
Wedelolactone Acts as Proteasome Inhibitor in Breast Cancer Cells

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
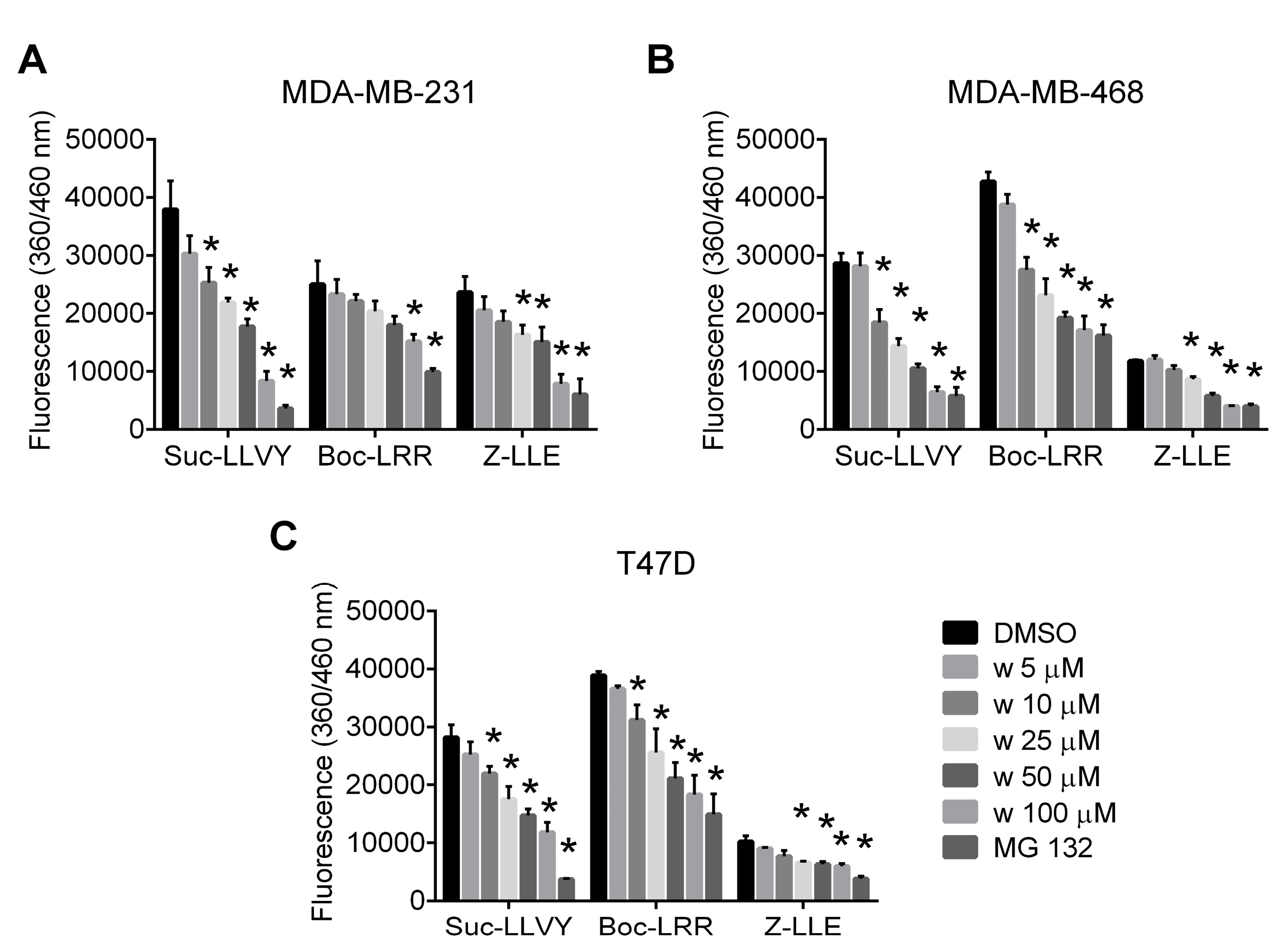
2.1. Wedelolactone Inhibits Proteolytic Activities of Proteasome in Breast Cancer Cell Lines
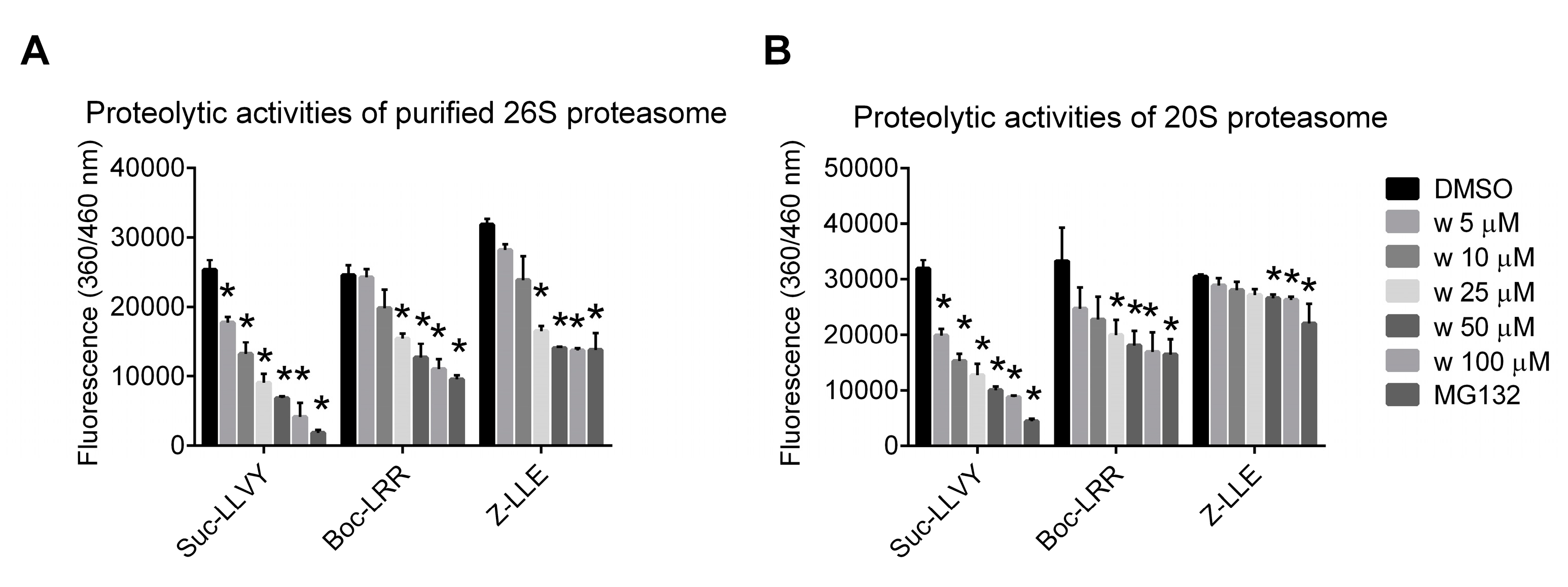
2.2. Wedelolactone Inhibits Proteolytic Activities of Purified 20S and 26S Proteasome Complexes In Vitro
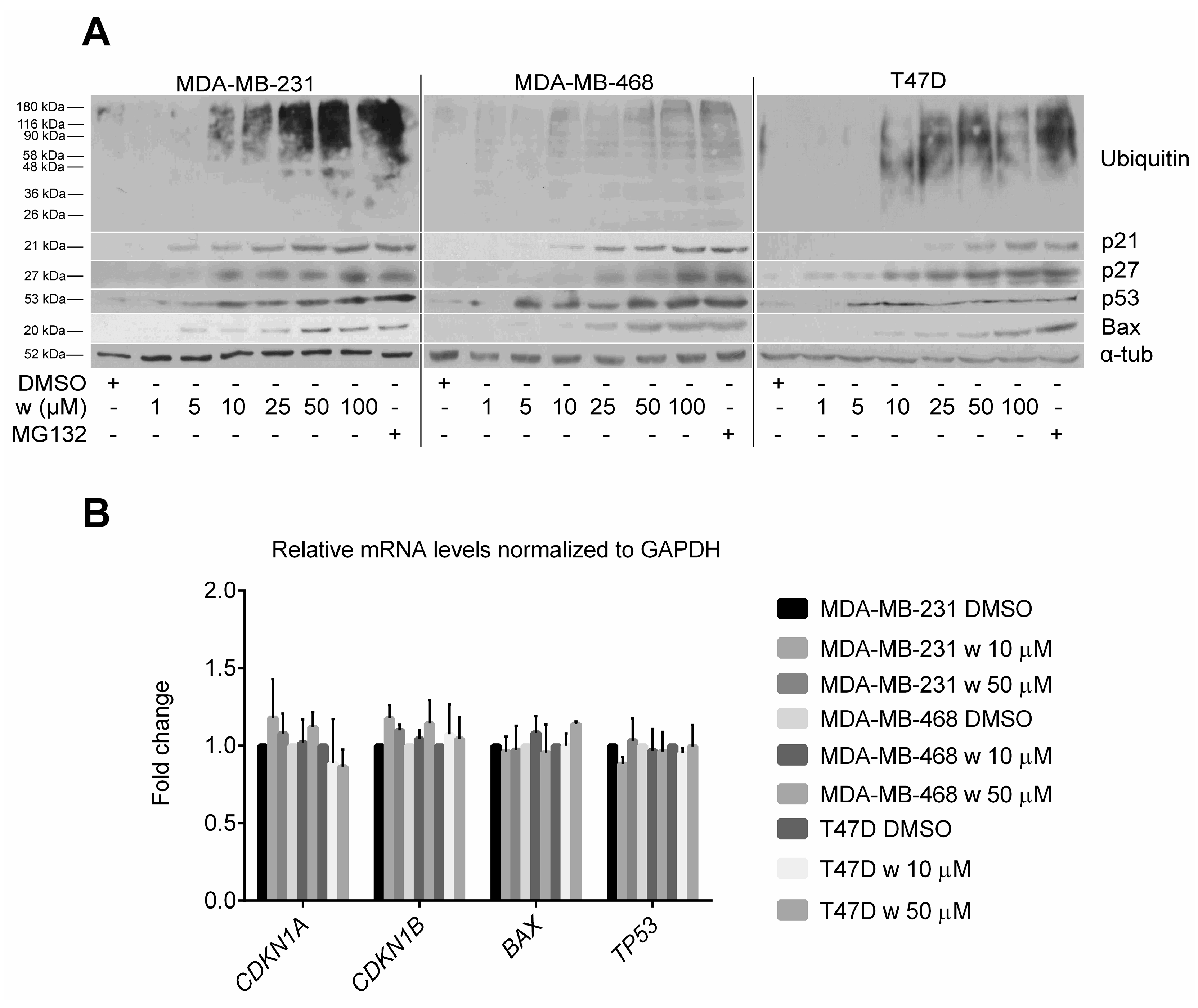
2.3. Wedelolactone Causes Accumulation of Polyubiquitinated and Short-Lived Proteins in Breast Cancer Cells
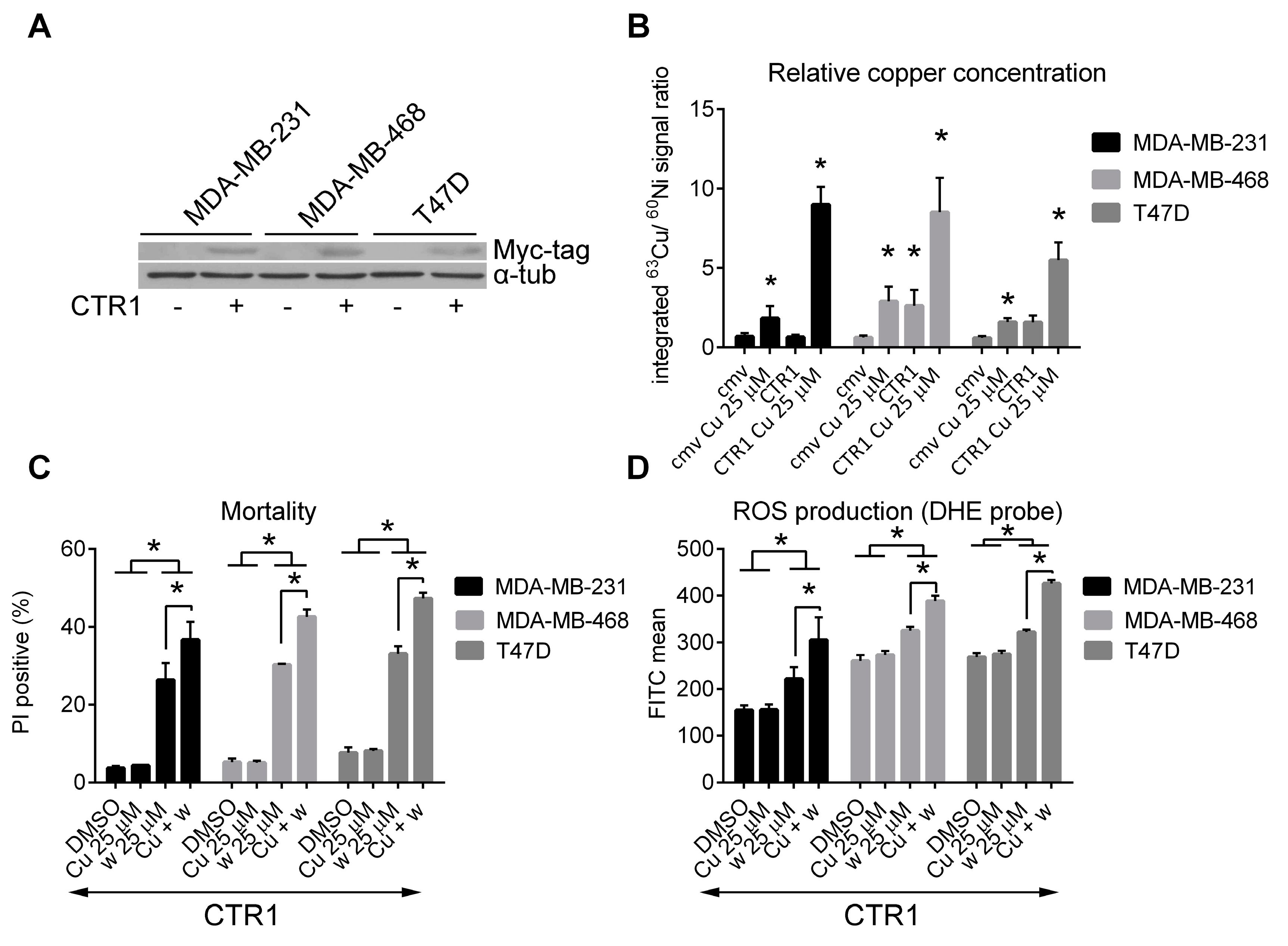
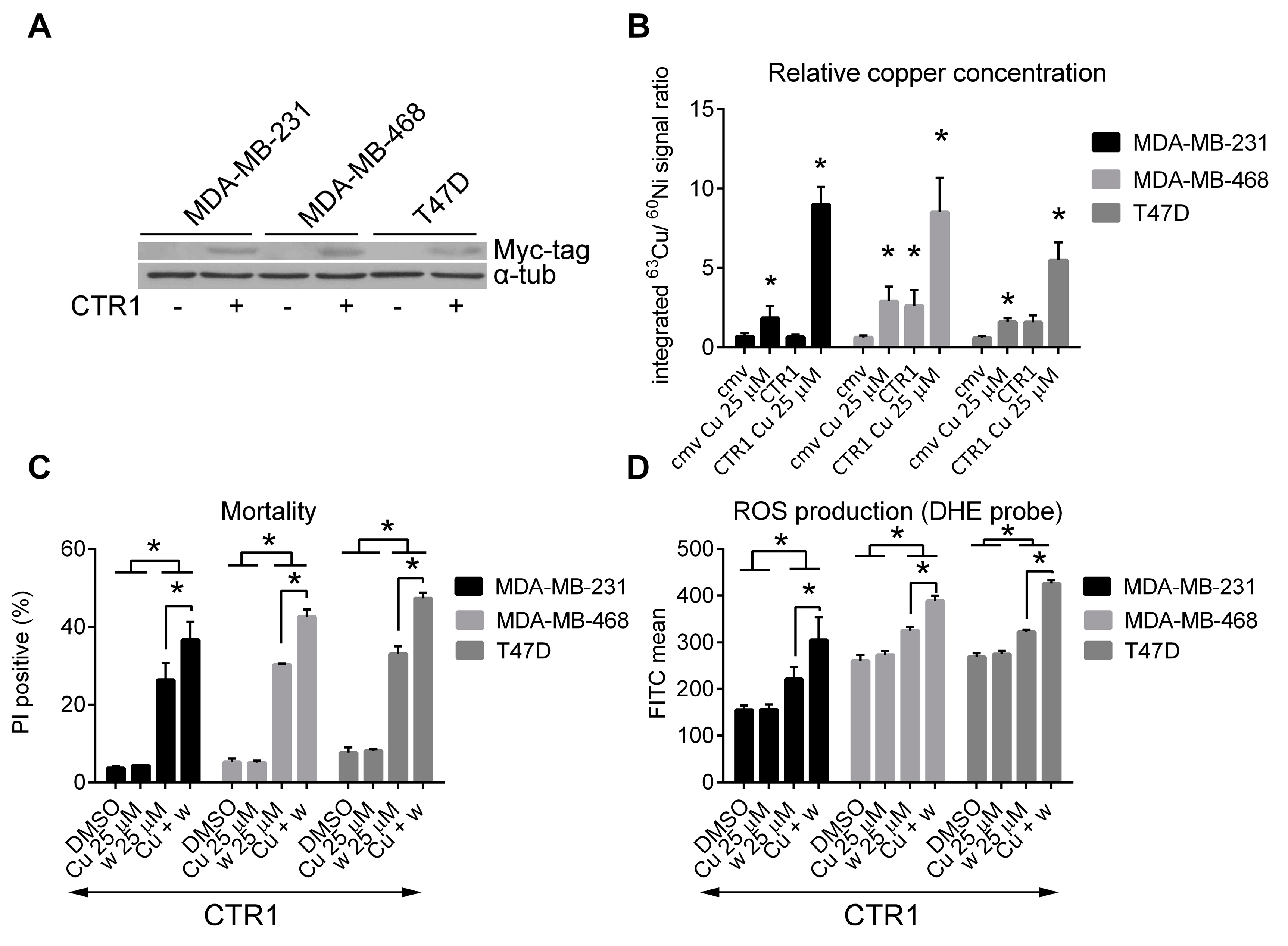
2.4. Cytotoxicity of Wedelolactone Increases for Cells with High Content of Intracellular Copper
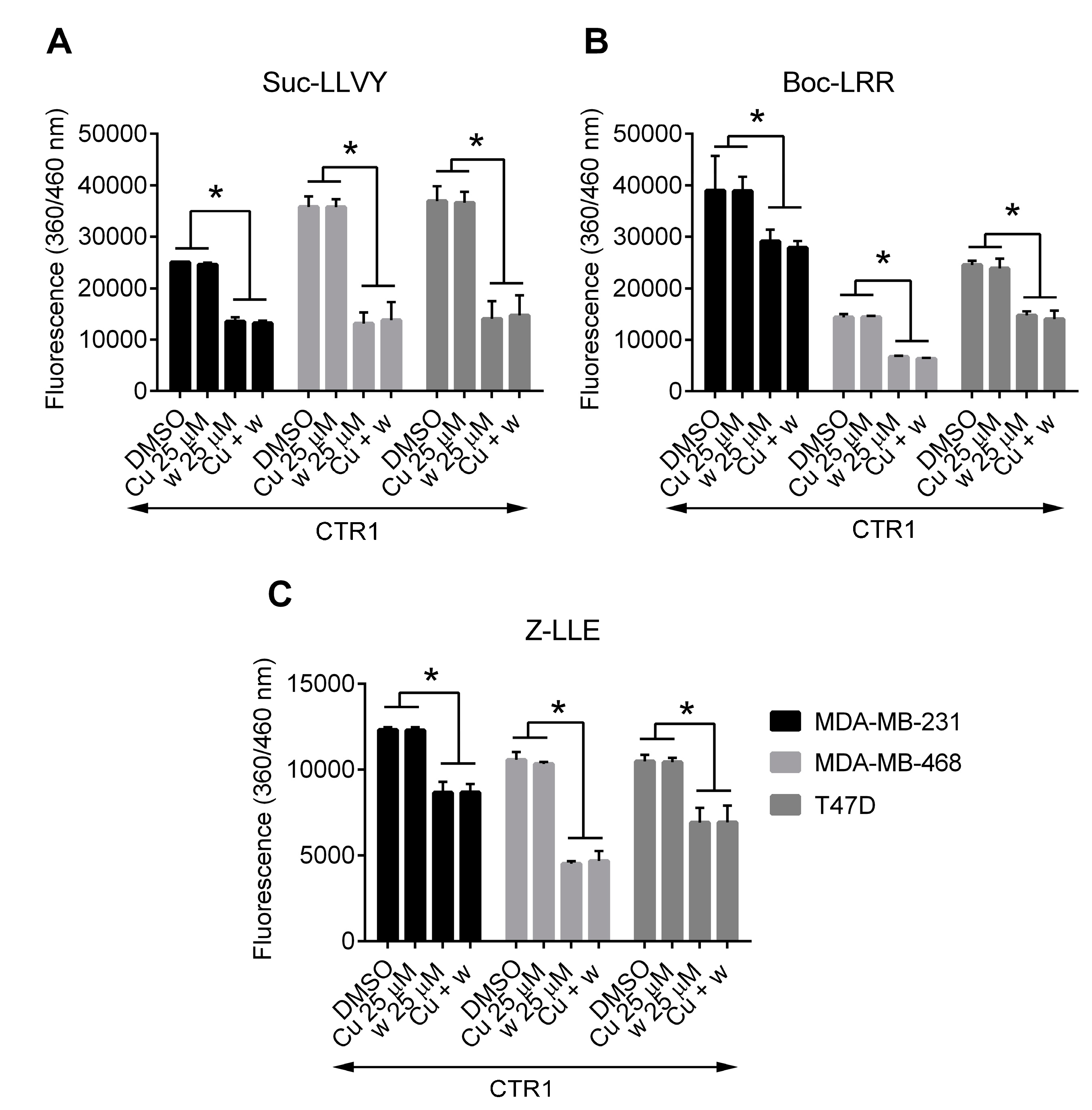
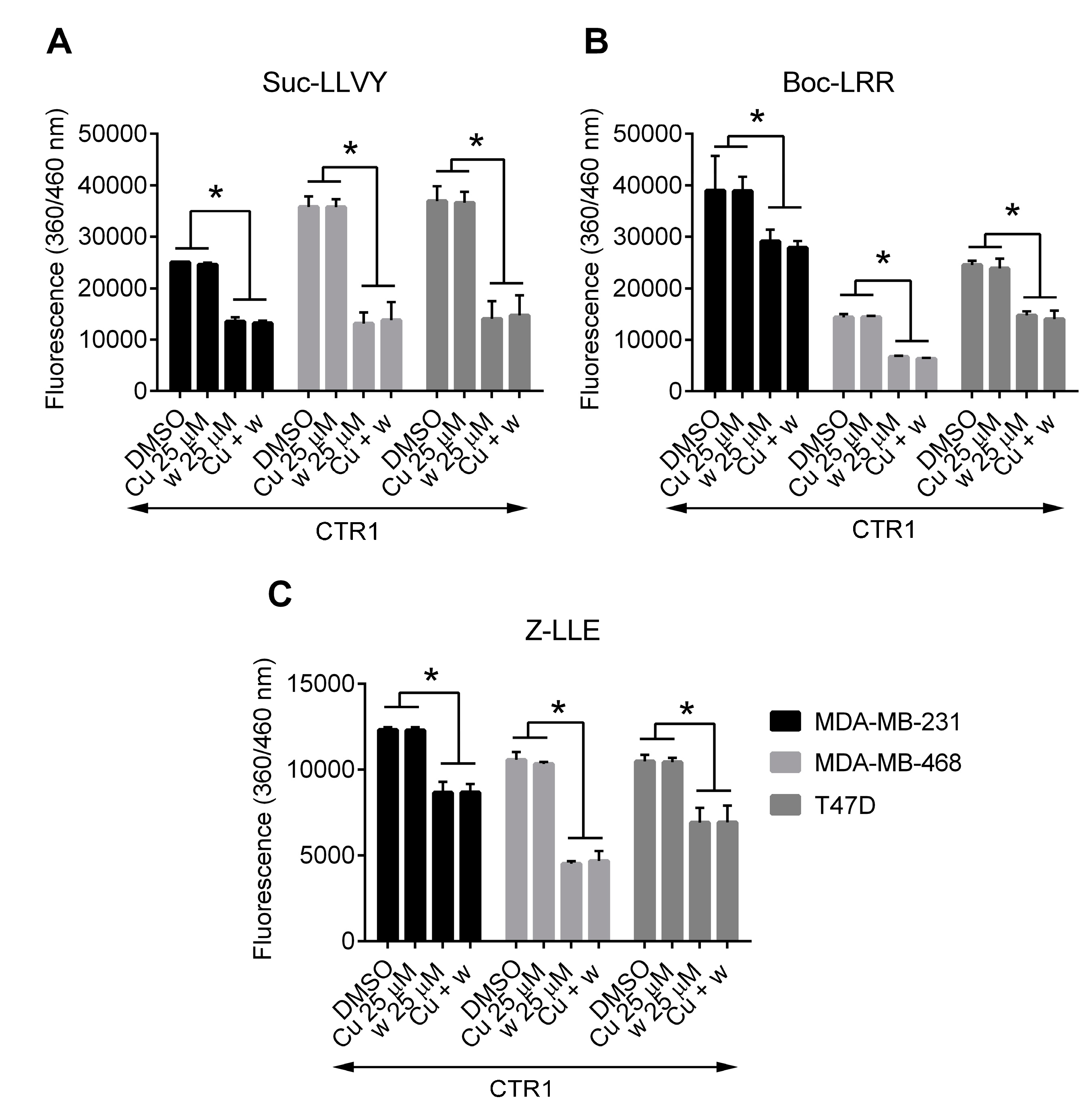
2.5. Copper Does Not Affect the Inhibition of Proteasome Activity by Wedelolactone in Breast Cancer Cell Lines
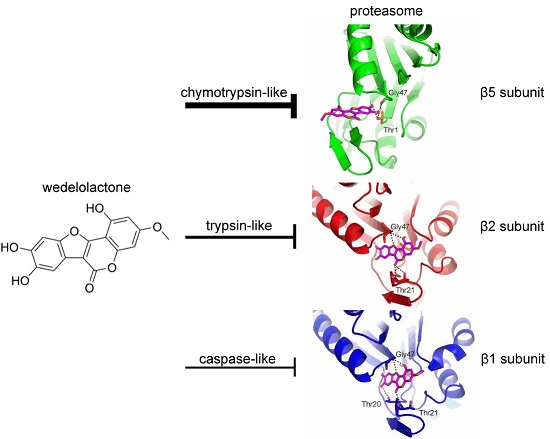
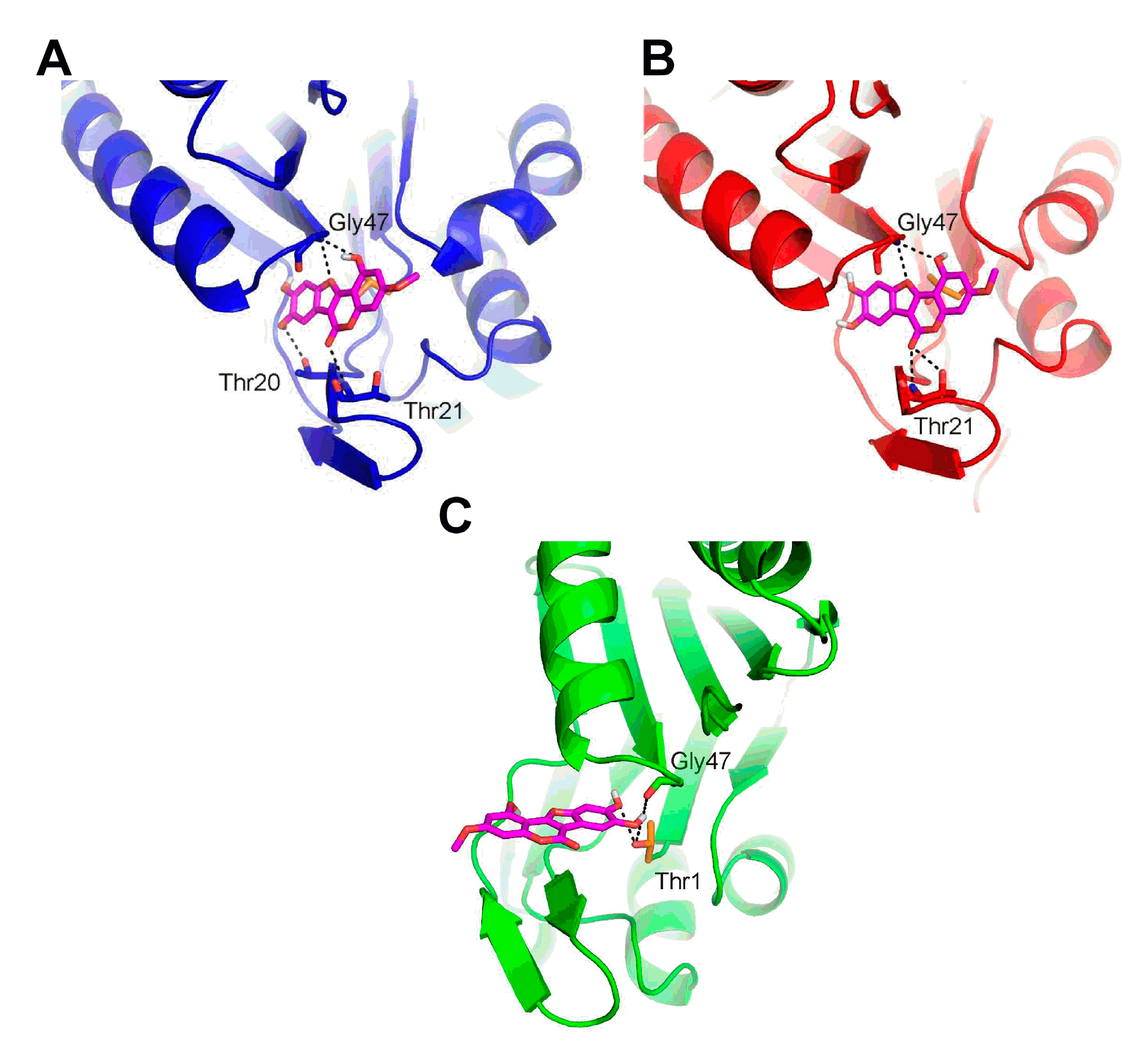
2.6. Molecular Docking of Wedelolactone to the Active Sites of Proteasome
3. Discussion
4. Material and Methods
4.1. Chemicals and Plasmids
4.2. Cell Culture
4.3. Proteasome Activity Assay
4.3.1. Purification of 26S Proteasome from MDA-MB-231 Cells
4.3.2. Proteasome Activity In Vitro
4.3.3. Proteasome Activity in Cancer Cell Lines
4.3.4. Proteasome Activity after Copper-Overloading
4.4. Immunoblotting
4.5. RNA Isolation, cDNA Synthesis and qPCR
4.6. Cell Mortality
4.7. Reactive Oxygen Species Production Analysis
4.8. Analysis of Copper Concentrations in Cells
4.9. Molecular Docking
4.10. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DHE | Dihydroethidium |
| DMSO | Dimethyl sulfoxide |
| ECL | Enhanced chemiluminescence |
| FCS | Fetal calf serum |
| CTR1 | Human copper transporter 1 |
| PDB | Protein data bank |
| PI | Propidium iodide |
| ROS | Reactive oxygen species |
| SALD ICP MS | Substrate-assisted laser desorption inductively-coupled plasma mass spectrometry |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| qPCR | Quantitative polymerase chain reaction |
| UPS | Ubiquitin-proteasome system |
References
- Groll, M.; Heinemeyer, W.; Jäger, S.; Ullrich, T.; Bochtler, M.; Wolf, D.H.; Huber, R. The catalytic sites of 20S proteasomes and their role in subunit maturation: A mutational and crystallographic study. Proc. Natl. Acad. Sci. USA 1999, 96, 10976–10983. [Google Scholar] [CrossRef] [PubMed]
- Heinemeyer, W.; Fischer, M.; Krimmer, T.; Stachon, U.; Wolf, D.H. The active sites of the eukaryotic 20S proteasome and their involvement in subunit precursor processing. J. Biol. Chem. 1997, 272, 25200–25209. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Schmitt, S.; Buac, D.; Duo, Q.P. Targeting the ubiquitin–proteasome system for cancer therapy. Expert Opin. Ther. Targets 2013, 17, 1091–1108. [Google Scholar] [CrossRef] [PubMed]
- Groll, M.; Huber, R. Purification, crystallization, and X-ray analysis of the yeast 20S proteasome. Methods Enzymol. 2005, 398, 329–336. [Google Scholar] [PubMed]
- Grigoreva, T.A.; Tribulovich, V.G.; Garabadzhiu, A.V.; Melino, G.; Barlev, N.A. The 26S proteasome is a multifaceted target for anti-cancer therapies. Oncotarget 2015, 6, 24733–24749. [Google Scholar] [CrossRef] [PubMed]
- Crawford, L.J.; Walker, B.; Irvine, A.E. Proteasome inhibitors in cancer therapy. J. Cell Commun. Signal. 2011, 5, 101–110. [Google Scholar] [PubMed]
- Arlt, A.; Bauer, I.; Schafmayer, C.; Tepel, J.; Müerköster, S.S.; Brosch, M.; Röder, C.; Kalthoff, H.; Hampe, J.; Moyer, M.P.; et al. Increased proteasome subunit protein expression and proteasome activity in colon cancer relate to an enhanced activation of nuclear factor E2-related factor 2 (Nrf2). Oncogene 2009, 28, 3983–3996. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.V.; Richardson, P.G.; Hideshima, T.; Anderson, K.C. Proteasome Inhibition As a Novel Therapeutic Target in Human Cancer. J. Cell. Oncol. 2005, 23, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Kubiczkova, L.; Pour, L.; Sedlarikova, L.; Hajek, R.; Sevcikova, S. Proteasome inhibitors—Molecular basis and current perspectives in multiple myeloma. J. Cell. Mol. Med. 2014, 18, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; van der Linden, W.A.; Overkleeft, H.S. Proteasome inhibitors: An expanding army attacking a unique target. Chem. Biol. 2012, 19, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Geyer, B.; Kiso, Y.; Hikino, H.; Rao, G.S. Coumestans as the main active principles of the liver drugs Eclipta alba and Wedelia calendulacea. Planta Med. 1986, 5, 370–374. [Google Scholar] [CrossRef]
- Benes, P.; Knopfova, L.; Trcka, F.; Nemajerova, A.; Pinheiro, D.; Soucek, K.; Fojta, M.; Smarda, J. Inhibition of topoisomerase IIα: Novel function of wedelolactone. Cancer Lett. 2011, 303, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, X.; Shen, S.; Zhang, H.; Ma, X.; Liu, J.; Kuang, S.; Yu, Q. Wedelolactone a naturally occurring coumestan, enhances interferon-G signaling through inhibiting STAT1 protein dephosphorylation. J. Biol. Chem. 2013, 288, 14417–14427. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.I.; Libouban, H.; Nyangoga, H.; Landao-Bassonga, E.; Chappard, D.; Ralston, S.H. Pharmacologic inhibitors of IkappaB kinase suppress growth and migration of mammary carcinosarcoma cells in vitro and prevent osteolytic bone metastasis in vivo. Mol. Cancer Ther. 2009, 8, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.M.; Chen, L.R.; Lin, E.H.; Ke, F.C.; Chen, H.Y.; Tsai, M.J.; Hsiao, P.W. Compounds from Wedelia chinensis synergistically suppress androgen activity and growth in prostate cancer cells. Carcinogenesis 2007, 28, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Nehybova, T.; Smarda, J.; Benes, P. Plant coumestans: Recent advances and future perspectives in cancer therapy. Anticancer Agents Med. Chem. 2014, 14, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Sukumari-Ramesh, S.; Bentley, J.N.; Laird, M.D.; Singh, N.; Vender, J.R.; Dhandapani, K.M. Dietary phytochemicals induce p53- and caspase-independent cell death in human neuroblastoma cells. Int. J. Dev. Neurosci. 2011, 29, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Lin, F.M.; Yang, Y.C.; Lee, M.T.; Cha, T.L.; Wu, G.J.; Hsieh, S.C.; Hsiao, P.W. Herbal extract of Wedelia chinensis attenuates androgen receptor activity and orthotopic growth of prostate cancer in nude mice. Clin. Cancer Res. 2009, 15, 5435–5444. [Google Scholar] [CrossRef] [PubMed]
- Vender, J.R.; Laird, M.D.; Dhandapani, K.M. Inhibition of NFκB reduces cellular viability in GH3 pituitary adenoma cells. Neurosurgery 2008, 62, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Sarveswaran, S.; Gautam, S.C.; Ghosh, J. Wedelolactone a medicinal plant-derived coumestan, induces caspase-dependent apoptosis in prostate cancer cells via downregulation of PKCe without inhibiting Akt. Int. J. Oncol. 2012, 41, 2191–2199. [Google Scholar] [PubMed]
- Sarweswaran, S.; Ghosh, R.; Parikh, R.; Ghosh, J. Wedelolactone, an Anti-inflammatory Botanical, Interrupts c-Myc Oncogenic Signaling and Synergizes with Enzalutamide to Induce Apoptosis in Prostate Cancer Cells. Mol. Cancer Ther. 2016, 15, 2791–2801. [Google Scholar] [CrossRef] [PubMed]
- Benes, P.; Alexova, P.; Knopfova, L.; Spanova, A.; Smarda, J. Redox state alters anti-cancer effects of wedelolactone. Environ. Mol. Mutagen. 2012, 53, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Daniel, K.G.; Gupta, P.; Harbach, R.H.; Guida, W.C.; Dou, Q.P. Organic copper complexes as a new class of proteasome inhibitors and apoptosis inducers in human cancer cells. Biochem. Pharmacol. 2004, 67, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.Q.; Liu, B.; Vaught, J.L.; Yamauchi, H.; Lind, S.E. Anticancer activity of the antibiotic clioquinol. Cancer Res. 2005, 65, 3389–3395. [Google Scholar] [PubMed]
- Arif, H.; Rehmani, N.; Farhan, M.; Ahmad, A.; Hadi, S.M. Mobilization of Copper ions by Flavonoids in Human Peripheral Lymphocytes Leads to Oxidative DNA Breakage: A Structure Activity Study. Int. J. Mol. Sci. 2015, 16, 26754–26769. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Khan, H.Y.; Oves, M.; Al-Harrasi, A.; Rehmani, N.; Arif, H.; Hadi, S.M.; Ahmad, A. Cancer Therapy by Catechins Involves Redox Cycling of Copper Ions and Generation of Reactive Oxygen species. Toxins 2016, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Faisal, M.; Ullah, M.F.; Farhan, M.; Sarkar, F.H.; Ahmad, A.; Hadi, S.M. Plant polyphenol induced cell death in human cancer cells involves mobilization of intracellular copper ions and reactive oxygen species generation: A mechanism for cancer chemopreventive action. Mol. Nutr. Food Res. 2014, 58, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Singh, S.; Naseem, I. Cytotoxic activity of soy phytoestrogen coumestrol against human breast cancer MCF-7 cells: Insights into the molecular mechanism. Food Chem. Toxicol. 2017, 99, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.D.; Lim, T.M. Dopamine (DA) induced irreversible proteasome inhibition via DA derived quinones. Free Radic. Res. 2009, 43, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Adams, J. The proteasome: Structure, function, and role in the cell. Cancer Treat. Rev. 2003, 29, 3–9. [Google Scholar] [CrossRef]
- Moore, B.S.; Eustáquio, A.S.; McGlinchey, R.P. Advances in and applications of proteasome inhibitors. Curr. Opin. Chem. Biol. 2008, 12, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Hochstrasser, M. Ubiquitin, proteasomes, and the regulation of intracellular protein degradation. Curr. Opin. Cell Biol. 1995, 7, 215–223. [Google Scholar] [CrossRef]
- Bae, S.H.; Ryoo, H.M.; Kim, M.K.; Lee, K.H.; Sin, J.I.; Hyun, M.S. Effects of the proteasome inhibitor bortezomib alone and in combination with chemotherapeutic agents in gastric cancer cell lines. Oncol. Rep. 2008, 19, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Smith, D.M.; Dou, P.D. Tannic Acid Potently Inhibits Tumor Cell Proteasome Activity, Increases p27 and Bax Expression, and Induces G1 Arrest and Apoptosis. Cancer Epidemiol. Biomark. Prev. 2001, 10, 1083–1088. [Google Scholar]
- Yang, H.; Landis-Piwowar, K.R.; Chen, D.; Milacic, V.; Dou, Q.P. Natural Compounds with Proteasome Inhibitory Activity for Cancer Prevention and Treatment. Curr. Protein Pept. Sci. 2008, 9, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Peña, M.M.; Nose, Y.; Thiele, D.J. Biochemical characterization of the human copper transporter Ctr1. J. Biol. Chem. 2002, 277, 4380–4387. [Google Scholar] [CrossRef] [PubMed]
- Nehybova, T.; Smarda, J.; Daniel, L.; Brezovsky, J.; Benes, P. Wedelolactone induces growth of breast cancer cells by stimulation of estrogen receptor signalling. J. Steroid. Biochem. Mol. Biol. 2015, 152, 76–83. [Google Scholar] [PubMed]
- Jancekova, B.; Ondrouskova, E.; Knopfova, L.; Smarda, J.; Benes, P. Enzymatically active cathepsin D sensitizes breast carcinoma cells to TRAIL. Tumour Biol. 2016, 37, 10685–10696. [Google Scholar] [CrossRef] [PubMed]
- Navratilova, J.; Hankeova, T.; Benes, P.; Smarda, J. Acidic pH of tumor microenvironment enhances cytotoxicity of the disulfiram/Cu2+ complex to breast and colon cancer cells. Chemotherapy 2013, 59, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Pes, O.; Jungova, P.; Vyhnanek, R.; Vaculovic, T.; Kanicky, V.; Preisler, J. Off-line coupling of capillary electrophoresis to substrate-assisted laser desorption inductively coupled plasma mass spectrometry. Anal. Chem. 2008, 80, 8725–8732. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC—A free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Delano, W.T. The PyMol Molecular Graphics System, Version 1.5; Schrödinger, LLC: New York, NY, USA, 2009. [Google Scholar]
- Durrant, J.; McCammon, J. NNScore 2.0: A Neural-Network Receptor-ligand Scoring Function. J. Chem. Inf. Model. 2011, 51, 2897–2903. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nehybová, T.; Šmarda, J.; Daniel, L.; Stiborek, M.; Kanický, V.; Spasojevič, I.; Preisler, J.; Damborský, J.; Beneš, P. Wedelolactone Acts as Proteasome Inhibitor in Breast Cancer Cells. Int. J. Mol. Sci. 2017, 18, 729. https://doi.org/10.3390/ijms18040729
Nehybová T, Šmarda J, Daniel L, Stiborek M, Kanický V, Spasojevič I, Preisler J, Damborský J, Beneš P. Wedelolactone Acts as Proteasome Inhibitor in Breast Cancer Cells. International Journal of Molecular Sciences. 2017; 18(4):729. https://doi.org/10.3390/ijms18040729
Chicago/Turabian StyleNehybová, Tereza, Jan Šmarda, Lukáš Daniel, Marek Stiborek, Viktor Kanický, Ivan Spasojevič, Jan Preisler, Jiří Damborský, and Petr Beneš. 2017. "Wedelolactone Acts as Proteasome Inhibitor in Breast Cancer Cells" International Journal of Molecular Sciences 18, no. 4: 729. https://doi.org/10.3390/ijms18040729