
Osteopontin Deficiency Suppresses Intestinal Tumor Development in Apc-Deficient Min Mice


Abstract
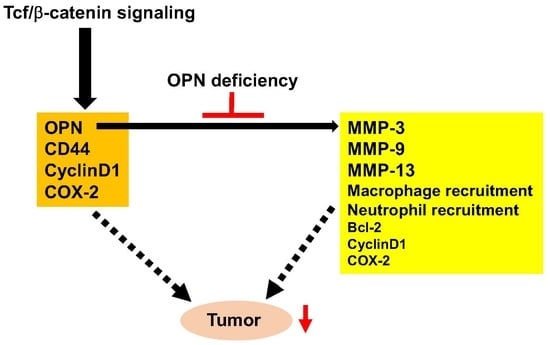
:
1. Introduction
2. Results
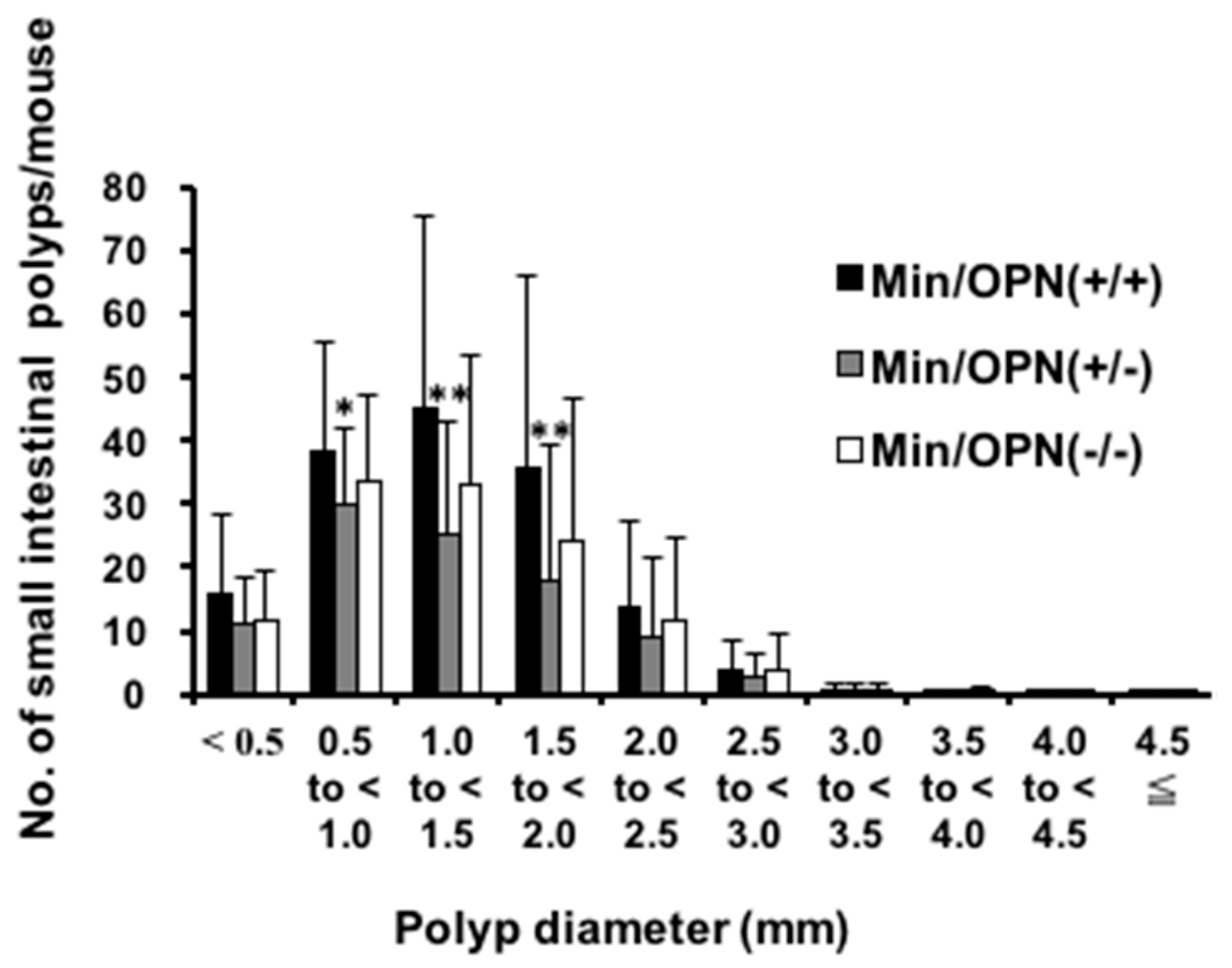
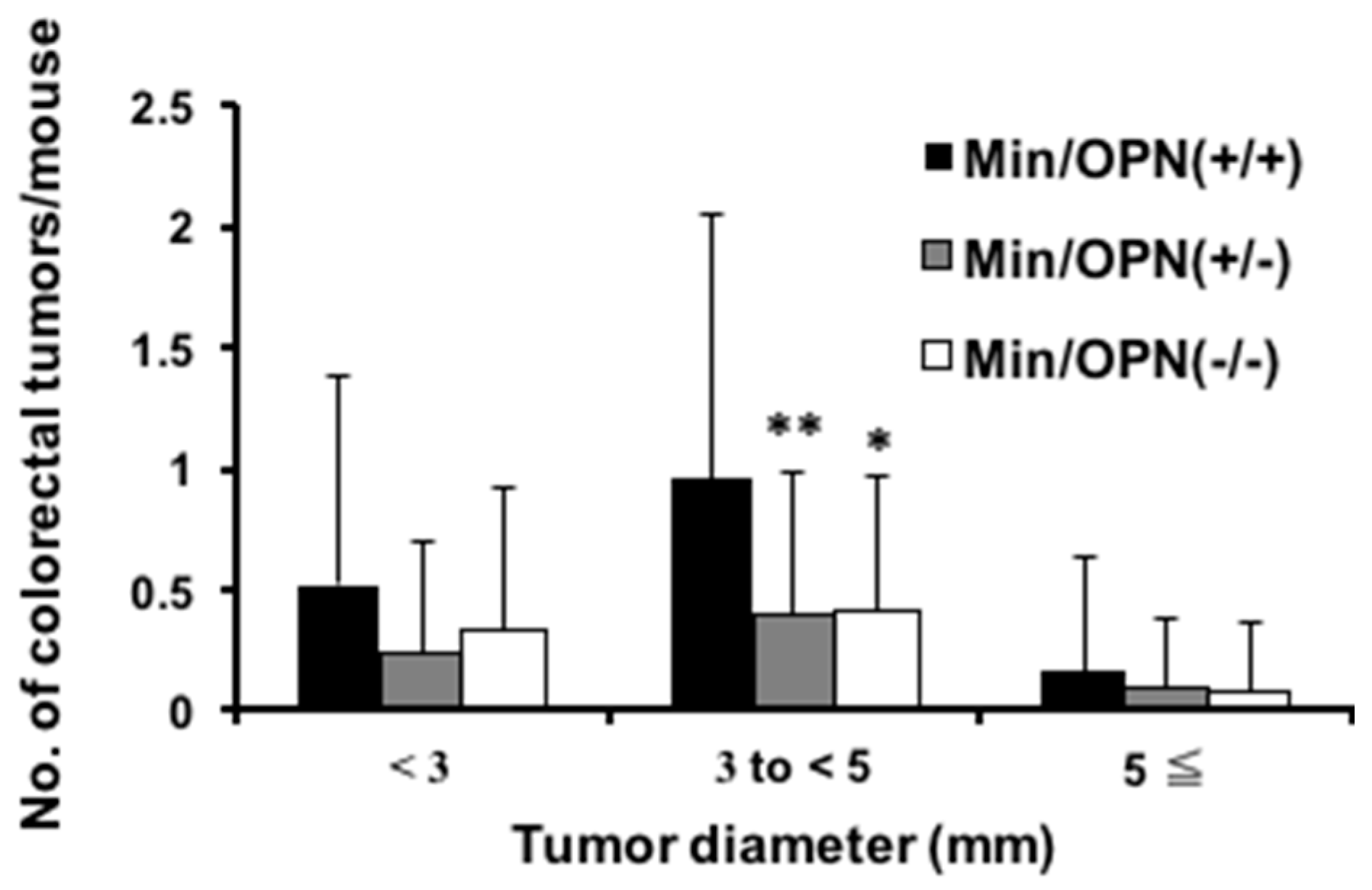
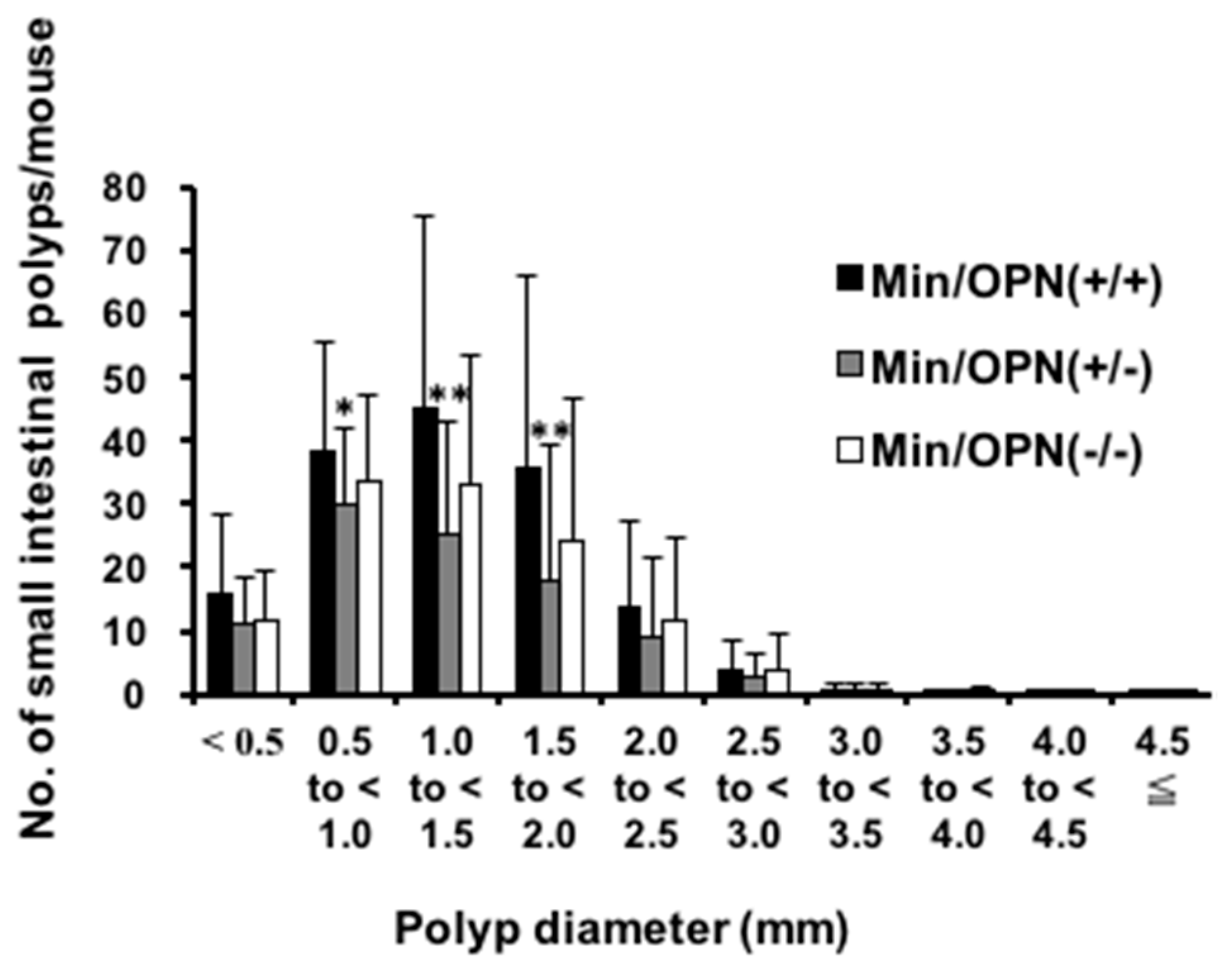
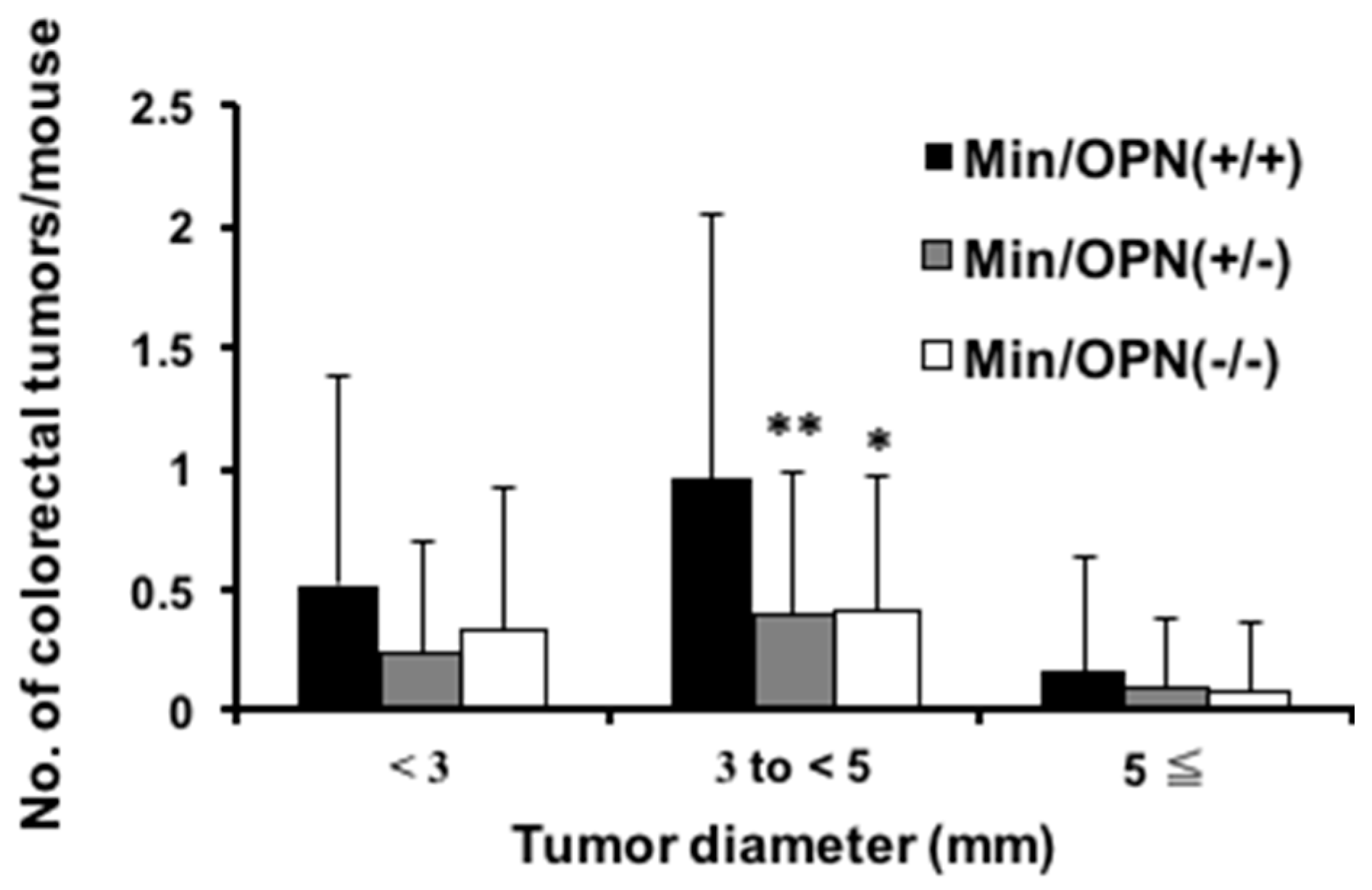
2.1. Effect of Osteopontin (OPN) Deficiency on Intestinal Polyp Formation in Min Mice
2.2. Serum Levels of OPN, Interleukin (IL)-6, and Triglycerides
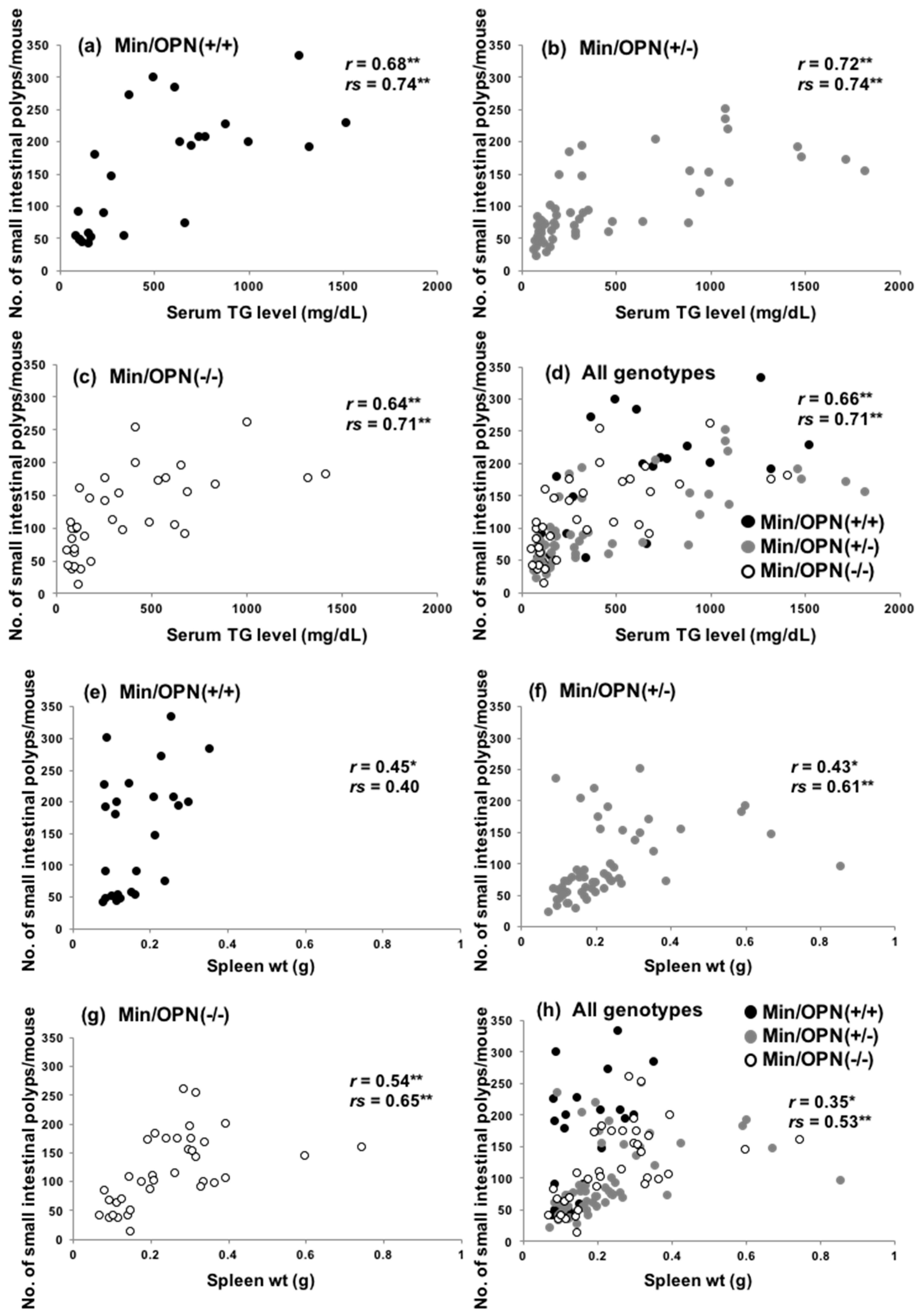
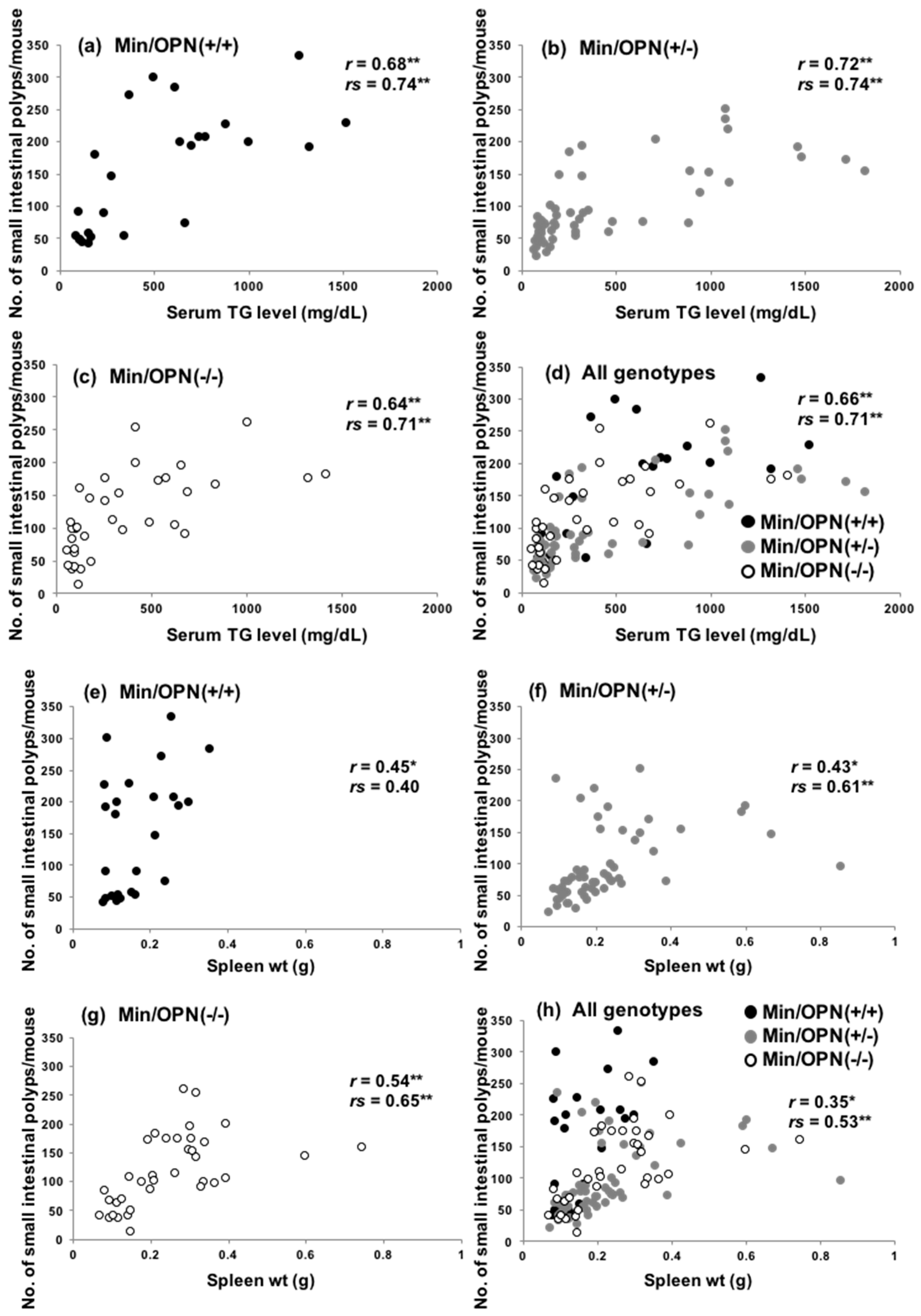
2.3. Correlation of Small Intestinal Polyp Numbers with Serum Levels of Triglycerides and Spleen Weights
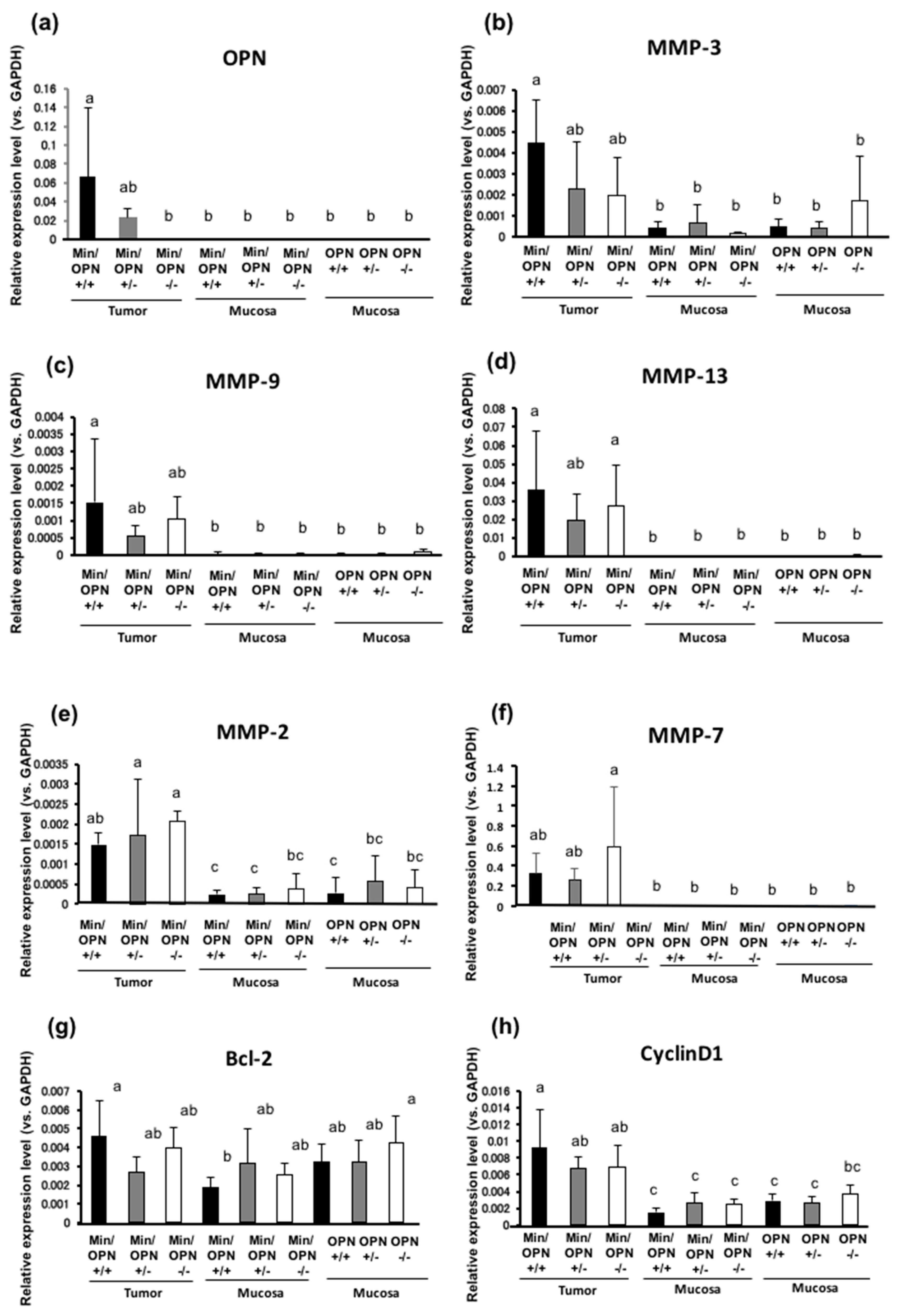
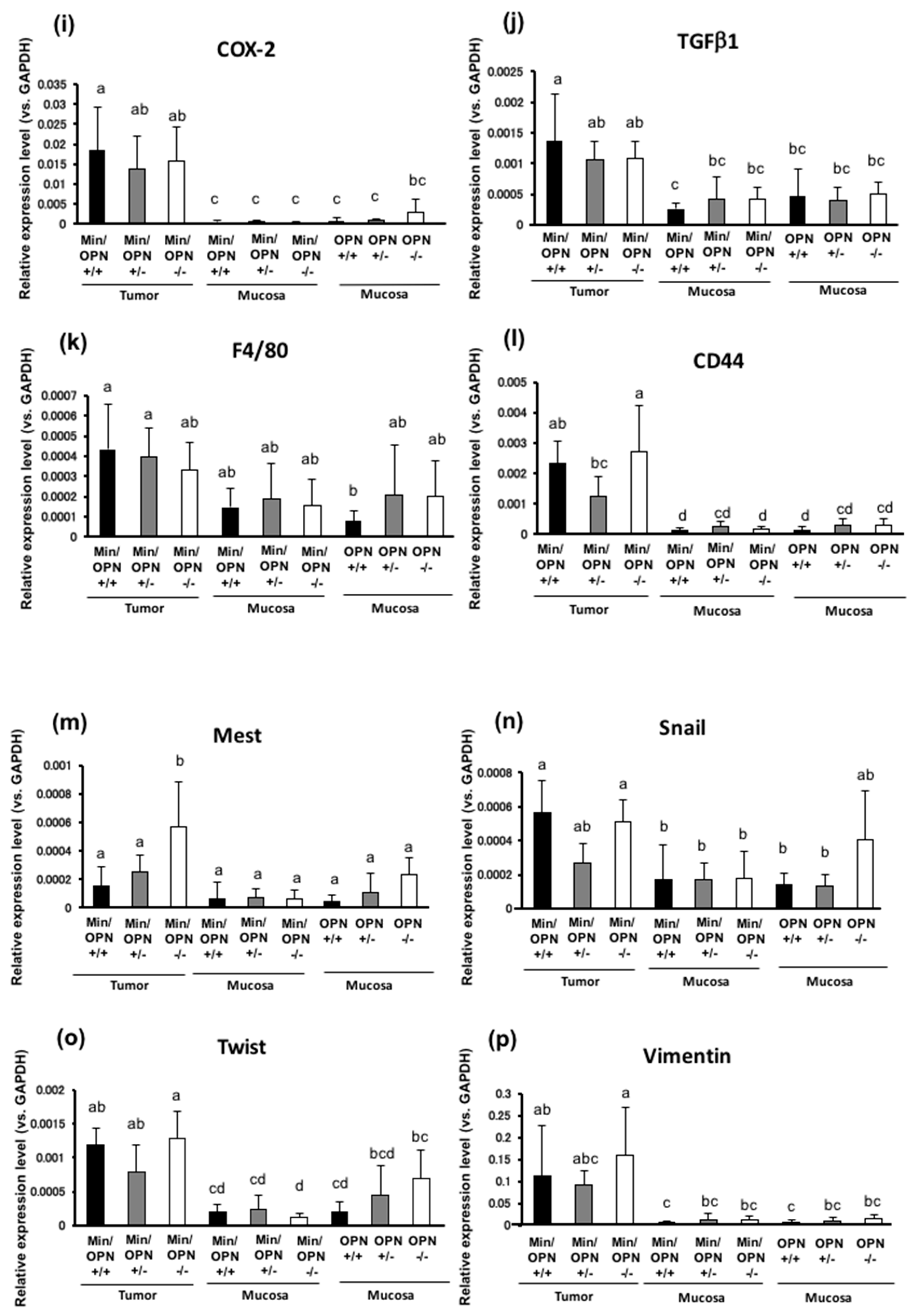
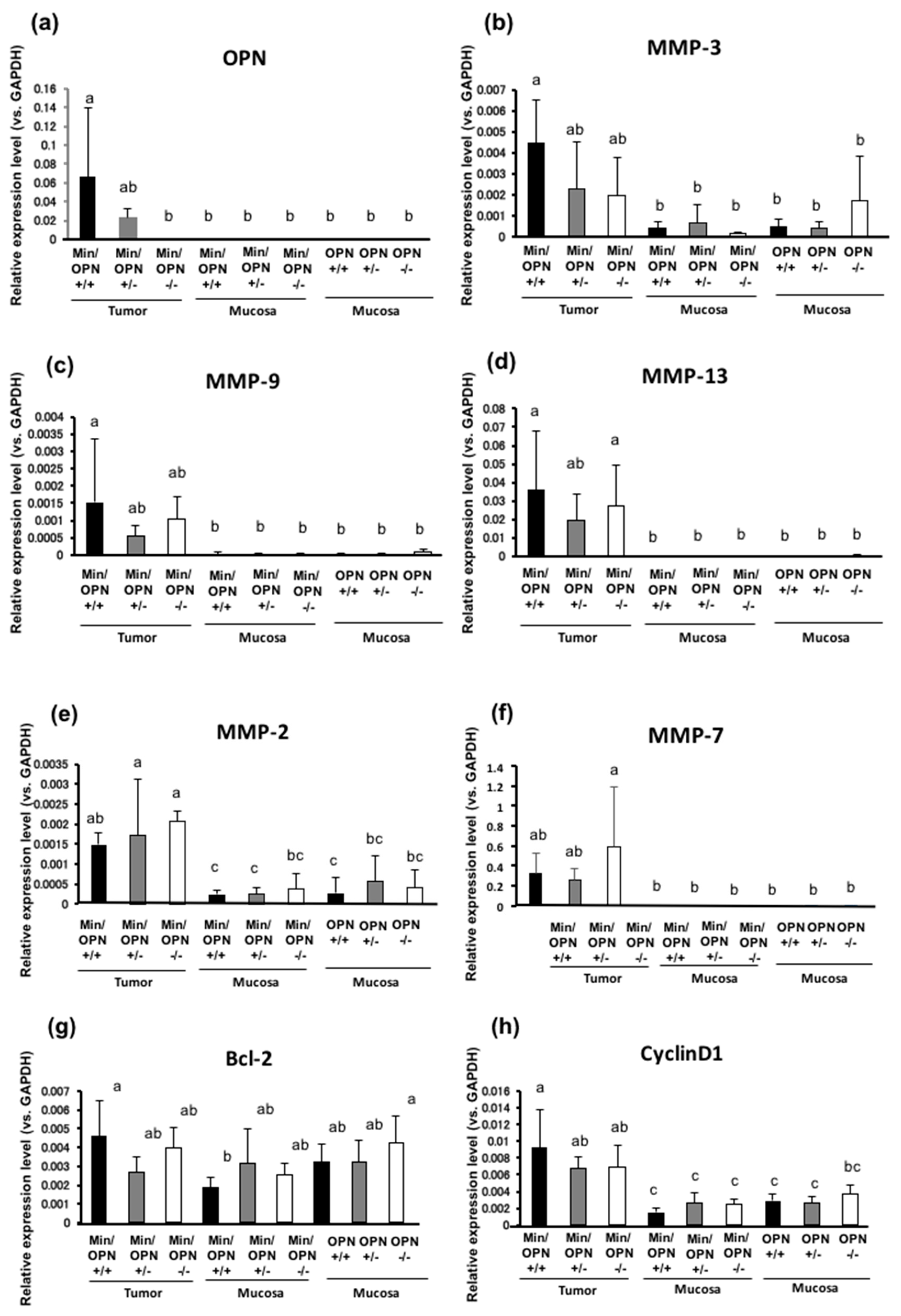
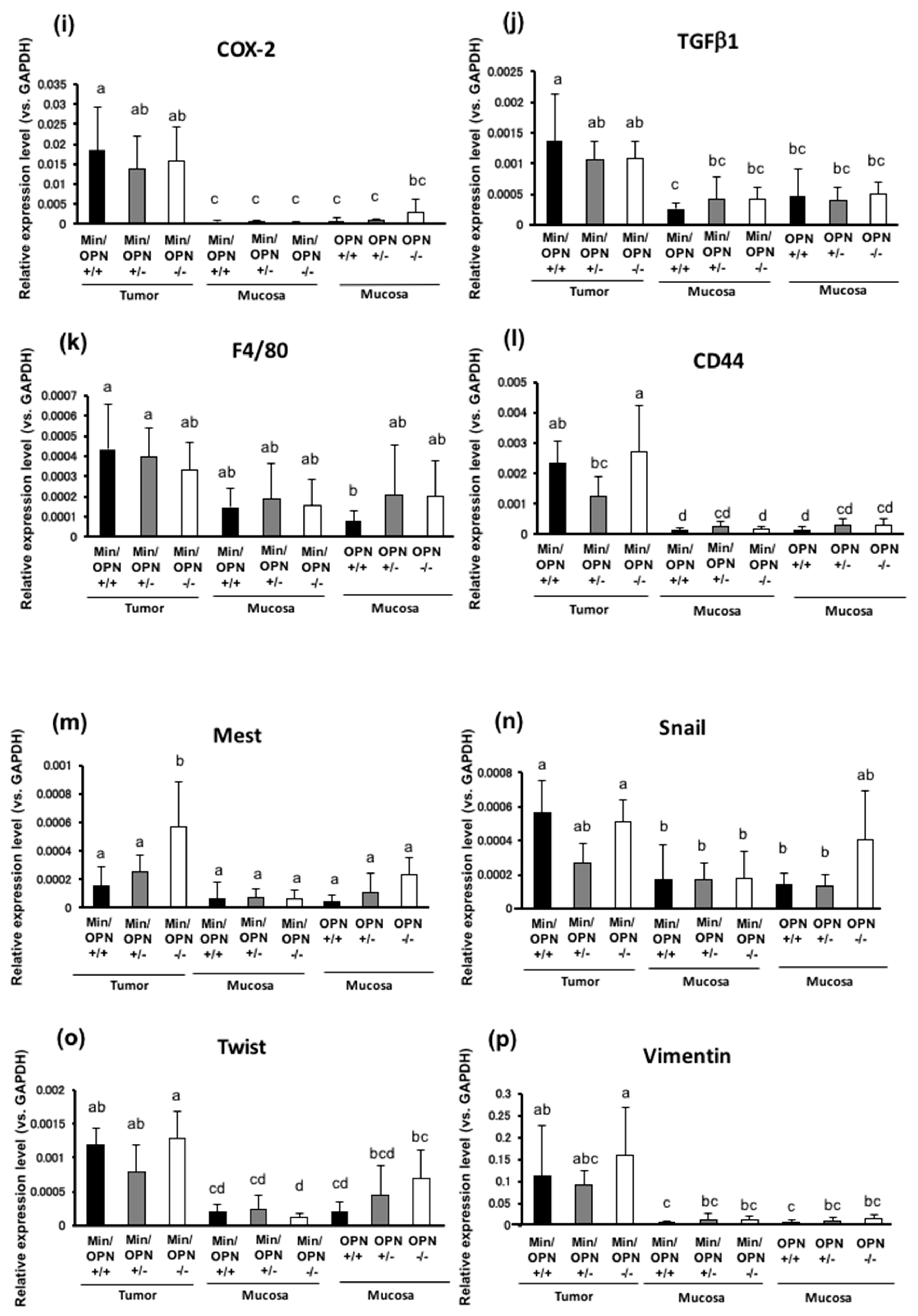
2.4. Effects of OPN deficiency on Gene Expression Levels in Colon Tumors
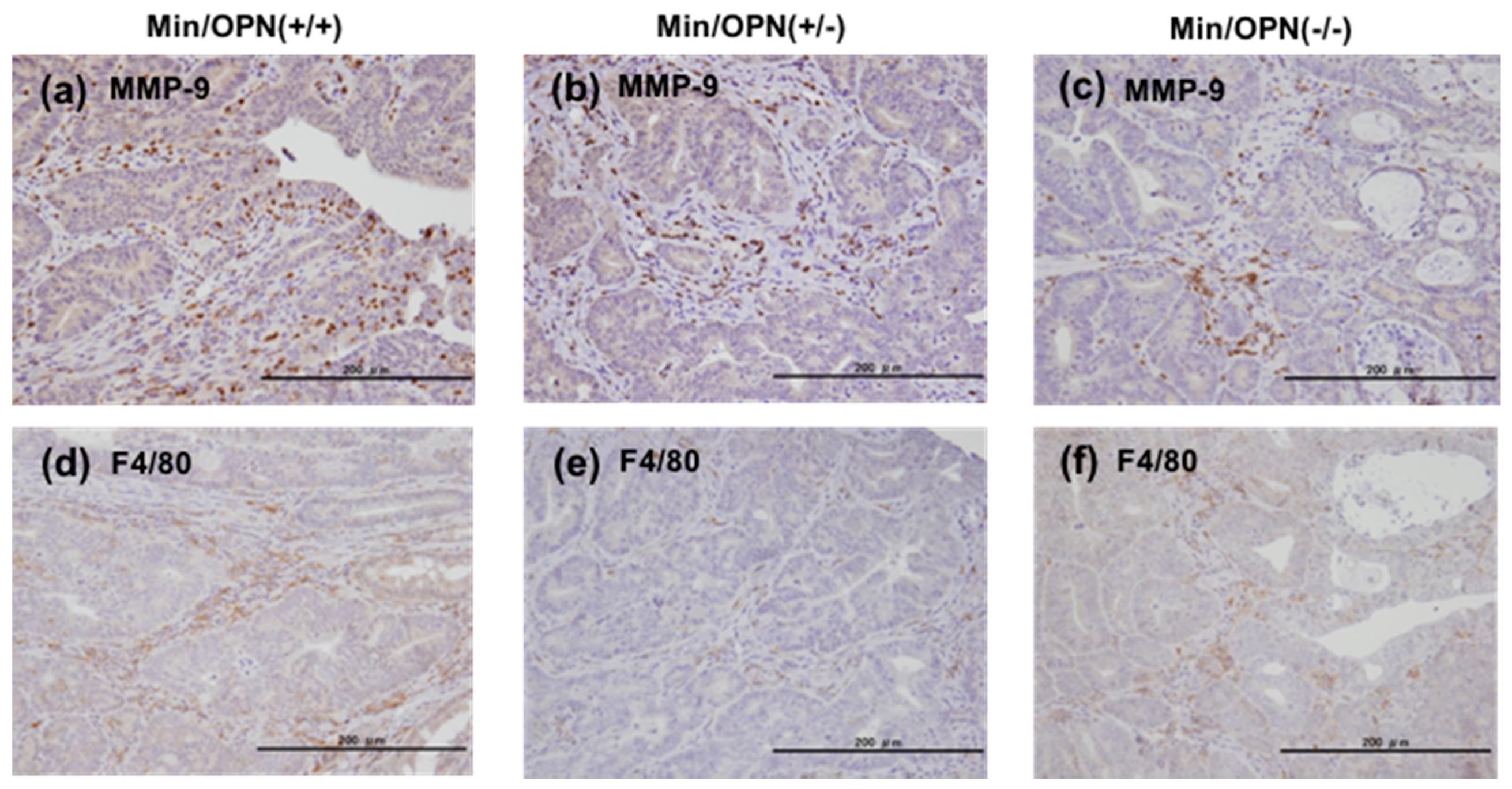
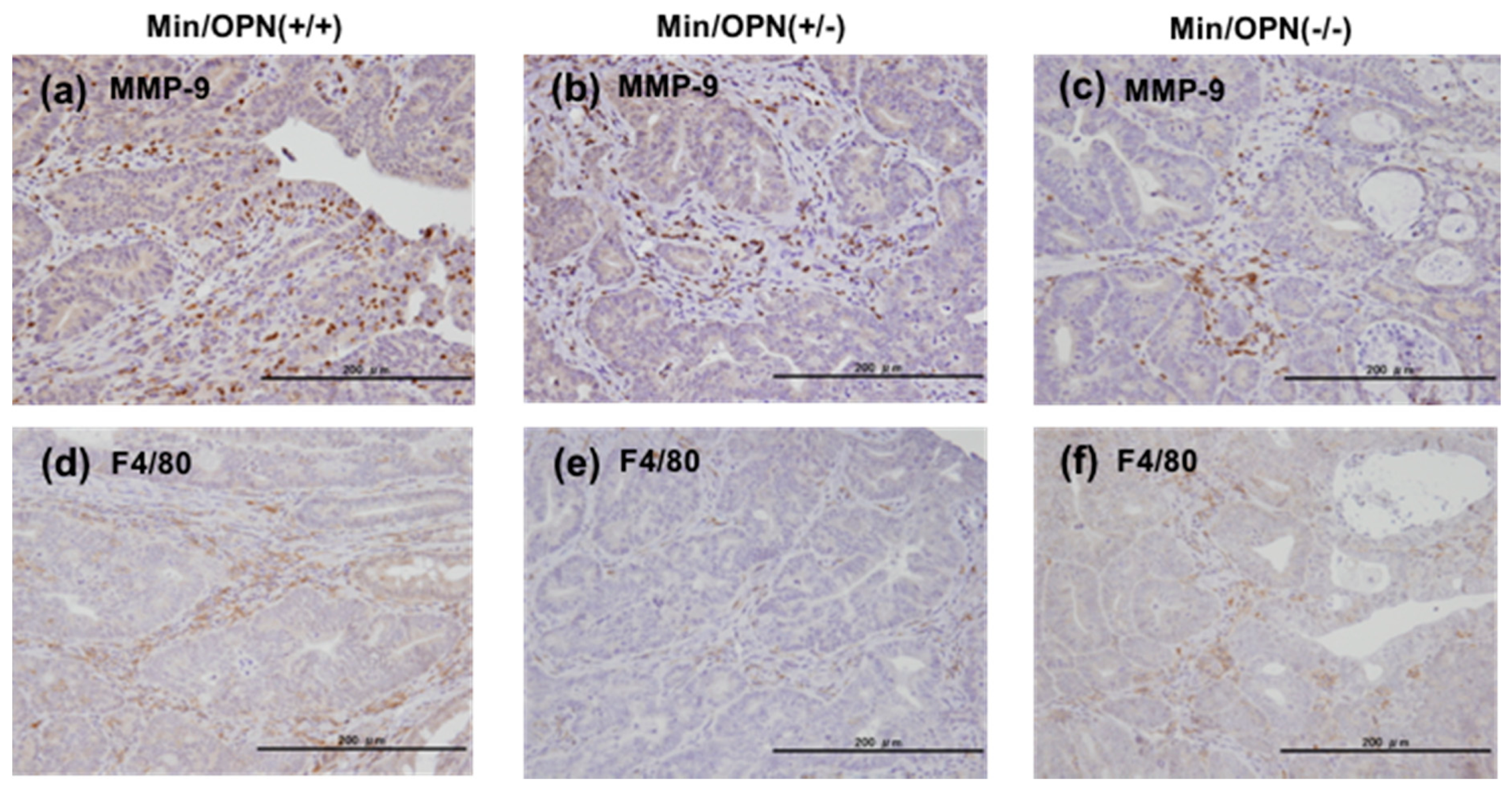
2.5. Protein Expression in Colon Tumors
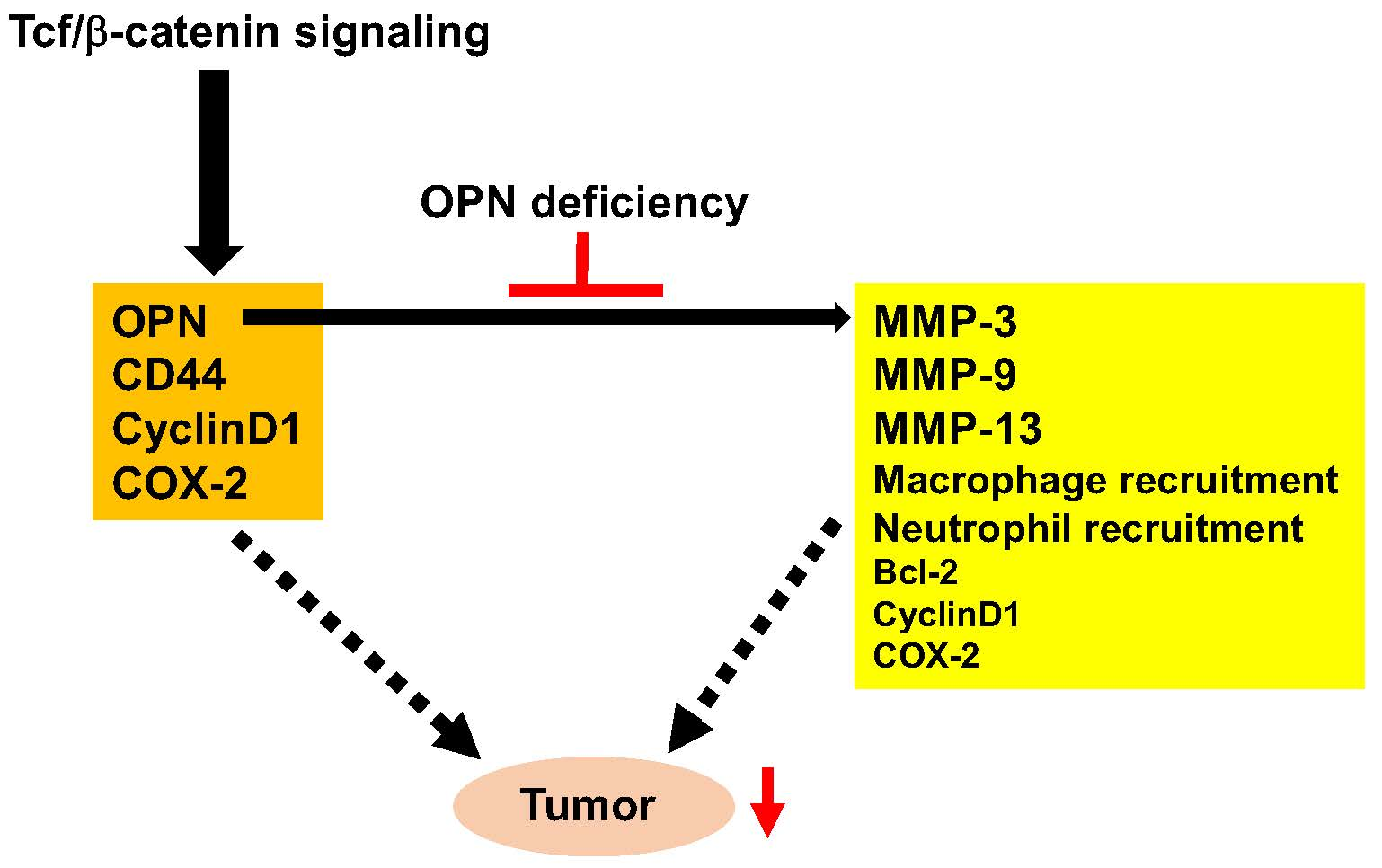
3. Discussion
4. Materials and Methods
4.1. Animals and Diets
4.2. Analysis of Intestinal Polyps
4.3. Measurement of Mouse Serum Parameter Levels
4.4. Quantitative RT-PCR Analysis
4.5. Immunohistochemical Staining of Colon Tumors
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, J.L.; Wang, M.J.; Sudhir, P.R.; Chen, G.D.; Chi, C.W.; Chen, J.Y. Osteopontin promotes integrin activation through outside-in and inside-out mechanisms: OPN-CD44V interaction enhances survival in gastrointestinal cancer cells. Cancer Res. 2007, 67, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell 2014, 14, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.F.; Papadopoulos-Sergiou, A.; Berse, B.; Manseau, E.J.; Tognazzi, K.; Perruzzi, C.A.; Dvorak, H.F.; Senger, D.R. Osteopontin expression and distribution in human carcinomas. Am. J. Pathol. 1994, 145, 610–623. [Google Scholar] [PubMed]
- Coppola, D.; Szabo, M.; Boulware, D.; Muraca, P.; Alsarraj, M.; Chambers, A.F.; Yeatman, T.J. Correlation of osteopontin protein expression and pathological stage across a wide variety of tumor histologies. Clin. Cancer Res. 2004, 10, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Fedarko, N.S.; Jain, A.; Karadag, A.; van Eman, M.R.; Fisher, L.W. Elevated serum bone sialoprotein and osteopontin in colon, breast, prostate, and lung cancer. Clin. Cancer Res. 2001, 7, 4060–4066. [Google Scholar] [PubMed]
- Cantor, H.; Shinohara, M.L. Regulation of T-helper-cell lineage development by osteopontin: The inside story. Nat. Rev. Immunol. 2009, 9, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Rittling, S.R. Osteopontin in macrophage function. Expert Rev. Mol. Med. 2011, 13, e15. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.X.; Denhardt, D.T. Osteopontin: role in immune regulation and stress responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Anborgh, P.H.; Mutrie, J.C.; Tuck, A.B.; Chambers, A.F. Role of the metastasis-promoting protein osteopontin in the tumour microenvironment. J. Cell. Mol. Med. 2010, 14, 2037–2044. [Google Scholar] [CrossRef] [PubMed]
- Rittling, S.R.; Chambers, A.F. Role of osteopontin in tumour progression. Br. J. Cancer 2004, 90, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Rohde, F.; Rimkus, C.; Friederichs, J.; Rosenberg, R.; Marthen, C.; Doll, D.; Holzmann, B.; Siewert, J.R.; Janssen, K.P. Expression of osteopontin, a target gene of de-regulated Wnt signaling, predicts survival in colon cancer. Int. J. Cancer 2007, 121, 1717–1723. [Google Scholar] [CrossRef] [PubMed]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin-A Master Regulator of Epithelial-Mesenchymal Transition. J. Clin. Med. 2016, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.P.; Shan, Y.S.; Lai, M.D.; Lin, P.W. Osteopontin-positive infiltrating tumor-associated macrophages in bulky ampullary cancer predict survival. Cancer Biol. Ther. 2010, 10, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Irby, R.B.; McCarthy, S.M.; Yeatman, T.J. Osteopontin regulates multiple functions contributing to human colon cancer development and progression. Clin. Exp. Metastasis 2004, 21, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Pan, C.; Hu, H.; Zheng, S.; Ding, L. Osteopontin-enhanced hepatic metastasis of colorectal cancer cells. PLoS ONE 2012, 7, e47901. [Google Scholar] [CrossRef] [PubMed]
- Wai, P.Y.; Mi, Z.; Guo, H.; Sarraf-Yazdi, S.; Gao, C.; Wei, J.; Marroquin, C.E.; Clary, B.; Kuo, P.C. Osteopontin silencing by small interfering RNA suppresses in vitro and in vivo CT26 murine colon adenocarcinoma metastasis. Carcinogenesis 2005, 26, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.L.; Lin, K.J.; Bai, A.P.; Wang, W.X.; Meng, X.K.; Su, X.L.; Hou, M.X.; Dong, P.D.; Zhang, J.J.; Wang, Z.Y.; et al. Osteopontin knockdown suppresses the growth and angiogenesis of colon cancer cells. World J. Gastroenterol. 2014, 20, 10440–10448. [Google Scholar] [CrossRef] [PubMed]
- Shevde, L.A.; Das, S.; Clark, D.W.; Samant, R.S. Osteopontin: an effector and an effect of tumor metastasis. Curr. Mol. Med. 2010, 10, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, M.; Bulbule, A.; Butti, R.; Chakraborty, G.; Ghorpade, P.; Ghosh, P.; Gorain, M.; Kale, S.; Kumar, D.; Kumar, S.; et al. Osteopontin as a therapeutic target for cancer. Expert Opin. Ther. Targets 2014, 18, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Johnston, N.I.; Gunasekharan, V.K.; Ravindranath, A.; O’Connell, C.; Johnston, P.G.; El-Tanani, M.K. Osteopontin as a target for cancer therapy. Front. Biosci. 2008, 13, 4361–4372. [Google Scholar] [CrossRef] [PubMed]
- Bardou, M.; Barkun, A.N.; Martel, M. Obesity and colorectal cancer. Gut 2013, 62, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Tsugane, S.; Inoue, M. Insulin resistance and cancer: epidemiological evidence. Cancer Sci. 2010, 101, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.; Miles, P.D.; Ofrecio, J.M.; Neels, J.G.; Yu, J.G.; Resnik, J.L.; Wilkes, J.; Talukdar, S.; Thapar, D.; Johnson, K.; et al. Osteopontin is required for the early onset of high fat diet-induced insulin resistance in mice. PLoS ONE 2010, 5, e13959. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Gollinger, K.; Todoric, J.; Kiefer, F.W.; Keck, M.; Aszmann, O.; Prager, G.; Zlabinger, G.J.; Petzelbauer, P.; Stulnig, T.M. Osteopontin is an activator of human adipose tissue macrophages and directly affects adipocyte function. Endocrinology 2011, 152, 2219–2227. [Google Scholar] [CrossRef] [PubMed]
- Lancha, A.; Rodriguez, A.; Catalan, V.; Becerril, S.; Sainz, N.; Ramirez, B.; Burrell, M.A.; Salvador, J.; Fruhbeck, G.; Gomez-Ambrosi, J. Osteopontin deletion prevents the development of obesity and hepatic steatosis via impaired adipose tissue matrix remodeling and reduced inflammation and fibrosis in adipose tissue and liver in mice. PLoS ONE 2014, 9, e98398. [Google Scholar] [CrossRef] [PubMed]
- Tardelli, M.; Zeyda, K.; Moreno-Viedma, V.; Wanko, B.; Grun, N.G.; Staffler, G.; Zeyda, M.; Stulnig, T.M. Osteopontin is a key player for local adipose tissue macrophage proliferation in obesity. Mol. Metab. 2016, 5, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, F.W.; Zeyda, M.; Gollinger, K.; Pfau, B.; Neuhofer, A.; Weichhart, T.; Saemann, M.D.; Geyeregger, R.; Schlederer, M.; Kenner, L.; et al. Neutralization of osteopontin inhibits obesity-induced inflammation and insulin resistance. Diabetes 2010, 59, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Catalan, V.; Gomez-Ambrosi, J.; Rodriguez, A.; Ramirez, B.; Izaguirre, M.; Hernandez-Lizoain, J.L.; Baixauli, J.; Marti, P.; Valenti, V.; Moncada, R.; et al. Increased Obesity-Associated Circulating Levels of the Extracellular Matrix Proteins Osteopontin, Chitinase-3 Like-1 and Tenascin C Are Associated with Colon Cancer. PLoS ONE 2016, 11, e0162189. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, J.W.; Go, D.M.; Kim, H.K.; Kwon, H.J.; Han, S.U.; Kim, D.Y. Ablation of osteopontin suppresses N-methyl-N-nitrosourea and Helicobacter pylori-induced gastric cancer development in mice. Carcinogenesis 2015, 36, 1550–1560. [Google Scholar] [PubMed]
- Lee, S.H.; Park, J.W.; Woo, S.H.; Go, D.M.; Kwon, H.J.; Jang, J.J.; Kim, D.Y. Suppression of osteopontin inhibits chemically induced hepatic carcinogenesis by induction of apoptosis in mice. Oncotarget 2016, 7, 87219–87231. [Google Scholar] [CrossRef] [PubMed]
- Miyaki, M.; Konishi, M.; Kikuchi-Yanoshita, R.; Enomoto, M.; Igari, T.; Tanaka, K.; Muraoka, M.; Takahashi, H.; Amada, Y.; Fukayama, M.; et al. Characteristics of somatic mutation of the adenomatous polyposis coli gene in colorectal tumors. Cancer Res. 1994, 54, 3011–3020. [Google Scholar] [PubMed]
- Morin, P.J.; Sparks, A.B.; Korinek, V.; Barker, N.; Clevers, H.; Vogelstein, B.; Kinzler, K.W. Activation of β-catenin-Tcf signaling in colon cancer by mutations in β-catenin or APC. Science 1997, 275, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Moser, A.R.; Pitot, H.C.; Dove, W.F. A dominant mutation that predisposes to multiple intestinal neoplasia in the mouse. Science 1990, 247, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Niho, N.; Takahashi, M.; Kitamura, T.; Shoji, Y.; Itoh, M.; Noda, T.; Sugimura, T.; Wakabayashi, K. Concomitant suppression of hyperlipidemia and intestinal polyp formation in Apc-deficient mice by peroxisome proliferator-activated receptor ligands. Cancer Res. 2003, 63, 6090–6095. [Google Scholar] [PubMed]
- Hodgson, A.; Wier, E.M.; Fu, K.; Sun, X.; Wan, F. Ultrasound imaging of splenomegaly as a proxy to monitor colon tumor development in Apcmin716/+ mice. Cancer Med 2016, 5, 2469–2476. [Google Scholar] [CrossRef] [PubMed]
- Mehl, K.A.; Davis, J.M.; Clements, J.M.; Berger, F.G.; Pena, M.M.; Carson, J.A. Decreased intestinal polyp multiplicity is related to exercise mode and gender in ApcMin/+ mice. J. Appl. Physioi. 2005, 98, 2219–2225. [Google Scholar] [CrossRef] [PubMed]
- Wielenga, V.J.; Smits, R.; Korinek, V.; Smit, L.; Kielman, M.; Fodde, R.; Clevers, H.; Pals, S.T. Expression of CD44 in Apc and Tcf mutant mice implies regulation by the WNT pathway. Am. J. Pathol. 1999, 154, 515–523. [Google Scholar] [CrossRef]
- Jung, H.; Lee, S.K.; Jho, E.H. Mest/Peg1 inhibits Wnt signalling through regulation of LRP6 glycosylation. Biochem. J. 2011, 436, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Washington, M.K.; Saxena, R.; Sheng, H. Heterozygous disruption of the PTEN promotes intestinal neoplasia in APCmin/+ mouse: roles of osteopontin. Carcinogenesis 2007, 28, 2476–2483. [Google Scholar] [CrossRef] [PubMed]
- Bekes, E.M.; Schweighofer, B.; Kupriyanova, T.A.; Zajac, E.; Ardi, V.C.; Quigley, J.P.; Deryugina, E.I. Tumor-recruited neutrophils and neutrophil TIMP-free MMP-9 regulate coordinately the levels of tumor angiogenesis and efficiency of malignant cell intravasation. Am. J. Pathol. 2011, 179, 1455–1470. [Google Scholar] [CrossRef] [PubMed]
- Atai, N.A.; Bansal, M.; Lo, C.; Bosman, J.; Tigchelaar, W.; Bosch, K.S.; Jonker, A.; de Witt Hamer, P.C.; Troost, D.; McCulloch, C.A.; et al. Osteopontin is up-regulated and associated with neutrophil and macrophage infiltration in glioblastoma. Immunology 2011, 132, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Aziz, M.; Yang, W.L.; Wang, Z.; Zhou, M.; Ochani, M.; Khader, A.; Wang, P. Neutralization of osteopontin attenuates neutrophil migration in sepsis-induced acute lung injury. Crit. Care 2015, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Urtasun, R.; Lopategi, A.; George, J.; Leung, T.M.; Lu, Y.; Wang, X.; Ge, X.; Fiel, M.I.; Nieto, N. Osteopontin, an oxidant stress sensitive cytokine, up-regulates collagen-I via integrin αvβ3 engagement and PI3K/pAkt/NFκB signaling. Hepatology 2012, 55, 594–608. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, W.; Wang, H.; Deng, Z.; Zeng, C.; Tu, M.; Li, L.; Xiao, W.; Gao, S.; Luo, W.; et al. Osteopontin promotes expression of matrix metalloproteinase 13 through NF-κB signaling in osteoarthritis. BioMed Res. Int. 2016, 2016, 6345656. [Google Scholar] [CrossRef] [PubMed]
- Roeb, E.; Arndt, M.; Jansen, B.; Schumpelick, V.; Matern, S. Simultaneous determination of matrix metalloproteinase (MMP)-7, MMP-1, -3, and -13 gene expression by multiplex PCR in colorectal carcinomas. Int. J. Colorectal Dis. 2004, 19, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Leeman, M.F.; McKay, J.A.; Murray, G.I. Matrix metalloproteinase 13 activity is associated with poor prognosis in colorectal cancer. J. Clin. Pathol. 2002, 55, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Kale, S.; Raja, R.; Thorat, D.; Soundararajan, G.; Patil, T.V.; Kundu, G.C. Osteopontin signaling upregulates cyclooxygenase-2 expression in tumor-associated macrophages leading to enhanced angiogenesis and melanoma growth via α9β1 integrin. Oncogene 2014, 33, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Behera, R.; Kumar, V.; Lohite, K.; Karnik, S.; Kundu, G.C. Activation of JAK2/STAT3 signaling by osteopontin promotes tumor growth in human breast cancer cells. Carcinogenesis 2010, 31, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Nunez, F.; Bravo, S.; Cruzat, F.; Montecino, M.; de Ferrari, G.V. Wnt/β-catenin signaling enhances cyclooxygenase-2 (COX2) transcriptional activity in gastric cancer cells. PLoS ONE 2011, 6, e18562. [Google Scholar] [CrossRef] [PubMed]
- Tetsu, O.; McCormick, F. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [PubMed]
- Takahashi, M.; Kamei, Y.; Ezaki, O. Mest/Peg1 imprinted gene enlarges adipocytes and is a marker of adipocyte size. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E117–E124. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Choi, Y.S.; Choi, J.H. Leptin promotes human endometriotic cell migration and invasion by up-regulating MMP-2 through the JAK2/STAT3 signaling pathway. Mol. Hum. Reprod. 2015, 21, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Avtanski, D.; Saxena, N.K.; Sharma, D. Leptin-induced epithelial-mesenchymal transition in breast cancer cells requires β-catenin activation via Akt/GSK3- and MTA1/Wnt1 protein-dependent pathways. J. Biol. Chem. 2012, 287, 8598–8612. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, R.; Rico, G.; Vega, G.; Basurto, L.; Cordova, L.; Galvan, R.; Hernandez, M.; Puello, E.; Zarate, A. Osteocalcin, under-carboxylated osteocalcin and osteopontin are not associated with gestational diabetes mellitus but are inversely associated with leptin in non-diabetic women. J. Endocrinol. Investig. 2015, 38, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Chang, Y.F. Serum interleukin-6 levels reflect the disease status of colorectal cancer. J. Surg. Oncol. 2003, 83, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Coletta, P.L.; Muller, A.M.; Jones, E.A.; Muhl, B.; Holwell, S.; Clarke, D.; Meade, J.L.; Cook, G.P.; Hawcroft, G.; Ponchel, F.; et al. Lymphodepletion in the ApcMin/+ mouse model of intestinal tumorigenesis. Blood 2004, 103, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Condomines, M.; Veyrune, J.L.; Larroque, M.; Quittet, P.; Latry, P.; Lugagne, C.; Hertogh, C.; Kanouni, T.; Rossi, J.F.; Klein, B. Increased plasma-immune cytokines throughout the high-dose melphalan-induced lymphodepletion in patients with multiple myeloma: a window for adoptive immunotherapy. J. Immunol. 2010, 184, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Noda, M.; Kurahashi, N.; Iwasaki, M.; Sasazuki, S.; Iso, H.; Tsugane, S. Impact of metabolic factors on subsequent cancer risk: results from a large-scale population-based cohort study in Japan. Eur. J. Cancer Prev. 2009, 18, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Iwasaki, M.; Ikeda, S.; Kozu, T.; Saito, H.; Mutoh, M.; Wakabayashi, K.; Tsugane, S. Serum triglycerides and colorectal adenoma in a case-control study among cancer screening examinees. Cancer Causes Control 2006, 17, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Takasu, S.; Mutoh, M.; Takahashi, M.; Nakagama, H. Lipoprotein lipase as a candidate target for cancer prevention/therapy. Biochem. Res. Int. 2012, 2012, 398697. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Wu, D.; Ma, Y.; Zhou, B.; Liu, Q. Suppression of Evi1 promotes the osteogenic differentiation and inhibits the adipogenic differentiation of bone marrow-derived mesenchymal stem cells in vitro. Int. J. Mol. Med. 2015, 36, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, F.J.; Zhao, W.J.; Xing, G.S.; Bai, X.; Wang, Y. Effects of parathyroid hormone-related protein on osteogenic and adipogenic differentiation of human mesenchymal stem cells. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1610–1617. [Google Scholar] [PubMed]
- Qadri, S.M.; Mahmud, H.; Lang, E.; Gu, S.; Bobbala, D.; Zelenak, C.; Jilani, K.; Siegfried, A.; Foller, M.; Lang, F. Enhanced suicidal erythrocyte death in mice carrying a loss-of-function mutation of the adenomatous polyposis coli gene. J. Cell. Mol. Med. 2012, 16, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Booker, C.D.; White, K.L., Jr. Benzo(a)pyrene-induced anemia and splenomegaly in NZB/WF1 mice. Food Chem. Toxicol. 2005, 43, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Youngster, I.; Weiss, M.; Drobot, A.; Eitan, A. An unusual presacral mass: extramedullary hematopoiesis. J. Gastrointest. Surg. 2006, 10, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Johnson, J.N.; Singh, K.; Singh, M. Impairment of myocardial angiogenic response in the absence of osteopontin. Microcirculation 2007, 14, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, P.; Kosunen, T.U.; Valle, J.; Riihela, M.; Seppala, K. Helicobacter pylori infection and chronic gastritis in gastric cancer. J. Clin. Pathol. 1992, 45, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Lee, S.H.; Go du, M.; Kim, H.K.; Kwon, H.J.; Kim, D.Y. Osteopontin depletion decreases inflammation and gastric epithelial proliferation during Helicobacter pylori infection in mice. Lab. Investig. 2015, 95, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Sharan, R.; Schoen, R.E. Cancer in inflammatory bowel disease. An evidence-based analysis and guide for physicians and patients. Gastroenterol. Clin. N. Am. 2002, 31, 237–254. [Google Scholar] [CrossRef]
- Solomon, M.J.; Schnitzler, M. Cancer and inflammatory bowel disease: Bias, epidemiology, surveillance, and treatment. World J. Surg. 1998, 22, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Nakase, H.; Takagi, T.; Bamba, S.; Okuyama, Y.; Kawamura, T.; Oki, T.; Obata, H.; Kawanami, C.; Katsushima, S.; et al. Risk factors for developing colorectal cancer in Japanese patients with ulcerative colitis: a retrospective observational study-CAPITAL (Cohort and Practice for IBD total management in Kyoto-Shiga Links) study I. BMJ Open Gastroenterol. 2016, 3, e000122. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, H.; Shen, Q.; Yuan, S.; Xu, L.; Cai, X.; Lian, J.; Chen, S.Y. Osteopontin: participation in inflammation or mucosal protection in inflammatory bowel diseases? Dig. Dis. Sci. 2013, 58, 1569–1580. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, K.; Hoffmann, U.; Witte, E.; Loddenkemper, C.; Sina, C.; Schreiber, S.; Hayford, C.; Holzlohner, P.; Wolk, K.; Tchatchou, E.; et al. Osteopontin as two-sided mediator of intestinal inflammation. J. Cell. Mol. Med. 2009, 13, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Komine-Aizawa, S.; Masuda, H.; Mazaki, T.; Shiono, M.; Hayakawa, S.; Takayama, T. Plasma osteopontin predicts inflammatory bowel disease activities. Int. Surg. 2015, 100, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Takahashi, Y.; Asai, S.; Hemmi, A.; Takayama, T. Osteopontin expression in ulcerative colitis is distinctly different from that in Crohn’s disease and diverticulitis. J. Gastroenterol. 2005, 40, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Mishima, R.; Takeshima, F.; Sawai, T.; Ohba, K.; Ohnita, K.; Isomoto, H.; Omagari, K.; Mizuta, Y.; Ozono, Y.; Kohno, S. High plasma osteopontin levels in patients with inflammatory bowel disease. J. Clin. Gastroenterol. 2007, 41, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nakai, T.; Tamura, N.; Okamoto, S.; Matsuoka, K.; Sakuraba, A.; Fukushima, T.; Uede, T.; Hibi, T. Osteopontin/Eta-1 upregulated in Crohn’s disease regulates the Th1 immune response. Gut 2005, 54, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.P.; Pollett, A.; Rittling, S.R.; Denhardt, D.T.; Sodek, J.; Zohar, R. Exacerbated tissue destruction in DSS-induced acute colitis of OPN-null mice is associated with downregulation of TNF-α expression and non-programmed cell death. J. Cell. Physiol. 2006, 208, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Eckhardt, E.R.; Oz, H.S.; Bruemmer, D.; de Villiers, W.J. Osteopontin deficiency protects mice from Dextran sodium sulfate-induced colitis. Inflamm. Bowel Dis. 2006, 12, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Liaw, L.; Birk, D.E.; Ballas, C.B.; Whitsitt, J.S.; Davidson, J.M.; Hogan, B.L. Altered wound healing in mice lacking a functional osteopontin gene (spp1). J. Clin. Investig. 1998, 101, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Nagate, T.; Chino, T.; Nishiyama, C.; Okuhara, D.; Tahara, T.; Maruyama, Y.; Kasahara, H.; Takashima, K.; Kobayashi, S.; Motokawa, Y.; et al. Diluted isoflurane as a suitable alternative for diethyl ether for rat anaesthesia in regular toxicology studies. J. Vet. Med. Sci. 2007, 69, 1137–1143. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | No. of Animals | Small Intestinal Polyps | |||
|---|---|---|---|---|---|
| Duodenum | Middle | Distal | Total | ||
| Male | |||||
| Min/OPN(+/+) | 15 | 5.1 ± 3.1 | 40.1 ± 33.9 | 106.6 ± 66.6 | 151.9 ± 101.8 |
| Min/OPN(+/−) | 29 | 5.0 ± 3.7 | 22.8 ± 16.9 | 72.7 ± 37.6 | 100.5 ± 53.4 |
| Min/OPN(−/−) | 18 | 6.0 ± 2.5 | 27.4 ± 21.9 | 77.1 ± 42.9 | 110.5 ± 63.9 |
| Female | |||||
| Min/OPN(+/+) | 10 | 7.1 ± 4.5 | 40.1 ± 27.8 | 106.9 ± 54.4 | 154.1 ± 85.2 |
| Min/OPN(+/−) | 27 | 4.5 ± 2.3 | 22.2 ± 17.5 * | 65.0 ± 44.7 * | 91.7 ± 62.0 * |
| Min/OPN(−/−) | 18 | 5.2 ± 3.3 | 28.2 ± 17.5 | 90.4 ± 44.3 | 123.7 ± 61.9 |
| Total | |||||
| Min/OPN(+/+) | 25 | 5.9 ± 3.8 | 40.1 ± 31.0 | 106.7 ± 60.8 | 152.8 ± 93.6 |
| Min/OPN(+/−) | 56 | 4.8 ± 3.1 | 22.5 ± 17.0 ** | 69.0 ± 41.0 ** | 96.3 ± 57.4 ** |
| Min/OPN(−/−) | 36 | 5.6 ± 2.9 | 27.8 ± 19.5 | 83.8 ± 43.5 | 117.1 ± 62.4 |
| Genotype | Adenoma | Adenocarcinoma | Total | |||
|---|---|---|---|---|---|---|
| Incidence (%) | Multiplicity a | Incidence (%) | Multiplicity a | Incidence (%) | Multiplicity a | |
| Male | ||||||
| Min/OPN(+/+) | 7/15 (47) | 1.0 ± 1.5 | 8/15 (53) | 0.7 ± 0.8 | 12/15 (80) | 1.7 ± 2.0 |
| Min/OPN(+/−) | 4/29 (14) * | 0.1 ± 0.4 ** | 13/29 (45) | 0.6 ± 0.8 | 16/29 (55) | 0.8 ± 0.8 * |
| Min/OPN(−/−) | 4/18 (22) | 0.2 ± 0.4 * | 8/18 (44) | 0.5 ± 0.6 | 11/18 (61) | 0.7 ± 0.7 |
| Female | ||||||
| Min/OPN(+/+) | 4/10 (40) | 0.5 ± 0.7 | 7/10 (70) | 1.0 ± 0.9 | 8/10 (80) | 1.5 ± 1.3 |
| Min/OPN(+/−) | 5/27 (19) | 0.2 ± 0.4 | 8/27 (30) * | 0.3 ± 0.6 * | 11/27 (41) * | 0.6 ± 0.7 * |
| Min/OPN(−/−) | 6/18 (33) | 0.4 ± 0.7 | 5/18 (28) * | 0.3 ± 0.6 * | 7/18 (39) * | 0.8 ± 1.2 |
| Total | ||||||
| Min/OPN(+/+) | 11/25 (44) | 0.8 ± 1.2 | 15/25 (60) | 0.8 ± 0.9 | 20/25 (80) | 1.6 ± 1.7 |
| Min/OPN(+/−) | 9/56 (16) ** | 0.2 ± 0.4 ** | 21/56 (38) | 0.5 ± 0.7 | 27/56 (48) ** | 0.6 ± 0.8 ** |
| Min/OPN(−/−) | 10/36 (28) | 0.3 ± 0.6 * | 13/36 (36) | 0.4 ± 0.6 | 18/36 (50) * | 0.8 ± 0.9 ** |
| Genotype | Serum OPN (ng/mL) | Serum IL-6 (pg/mL) | Serum TGs (mg/dL) |
|---|---|---|---|
| Min/OPN(+/+) | 456.7 ± 144.7 a | 50.5 ± 16.7 a | 464 ± 383 a |
| Min/OPN(+/−) | 250.4 ± 73.4 b | 54.8 ± 27.7 a | 401 ± 454 a |
| Min/OPN(−/−) | 0 d | 22.8 ± 28.3 b | 360 ± 349 a |
| OPN(+/+) | 414.5 ± 192.9 a | 0 b | 94 ± 20 b |
| OPN(+/−) | 182.3 ± 88.2 c | 5.0 ± 14.6 b | 91 ± 25 b |
| OPN(−/−) | 0 d | 0 b | 97 ± 61 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishigamori, R.; Komiya, M.; Takasu, S.; Mutoh, M.; Imai, T.; Takahashi, M. Osteopontin Deficiency Suppresses Intestinal Tumor Development in Apc-Deficient Min Mice. Int. J. Mol. Sci. 2017, 18, 1058. https://doi.org/10.3390/ijms18051058
Ishigamori R, Komiya M, Takasu S, Mutoh M, Imai T, Takahashi M. Osteopontin Deficiency Suppresses Intestinal Tumor Development in Apc-Deficient Min Mice. International Journal of Molecular Sciences. 2017; 18(5):1058. https://doi.org/10.3390/ijms18051058
Chicago/Turabian StyleIshigamori, Rikako, Masami Komiya, Shinji Takasu, Michihiro Mutoh, Toshio Imai, and Mami Takahashi. 2017. "Osteopontin Deficiency Suppresses Intestinal Tumor Development in Apc-Deficient Min Mice" International Journal of Molecular Sciences 18, no. 5: 1058. https://doi.org/10.3390/ijms18051058