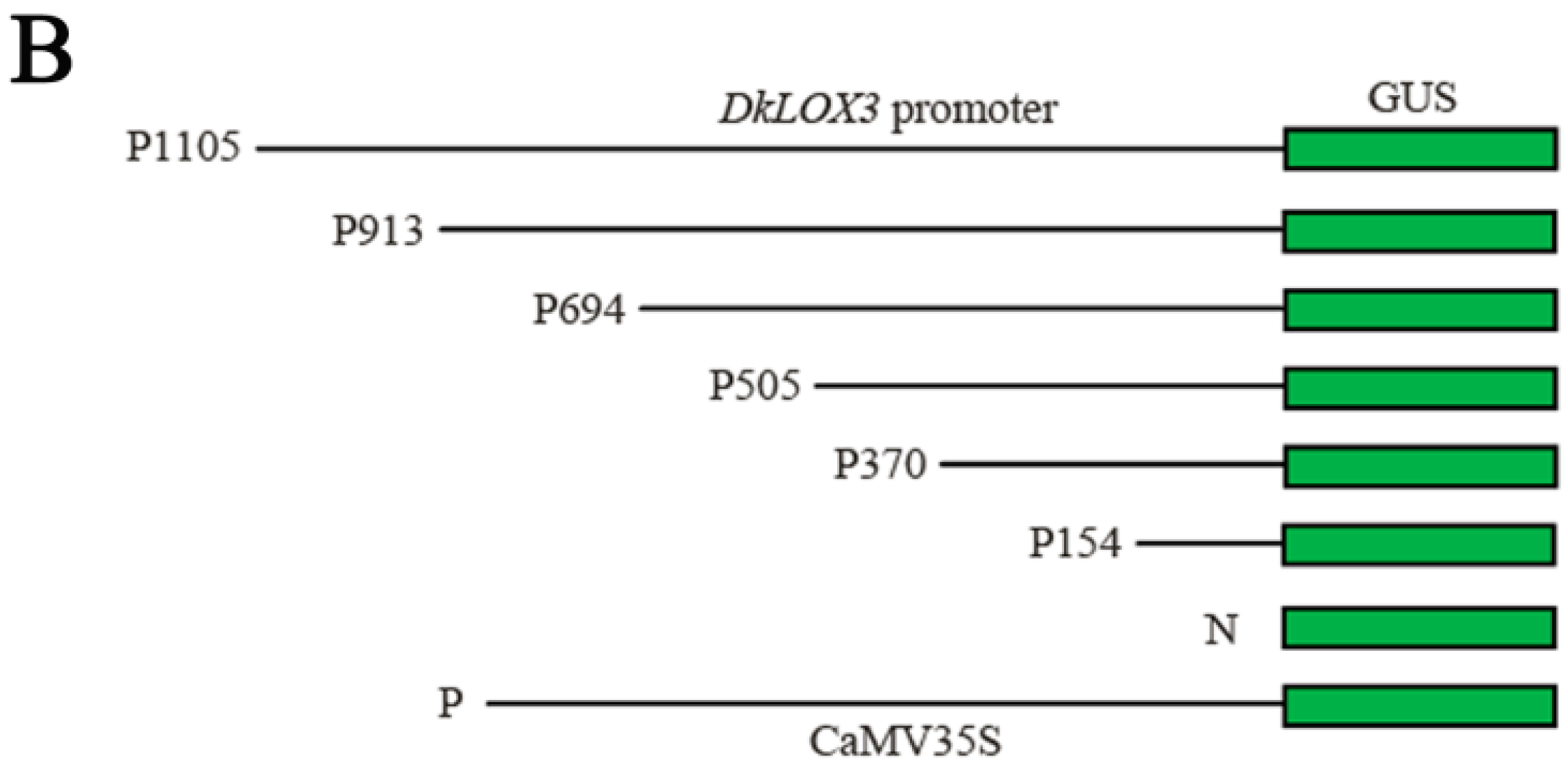
2.6. Characterization of DkLOX3 Promoter Activity in Tobacco Leaves
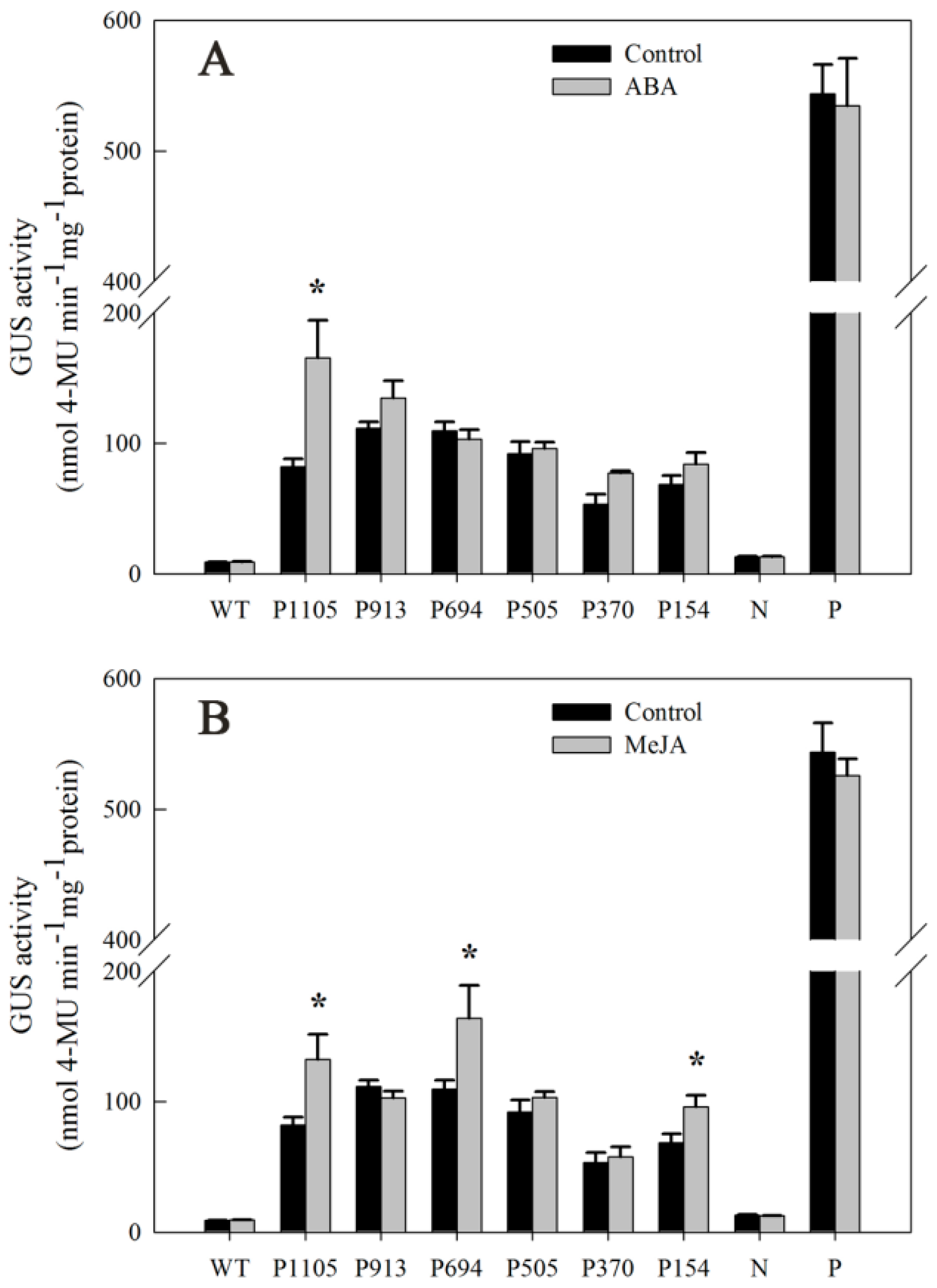
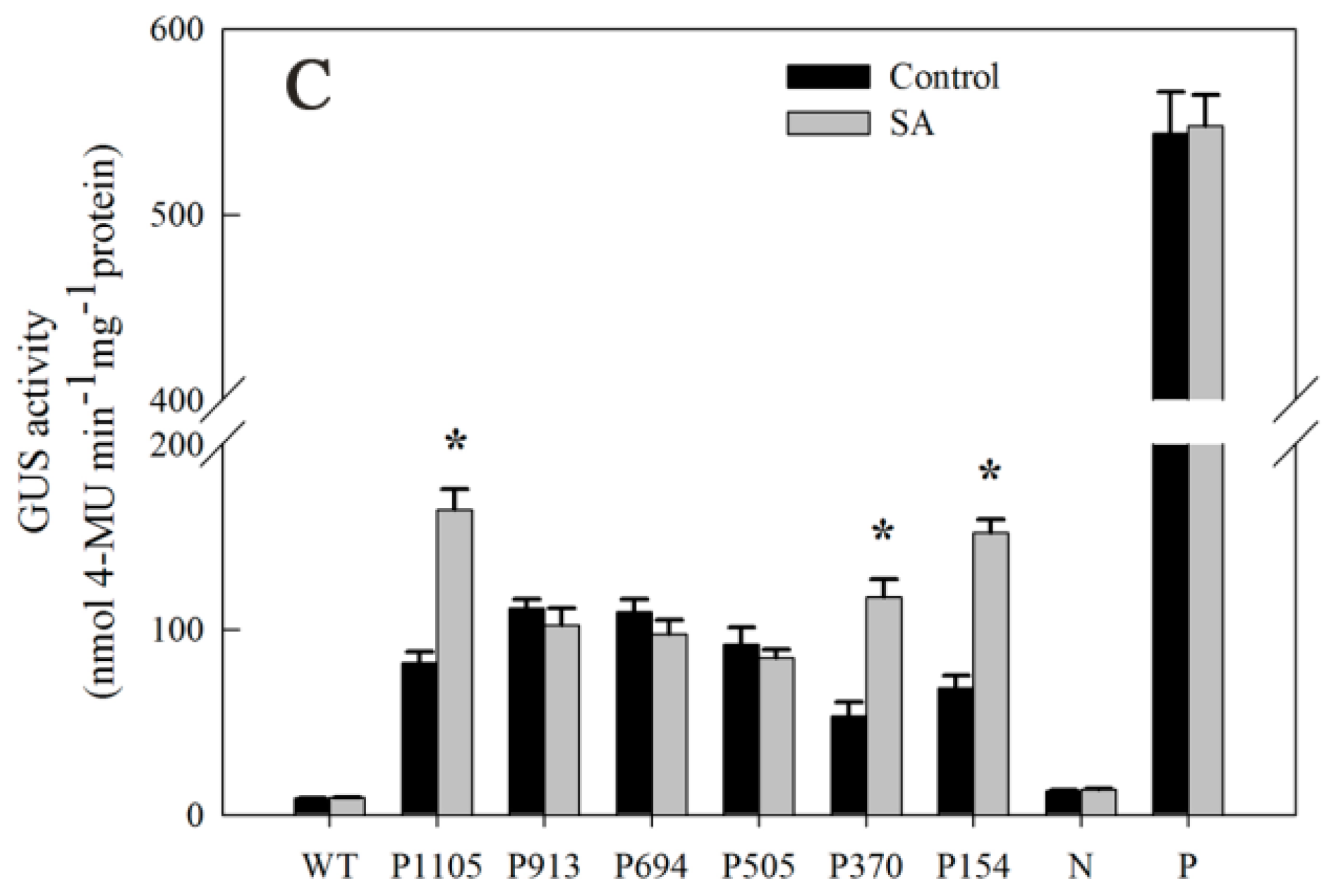
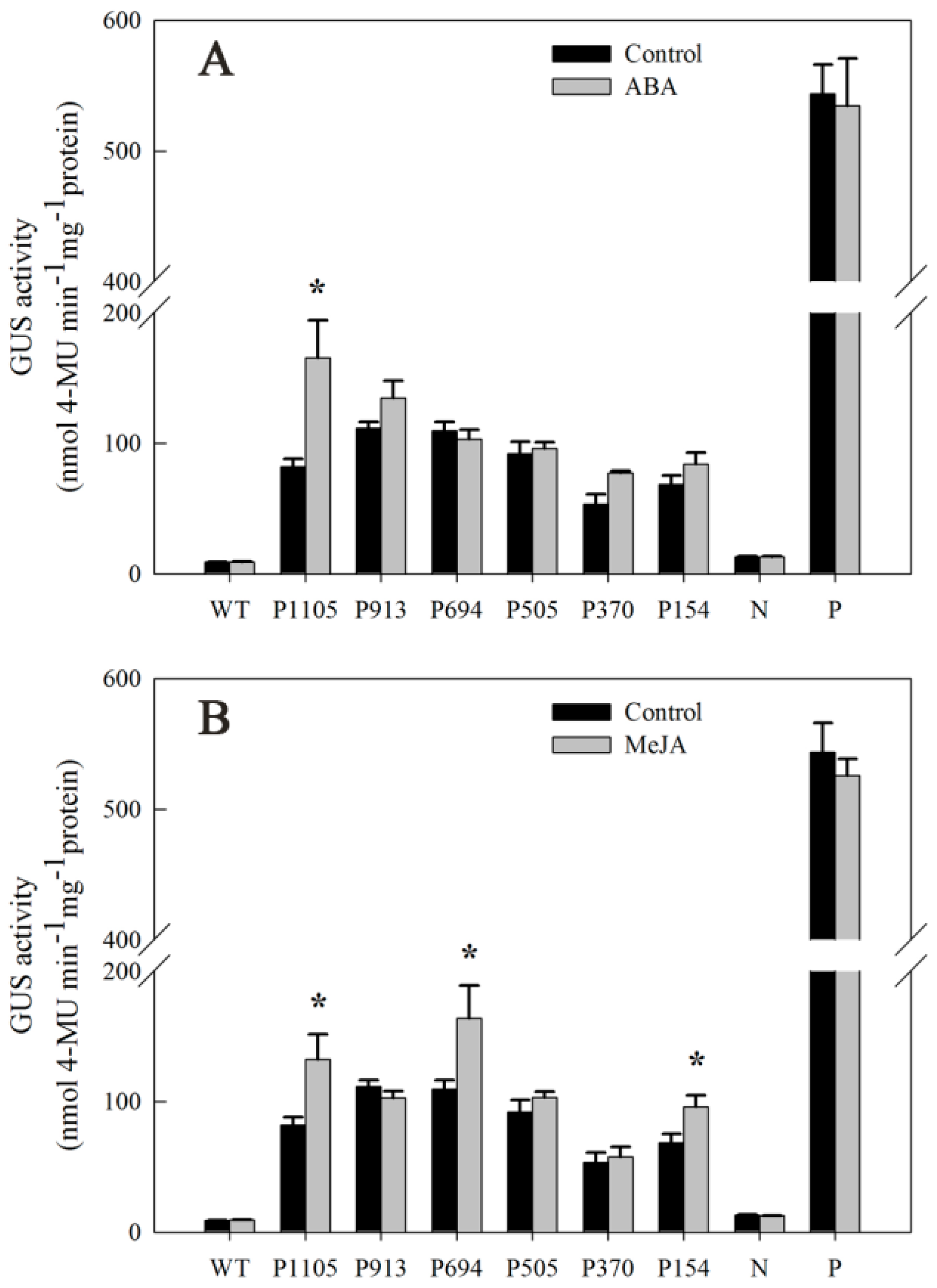
2.7. Responsiveness of DkLOX3 Promoters to Hormonal Stress
3. Discussion
3.1. DkLOX3 May Have a Positive Role in Ultrastructural Changes during Fruit Ripening
3.2. DkLOX3 May Play an Important Role in Hormonal Stress Response during Fruit Ripening
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Fruit Firmness, Ethylene Production and Weight Loss
4.3. Structural Analyses
4.4. Immunolabelling for Transmission Electron Microscopy (TEM)
4.5. Quantitative Real-Time PCR Analysis
4.6. Cloning of the 5′ Flanking Region of DkLOX3
4.7. Construction of the Promoter-GUS Fusion
4.8. Agrobacterium-Mediated Transient Expression Assays in Tobacco Plants
4.9. Histochemical and Fluorometric Assays for GUS Activity
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Z.; Fu, R.; Huber, D.J.; Rao, J.; Chang, X.; Hu, M.; Zhang, Y.; Jiang, N. Expression of expansin gene (CDK-Exp3) and its modulation by exogenous gibberellic acid during ripening and softening of persimmon fruit. HortScience 2012, 47, 378–381. [Google Scholar]
- Nakano, R.; Ogura, E.; Kubo, Y.; Inaba, A. Ethylene biosynthesis in detached young persimmon fruit is initiated in calyx and modulated by water loss from the fruit. Plant Physiol. 2003, 131, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Kader, A.A. Fruit maturity, ripening, and quality relationships. Acta Hortic. 1999, 485, 203–208. [Google Scholar] [CrossRef]
- Giovannoni, J. Molecular biology of fruit maturation and ripening. Annu. Rev. Plant Physiol. Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.M.; Chin, L.H.; Lazan, H. A comparative study on wall degrading enzymes, pectin modifications and softening during ripening of selected tropical fruits. Plant Sci. 2004, 167, 317–327. [Google Scholar] [CrossRef]
- Gapper, N.E.; McQuinn, R.P.; Giovannoni, J. Molecular and genetic regulation of fruit ripening. Plant Mol. Biol. 2013, 82, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Payasi, A.; Mishra, N.N.; Chaves, A.L.S.; Singh, R. Biochemistry of fruit softening: An overview. Physiol. Mol. Biol. Plants 2009, 15, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Andreou, A.; Feussner, I. Lipoxygenases-structure and reaction mechanism. Biochemistry 2009, 70, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Porta, H.; Rocha-Sosa, M. Plant lipoxygenases: Physiological and molecular features. Plant Physiol. 2002, 130, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.; Heydeck, D.; Hofheinz, K.; Roffeis, J.; O’Donnell, V.B.; Kuhn, H.; Walther, M. Molecular enzymology of lipoxygenases. Arch. Biochem. Biophys. 2010, 503, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Kolomiets, M.V.; Hannapel, D.J.; Chen, H.; Tymeson, M.; Gladon, R.J. Lipoxygenase is involved in the control of potato tuber development. Plant Cell 2001, 13, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Lian, Q.; Xin, H.; Zhong, X.; Zhang, Z.; Li, X.; Yuan, X.; Han, H.; He, X.; Yi, M. Cloning, characterization and expression analysis of a 9-lipoxygenase gene in Gladiolus hybridus. Sci. Hortic. 2011, 130, 468–475. [Google Scholar] [CrossRef]
- Wongshereea, T.; Ketsa, S.; Doorn, W.G. The relationship between chilling injury and membrane damage in lemon basil (Ocimum × citriodourum) leaves. Postharvest Biol. Technol. 2009, 51, 91–96. [Google Scholar] [CrossRef]
- Bhardwaj, P.K.; Kaur, J.; Sobti, R.C.; Ahuja, P.S.; Kumar, S. Lipoxygenase in Caragana jubata responds to low temperature, abscisic acid, methyl jasmonate and salicylic acid. Gene 2011, 483, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Cascón, T.; Vicedo, B.; García-Agustín, P.; Hamberg, M.; Castresana, C. Role of 9-lipoxygenase and α-dioxygenase oxylipin pathways as modulators of local and systemic defense. Mol. Plant 2012, 5, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene CaLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010, 152, 948–967. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Zeng, H.; Hu, Z.; Qv, X.; Chen, G. Overexpression of the tomato 13-lipoxygenase gene TomloxD increases generation of endogenous jasmonic acid and resistance to Cladosporium fulvum and high temperature. Plant Mol. Biol. Rep. 2013, 31, 1141–1149. [Google Scholar] [CrossRef]
- Podolyan, A.; White, J.; Jordan, B.; Winefield, C. Identification of the lipoxygenase gene family from Vitis vinifera and biochemical characterisation of two 13-lipoxygenases expressed in grape berries of Sauvignon Blanc. Funct. Plant Biol. 2010, 37, 67–784. [Google Scholar] [CrossRef]
- Yan, L.; Zhai, Q.; Wei, J.; Li, S.; Wang, B.; Huang, T.; Du, M.; Sun, J.; Kang, L.; Li, C.B.; Li, C. Role of tomato lipoxygenase D in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genet. 2013. [Google Scholar] [CrossRef] [PubMed]
- Padilla, M.N.; Hernández, M.L.; Sanz, C.; Martínez-Rivas, J.M. Molecular cloning, functional characterization and transcriptional regulation of a 9-lipoxygenase gene from olive. Phytochemistry 2012, 74, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, A.; Caldelari, D.; Wolfender, J.L.; Farmer, E.E. Four 13-lipoxygenases contribute to rapid jasmonate synthesis in wounded Arabidopsis thalianaleaves: A role for lipoxygenase 6 in responses to long-distance wound signals. New Phytol. 2013, 197, 566–575. [Google Scholar] [CrossRef]
- Bae, K.; Rahimi, S.; Kim, Y.; Renuka Devi, B.S.; Khorolragchaa, A.; Sukweenadhi, J.; Silva, J.; Myagmarjav, D.; Yang, D. Molecular characterization of lipoxygenase genes and their expression analysis against biotic and abiotic stresses in Panax ginseng. Eur. J. Plant Pathol. 2016, 145, 331–343. [Google Scholar] [CrossRef]
- Yang, X.Y.; Jiang, W.J.; Yu, H.J. The expression profiling of the lipoxygenase (LOX) family genes during fruit development, abiotic stress and hormonal treatments in cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2012, 13, 2481–2500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Jin, Y.Z.; Liu, J.Y.; Tang, Y.F.; Cao, S.X.; Qi, H.Y. The phylogeny and expression profiles of the lipoxygenase (LOX) family genes in the melon (Cucumis melo L.) genome. Sci. Hortic. 2014, 170, 94–102. [Google Scholar] [CrossRef]
- Han, M.; Zhang, T.; Zhao, C.; Zhi, J. Regulation of the expression of lipoxygenase genes in Prunus persica fruit ripening. Acta Physiol. Plant 2011, 33, 1345–1352. [Google Scholar] [CrossRef]
- Mariutto, M.; Duby, F.; Adam, A.; Bureau, C.; Fauconnier, M.L.; Ongena, M.; Thonart, P.; Dommes, J. The elicitation of a systemic resistance by Pseudomonas putida BTP1 in tomato involves the stimulation of two lipoxygenase isoforms. BMC Plant Biol. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Tieman, D.; Bliss, P.; McIntyre, L.M.; Blandon-Ubeda, A.; Bies, D.; Odabasi, A.Z.; Rodríguez, G.R.; van der Knaap, E.; Taylor, M.G.; Goulet, C.; et al. The chemical interactions underlying tomato flavor preferences. Curr. Biol. 2012, 22, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Tieman, D.; Jones, J.B.; Taylor, M.G.; Schmelz, E.; Huffaker, A.; Bies, D.; Chen, K.; Klee, H.J. A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 flavour volatiles in tomato. J. Exp. Bot. 2014, 65, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, K.; Bowen, J.; Allen, A.; Espley, R.; Karunairetnam, S.; Ferguson, I. Differential expression within the LOX gene family in ripening kiwifruit. J. Exp. Bot. 2006, 57, 3825–3836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yin, X.; Li, X.; Yang, S.; Ferguson, I.B.; Chen, K. Lipoxygenase Gene Expression in Ripening Kiwifruit in Relation to Ethylene and Aroma Production. J. Agric. Food Chem. 2009, 57, 2875–2881. [Google Scholar] [CrossRef] [PubMed]
- Rogiers, S.Y.; Kumar, G.M.; Knowles, N.R. Maturation and ripening of fruit of Amelanchier alnifolia Nutt. are accompanied by increasing oxidative stress. Ann. Bot. 1998, 81, 203–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.S.; Zhang, S.L.; Ferguson, I. The role of salicylic acid in postharvest ripening of kiwifruit. Postharvest Biol. Technol. 2003, 28, 67–74. [Google Scholar] [CrossRef]
- Kondo, S.; Meemak, S.; Ban, Y.; Moriguchi, T.; Harada, T. Effects of auxin and jasmonates on 1-aminocyclopropane-1-carboxylate (ACC) synthase and ACC oxidase gene expression during ripening of apple fruit. Postharvest Biol. Technol. 2009, 51, 281–284. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Zheng, P.; Varanasi, V.; Shin, S.B.; Main, D.; Curry, E.; Mattheis, J.P. Multiple plant hormones and cell wall metabolism regulate apple fruit maturation patterns and texture attributes. Tree Genet. Genomes 2012, 8, 1389–1406. [Google Scholar] [CrossRef]
- Davarynejad, G.H.; Zarei, M.; Nasrabadi, M.E.; Ardakani, E. Effects of salicylic acid and putrescine on storability, quality attributes and antioxidant activity of plum cv. “Santa Rosa”. J. Food Sci. Technol. 2015, 52, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chen, P.; Duan, C.R.; Tao, P.; Wang, Y.P.; Ji, K.; Hu, Y.; Li, Q.; Dai, S.J.; Wu, Y.; et al. Transcriptional regulation of genes encoding key enzymes of abscisic acid metabolism during melon (Cucumis melo L.) fruit development and ripening. J. Plant Growth Regul. 2013, 32, 233–244. [Google Scholar] [CrossRef]
- Zaharah, S.S.; Singh, Z.; Symons, G.M.; Reid, J.B. Mode of action of abscisic acid in triggering ethylene biosynthesis and softening during ripening in mango fruit. Postharvest Biol. Technol. 2013, 75, 37–44. [Google Scholar] [CrossRef]
- Jia, H.F.; Chai, Y.M.; Li, C.L.; Lu, D.; Luo, J.J.; Qin, L.; Shen, Y.Y. Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant Physiol. 2011, 157, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.S.; Li, D.D.; Bu, J.W.; Jiang, Y.Y.; Khan, Z.U.; Luo, Z.S.; Mao, L.C.; Ying, T.J. Comprehensive analysis of ABA effects on ethylene biosynthesis and signaling during tomato fruit ripening. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Wang, Y.; Ji, K.; Dai, S.J.; Hu, Y.; Sun, L.; Li, Q.; Chen, P.; Sun, Y.; Duan, C.; et al. The role of abscisic acid in regulating cucumber fruit development and ripening and its transcriptional regulation. Plant Physiol. Biochem. 2013, 64, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Mukkun, L.; Singh, Z. Methyl jasmonate plays a role in fruit ripening of “Pajaro” strawberry through stimulation of ethylene biosynthesis. Sci. Hortic. 2009, 123, 5–10. [Google Scholar] [CrossRef]
- Wasternack, C.; Hanse, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Avanci, N.C.; Luche, D.D.; Goldman, G.H.; Goldman, M.H.S. Jasmonates are phytohormones with multiple functions, including plant defense and reproduction. Genet. Mol. Biol. 2010, 9, 484–505. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi Pirbalouti, A.; Sajjadi, S.E.; Parang, K. A review (research and patent) on jasmonic acid and its derivatives. Arch. Pharm. 2014, 347, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.F.; Zhang, C.; Pervaiz, T.; Zhao, P.C.; Liu, Z.J.; Wang, B.J.; Wang, C.; Zhang, L.; Fang, J.G.; Qian, J.P. Jasmonic acid involves in grape fruit ripening and resistant against Botrytis cinerea. Funct. Integr. Genom. 2016, 16, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Glowacz, M.; Rees, D. Using jasmonates and salicylates to reduce losses within the fruit supply chain. Eur. Food Res. Technol. 2016, 242, 143–156. [Google Scholar] [CrossRef]
- Luo, Z.S.; Chen, C.; Xie, J. Effect of salicylic acid treatment on alleviating postharvest chilling injury of “Qingnai” plum fruit. Postharvest Biol. Technol. 2011, 62, 115–120. [Google Scholar] [CrossRef]
- Kazemi, M.; Aran, M.; Zamani, S. Effect of salicylic acid treatments on quality characteristics of apple fruits during storage. Am. J. Plant Physiol. 2011, 6, 113–119. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; van Wees, S.C.M.; van Pelt, J.A.; Knoester, M.; Laan, R.; Gerrits, N.; Weisbeek, P.J.; van Loon, L.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 1998, 10, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Rao, J.; Zhu, Y.; Chang, X.; Hou, Y.; Zhu, Q. Cloning and expression of lipoxygenase genes and enzyme activity in ripening persimmon fruit in response to GA and ABA treatments. Postharvest Biol. Technol. 2014, 92, 54–61. [Google Scholar] [CrossRef]
- Meng, K.; Hou, Y.L.; Huber, D.J.; Dong, X.Q.; Ban, Q.Y.; Chang, X.X.; Zhang, T.; Rao, J.P. Molecular cloning, structural characterization, and ripening-related expression of lipoxygenase genes from three persimmon cultivars differing in postharvest ripening rate. Plant Mol. Biol. Rep. 2016, 34, 854–868. [Google Scholar] [CrossRef]
- Hou, Y.L.; Meng, K.; Han, Y.; Ban, Q.Y.; Wang, B.; Suo, J.T.; Lv, J.Y.; Rao, J.P. The persimmon 9-lipoxygenase gene DkLOX3 plays positive roles in both promoting senescence and enhancing tolerance to abiotic stress. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic. Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Pastuglia, M.; Roby, D.; Dumas, C.; Cock, J.M. Rapid induction by wounding and bacterial infection of an S gene family receptor-like kinase gene in Brassica oleracea. Plant Cell 1997, 9, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.C.; Howard, E.A.; Dennis, E.S.; Peacock, W.J. DNA sequences required for anaerobic expression of the maize alcohol dehydrogenase 1 gene. Proc. Natl. Acad. Sci. USA 1987, 84, 6624–6628. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Suzuki, A.; Washida, H.; Takaiwa, F. The GCN4 motif in a rice glutelin gene is essential for endosperm-specific gene expression and is activated by Opaque-2 in transgenic rice plants. Plant J. 1998, 14, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Torres, J.T.; Parniske, M.; Wernert, P.; Hahlbrock, K.; Somssich, I.E. Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes. EMBO J. 1996, 15, 5690–5700. [Google Scholar] [PubMed]
- Fink, J.S.; Verhave, M.; Kasper, S.; Tsukada, T.; Mandel, G.; Goodman, R.H. The CGTCA sequence motif is essential for biological activity of the vasoactive intestinal peptide gene cAMP-regulated enhancer. Proc. Natl. Acad. Sci. USA 1988, 85, 6662–6666. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Kim, Y.; An, G. Identification of methyl jasmonate and salicylic acid response elements from nopaline synthase (NOS) promoter. Plant Physiol. 1993, 103, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.; Izawa, T.; Chua, N.H. Plant bZIP proteins gather at ACGT elements. FASEB J. 1994, 8, 192–200. [Google Scholar] [PubMed]
- Aguilera, J.M.; Stanley, D.W. Microstructural Principles of Food Processing and Engineering; An Aspen Publication: New York, NY, USA, 1999; Chapter 1. [Google Scholar]
- Domínguez, E.; Cuartero, J.; Heredia, A. An overview on plant cuticle biomechanics. Plant Sci. 2011, 181, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lara, I.; Belge, B.; Goulao, L.F. The fruit cuticle as a modulator of postharvest quality. Postharvest Biol. Technol. 2014, 87, 103–112. [Google Scholar] [CrossRef]
- Konaraka, A. The structure of the fruit peel in two varieties of Malus domestica Borkh. (Rosaceae) before and after storage. Protoplasma 2013, 250, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Curry, E.A. Growth-induced microcracking and repair mechanisms of fruit cuticles. Plant Physiology. In Proceedings of the SEM Annual Conference, Albuquereque, NM, USA, 1–4 June 2009.
- Veraverbeke, E.A.; Verboven, P.; van Oostveldt, P.; Nicolai, B.M. Prediction of moisture loss across the cuticle of apple (Malus sylvestris subsp. Mitis (Wallr.)) during storage: Part 1. Model development and determination of diffusion coefficients. Postharvest Biol. Technol. 2003, 30, 75–88. [Google Scholar] [CrossRef]
- Riederer, M.; Müller, C. Cuticular transpiration. In Annual Plant Reviews Volume 23: Biology of the Plant Cuticle; Blackwell Publishing Ltd.: Oxford, UK, 2007; Chapter 9. [Google Scholar]
- Maguire, K.M.; Lang, A.; Banks, N.H.; Hall, A.; Hopcroft, D.; Benneti, R. Relationship between water vapour permeance of apples and micro-cracking of the cuticle. Postharvest Biol. Technol. 1999, 17, 89–96. [Google Scholar] [CrossRef]
- Farmaki, T.; Sanmartín, M.; Jiménez, P.; Paneque, M.; Sanz, C.; Vancanneyt, G.; León, J.; Sánchez-Serrano, J.J. Differential distribution of the lipoxygenase pathway enzymes within potato chloroplasts. J. Exp. Bot. 2007, 58, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Creelman, B.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar] [CrossRef] [PubMed]
- Bapat, V.A.; Trivedi, P.K.; Ghosh, A.; Sane, V.A.; Ganapathi, T.R.; Nath, P. Ripening of fleshy fruit: Molecular insight and the role of ethylene. Biotechnol. Adv. 2010, 28, 94–107. [Google Scholar] [CrossRef] [PubMed]
- McAtee, P.; Karim, S.; Schaffer, R.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 16, 4561–4575. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Martinelli, A.P.; Bouchard, R.A.; Finer, J.J. A soybean (Glycine max) polyubiquitin promoter gives strong constitutive expression in transgenic soybean. Plant Cell Rep. 2009, 28, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Sun, Y.; Zhang, M.; Wang, L.; Ren, J.; Cui, M.; Wang, Y.; Ji, K.; Li, P.; Li, Q.; et al. Suppression of 9-cis-epoxycarotenoid dioxygenase, which encodes a key enzyme in abscisic acid biosynthesis, alters fruit texture in transgenic tomato. Plant Physiol. 2012, 158, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yuan, B.; Leng, P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Tareen, M.J.; Abbasi, N.A.; Hafiz, I.A. Postharvest application of salicylic acid enhanced antioxidant enzyme activity and maintained quality of peach cv. ‘Flordaking’ fruit during storage. Sci. Hortic. 2012, 142, 221–228. [Google Scholar] [CrossRef]
- Shafiee, M.; Taghavi, T.S.; Babalar, M. Addition of salicylic acid to nutrient solution combined with postharvest treatments (hot water salicylic acid, and calcium dipping) improved postharvest fruit quality of strawberry. Sci. Hortic. 2010, 124, 40–45. [Google Scholar] [CrossRef]
- Ding, C.K.; Wang, C.Y. The dual effects of methyl salicylate on ripening and expression of ethylene biosynthetic genes in tomato fruit. Plant Sci. 2003, 164, 589–596. [Google Scholar] [CrossRef]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Zapata, P.J.; Castillo, S.; Serrano, M. Postharvest methyl salicylate treatments delay ripening and maintain quality attributes and antioxidant compounds of ‘Early Lory’ sweet cherry. Postharvest Biol. Technol. 2016, 117, 102–109. [Google Scholar] [CrossRef]
- Rao, J.; Ren, X. Store and Transport of Horticulture Production; Shaanxi People’s Publishing House: Xi’an, China, 2003; Chapter 1. [Google Scholar]
- Khaliq, G.; Mohamed, M.T.M.; Ali, A.; Ding, P.; Ghazali, H.M. Effect of gum arabic coating combined with calcium chloride on physico-chemical and qualitative properties of mango (Mangifera indica L.) fruit during low temperature storage. Sci. Hortic. 2015, 190, 187–194. [Google Scholar] [CrossRef]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, P.; Hallett, I.; Redgwell, R.; Benhamou, N.; MacRae, E. Localization of cell wall polysaccharides during kiwifruit (Actinidia deliciosa) ripening. Int. J. Plant Sci. 1999, 160, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Wilkins, T.A. A modified hot borate method significantly enhances the yield of high-quality RNA from cotton (Gossypium hirsutum L.). Anal. Biochem. 1994, 223, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Xu, W.R.; Wang, J.; Wang, L.; Yao, W.K.; Xu, Y.; Ding, J.H.; Wang, Y.J. A core functional region of the RFP1 promoter from Chinese wild grapevine is activated by powdery mildew pathogen and heat stress. Planta 2013, 237, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′–3′) | Purpose |
|---|---|---|
| pLOX3-SP1 | AACTCAGTGTGGTGAAGATTGCGGATG | |
| pLOX3-SP2 | AAGGCGACATCGAAAGCTGAATCTCC | DkLOX3 promoter clone |
| pLOX3-SP3 | CAAGTAAGCTGCCTTCCCAAGCTTCC | |
| DkLOX3-PF | CCCAAGCTTCCACAATTATTCTTTGGTTATTTCG | DkLOX3 full-length promoter clone |
| DkLOX3-PR | GCTCTAGACTTCTTCTTCTTCTTCTTCTTCTGC | |
| DkLOX3-PF5 | CCCAAGCTTTATAGTATGGTCTATCATTCTCATACG | DkLOX3 promoter deletion derivatives construct |
| DkLOX3-PF4 | CCCAAGCTTGTACCCCATGTTTGATACAACTCT | |
| DkLOX3-PF3 | CCCAAGCTTGCATTTCTATGTATGCTCTCTTATG | |
| DkLOX3-PF2 | CCCAAGCTTCTGGATCTAGTTCAACCCATAAG | |
| DkLOX3-PF1 | CCCAAGCTTTTATCAGGGATCTGATTTGTCTTA | |
| DkLOX3qF | CACTGCTCTTCCCTACCA | DkLOX3 qRT-PCR |
| DkLOX3qR | CAGAGGGAGAAATCAGTGATACAC | |
| ActinqF | GGATTCTGGTGATGGTGTTAG | Actin qRT-PCR |
| ActinqR | CAGCAGTTGTTGTGAAGGAGT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, K.; Hou, Y.; Han, Y.; Ban, Q.; He, Y.; Suo, J.; Rao, J. Exploring the Functions of 9-Lipoxygenase (DkLOX3) in Ultrastructural Changes and Hormonal Stress Response during Persimmon Fruit Storage. Int. J. Mol. Sci. 2017, 18, 589. https://doi.org/10.3390/ijms18030589
Meng K, Hou Y, Han Y, Ban Q, He Y, Suo J, Rao J. Exploring the Functions of 9-Lipoxygenase (DkLOX3) in Ultrastructural Changes and Hormonal Stress Response during Persimmon Fruit Storage. International Journal of Molecular Sciences. 2017; 18(3):589. https://doi.org/10.3390/ijms18030589
Chicago/Turabian StyleMeng, Kun, Yali Hou, Ye Han, Qiuyan Ban, Yiheng He, Jiangtao Suo, and Jingping Rao. 2017. "Exploring the Functions of 9-Lipoxygenase (DkLOX3) in Ultrastructural Changes and Hormonal Stress Response during Persimmon Fruit Storage" International Journal of Molecular Sciences 18, no. 3: 589. https://doi.org/10.3390/ijms18030589