
Function and Clinical Implications of Long Non-Coding RNAs in Melanoma
by
 ,
,
Georg Richtig
1,2 ,
,
Barbara Ehall
3,4,
Erika Richtig
2,
Ariane Aigelsreiter
3,
Tony Gutschner
5,* and
Martin Pichler
6,* 1
Institute of Experimental and Clinical Pharmacology, Medical University of Graz, Graz 8010, Austria
2
Department of Dermatology, Medical University of Graz, Graz 8036, Austria
3
Institute for Pathology, Medical University of Graz, Graz 8036, Austria
4
Division of Hematology, Department of Internal Medicine, Medical University of Graz, Graz 8036, Austria
5
Faculty of Medicine, Martin-Luther-University Halle-Wittenberg, Halle (Saale) 06120, Germany
6
Division of Clinical Oncology, Department of Medicine, Medical University of Graz, Graz 8036, Austria
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(4), 715; https://doi.org/10.3390/ijms18040715
Submission received: 19 February 2017
/
Revised: 16 March 2017
/
Accepted: 17 March 2017
/
Published: 28 March 2017
(This article belongs to the Collection Regulation by Non-coding RNAs)
Abstract
:Metastatic melanoma is the most deadly type of skin cancer. Despite the success of immunotherapy and targeted agents, the majority of patients experience disease recurrence upon treatment and die due to their disease. Long non-coding RNAs (lncRNAs) are a new subclass of non-protein coding RNAs involved in (epigenetic) regulation of cell growth, invasion, and other important cellular functions. Consequently, recent research activities focused on the discovery of these lncRNAs in a broad spectrum of human diseases, especially cancer. Additional efforts have been undertaken to dissect the underlying molecular mechanisms employed by lncRNAs. In this review, we will summarize the growing evidence of deregulated lncRNA expression in melanoma, which is linked to tumor growth and progression. Moreover, we will highlight specific molecular pathways and modes of action for some well-studied lncRNAs and discuss their potential clinical implications.
1. Introduction
Although melanoma only represents a small fraction of all skin cancer types, it is responsible for the majority of skin cancer related deaths [1]. Therefore, it is the most lethal cutaneous neoplasm. Unfortunately, melanoma incidence is rising worldwide and despite the development of new treatment options, metastatic disease in melanoma is still associated with high rates of mortality [2]. Prognoses of melanoma patients depend on the tumor thickness, ulceration and metastatic spread, with the 10-year survival varying from 93% to 39%, respectively [3]. The major risk factor for melanoma is ultraviolet (UV) radiation, which is responsible for a characteristic genetic signature [4]. Effectively mutational rate in melanoma is the highest within all cancer types and considered responsible for the success of immunotherapy [4]. The two main pathways of interest in melanoma nowadays are the PI3K/PTEN/Akt/mTOR signaling pathway and the RAS/RAF/MEK/ERK signal transduction cascade (Mitogen-activated protein kinase (MAPK) Signaling Pathway) [5]. The most common genetic mutation suitable for targeted therapy within the MAPK pathway, is located in the B-Raf proto-oncogene, serine/threonine kinase (BRAF) accounting for approximately 40–60% of all melanomas [6,7]. The mutation occurs at codon 600 of the BRAF gene in approximately 95% of all cases, the most common exchange being a valine to glutamic acid referred as BRAFV600E mutation [8]. The majority of BRAF mutations lead to a hyper-activation of the MAPK pathway, resulting in enhanced cell growth and survival [9,10]. Usually Rat sarcoma (RAS) activates Rapidly Accelerated Fibrosarcoma (RAF), which propagates down-stream signaling through MAPK/ERK Kinase (MEK) to extracellular signal-regulated kinase (ERK). When a high-kinase activity mutation in BRAF occurs, it can independently activate the MAPK pathway and in fact BRAFV600E has a ~500-fold increased activity compared to BRAFwt [11].
The second most common mutation found in melanoma is affecting the NRAS proto-oncogene, GTPase (NRAS) gene with a frequency of ~20% [7,12]. Therefore, the MAPK pathway is one of the major oncogenic drivers in melanoma. However, recent studies revealed additional mutations involved in growth and progression of malignant melanoma, e.g., KIT proto-oncogene receptor tyrosine kinase (KIT) gene in mucosal melanoma, telomerase reverse transcriptase (TERT) gene, germline cyclin dependent kinase inhibitor 2A (CDKN2A) gene, tumor protein p53 (TP53) gene, neurofibromin 1 (NF1) gene and others [13,14,15].
1.1. Therapeutic Modalities
Currently, cutaneous melanoma are classified into four subgroups: BRAFmt, NRASmt, NF1mt and triple wild type [16,17]. Therefore, patients with metastatic diseases have three pharmacological therapeutic options: (i) immunotherapy; (ii) targeted therapy, focusing on the mutational status of melanoma cells; and (iii) conventional chemotherapy, if the first two options are not suitable or available [18,19,20,21,22].
1.1.1. Targeted Therapy
BRAF Mutated Melanoma
There are two BRAF inhibitors, which are widely used for systemic treatment in metastatic melanoma: vemurafenib and dabrafenib [23,24]. In the BRAF Inhibitor in Melanoma 3 (BRIM-3) study, vemurafenib was more effective in BRAFV600E mutated melanoma compared to conventional chemotherapy (dacarbazine and DTIC) [21]. Similar results could be obtained when another BRAF inhibitor, dabrafenib, was used in patients with metastatic melanoma and further BRAF inhibitors (e.g., encorafenib) are now in clinical trials [25,26]. However, a major problem of targeted therapy is the emergence of receptor tyrosine kinase (RTK) mutations upon treatment with BRAF inhibitors [27].
Another common side effect of BRAF inhibitors is the development of new BRAFwt and RASmt melanoma, which may be explained by a paradoxical activation of the MAPK pathway in BRAFwt melanoma [28,29]. To increase the efficiency of MAPK pathway blockage, MEK inhibitors have been developed in combination with BRAF inhibitors [30,31,32]. Effectively, dual inhibition has improved progression-free survival, overall survival and response rates [19,33]. Therefore, dual inhibition is now considered the standard of care in BRAFV600 mutated melanoma.
NRAS Mutated Melanoma
Approximately twenty percent of all melanomas have a hotspot mutation in the NRAS gene [34]. Due to its downstream signaling to the MAPK pathway and the knowledge that BRAF inhibitors lead to enhanced growth of BRAFwt melanomas, several trials investigated the efficiency of MEK inhibition in this subset of melanoma [29]. However, the response rate to MEK inhibitors is variable, suggesting that NRAS signaling does not solely act on the MAPK pathway [35]. In the literature, new mutations in the MAPK pathway and other pathways involved in proliferation and growth have been suggested as putative escape mechanism, but it might also be possible that there are some epigenetic modifications involved [36].
NF1 Mutated Melanoma
The Cancer Genome Atlas proposed that NF1 mutations should be included for classification in BRAFwt/NRASwt melanoma, since reports were available showing that mutations in NF1 led to hyper-activation of the MAPK pathway and were responsive to MEK inhibition [16,37,38]. On the downside, another study suggested loss of NF1 mediates resistance to MEK inhibition (selumetinib) [39]. However, Krauthammer and colleagues showed that not all NF1 mutated melanoma were responsive to MEK inhibition and that many NF1 mutated melanoma had concurrent mutations in RAS related genes [40]. Therefore it seems that there might be some patients with NF1mt melanoma suitable for MEK inhibitor treatment, although further studies are necessary to elucidate the role of NF1.
KIT Mutated Melanoma
The KIT proto-oncogene receptor tyrosine kinase gene is frequently mutated in acral and mucosal melanoma as well as in melanomas of chronically sun-damaged skin [41]. KIT acts as a bona fide oncogene, resulting in higher cell proliferation, cell migration and progression through the activation of the MAPK pathway and the PI3K/mTOR pathway [42,43]. Imatinib-a receptor tyrosine kinase inhibitor-showed high response rates and beneficial effects in patients harboring a melanoma with mutations in the exons 11 and 13 of the KIT gene [44]. In addition, nilotinib, a tyrosine kinase inhibitor used in imatinib-resistant chronic myeloid leukaemia, as well as sunitinib, provided promising results in the treatment of KIT mutated metastatic melanoma [45,46]. However, it has to be mentioned that this is confined to a subset of an already small subpopulation of patients.
1.1.2. Immunotherapy
The second pharmacological option in metastatic melanoma nowadays is immunotherapy. Boosting the immune system of patients with metastatic melanoma has a long history despite low success rates [47].
However, the first immunotherapy approved for metastatic melanoma was Ipilimumab in 2011 [48]. Ipilimumab improved the overall survival of patients with advanced melanoma (compared to dacarbazine) and showed a durable long-term survival of approximately 20% [49]. However, one of the major disadvantages of cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) monoclonal antibody (mAb) was the low response rate of ~20%. A major contribution to immunotherapy was the discovery of the immune evasion mechanism, mediated through the expression of B7-H1 on the surface of melanoma cells, which was later described as programmed cell death ligand-1 (PD-L1) [50].
PD-L1 was able to suppress in vivo activated T-cells, which in turn reduced the immune response. When PD-L1 was blocked by a mAb, immune response was restored and a significant reduction in tumor size was achieved [50,51]. In 2012 nivolumab-a mAb against programmed cell death 1 (PD-1)-improved progression-free survival, overall survival, demonstrated an increased response rate and could achieve an up to 35% long term survival in patients with metastatic melanoma [52,53]. However, the response rate was raised up to 53%, when ipilimumab was added to nivolumab [54]. Although the rate of adverse side effects was rising as suspected, it was manageable [55].
Immunotherapy is now a major field of anti-cancer treatment and is evolving at high speed, with new substances constantly being tested. There are ongoing clinical trials for mAb against T-cell immunoglobulin and mucin-domain containing-3 (TIM-3), OX40 and CD73 [56,57]. However, similar to targeted therapy, a significant proportion of patients developed resistance upon treatment, whereas the mechanism(s) are still widely unknown.
2. Long Non-Coding RNAs
The ENCyclopedia of DNA Elements (ENCODE) project revealed that a surprisingly large fraction (70%–90%) of the human genome is transcribed into RNA. However, only 1%–3% of the transcriptome carries the blueprint for the synthesis of proteins, leaving the question whether or not the remaining non-coding RNA (ncRNA) transcripts are just “nature’s trash” [58,59].
NcRNAs can be classified into small ncRNAs (<200 nucleotides (nt)) and long ncRNAs (lncRNAs), depending on their size. Multiple types of small ncRNA (microRNAs (miRNAs), small interfering RNAs (siRNAs) and PIWI-interacting RNAs (piRNAs)) have been studied extensively, especially their effects on cancer development and their involvement in regulation of immune checkpoint pathways [60,61,62,63]. Long ncRNAs represent a highly heterogeneous group of RNAs, which have an extensive variability in their cellular effects, as well as their molecular influences. They can be classified by their length (>200 nt) and by their lack of a functional open reading frame, meaning they encompass less than 100 amino acids [64,65,66,67]. Hence, lncRNAs constitute a very heterogeneous group of RNA molecules, which allows them to cover a broad spectrum of molecular and cellular functions by implementing different modes of action [68,69,70,71]. From an evolutionary perspective, circumventing the energy intensive protein translation by regulating various processes through ncRNAs seems quite reasonable [72]. This regulation by lncRNA can occur via multiple mechanisms, which Wang et al. divided into four types, which we will discuss here briefly [73].
2.1. Signal LncRNA
lncRNAs are generally transcribed by polymerase II and their expression and stimulus response are very cell type specific, indicating a strong transcriptional control [74]. Since they are under transcriptional control, they can be seen as a signal able to detect the chromatin state of regulatory elements, or simply, the expression of associated genes. An interesting point to consider is that the cell saves resources and time by producing regulatory RNAs and circumventing protein translation. This category sees lncRNAs as a signal of gene expression patterns and is associated with time, location and developmental state. One lncRNA in this category, which is regulated by external stimuli—namely, DNA damage—is LincRNA-p21, which plays a key regulatory role in p53 transcriptional response. It acts as a transcriptional repressor of the p53 pathway. p53 directly regulates LincRNA-p21 expression by binding to the promotor of LincRNA-p21 [75].
2.2. Decoy LncRNA
One major mechanism of transcription regulation involves lncRNAs acting as decoys, by binding to various transcription factors, chromatin modifiers and other regulatory factors, to prevent them from executing their designated function [76]. One described mode of action involves lncRNAs acting as “miRNA sponges”, by sequestering and thus down-regulating respective miRNAs. Consequently target genes of these miRNAs are influenced significantly in their expression [77]. Experiments involving gene knockout of lncRNA should therefore indicate increased effects of the speculated bound effector molecule [78]. For example, depletion of Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) in tumor cells resulted in a reduction of tumorigenicity, while transient overexpression induced tumor proliferation and formation [79]. Based on studies by Tripathi et al. double knockdown of lncRNA MALAT1 and its suspected effectors with siRNA resulted in a rescue phenotype [80].
2.3. Guide LncRNA
Guide lncRNAs are categorized by their ability to bind proteins and afterwards direct the complex to a specific location. Gene expression changes can occur either in cis (on neighboring genes) or in trans (more distant genes), although this is impossible to predict based solely on the lncRNA sequence [81]. Mechanisms in cis could include a co-transcriptional chromatin change alongside the RNA polymerase, or serve as complementary targets for small regulatory RNAs. The in trans guidance is thought to be based upon lncRNAs binding to target DNA as heteroduplexes or RNA:DNA:DNA triplexes, or even RNA recognition sites on the surface of specific chromatin features. Whatever the mechanism, the result of lncRNA guidance is to regulate and bring about epigenetic changes in target genes [82].
This class/archetype encompasses lncRNAs such as Homebox (HOX) transcript antisense RNA (HOTAIR) and would be characterizable in knockdown lncRNA experiments by the missing functionality of the effector, due to its malfunctioning localization or even a phenotype resembling an effector knockout. In comparison to the decoy archetype, the guide archetype would exhibit an exacerbated phenotype instead of a rescue.
2.4. Scaffold LncRNA
Until now, it was thought that only proteins play a key role in scaffolding complexes and therefore the control of intermolecular interactions and signaling [83]. Recent evidence has shown, that lncRNAs may possess similar functions [84,85]. The lncRNA in this class is characterized by its ability to bind multiple effector molecules over specific domains, which brings them closer together and supports their specific functions, such as activation or repression of gene transcription. By furthering our understanding of these complex interactions and their regulation, we would then be able to exploit these mechanisms to influence cells according to our needs. This archetype would be characterizable in knockdown of the lncRNA by malfunctioning of the involved pathway or even a loss of function due to interference in the lncRNA-effector-scaffold assembly. Any further knockout of effectors would likely exacerbate the phenotype instead of saving it. When manipulating the specific lncRNA domains, there might be effects on different effectors and functions [73]. One scaffold lncRNA is the antisense lncRNA in INK4 locus (ANRIL), which directly interacts with polycomb repressive complexes (PRC1 and PRC2). Interaction with ANRIL interrupts transcriptional repression of the INK4b locus [73,86].
2.5. Multifunctionality and Novel Mechanisms
Classification of one lncRNA into multiple archetypes is not unlikely, as already described for cold induced long antisense intragenic RNA (COOLAIR) and Homebox A (HOXA) transcript at the distal tip (HOTTIP), which both fall into the signal as well as the guide class [87]. HOX transcript antisense RNA (HOTAIR) is also a multifunctional lncRNA transcribed in distal and posterior cells, thus being an anatomically specific signal, which is also involved in both PRC2 and Lysine-specific histone demethylase 1 (LSD1) complex assembly, therefore fitting the scaffold category and the localization of PRC2 as a guide [88].
However, ongoing research efforts will very likely identify lncRNAs with new modes of action that cannot be included in the aforementioned subtypes. For example, a new mechanism how lncRNA can affect metabolism and regulatory processes has recently been described by Liu and colleagues [89]. They demonstrated that the lncRNA neighbor of BRCA1 gene 2 (NBR2) directly targets the adenosine monophosphate-activated protein kinase (AMPK) during energy-stress periods. Furthermore, NBR2 depletion led to altered apoptosis/autophagy and unchecked cell cycling with increased tumor development in vivo [89,90].
These examples highlight the diverse and complex biological functions that can be executed or mediated by lncRNAs. Consequently, lncRNA deregulation can have a severe impact on cellular behavior and is often found in human diseases, especially cancer. In the following paragraph, we highlight examples of lncRNAs, whose expression is altered in human melanoma and which have been shown to functionally contribute to melanoma development and progression.
3. Long Non-Coding RNAs in Melanoma
3.1. ANRIL
In the well-studied INK4 locus the interplay between ANRIL and chromatin-modifying complexes can be observed, in which ANRIL serves as a scaffold lncRNA. ANRIL was found in the INK4B/ARF/INK4A locus, has 19 exons and spans 126.3 kb. Due to alternative splicing, several long, short and circular isoforms of ANRIL exist [91]. ANRIL expression has been linked to several conditions, including the risk of melanoma [92,93].
It has been proposed that ANRIL negatively regulates INK4b/ARF/INK4a in cis through chromatin remodeling. It achieves this by binding to PRC1 and PRC2, which in turn controls lysine 27 methylation of histone H3 in the INK4B/ARF/INK4A tumor suppressor locus (Figure 1A) [94]. This implies that ANRIL is significantly involved in cell proliferation and furthermore in cell proliferation after DNA damage repair (see Table 1) [94,95]. Xu et al. indicated that ANRIL was overexpressed in cutaneous melanoma and uveal melanoma compared to normal tissue [96]. Knockdown of ANRIL by siRNA restored the ability of two tumor cell lines (A375 and OM431) to transcribe INK4A and INK4B. This reduced the cell’s ability to migrate and form colonies and ANRIL might therefore be a valid therapeutic target [96].
3.2. BANCR
BRAF-activated non-coding RNA (BANCR) is a 693nt lncRNA, which is encoded by chromosome 9 and acts as a decoy lncRNA [97]. BANCR is notable, because it is highly upregulated in human primary malignant melanoma and induced by BRAFV600E in comparison to BRAFwt melanoma. Depletion experiments demonstrated that BANCR has a regulatory function in melanoma cell migration, whereas its absence significantly decreases cellular migration. In BANCR depleted cells CXCL11 could be identified as a factor simultaneously down-regulated. When the authors supplied CXCL11 to BANCR deficient cells, CXCL11 was capable of rescuing and restoring the migratory abilities of BANCR-depleted cells [97].
Another study revealed that BANCR expression directly correlates with tumor stage and might contribute to the development of melanoma [98]. Li and colleagues demonstrated that BANCR can activate ERK1/2, its upstream molecule CRAF and JNK in-vitro and in-vivo, which led to proliferation of melanoma cells (summarized in Figure 1A and Table 1). They concluded that the link between these pathways indicates a novel regulation mechanism in melanoma proliferation [98].
3.3. CASC15
The lncRNA cancer susceptibility candidate 15 (CASC15) spans ~530 kb and is located on chromosome 6 between the SOX4 and PRL genes [99]. It was frequently expressed in metastatic melanoma cell lines independent of their BRAF mutational status and was absent in normal melanocytes. Furthermore, brain metastases showed significantly higher CASC15 expression levels compared to the cutaneous cell lines [99]. Intriguingly, CASC15 expression in patient-derived FFPE samples from brain and lung metastases was found to be significantly higher compared to normal tissue and naevi. Analysis of the 10-year disease-free survival rates and CASC15 expression in stage III melanoma lymph node metastases revealed that patients with high CASC15 expression had a significantly reduced DFS. Additional in vitro experiments could show that CASC15 regulates melanoma cell phenotype switching between proliferative and invasive states (Table 1) [99].
3.4. GAS5
The lncRNA growth arrest-specific transcript 5 (GAS5) is located on chromosome 1 and consists of 650 bases (12 exons) [100]. Chen et al. investigated the functional role of GAS5 in melanoma and revealed that some cell lines had a reduced expression of GAS5. Moreover, GAS5-depleted cells showed a higher ability to migrate, whereas induced overexpression in such cells reduced their migratory ability and went along with decreased levels of matrix metalloproteinase (MMP) 2 protein production (Figure 1B and Table 1) [101].
3.5. HOTAIR
The lncRNA HOTAIR is transcribed from the HOXC cluster and regulates the transcription of the HOXD cluster (including HOXD8, HOXD9, HOXD10, and HOXD11) located on chromosome 2 [87].
HOTAIR expression was found to be significantly higher in lymph node metastases compared to primary melanoma, whereas several other lncRNAs, including MALAT1, Urothelial carcinoma-associated 1 (UCA1), and nuclear-enriched transcript 1 (NEAT1), showed no change in their expression patterns [102]. This result supports the idea that HOTAIR contributes to the metastatic behavior in melanoma, which is in line with findings from other groups that established HOTAIR as a crucial regulator of metastases in several cancer types [88,103]. In fact, HOTAIR knockdown experiments led to a decreased melanoma cell motility and invasiveness in conjunction with a reduced capability to degrade the extracellular matrix [102].
3.6. Llme23
Llme23 was found exclusively in human melanoma cell lines and it was shown to act as a decoy lncRNA, binding to the protein associated splicing factor (PSF), a known tumor suppressor [105]. Competitive binding of Llme23 to PSF prevents this negative regulatory protein from binding to the promotor of the proto-oncogene RAB23, a RAS-related small GTPase (Figure 1A and Table 1). Importantly, expression levels of RAB23 and Llme23 have been reported to be concordant [106].
3.7. MALAT1
MALAT1 is also known as nuclear-enriched transcript 2 (NEAT2) and has a length of ~8000 nt. It was discovered as a prognostic marker for lung cancer metastasis, although it has also been linked to multiple other human tumors [107,108]. MALAT1 knockdown resulted in impaired melanoma migration, implying possible effects on tumor dissemination. Patient-derived melanoma samples demonstrated a significantly higher MALAT1 expression in lymph node metastases, compared to the primary tumor and to adjacent tissue [109].
The molecular mode of action of MALAT1 is still not fully understood, but might involve transcriptional and epigenetic mechanisms [79,108,110]. In addition, recent studies suggest that MALAT1 may act as a competing endogenous RNA (ceRNA) by binding to tumor-suppressive miRNAs [111]. For example, MALAT1 acts as decoy lncRNA by targeting miR-22. Its depletion leads to increased cell migration and proliferation and miR-22 levels inversely indirectly correlate with MALAT1 levels. Functionally, miR-22 binds to MMP14 and SNAIL and suppresses their oncogenic function [112]. In uveal melanoma, Sun et al. could demonstrate that MALAT1 plays a similar role by targeting miR-140, which was down-regulated in tumor samples compared to normal tissue (summarized in Table 1) [113].
3.8. PAUPAR
The lncRNA PAX6 upstream antisense RNA (PAUPAR) was found in uveal melanoma tissues and uveal melanoma cell lines at low levels, suggesting that it might act as a tumor suppressor lncRNA [114]. Effectively it impacts tumorigenesis in vitro and in vivo by reducing tumor metastases significantly. Further experiments showed that PAUPAR acts as a guide lncRNA by inducing the silencing of the transcription factor hairy and enhancer of split-1(HES1) by inhibiting histone H3K4 tri-methylation at the HES1 locus (Figure 1B and Table 1) [114].
3.9. RMEL3
RMEL3 was first described together with RMEL1 and RMEL2 by Sousa et al. who revealed that all RMEL lncRNAs were almost exclusively expressed in melanocytes and melanoma [115]. Interestingly, RMEL3 was significantly higher in BRAFmt melanoma compared to triplewt (RAS/BRAF/NF1) melanoma and expression levels were negatively correlated with melanoma progression [115].
However, RMEL3 experiments could not confirm these clinical findings and suggested that RMEL3 plays a pro-oncogenic role, since knockdown experiments resulted in a 95% decrease of colony formation in different BRAFV600E melanoma cell lines. Additionally, MAPK and PI3K pathway activators and effectors were also impacted negatively. Multiple genes for these activators and effectors were found to be correlated with RMEL3, thus indicating the requirement of RMEL3 for MAPK and PI3K signaling (see Figure 1A). Because RMEL3 depletion decrease cell survival and proliferation in BRAFV600E melanoma cell lines, it might represent a potential therapeutic target gene in this subset of melanomas (summarized in Table 1) [116].
3.10. SAMMSON
Survival associated mitochondrial melanoma-specific oncogenic non-coding RNA (SAMMSON) is located 30 kb downstream of the melanoma-specific oncogene melanogenesis associated transcription factor (MITF) and is co-amplified in around 10% of all melanoma cases, although studies demonstrated that SAMMSON acts in trans as a decoy lncRNA by targeting p32. In melanoma, SAMMSON is the target of the melanoblast/melanoma-specific transcription factor SOX10 and its co-factor (Figure 1C). Expression of SAMMSON was detectable in over 90% of human primary melanoma and metastasis, whereas SAMMSON was undetectable in normal healthy tissue. Knockdown experiments established a role for this lncRNA in melanoma cell viability and growth, irrespective of their mutational status (BRAF, NRAS, TP53) [117]. By targeting SAMMSON, cell sensitivity towards MAPK-targeting therapeutics could be enhanced and MAPK-resistant cell lines were still susceptible to SAMMSON targeting (summarized in Table 1) [117].
3.11. SNHG5
SnoRNA host gene 5 (SNHG5) is part of the non-coding multiple small nucleolar RNA host gene family and encompasses 524 base pairs [118]. The site is known to be involved in human β-cell lymphoma [118]. Serum levels of SNGH5 were significantly increased in malignant melanoma patients of all stages compared to normal subjects and patients with squamous cell carcinoma. This indicates that SNGH5 may play a role in melanoma genesis (see Table 1) [119].
3.12. SPRY4-IT1
SPRY4 intronic transcript 1 (SPRY4-IT1; also known as SPRIGHTLY) is initiated in intron 1 of the SPRY4 gene and extends to exon 3 [120]. Even though SPRY4-IT1 is located within an intron of SPRY4, both genes were found to be functionally and transcriptionally independent [121].
SPRY4-IT1 was shown to be over-proportionally represented in melanoma cell lines as well as in human melanoma samples compared to melanocytes [122]. It is mainly located in the cytoplasm of melanoma cells and was shown to be associated with polysomes [120,121]. Furthermore, SPRY4-IT1 reduces the abundance of the lipid phosphatase lipin 2. Hence, it may impair apoptosis, because of lipin 2-mediated alterations in lipid metabolism and the resulting lipotoxicity (see Figure 1C) [120]. Additional studies reported a decreased cell growth, invasion and differentiation, but increased apoptosis in SPRY4-IT1-depleted melanoma cells [120,121,123]. Zhao et al. demonstrated that SPRY4-IT1 expression is lower in melanocytes compared to melanoma. Interestingly, one month after ectopic overexpression of SPRY4-IT1 in melanocytes induced dendritic-like cell morphologies together with enlarged nuclei in 25% of the cells. These melanocytes were more proliferative, invasive, and formed anchorage-independent colonies, suggesting that SPRY4-IT1 plays an important role in melanomagenesis and progression [122]. Melanoma patients had significantly higher SPRY4-IT1 expression in comparison to healthy patients and furthermore higher SPRY4-IT1 levels were associated with lower overall survival as well as higher tumor stage (summarized in Table 1) [123].
3.13. UCA1
UCA1 was first discovered in bladder transitional cell carcinoma and so far two isoforms have been identified [124].
A study comparing the expression of six cancer-implicated lncRNAs in melanoma to their respective expression in paired adjacent healthy tissue, found elevated expression of UCA1 in melanoma, especially at advanced stages [109,125]. Knockdown of UCA1 inhibited melanoma cell migration suggest that it might be involved in tumor dissemination [109]. In fact, UCA1 might act as a decoy lncRNA targeting miR-507, an inhibitor of the pro-oncogenic transcription factor FoxM1 (Figure 1A). Depletion of UCA1 increased the levels of miR-507 and reduced FoxM1 levels which led to cell cycle progression defects [125]. In turn, this led to decreased cell proliferation through G1 cell cycle arrest (functionally summarized in Table 1).
4. Conclusions and Outlook
Long noncoding RNAs are now widely recognized as contributing factors which play diverse and complex roles in cancer. Moreover, they are gaining increasing attention as potential biomarkers and represent a novel class of target molecules. However, we are only beginning to understand the complexity of tumorigenic processes and the role of lncRNAs in melanoma as well as in other cancer types. The clinical integration of lncRNAs as prognostic and predictive biomarkers in conjunction with additional cancer targets, could provide a chance to increase the therapeutic benefit.
Nevertheless, several challenges lie ahead of us. First of all, we have to learn more about the molecular mechanisms and processes employed and controlled by lncRNAs. This task is not trivial and requires multiple techniques as well as the development of novel methods that allow us to observe and capture the lncRNAs and their respective interaction partners and sites in a highly specific manner. For example, in addition to our current understanding that lncRNAs exert their biological effect through interaction with DNA/RNA or proteins, it might be possible that lncRNAs directly interact with other metabolites, such as lipids or sugars [126]. Identifying and studying these lncRNAs will require novel technologies and an open mind.
Secondly, the in vivo function of lncRNAs is difficult to study, due to the low conservation of most lncRNAs [127]. Additionally, multiple strategies have to be considered to generate knockout mouse models for lncRNAs [128]. Once established, these models should be combined with melanoma mouse models to investigate the functional relevance and molecular mechanisms of the lncRNA under investigation.
Finally, these novel animal models could represent valuable tools to develop effective therapeutic reagents against lncRNAs, which are currently considered difficult to target. Reducing the expression or blocking the function of oncogenic lncRNAs with, e.g., small molecules, might pave the way to novel treatment strategies and their clinical application in the future.
Acknowledgments
Georg Richtig received funding from the Austrian Science Fund FWF (W1241) and the Medical University Graz through the PhD Program Molecular Fundamentals of Inflammation (DK-MOLIN). We gratefully thank Jessica Damberger for linguistic help.
Author Contributions
Georg Richtig wrote the article; Barbara Ehall contributed to the article; Erika Richtig contributed her expert knowledge about melanoma biology; Ariane Aigelsreiter contributed her knowledge about molecular pathology; Tony Gutschner and Martin Pichler provided valuable insights about long non-coding RNA biology as well as conceptual advice and helped writing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AMPK | adenosine monophosphate-activated protein kinase |
| ANRIL | antisense lncRNA in INK4 locus |
| BANCR | BRAF-activated non-coding RNA |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase |
| BRIM-3 | BRAF Inhibitor in Melanoma 3 |
| CASC15 | cancer susceptibility candidate 15 |
| CDKN2A | cyclin dependent kinase inhibitor 2A |
| ceRNA | competing endogenous RNA |
| COOLAIR | cold induced long antisense intragenic RNA |
| CTLA-4 | cytotoxic T-lymphocyte-associated protein 4 |
| DTIC | dacarbazine |
| ENCODE | ENCyclopedia of DNA Elements |
| ERK | extracellular signal-regulated kinase |
| GAS5 | growth arrest-specific transcript 5 |
| HES1 | hairy and enhancer of split-1 |
| HOTAIR | HOX transcript antisense RNA |
| HOTTIP | HOXA distal transcript antisense RNA |
| HOX | homebox |
| HOXA | homebox A |
| KIT | KIT proto-oncogene receptor tyrosine kinase |
| lncRNA | long non-coding RNA |
| LSD1 | lysine-specific histone demethylase 1 |
| mAb | monoclonal antibody |
| MALAT1 | metastasis-associated lung adenocarcinoma transcript 1 |
| MAPK | mitogen-activated protein kinase |
| MEK | MAPK/ERK Kinase |
| miRNA | microRNA |
| MITF | melanogenesis associated transcription factor |
| MMP | matrix metalloproteinase |
| NBR2 | lncRNA neighbor of BRCA1 gene 2 |
| ncRNA | non-coding RNA |
| NEAT | nuclear-enriched transcript |
| NF1 | neurofibromin 1 |
| NRAS | neuroblastoma RAS viral (v-ras) oncogene homolog |
| nt | nucleotides |
| PAUPAR | PAX6 upstream antisense RNA |
| PD-1 | programmed cell death 1 |
| PD-L1 | programmed cell death ligand-1 |
| piRNA | PIWI-interacting RNA |
| PRC | polycomb repressive complexe |
| PSF | protein associated splicing factor |
| RAF | rapidly Accelerated Fibrosarcoma |
| RAS | rat sarcoma |
| RTK | receptor tyrosine kinase |
| SAMMSON | survival associated mitochondrial melanoma-specific oncogenic non-coding RNA |
| siRNA | small interfering RNA |
| SNHG5 | SnoRNA host gene 5 |
| SPRY4-IT1 | SPRY4 intronic transcript 1 |
| TERT | telomerase reverse transcriptase |
| TIM-3 | T-cell immunoglobulin and mucin-domain containing-3 |
| TP53 | tumor protein p53 |
| UCA1 | urothelial carcinoma-associated 1 |
| UV | ultraviolet |
References
- Deeks, E.D. Nivolumab: A review of its use in patients with malignant melanoma. Drugs 2014, 74, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Richtig, G.; Richtig, E.; Massone, C.; Hofmann-Wellenhof, R. Analysis of clinical, dermoscopic and histopathological features of primary melanomas of patients with metastatic disease—A retrospective study at the Department of Dermatology, Medical University of Graz, 2000–2010. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1776–1781. [Google Scholar] [CrossRef] [PubMed]
- Balch, C.M.; Gershenwald, J.E.; Soong, S.J.; Thompson, J.F.; Atkins, M.B.; Byrd, D.R.; Buzaid, A.C.; Cochran, A.J.; Coit, D.G.; Ding, S.; et al. Final version of 2009 AJCC melanoma staging and classification. J. Clin. Oncol. 2009, 27, 6199–6206. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational heterogeneity in cancer and the search for new cancer-associated genes. Nature 2013, 499, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Masliah-Planchon, J.; Garinet, S.; Pasmant, E. RAS-MAPK pathway epigenetic activation in cancer: miRNAs in action. Oncotarget 2015, 7, 38892–38907. [Google Scholar]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Colombino, M.; Capone, M.; Lissia, A.; Cossu, A.; Rubino, C.; De Giorgi, V.; Massi, D.; Fonsatti, E.; Staibano, S.; Nappi, O.; et al. BRAF/NRAS mutation frequencies among primary tumors and metastases in patients with melanoma. J. Clin. Oncol. 2012, 30, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Long, G.V.; Menzies, A.M.; Nagrial, A.M.; Haydu, L.E.; Hamilton, A.L.; Mann, G.J.; Hughes, T.M.; Thompson, J.F.; Scolyer, R.A.; Kefford, R.F. Prognostic and clinicopathologic associations of oncogenic BRAF in metastatic melanoma. J. Clin. Oncol. 2011, 29, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed]
- Richtig, G.; Aigelsreiter, A.; Kashofer, K.; Talakic, E.; Kupsa, R.; Schaider, H.; Richtig, E. Two case reports of rare BRAF mutations in exon 11 and exon 15 with discussion of potential treatment options. Case Rep. Oncol. 2016, 9, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Richtig, G.; Hoeller, C.; Kashofer, K.; Aigelsreiter, A.; Heinemann, A.; Kwong, L.N.; Pichler, M.; Richtig, E. Beyond the BRAFV600E hotspot-biology and clinical implications of rare BRAF gene mutations in melanoma patients. Br. J. Dermatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ellerhorst, J.A.; Greene, V.R.; Ekmekcioglu, S.; Warneke, C.L.; Johnson, M.M.; Cooke, C.P.; Wang, L.E.; Prieto, V.G.; Gershenwald, J.E.; Wei, Q.; et al. Clinical correlates of NRAS and BRAF mutations in primary human melanoma. Clin. Cancer Res. 2011, 17, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K.H.; et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Van Raamsdonk, C.D.; Griewank, K.G.; Crosby, M.B.; Garrido, M.C.; Vemula, S.; Wiesner, T.; Obenauf, A.C.; Wackernagel, W.; Green, G.; Bouvier, N.; et al. Mutations in GNA11 in uveal melanoma. N. Engl. J. Med. 2010, 363, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Shain, A.H.; Yeh, I.; Kovalyshyn, I.; Sriharan, A.; Talevich, E.; Gagnon, A.; Dummer, R.; North, J.; Pincus, L.; Ruben, B.; et al. The genetic evolution of melanoma from precursor lesions. N. Engl. J. Med. 2015, 373, 1926–1936. [Google Scholar] [CrossRef] [PubMed]
- Akbani, R.; Akdemir, K.C.; Aksoy, B.A.; Albert, M.; Ally, A.; Amin, S.B.; Arachchi, H.; Arora, A.; Auman, J.T.; Ayala, B.; et al. Genomic classification of cutaneous melanoma. Cell 2015, 161, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.; Watson, I.R. Molecular characterisation of cutaneous melanoma: Creating a framework for targeted and immune therapies. Br. J. Cancer 2016, 115, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.S.; D’Angelo, S.P.; Minor, D.; Hodi, F.S.; Gutzmer, R.; Neyns, B.; Hoeller, C.; Khushalani, N.I.; Miller, W.H.; Lao, C.D.; et al. Nivolumab versus chemotherapy in patients with advanced melanoma who progressed after anti-CTLA-4 treatment (CheckMate 037): A randomised, controlled, open-label, phase 3 trial. Lancet Oncol. 2015, 16, 375–384. [Google Scholar] [CrossRef]
- Robert, C.; Karaszewska, B.; Schachter, J.; Rutkowski, P.; Mackiewicz, A.; Stroiakovski, D.; Lichinitser, M.; Dummer, R.; Grange, F.; Mortier, L.; et al. Improved overall survival in melanoma with combined dabrafenib and trametinib. N. Engl. J. Med. 2014, 372, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Schachter, J.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus ipilimumab in advanced melanoma. N. Engl. J. Med. 2015, 372, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Chapman, P.B.; Hauschild, A.; Robert, C.; Haanen, J.B.; Ascierto, P.; Larkin, J.; Dummer, R.; Garbe, C.; Testori, A.; Maio, M.; et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 2011, 364, 2507–2516. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.B.; Kefford, R.; Pavlick, A.C.; Infante, J.R.; Ribas, A.; Sosman, J.A.; Fecher, L.A.; Millward, M.; McArthur, G.A.; Hwu, P.; et al. Phase II study of the MEK1/MEK2 inhibitor trametinib in patients with metastatic BRAF-mutant cutaneous melanoma previously treated with or without a BRAF inhibitor. J. Clin. Oncol. 2013, 31, 482–489. [Google Scholar] [CrossRef] [PubMed]
- McGettigan, S. Dabrafenib: A new therapy for use in BRAF-mutated metastatic melanoma. J. Adv. Pract. Oncol. 2014, 5, 211–215. [Google Scholar] [PubMed]
- Livingstone, E.; Zimmer, L.; Piel, S.; Schadendorf, D. PLX4032: Does it keep its promise for metastatic melanoma treatment? Expert Opin. Investig. Drugs 2010, 19, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Hauschild, A.; Grob, J.J.; Demidov, L.V.; Jouary, T.; Gutzmer, R.; Millward, M.; Rutkowski, P.; Blank, C.U.; Miller, W.H.; Kaempgen, E.; et al. Dabrafenib in BRAF-mutated metastatic melanoma: A multicentre, open-label, phase 3 randomised controlled trial. Lancet 2012, 380, 358–365. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, K.; Zhu, X.; Lin, G.; Song, F.; Zhao, Y.; Piao, Y.; Liu, J.; Cheng, W.; Bi, X.; et al. Encorafenib (LGX818), a potent BRAF inhibitor, induces senescence accompanied by autophagy in BRAFV600E melanoma cells. Cancer Lett. 2016, 370, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Nazarian, R.; Shi, H.; Wang, Q.; Kong, X.; Koya, R.C.; Lee, H.; Chen, Z.; Lee, M.-K.; Attar, N.; Sazegar, H.; et al. Melanomas acquire resistance to B-RAFV600E inhibition by RTK or N-RAS upregulation. Nature 2010, 468, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, L.; Hillen, U.; Livingstone, E.; Lacouture, M.E.; Busam, K.; Carvajal, R.D.; Egberts, F.; Hauschild, A.; Kashani-Sabet, M.; Goldinger, S.M.; et al. Atypical melanocytic proliferations and new primary melanomas in patients with advanced melanoma undergoing selective BRAF Inhibition. J. Clin. Oncol. 2012, 30, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Hatzivassiliou, G.; Song, K.; Yen, I.; Brandhuber, B.J.; Anderson, D.J.; Alvarado, R.; Ludlam, M.J.; Stokoe, D.; Gloor, S.L.; Vigers, G.; et al. RAF inhibitors prime wild-type RAF to activate the MAPK pathway and enhance growth. Nature 2010, 464, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.T.; Infante, J.R.; Daud, A.; Gonzalez, R.; Kefford, R.F.; Sosman, J.; Hamid, O.; Schuchter, L.; Cebon, J.; Ibrahim, N.; et al. Combined BRAF and MEK inhibition in melanoma with BRAFV600 mutations. N. Engl. J. Med. 2012, 367, 1694–1703. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Marincola, F.M.; Atkins, M.B. What’s new in melanoma? Combination! J. Transl. Med. 2015, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Ribas, A.; Gonzalez, R.; Pavlick, A.; Hamid, O.; Gajewski, T.F.; Daud, A.; Flaherty, L.; Logan, T.; Chmielowski, B.; Lewis, K.; et al. Combination of vemurafenib and cobimetinib in patients with advanced BRAFV600-mutated melanoma: A phase 1B study. Lancet Oncol. 2014, 15, 954–965. [Google Scholar] [CrossRef]
- Larkin, J.; Ascierto, P.A.; Dréno, B.; Atkinson, V.; Liszkay, G.; Maio, M.; Mandalà, M.; Demidov, L.; Stroyakovskiy, D.; Thomas, L.; et al. Combined vemurafenib and cobimetinib in BRAF-mutated melanoma. N. Engl. J. Med. 2014, 371, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Puzanov, I. Treatment of NRAS-mutant melanoma. Curr. Treat. Options Oncol. 2015, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Schadendorf, D.; Berking, C.; Agarwala, S.S.; van Herpen, C.M.L.; Queirolo, P.; Blank, C.U.; Hauschild, A.; Beck, J.T.; St-Pierre, A.; et al. MEK162 for patients with advanced melanoma harbouring NRAS or Val600 BRAF mutations: A non-randomised, open-label phase 2 study. Lancet Oncol. 2013, 14, 249–256. [Google Scholar] [CrossRef]
- Richtig, G.; Richtig, E.; Kashofer, K.; Koch, L.; Winter, G.; Hoefler, G.; Pichler, M.; Ehall, B.; Grübler, M.R.; Heinemann, A.; et al. Testing and clinical implications for non-V600 BRAF mutations in metastatic NRAS(mt) melanoma. Br. J. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Nissan, M.H.; Pratilas, C.A.; Jones, A.M.; Ramirez, R.; Won, H.; Liu, C.; Tiwari, S.; Kong, L.; Hanrahan, A.J.; Yao, Z.; Merghoub, T.; et al. Loss of NF1 in cutaneous melanoma is associated with RAS activation and MEK dependence. Cancer Res. 2014, 74, 2340–2350. [Google Scholar] [CrossRef] [PubMed]
- Ranzani, M.; Alifrangis, C.; Perna, D.; Dutton-Regester, K.; Pritchard, A.; Wong, K.; Rashid, M.; Robles-Espinoza, C.D.; Hayward, N.K.; Mcdermott, U.; et al. BRAF/NRAS wild-type melanoma, NF1 status and sensitivity to trametinib. Pigment Cell Melanoma Res. 2015, 28, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, S.R.; Theurillat, J.-P.; van Allen, E.; Wagle, N.; Hsiao, J.; Cowley, G.S.; Schadendorf, D.; Root, D.E.; Garraway, L.A. A genome-scale RNA interference screen implicates NF1 loss in resistance to RAF inhibition. Cancer Discov. 2013, 3, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Krauthammer, M.; Kong, Y.; Bacchiocchi, A.; Evans, P.; Pornputtapong, N.; Wu, C.; McCusker, J.P.; Ma, S.; Cheng, E.; Straub, R.; et al. Exome sequencing identifies recurrent mutations in NF1 and RASopathy genes in sun-exposed melanomas. Nat. Genet. 2015, 47, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.A.; Busam, K.; Pinkel, D.; Bastian, B.C. Somatic activation of KIT in distinct subtypes of melanoma. J. Clin. Oncol. 2006, 24, 4340–4346. [Google Scholar] [CrossRef] [PubMed]
- Bastian, B.C.; Esteve-Puig, R. Targeting activated KIT signaling for melanoma therapy. J. Clin. Oncol. 2013, 31, 3288–3290. [Google Scholar] [CrossRef] [PubMed]
- Posch, C.; Moslehi, H.; Sanlorenzo, M.; Green, G.; Panzer-Grümayer, R.; Rappersberger, K.; Ortiz-Urda, S. Pharmacological inhibitors of c-KIT block mutant c-KIT mediated migration of melanocytes and melanoma cells in vitro and in vivo. Oncotarget 2016, 7, 45916–45925. [Google Scholar] [CrossRef] [PubMed]
- Hodi, F.S.; Corless, C.L.; Giobbie-Hurder, A.; Fletcher, J.A.; Zhu, M.; Marino-Enriquez, A.; Friedlander, P.; Gonzalez, R.; Weber, J.S.; Gajewski, T.F.; et al. Imatinib for melanomas harboring mutationally activated or amplified kit arising on mucosal, acral, and chronically sun-damaged skin. J. Clin. Oncol. 2013, 31, 3182–3190. [Google Scholar] [CrossRef] [PubMed]
- Minor, D.R.; Kashani-Sabet, M.; Garrido, M.; O’Day, S.J.; Hamid, O.; Bastian, B.C. Sunitinib therapy for melanoma patients with KIT mutations. Clin. Cancer Res. 2012, 18, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, R.D.; Lawrence, D.P.; Weber, J.S.; Gajewski, T.F.; Gonzalez, R.; Lutzky, J.; O’Day, S.J.; Hamid, O.; Wolchok, J.D.; Chapman, P.B.; et al. Phase II study of nilotinib in melanoma harboring KIT alterations following progression to prior KIT inhibition. Clin. Cancer Res. 2015, 21, 2289–2296. [Google Scholar] [CrossRef] [PubMed]
- Atkins, M.B.; Kunkel, L.; Sznol, M.; Rosenberg, S.A. High-dose recombinant interleukin-2 therapy in patients with metastatic melanoma: Long-term survival update. Cancer J. Sci. Am. 2000, 6, 11–14. [Google Scholar]
- Robert, C.; Thomas, L.; Bondarenko, I.; O’Day, S.; Weber, J.; Garbe, C.; Lebbe, C.; Baurain, J.-F.; Testori, A.; Grob, J.-J.; et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N. Engl. J. Med. 2011, 364, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Schadendorf, D.; Hodi, F.S.; Robert, C.; Weber, J.S.; Margolin, K.; Hamid, O.; Patt, D.; Chen, T.-T.; Berman, D.M.; Wolchok, J.D. Pooled analysis of long-term survival data from phase II and phase III trials of ipilimumab in unresectable or metastatic melanoma. J. Clin. Oncol. 2015, 33, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.; Kuball, J.; Voelkl, S.; Wiendl, H.; Becker, B.; Walter, B.; Majdic, O.; Gajewski, T.F.; Theobald, M.; Andreesen, R.; et al. Blockade of PD-L1 (B7-H1) augments human tumor-specific T cell responses in vitro. Int. J. Cancer 2006, 119, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Carvajal, R.D.; Sosman, J.A.; Atkins, M.B.; et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N. Engl. J. Med. 2012, 366, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Hodi, F.S.; Kluger, H.; Sznol, M.; Carvajal, R.; Lawrence, D.; Atkins, M.; Powderly, J.; Sharfman, W.; Puzanov, I.; Smith, D.; et al. Durable, Long-Term Survival in Previously Treated Patients with Advanced Melanoma (MEL) Who Received Nivolumab (NIVO) Monotherapy in a Phase I Trial. Available online: http://www.abstractsonline.com/Plan/ViewAbstract.aspx?mID=4017&sKey=371fa616-a0cf-4bf-993d-ce424853b52c&cKey=616f965e-a236-4bd2-9f7a-6399bd6f3f6c&mKey=1d10d749-4b6a-4ab3-bcd4-f80fb1922267 (accessed on 22 January 2017).
- Galluzzi, L.; Eggermont, A.; Kroemer, G. Doubling the blockade for melanoma immunotherapy. Oncoimmunology 2016, 5, e1106127. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Kluger, H.; Callahan, M.K.; Postow, M.A.; Rizvi, N.A.; Lesokhin, A.M.; Segal, N.H.; Ariyan, C.E.; Gordon, R.-A.; Reed, K.; et al. Nivolumab plus ipilimumab in advanced melanoma. N. Engl. J. Med. 2013, 369, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Romero, D. Immunotherapy: PD-1 says goodbye, TIM-3 says hello. Nat. Rev. Clin. Oncol. 2016, 13, 202–203. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Sinha, D.; Barkauskas, D.; Young, A.; Kalimutho, M.; Stannard, K.; Caramia, F.; Haibe-Kains, B.; Stagg, J.; Khanna, K.K.; et al. Adenosine 2B receptor expression on cancer cells promotes metastasis. Cancer Res. 2016, 76, 4372–4382. [Google Scholar] [CrossRef] [PubMed]
- Knowling, S.; Morris, K.V. Non-coding RNA and antisense RNA. Nature’s trash or treasure? Biochimie 2011, 93, 1922–1927. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project; Bernstein, B.E.; Birney, E.; Dunham, I.; Green, E.D.; Gunter, C.; Snyder, M. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]
- Smolle, M.A.; Calin, H.N.; Pichler, M.; Calin, G.A. Noncoding RNAs and immune checkpoints: Clinical implications as cancer therapeutics. FEBS J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pichler, M.; Stiegelbauer, V.; Vychytilova-Faltejskova, P.; Ivan, C.; Ling, H.; Winter, E.; Zhang, X.; Goblirsch, M.; Wulf-Goldenberg, A.; Ohtsuka, M.; et al. Genome-wide miRNA analysis identifies miR-188-3p as a novel prognostic marker and molecular factor involved in colorectal carcinogenesis. Clin. Cancer Res. 2016, 23, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Pichler, M.; Calin, G.A. MicroRNAs in cancer: From developmental genes in worms to their clinical application in patients. Br. J. Cancer 2015, 113, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015. [Google Scholar] [CrossRef] [PubMed]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Mattick, J.S. Differentiating protein-coding and noncoding RNA: Challenges and ambiguities. PLoS Comput. Biol. 2008, 4, e1000176. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Carmichael, G.G. Long noncoding RNAs in mammalian cells: What, where, and why? Wiley Interdiscip. Rev. RNA 2010, 1, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Lipovich, L.; Johnson, R.; Lin, C.Y. MacroRNA underdogs in a microRNA world: Evolutionary, regulatory, and biomedical significance of mammalian long non-protein-coding RNA. Biochim. Biophys. Acta Gene Regul. Mech. 2010, 1799, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Cerk, S.; Schwarzenbacher, D.; Adiprasito, J.B.; Stotz, M.; Hutterer, G.C.; Gerger, A.; Ling, H.; Calin, G.A.; Pichler, M. Current status of long non-coding RNAs in human breast cancer. Int. J. Mol. Sci. 2016, 17, 1485. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, M.; Ling, H.; Ivan, C.; Pichler, M.; Matsushita, D.; Goblirsch, M.; Stiegelbauer, V.; Shigeyasu, K.; Zhang, X.; Chen, M.; et al. H19 noncoding RNA, an independent prognostic factor, regulates essential Rb-E2F and CDK8-β-catenin signaling in colorectal cancer. EBioMedicine 2016, 13, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Seles, M.; Hutterer, G.; Kiesslich, T.; Pummer, K.; Berindan-Neagoe, I.; Perakis, S.; Schwarzenbacher, D.; Stotz, M.; Gerger, A.; Pichler, M. Current insights into long non-coding RNAs in renal cell carcinoma. Int. J. Mol. Sci. 2016, 17, 573. [Google Scholar] [CrossRef] [PubMed]
- Smolle, M.A.; Bullock, M.D.; Ling, H.; Pichler, M.; Haybaeck, J. Long non-coding RNAs in endometrial carcinoma. Int. J. Mol. Sci. 2015, 16, 26463–26472. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.F. Non-coding RNAs: Meet thy masters. BioEssays 2010, 32, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010, 142, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Guenther, M.G.; Levine, S.S.; Boyer, L.A.; Jaenisch, R.; Young, R.A. A chromatin landmark and transcription initiation at most promoters in human cells. Cell 2007, 130, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Wu, H.; Ni, P.; Gu, Z.; Qiao, Y.; Chen, N.; Sun, F.; Fan, Q. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucl. Acids Res. 2010, 38, 5366–5383. [Google Scholar] [CrossRef] [PubMed]
- Martianov, I.; Ramadass, A.; Serra Barros, A.; Chow, N.; Akoulitchev, A. Repression of the human dihydrofolate reductase gene by a non-coding interfering transcript. Nature 2007, 445, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Hämmerle, M.; Eißmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Groß, M.; et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Shen, Z.; Chakraborty, A.; Giri, S.; Freier, S.M.; Wu, X.; Zhang, Y.; Gorospe, M.; Prasanth, S.G.; Lal, A.; et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB. PLoS Genet. 2013, 9, e1003368. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.; Chang, H.Y. Long noncoding RNA in genome regulation: Prospects and mechanisms. RNA Biol. 2011, 7, 582–585. [Google Scholar] [CrossRef]
- Good, M.C.; Zalatan, J.G.; Lim, W.A. Scaffold proteins: Hubs for controlling the flow of cellular information. Science 2011, 332, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, X.; Zhou, J.; Hu, J.; Zhang, D.; Liu, J.; Qiao, Y.; Zhan, Q. LncRNA HULC modulates the phosphorylation of YB-1 through serving as a scaffold of ERK and YB-1 to enhance hepatocarcinogenesis. Hepatology 2016. [Google Scholar] [CrossRef]
- Sun, C.-C.; Li, S.-J.; Li, G.; Hua, R.-X.; Zhou, X.-H.; Li, D.-J. Long intergenic noncoding RNA 00511 acts as an oncogene in non-small-cell lung cancer by binding to EZH2 and suppressing p57. Mol. Ther. Acids 2016, 5, e385. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15 (INK4B) tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xiao, Z.-D.; Han, L.; Zhang, J.; Lee, S.-W.; Wang, W.; Lee, H.; Zhuang, L.; Chen, J.; Lin, H.-K.; et al. LncRNA NBR2 engages a metabolic checkpoint by regulating AMPK under energy stress. Nat. Cell Biol. 2016, 18, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xiao, Z.D.; Gan, B. An lncRNA switch for AMPK activation. Cell Cycle 2016, 15, 1948–1949. [Google Scholar] [CrossRef] [PubMed]
- Pasmant, E.; Laurendeau, I.; Héron, D.; Vidaud, M.; Vidaud, D.; Bièche, I. Characterization of a germ-line deletion, including the entire INK4/ARF locus, in a melanoma-neural system tumor family: Identification of ANRIL, an antisense noncoding RNA whose expression coclusters with ARF. Cancer Res. 2007, 67, 3963–3969. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.T.; Demenais, F.; Iles, M.M.; Harland, M.; Taylor, J.C.; Corda, E.; Randerson-Moor, J.; Aitken, J.F.; Avril, M.F.; Azizi, E.; et al. Genome-wide association study identifies three loci associated with melanoma risk. Nat. Genet. 2009, 41, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, R.; Smith, E.; Graham, G. Multiple neurofibromas as the presenting feature of familial atypical multiple malignant melanoma (FAMMM) syndrome. Am. J. Med. Genet. Part A 2013, 161, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Cunnington, M.S.; Koref, M.S.; Mayosi, B.M.; Burn, J.; Keavney, B. Chromosome 9p21 SNPs associated with multiple disease phenotypes correlate with ANRIL expression. PLoS Genet. 2010, 6, e1000899. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, H.; Pan, H.; Shi, Y.; Li, T.; Ge, S.; Jia, R.; Zhang, H.; Fan, X. ANRIL lncRNA triggers efficient therapeutic efficacy by reprogramming the aberrant INK4-hub in melanoma. Cancer Lett. 2016, 381, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Flockhart, R.J.; Webster, D.E.; Qu, K.; Mascarenhas, N.; Kovalski, J.; Kretz, M.; Khavari, P.A. BRAFV600E remodels the melanocyte transcriptome and induces BANCR to regulate melanoma cell migration. Genome Res. 2012, 22, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Jia, L.; Duan, Y.; Li, Y.; Bao, L.; Sha, N. Long non-coding RNA BANCR promotes proliferation in malignant melanoma by regulating MAPK pathway activation. PLoS ONE 2014, 9, e100893. [Google Scholar] [CrossRef] [PubMed]
- Lessard, L.; Liu, M.; Marzese, D.M.; Wang, H.; Chong, K.; Kawas, N.; Donovan, N.C.; Kiyohara, E.; Hsu, S.; Nelson, N.; et al. The CASC15 long intergenic noncoding RNA locus is involved in melanoma progression and phenotype switching. J. Investig. Dermatol. 2015, 135, 2464–2474. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Steitz, J.A. Classificatiocelln of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5′-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell. Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, H.; Xiao, Y.; Tang, X.; Li, Y.; Han, Q.; Fu, J.; Yang, Y.; Zhu, Y. Lentiviral-mediated overexpression of long non-coding RNA GAS5 reduces invasion by mediating MMP2 expression and activity in human melanoma cells. Int. J. Oncol. 2016, 48, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zhang, W.; Su, B.; Yu, B. Long noncoding RNA HOTAIR is associated with motility, invasion, and metastatic potential of metastatic melanoma. BioMed Res. Int. 2013, 2013, 251098. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhou, L.; Wu, L.-M.; Lai, M.-C.; Xie, H.-Y.; Zhang, F.; Zheng, S.-S. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol. 2011, 18, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Murat, P.; Matak-Vinkovic, D.; Murrell, A.; Balasubramanian, S. Binding interactions between long noncoding RNA HOTAIR and PRC2 proteins. Biochemistry 2013, 52, 9519–9527. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Sui, A.; Garen, A. Binding of mouse VL30 retrotransposon RNA to PSF protein induces genes repressed by PSF: Effects on steroidogenesis and oncogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Tan, G.H.; Ma, C.C.; Li, L. The non-coding RNA llme23 drives the malignant property of human melanoma cells. J. Genet. Genom. 2013, 40, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin β4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Hämmerle, M.; Diederichs, S. MALAT1—A paradigm for long noncoding RNA function in cancer. J. Mol. Med. 2013, 91, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, X.; Hao, Y.; Fang, Z.; He, Y. Potential roles of abnormally expressed long noncoding RNA UCA1 and Malat-1 in metastasis of melanoma. Melanoma Res. 2014, 24, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Sirokman, K.; McDonel, P.; Shishkin, A.A.; Surka, C.; Russell, P.; Grossman, S.R.; Chow, A.Y.; Guttman, M.; Lander, E.S. RNA-RNA interactions enable specific targeting of noncoding RNAs to nascent pre-mRNAs and chromatin sites. Cell 2014, 159, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Arancio, W.; Pizzolanti, G.; Genovese, S.I.; Baiamonte, C.; Giordano, C. Competing endogenous RNA and interactome bioinformatic analyses on human telomerase. Rejuvenation Res. 2014, 17, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.; Li, L.; Shi, Y.; Bu, X.; Xia, Y.; Wang, J.; Djangmah, H.S.; Liu, X.; You, Y.; Xu, B.; et al. Long non-coding RNA MALAT1 acts as a competing endogenous RNA to promote malignant melanoma growth and metastasis by sponging miR-22. Oncotarget 2016, 7, 63901–63912. [Google Scholar] [CrossRef] [PubMed]
- Silayoi, P.; Speece, M. The importance of packaging attributes: A conjoint analysis approach. Eur. J. Mark 2007, 41, 1495–1517. [Google Scholar] [CrossRef]
- Ding, X.; Wang, X.; Lin, M.; Xing, Y.; Ge, S.; Jia, R.; Zhang, H.; Fan, X.; Li, J. PAUPAR lncRNA suppresses tumourigenesis by H3K4 demethylation in uveal melanoma. FEBS Lett. 2016, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.F.; Torrieri, R.; Silva, R.R.; Pereira, C.G.; Valente, V.; Torrieri, E.; Peronni, K.C.; Martins, W.; Muto, N.; Francisco, G.; et al. Novel primate-specific genes, RMEL 1, 2 and 3, with highly restricted expression in melanoma, assessed by new data mining tool. PLoS ONE 2010, 5, e13510. [Google Scholar] [CrossRef] [PubMed]
- Goedert, L.; Pereira, C.G.; Roszik, J.; Plaça, J.R.; Cardoso, C.; Chen, G.; Deng, W.; Yennu-Nanda, V.G.; Silva, W.A., Jr.; Davies, M.A.; et al. RMEL3, a novel BRAFV600E-associated long noncoding RNA, is required for MAPK and PI3K signaling in melanoma. Oncotarget 2016, 7, 36711–36718. [Google Scholar] [PubMed]
- Leucci, E.; Vendramin, R.; Spinazzi, M.; Laurette, P.; Fiers, M.; Wouters, J.; Radaelli, E.; Eyckerman, S.; Leonelli, C.; Vanderheyden, K.; et al. Melanoma addiction to the long non-coding RNA SAMMSON. Nature 2016, 531, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Satoh, H.; Moriyama, M.; Satoh, K.; Morishita, Y.; Yoshida, S.; Watanabe, T.; Nakamura, Y.; Mori, S. Intronic U50 small-nucleolar-RNA (snoRNA) host gene of no protein-coding potential is mapped at the chromosome breakpoint t(3;6)(q27;q15) of human B-cell lymphoma. Genes Cells 2000, 5, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Ichigozaki, Y.; Fukushima, S.; Jinnin, M.; Miyashita, A.; Nakahara, S.; Tokuzumi, A.; Yamashita, J.; Kajihara, I.; Aoi, J.; Masuguchi, S.; et al. Serum long non-coding RNA, snoRNA host gene 5 level as a new tumor marker of malignant melanoma. Exp. Dermatol. 2016, 25, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Mazar, J.; Zhao, W.; Khalil, A.M.; Lee, B.; Shelley, J.; Govindarajan, S.S.; Yamamoto, F.; Ratnam, M.; Aftab, M.N.; Collins, S.; et al. The functional characterization of long noncoding RNA SPRY4-IT1 in human melanoma cells. Oncotarget 2014, 5, 8959–8969. [Google Scholar] [CrossRef] [PubMed]
- Khaitan, D.; Dinger, M.E.; Mazar, J.; Crawford, J.; Smith, M.A.; Mattick, J.S.; Perera, R.J. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res. 2011, 71, 3852–3862. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Mazar, J.; Lee, B.; Sawada, J.; Li, J.L.; Shelley, J.; Govindarajan, S.; Towler, D.; Mattick, J.S.; Komatsu, M.; et al. The long noncoding RNA SPRIGHTLY regulates cell proliferation in primary human melanocytes. J. Investig. Dermatol. 2016, 136, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Shen, S.-K.; Xiong, J.-G.; Xu, Y.; Zhang, H.-Q.; Liu, H.-J.; Lu, Z.-G. Clinical significance of long noncoding RNA SPRY4-IT1 in melanoma patients. FEBS Open Bio 2016, 6, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, W.; Yang, C.; Wu, W.; Wu, S.; Qin, X.; Li, X. Long non-coding RNA UCA1a(CUDR) promotes proliferation and tumorigenesis of bladder cancer. Int. J. Oncol. 2012, 41, 276–284. [Google Scholar] [PubMed]
- Wei, Y.; Sun, Q.; Zhao, L.; Wu, J.; Chen, X.; Wang, Y.; Zang, W.; Zhao, G. LncRNA UCA1-miR-507-FOXM1 axis is involved in cell proliferation, invasion and G0/G1 cell cycle arrest in melanoma. Med. Oncol. 2016, 33, 88. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.-D.; Zhuang, L.; Gan, B. Long non-coding RNAs in cancer metabolism. BioEssays 2016, 38, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, S. The four dimensions of noncoding RNA conservation. Trends Genet. 2014, 30, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, M.; Gutschner, T. Long non-coding RNAs in cancer and development: Where do we go from here? Int. J. Mol. Sci. 2015, 16, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
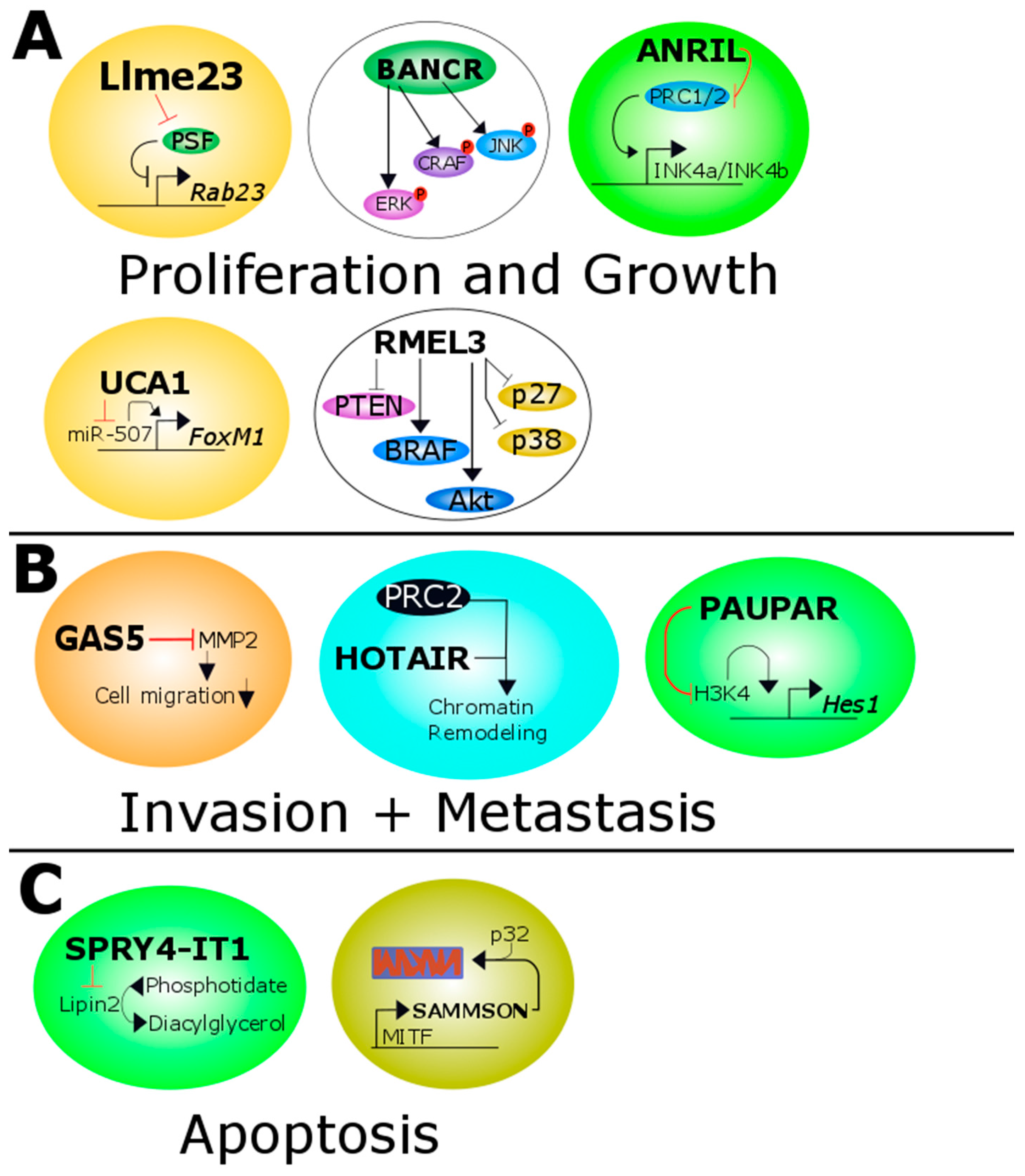
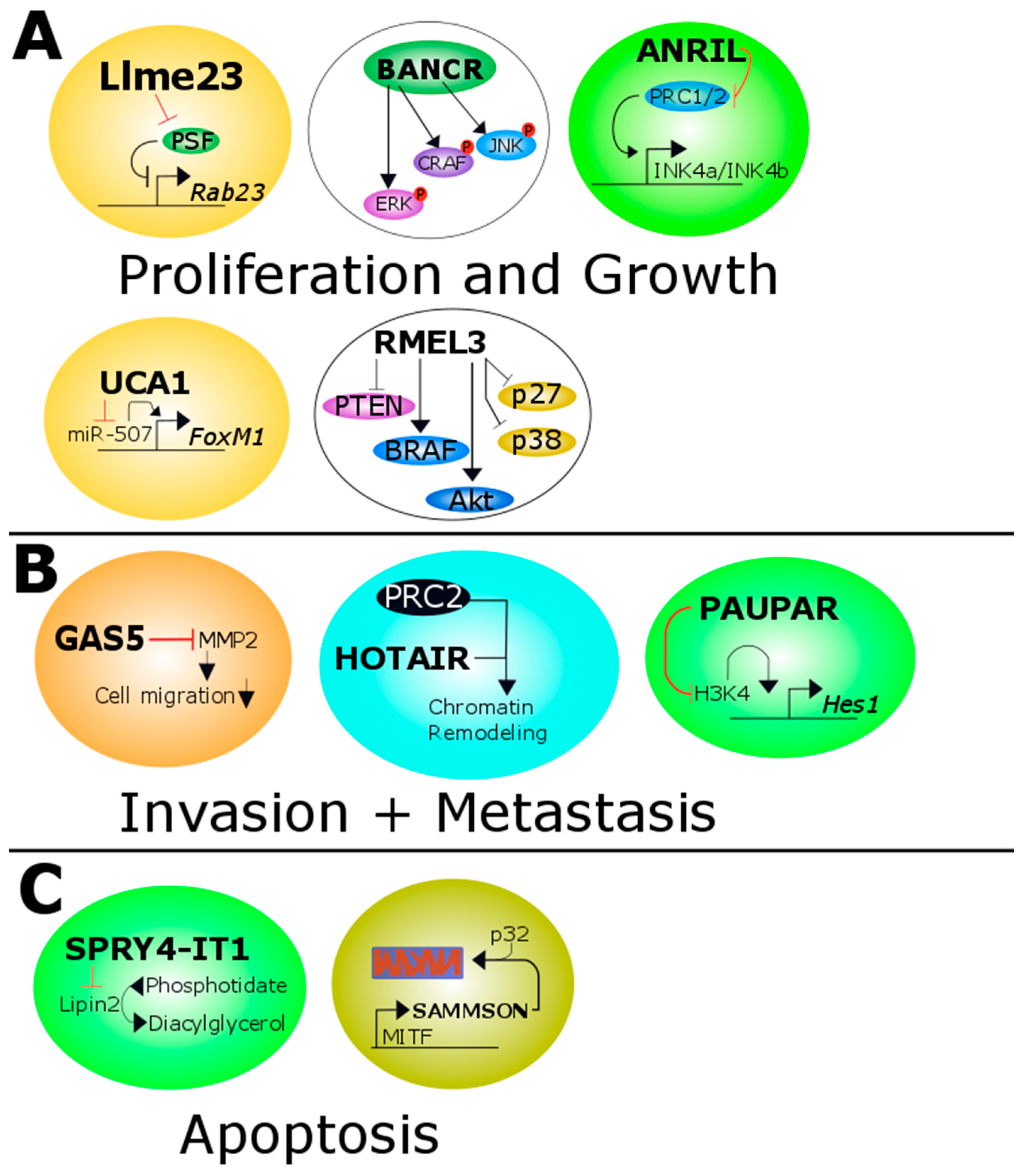
Figure 1.
Overview of all melanoma related lncRNAs. (A) Llme23 binds the protein associated splicing factor which in turn can not bind to the promoter of the proto-oncogene RAB23. BANCR increases the activity of ERK, CRAF and JNK. ANRIL inhibits the transcription of p14/p15/p16 expression by methylation of the histone H3 of the INK4b/ARF/INK4a tumor suppressor locus. UCA1 targets and inactivates miR-507 and this leads to increased levels of the pro-oncogenic transcription factor FoxM1. RMel3 decreases the level of PTEN, p27 and p38 and increases the activity of BRAF and Akt; (B) GAS5 inhibits the matrixmetalloprotease 2 which in turn decreases the ability of migration of melanoma cells. HOTAIR leads together with PRC2 to increased chromatin remodeling. MALAT1 binds to miR-22 in cutaneous melanoma promoting MMP14 and SNAIL expression. In uveal melanoma it binds to miR-140 decreasing SLUG and ADAM10 expression. PAUPAR represses HES1 expression by inhibiting histone H3K4 demethylation; (C) SPRY4-IT1 inhibits Lipin2 which converts phosphatidate to diacylglycerol. SAMMSON is co-amplified with MITF and forms a complex with p32 which stabilized mitochondrial biogenesis. Red and black T shaped bars signify reduced or downregulated proteins/miRNAs or genes. Black arrows indicate overexpressed or upregulated proteins or genes. ERK: extracellular signal-regulated kinase; CRAF: C-Raf proto-oncogene, serine/threonine kinase; JNK: JUN N-terminal kinase; ANRIL: antisense lncRNA in INK4 locus; UCA1: Urothelial carcinoma-associated 1; PTEN: phosphatase and tensin homolog; BRAF: B-Raf proto-oncogene, serine/threonine kinase; GAS5: growth arrest-specific transcript 5; HOTAIR: HOX transcript antisense RNA; PRC2: polycomb repressive complexe 2; MMP14: matrix metalloproteinase 14; SNAIL: snail family transcriptional repressor; SLUG: snail family transcriptional repressor 2; ADAM10: ADAM metallopeptidase domain 10; PAUPAR: PAX6 upstream antisense RNA; SPRY4-IT1: SPRY4 intronic transcript 1; SAMMSON: Survival associated mitochondrial melanoma-specific oncogenic non-coding RNA; MITF: melanogenesis associated transcription factor.
Figure 1.
Overview of all melanoma related lncRNAs. (A) Llme23 binds the protein associated splicing factor which in turn can not bind to the promoter of the proto-oncogene RAB23. BANCR increases the activity of ERK, CRAF and JNK. ANRIL inhibits the transcription of p14/p15/p16 expression by methylation of the histone H3 of the INK4b/ARF/INK4a tumor suppressor locus. UCA1 targets and inactivates miR-507 and this leads to increased levels of the pro-oncogenic transcription factor FoxM1. RMel3 decreases the level of PTEN, p27 and p38 and increases the activity of BRAF and Akt; (B) GAS5 inhibits the matrixmetalloprotease 2 which in turn decreases the ability of migration of melanoma cells. HOTAIR leads together with PRC2 to increased chromatin remodeling. MALAT1 binds to miR-22 in cutaneous melanoma promoting MMP14 and SNAIL expression. In uveal melanoma it binds to miR-140 decreasing SLUG and ADAM10 expression. PAUPAR represses HES1 expression by inhibiting histone H3K4 demethylation; (C) SPRY4-IT1 inhibits Lipin2 which converts phosphatidate to diacylglycerol. SAMMSON is co-amplified with MITF and forms a complex with p32 which stabilized mitochondrial biogenesis. Red and black T shaped bars signify reduced or downregulated proteins/miRNAs or genes. Black arrows indicate overexpressed or upregulated proteins or genes. ERK: extracellular signal-regulated kinase; CRAF: C-Raf proto-oncogene, serine/threonine kinase; JNK: JUN N-terminal kinase; ANRIL: antisense lncRNA in INK4 locus; UCA1: Urothelial carcinoma-associated 1; PTEN: phosphatase and tensin homolog; BRAF: B-Raf proto-oncogene, serine/threonine kinase; GAS5: growth arrest-specific transcript 5; HOTAIR: HOX transcript antisense RNA; PRC2: polycomb repressive complexe 2; MMP14: matrix metalloproteinase 14; SNAIL: snail family transcriptional repressor; SLUG: snail family transcriptional repressor 2; ADAM10: ADAM metallopeptidase domain 10; PAUPAR: PAX6 upstream antisense RNA; SPRY4-IT1: SPRY4 intronic transcript 1; SAMMSON: Survival associated mitochondrial melanoma-specific oncogenic non-coding RNA; MITF: melanogenesis associated transcription factor.

{kind=link}
Table 1.
Overview of lncRNA function(s) in melanoma.
| LncRNA Name | Function | References |
|---|---|---|
| ANRIL | Represses the transcription of CDKN2A/B which leads to perturbation in the cell cycle, increased migration and colony formation. | [94,95,96] |
| BANCR | High levels of BANCR lead to increased migration (by targeting CXCL11) and proliferation. High levels of BANCR directly correlated with tumor stage and indirectly with survival. | [97,98] |
| CASC15 | Promotes melanoma progression and invasiveness. Direct correlation between tumor stage and expression levels. | [99] |
| GAS5 | Indirectly correlates with melanoma migration and invasiveness over reduced levels of MMP2. | [101] |
| HOTAIR | HOTAIR is up-regulated in metastases compared to the primary tumor, favoring a pro-metastatic role. | [102] |
| Llme23 | Llme23 promotes the expression of the proto-oncogenic RAS-related small GTPase Rab23. | [106] |
| MALAT1 | Possibly involved in cell proliferation and invasion. It does this by targeting miR-22 in cutaneous melanoma and miR-140 in uveal melanoma. | [109,112,113] |
| PAUPAR | It is a tumor suppressor lncRNA and reduces cell migration and metastases. | [114] |
| RMEL3 | Depletion led to decreased cell survival and proliferation in BRAFV600E melanoma cell lines. | [116] |
| SAMMSON | Promotes cell viability and growth irrespective of melanomas mutational status. | [117] |
| SNGH5 | Increased serum levels in patients with melanoma. | [119] |
| SPRTY4-IT1 (SPRIGHTLY) | Associated with melanoma-genesis; Associated with higher tumor stage and worse prognosis. | [122,123] |
| UCA1 | Promotes invasion and cell proliferation. | [109,125] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Richtig, G.; Ehall, B.; Richtig, E.; Aigelsreiter, A.; Gutschner, T.; Pichler, M. Function and Clinical Implications of Long Non-Coding RNAs in Melanoma. Int. J. Mol. Sci. 2017, 18, 715. https://doi.org/10.3390/ijms18040715
AMA Style
Richtig G, Ehall B, Richtig E, Aigelsreiter A, Gutschner T, Pichler M. Function and Clinical Implications of Long Non-Coding RNAs in Melanoma. International Journal of Molecular Sciences. 2017; 18(4):715. https://doi.org/10.3390/ijms18040715
Chicago/Turabian StyleRichtig, Georg, Barbara Ehall, Erika Richtig, Ariane Aigelsreiter, Tony Gutschner, and Martin Pichler. 2017. "Function and Clinical Implications of Long Non-Coding RNAs in Melanoma" International Journal of Molecular Sciences 18, no. 4: 715. https://doi.org/10.3390/ijms18040715
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.