
First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis

 , ,
, ,
Abstract
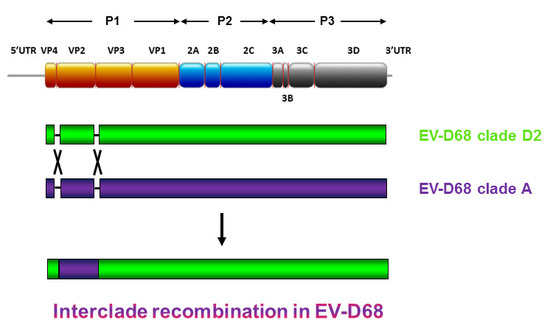
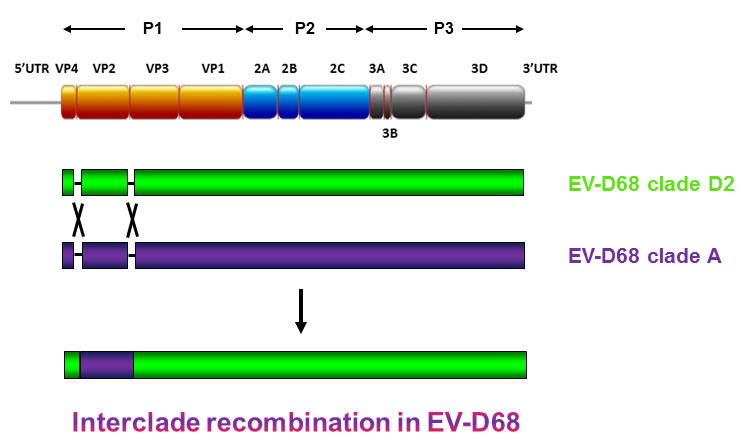
:
1. Introduction
2. Results
2.1. Clinical Characteristics of Patients with Enterovirus D68 (EV-D68) Infections
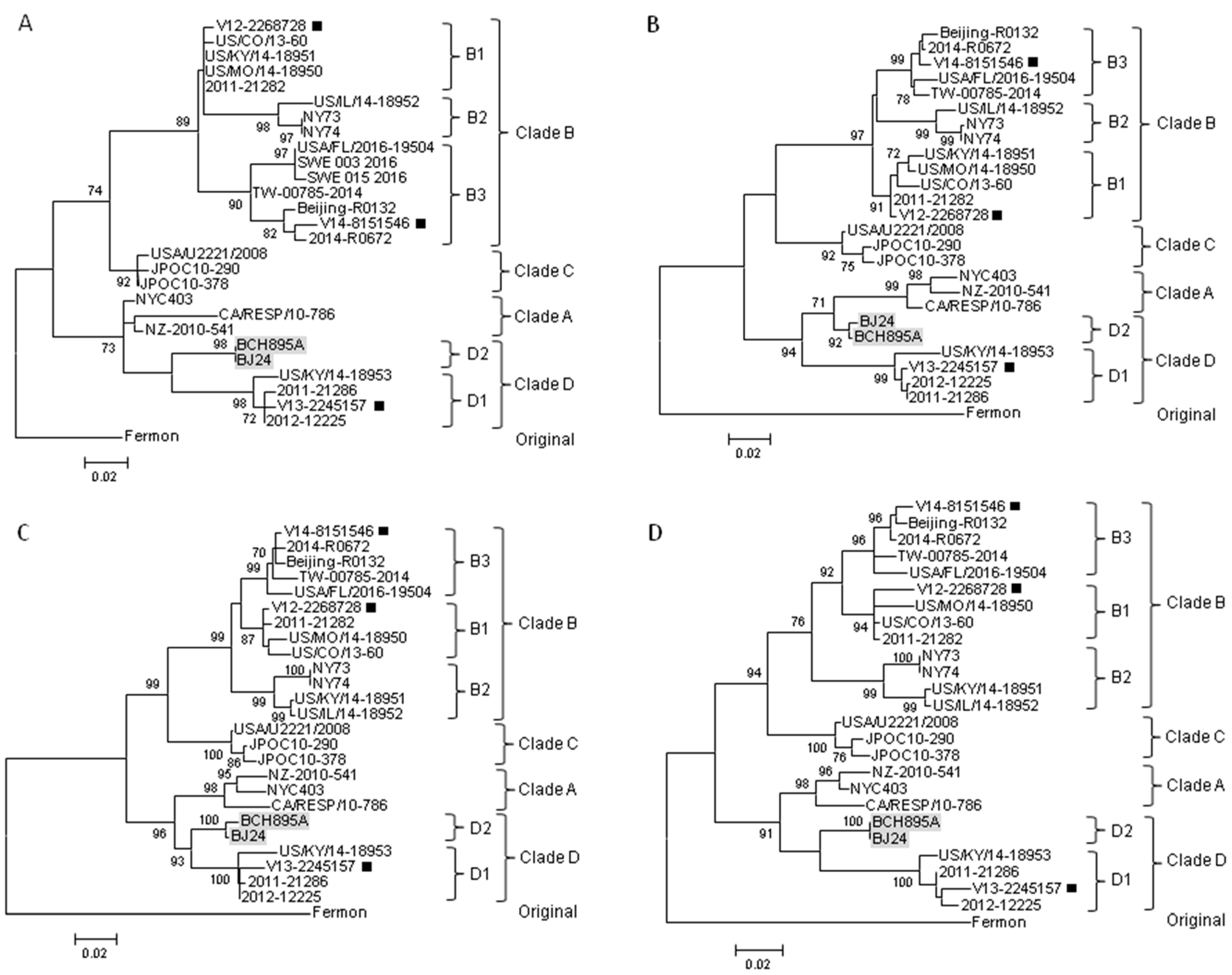
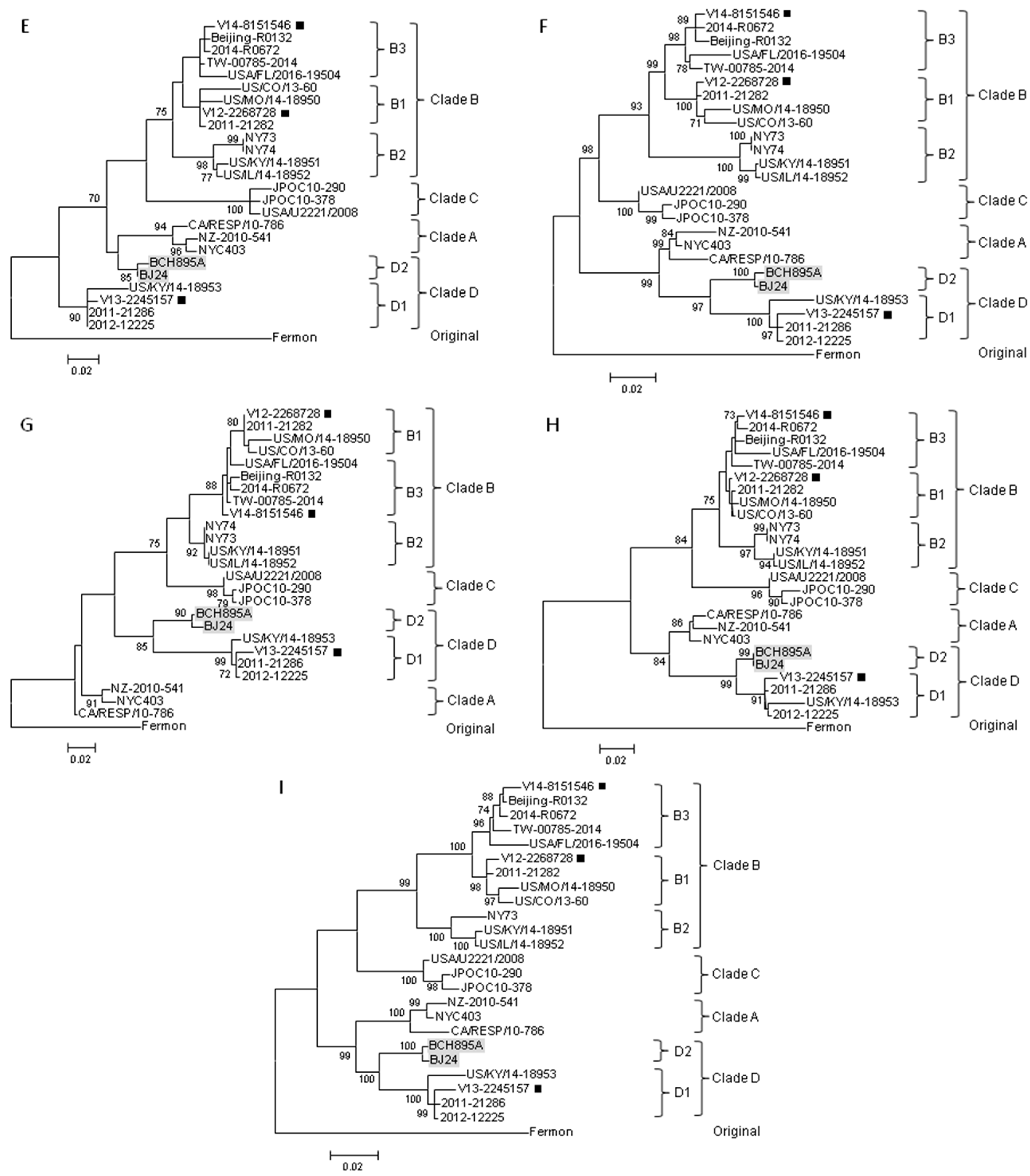
2.2. Complete VP1 Gene Sequence Analysis
2.3. Partial 5′UTR and 2C Sequence Analysis
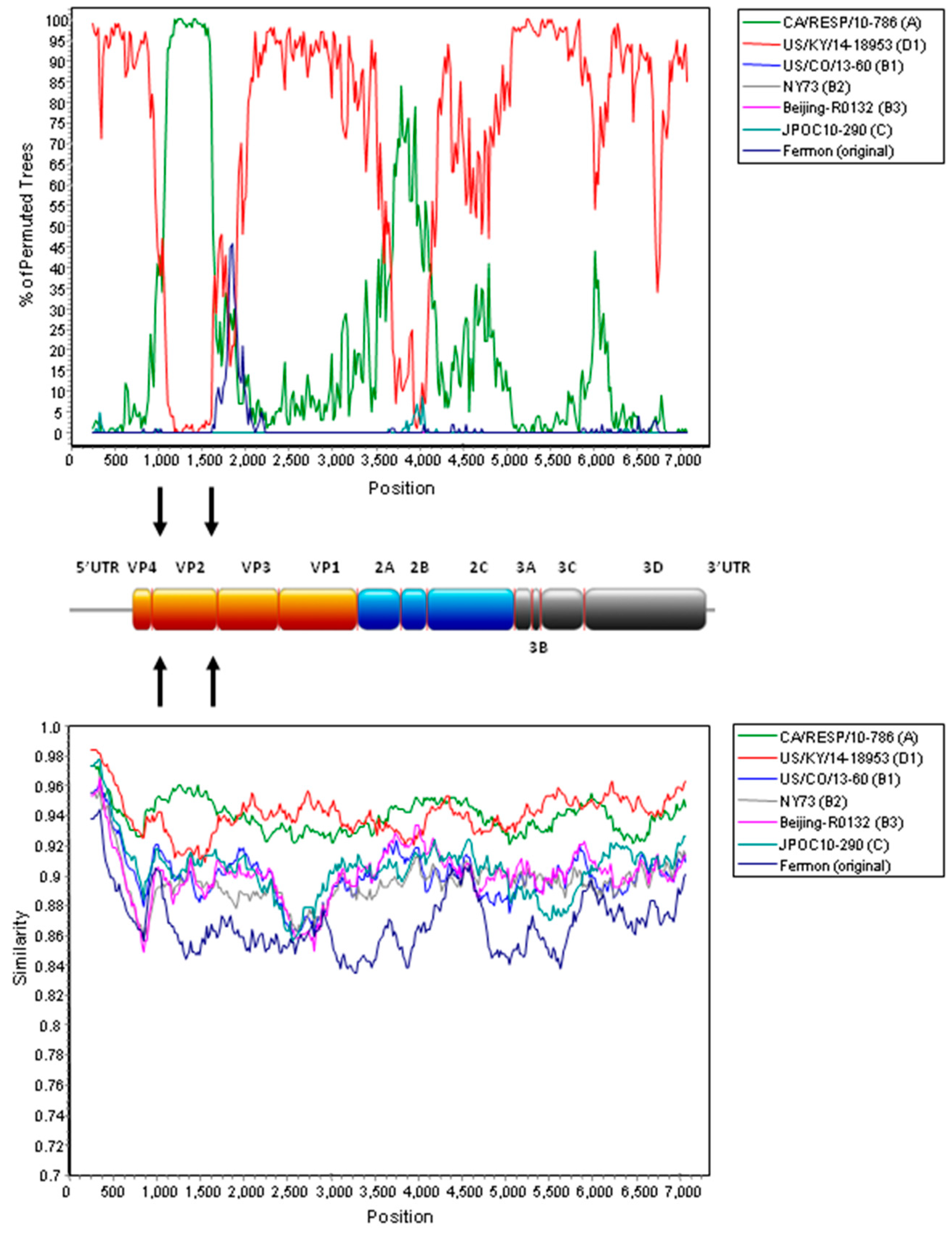
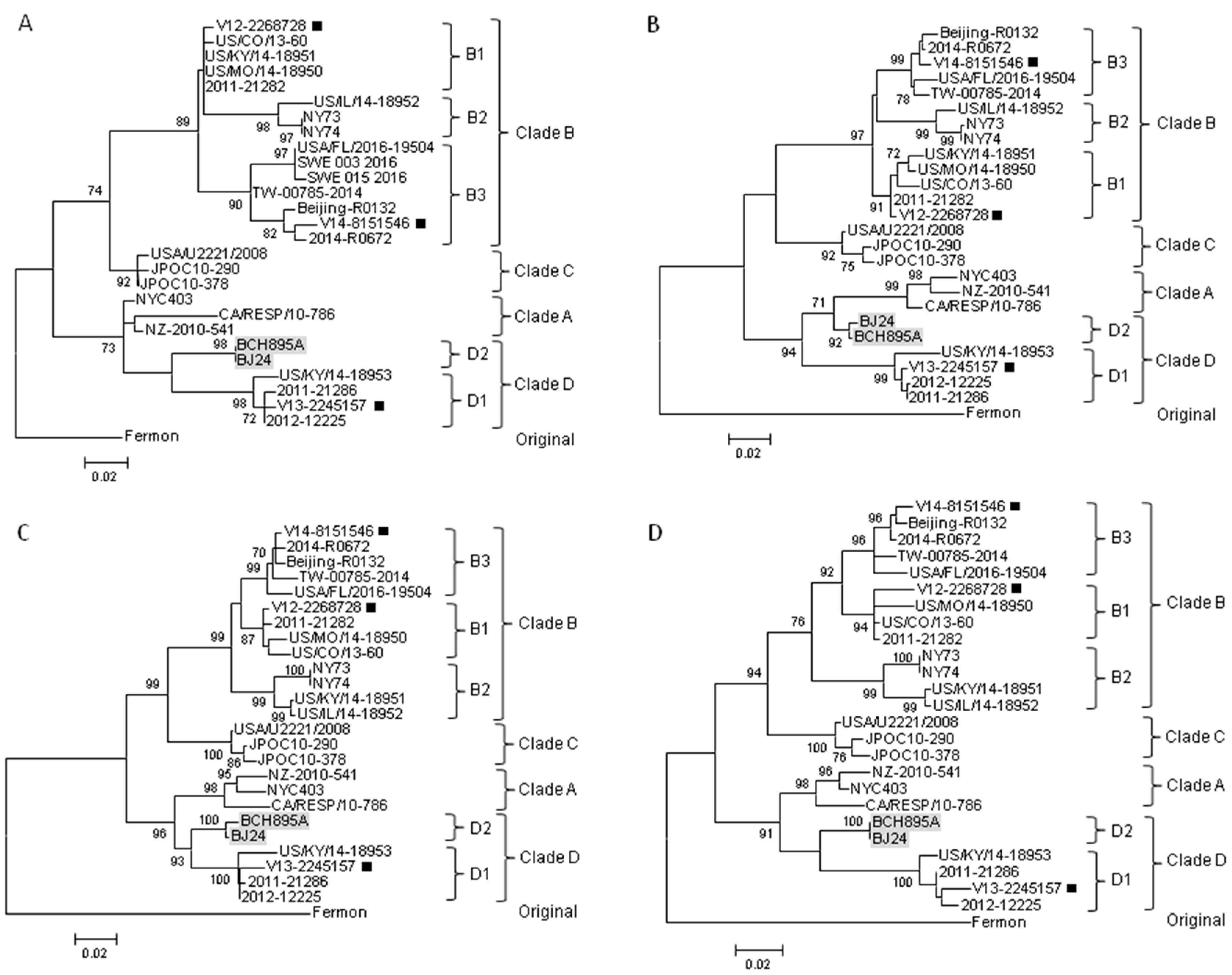
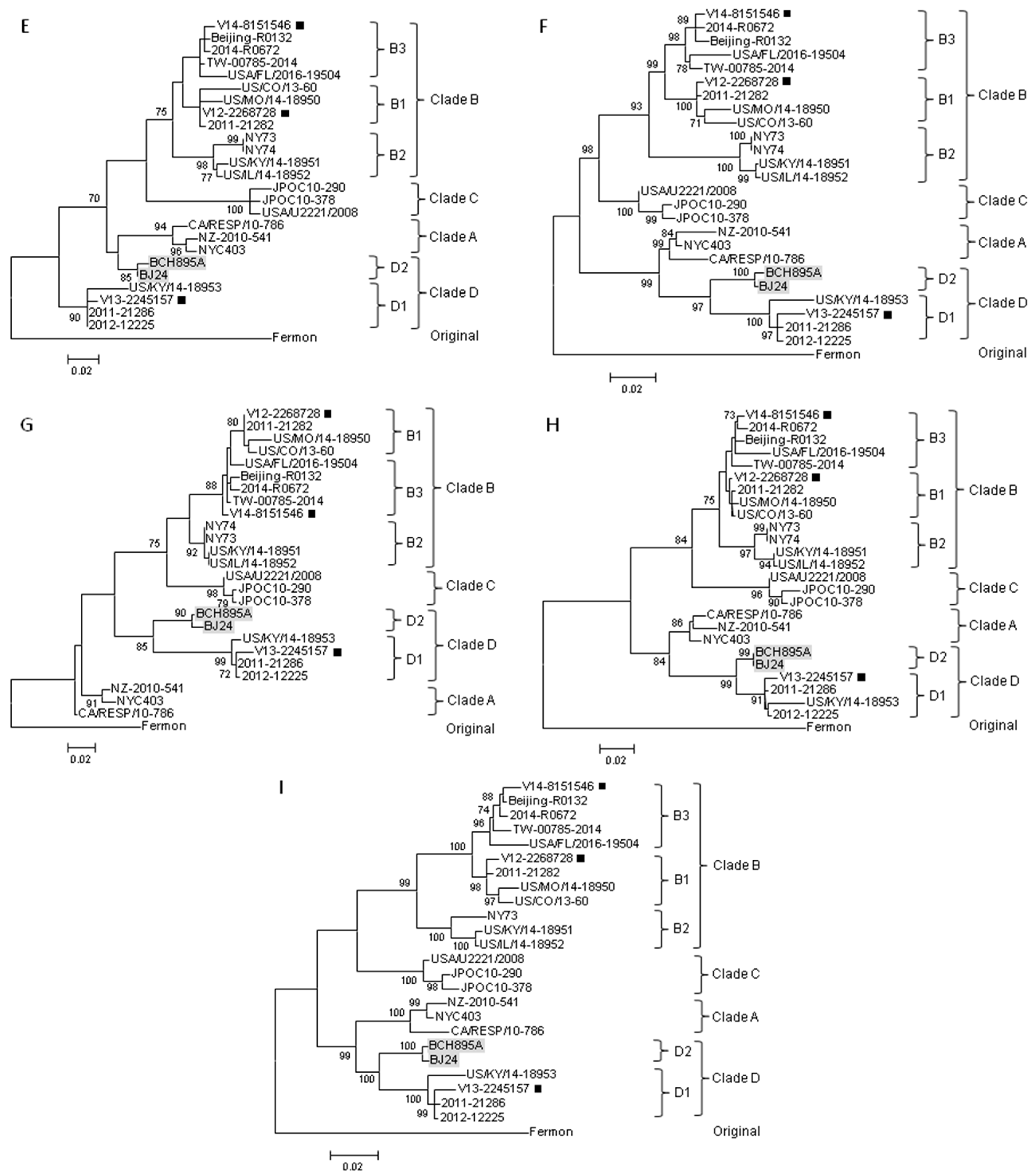
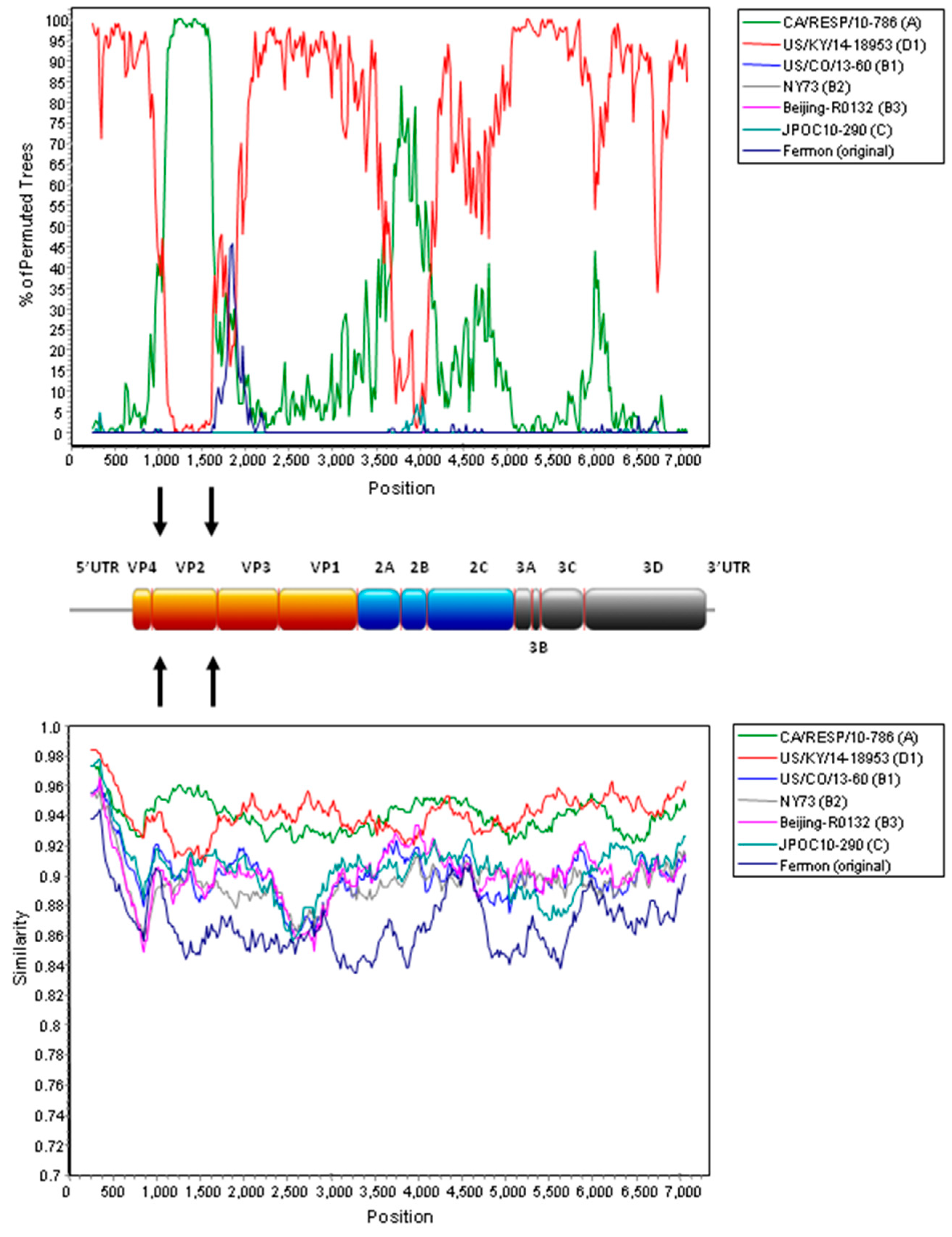
2.4. Complete Genome Analysis of Local and Regional EV-D68 Strains
3. Discussion
4. Materials and Methods
4.1. Clinical Specimens
4.2. RNA Extraction
4.3. Reverse-Transcription Polymerase Chain Reaction (RT-PCR) and Sequencing of VP1 of EV-D68 for Clade Determination
4.4. RT-PCR and Sequencing of Partial 5′UTR and 2C of EV-D68 and Phylogenetic Analysis
4.5. Complete Genome Sequencing of EV-D68
4.6. Genome Analysis
4.7. Nucleotide Sequence Accession Numbers
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AFM | Acute flaccid myelitis |
| CVA | Cerebrovascular accident |
| CSF | Cerebrospinal fluid |
| EV-D | Enterovirus species D |
| HT | Hypertension |
| kbp | Kilobase pairs |
| ML | Maximum likelihood |
| MRMP | Macrolide-resistant Mycoplasma pneumoniae |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| NPA | Nasopharyngeal aspirate |
| NPS | Nasopharyngeal swab |
| RACE | Rapid amplification of cDNA ends |
| RD | Rhabdomyosarcoma |
| RDP | Recombination Detection Program |
| RSV | Respiratory syncytial virus |
| RT-PCR | Reverse transcription polymerase chain reaction |
| URTI | Upper respiratory tract infection |
| US | United States |
| UTR | Untranslated region |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Year of Isolation | Place of Isolation | Clade * | Source | GenBank Accession No. |
|---|---|---|---|---|---|
| Fermon (prototype) | 1962 | USA | Original | GenBank | AY426531 |
| CA/RESP/10-786 | 2013 | USA | A | GenBank | KM892500 |
| NYC403 | 2009 | USA | A | GenBank | JX101846 |
| NZ-2010-541 | 2010 | NZ | A | GenBank | JX070222 |
| HK2/2010 | 2010 | HK | A | GenBank | KT959174 (VP1), |
| KT959114 (2C) | |||||
| TW-02378_2010 | 2010 | TW | A | GenBank | KP657713 (VP1) |
| ITA/19391/12 | 2012 | Italy | A | GenBank | KC763159 (VP1) |
| 2011-21282 | 2011 | China | B1 | GenBank | KT285320 |
| HK8/2011 | 2011 | HK | B1 | GenBank | KT959180 (VP1), |
| KT959120 (2C) | |||||
| HK14/2012 | 2012 | HK | B1 | GenBank | KT959186 (VP1) |
| KT959126 (2C) | |||||
| HK26/2014 | 2014 | HK | B1 | GenBank | KT959198 (VP1), |
| KT959138 (2C) | |||||
| NY316 | 2014 | USA | B1 | GenBank | KP745764 |
| STL_2014_12 | 2014 | USA | B1 | GenBank | KM881710 |
| US/CO/13-60 | 2013 | USA | B1 | GenBank | KP100794 |
| US/CO/14-93 | 2014 | USA | B1 | GenBank | KP126911 |
| US/MO/14-18950 | 2014 | USA | B1 | GenBank | KM851228 |
| NY73 | 2014 | USA | B2 | GenBank | KP745768 |
| NY74 | 2014 | USA | B2 | GenBank | KP745769 |
| US/KY/14-18951 | 2014 | USA | B2 | GenBank | KM851229 |
| US/IL/14-18952 | 2014 | USA | B2 | GenBank | KM851230 |
| 2014-R0672 | 2014 | China | B3 | GenBank | KT280500 |
| Beijing-R0132 | 2014 | China | B3 | GenBank | KP240936 |
| HK25/2014 | 2014 | HK | B3 | GenBank | KT959197 (VP1), |
| KT959137 (2C) | |||||
| HK28/2014 | 2014 | HK | B3 | GenBank | KT959200 (VP1), |
| KT959140 (2C) | |||||
| TW-00932-2014 | 2014 | TW | B3 | GenBank | KT711081 |
| TW-00785-2014 | 2014 | TW | B3 | GenBank | KT711083 |
| NY10_16 | 2016 | USA | B3 | GenBank | KX957754 |
| USA/FL/2016-19504 | 2016 | USA | B3 | GenBank | KX675261 |
| SWE_003_2016 | 2016 | SWE | B3 | GenBank | KY215829 (VP4) |
| SWE_015_2016 | 2016 | SWE | B3 | GenBank | KY215841 (VP4) |
| USA/U2221/2008 | 2008 | USA | C | GenBank | KX255371 |
| USA/MO93/2009 | 2009 | USA | C | GenBank | KX261814 |
| JPOC10-290 | 2010 | Japan | C | GenBank | AB601882 |
| JPOC10-378 | 2010 | Japan | C | GenBank | AB601883 |
| 2011-21286 | 2011 | China | D1 | GenBank | KT306743 |
| 2012-12225 | 2012 | China | D1 | GenBank | KT285319 |
| 2013-1017-26 | 2013 | China | D1 | GenBank | KT280501 |
| HK5/2011 | 2011 | HK | D1 | GenBank | KT959177 (VP1), |
| KT959117 (2C) | |||||
| HK13/2012 | 2012 | HK | D1 | GenBank | KT959185 (VP1), |
| KT959125 (2C) | |||||
| HK21/2013 | 2013 | HK | D1 | GenBank | KT959193 (VP1), |
| KT959133 (2C) | |||||
| US/KY/14-18953 | 2014 | USA | D1 | GenBank | KM851231 |
| TW-02722_2013 | 2013 | TW | D1 | GenBank | KP657720 (VP1) |
| BCH895A | 2008 | China | D2 | GenBank | KF726085 |
| BJ24 | 2008 | China | D2 | GenBank | KU242683 |
| 1737-Yamagata-2008 | 2008 | Japan | D2 | GenBank | AB667899 (VP1) |
| TW-01166_2007 | 2007 | TW | D2 | GenBank | KP657701 (VP1) |
| TW-11134_2009 | 2009 | TW | D2 | GenBank | KP657708 (VP1) |
| V12-2240471 | 2012 | HK | B1 | This study | KY767823 (VP1), |
| KY767829 (5′UTR), | |||||
| KY767836 (2C) | |||||
| V12-2268728 | 2012 | HK | B1 | This study | KY767820 |
| V13-2245157 | 2013 | HK | D1 | This study | KY767821 |
| V14-8133616 | 2014 | HK | B3 | This study | KY767824 (VP1), |
| KY767830 (5′UTR), | |||||
| KY767837 (2C) | |||||
| V14-8135150 | 2014 | HK | B3 | This study | KY767825 (VP1), |
| KY767831 (5′UTR), | |||||
| KY767838 (2C) | |||||
| V14-8143594 | 2014 | HK | B3 | This study | KY767826 (VP1), |
| KY767832 (5′UTR), | |||||
| KY767839 (2C) | |||||
| V14-8143833 | 2014 | HK | B3 | This study | KY767827 (VP1), |
| KY767833 (5′UTR), | |||||
| KY767840 (2C) | |||||
| V14-8151518 | 2014 | HK | B3 | This study | KY767828 (VP1), |
| KY767834 (5′UTR), | |||||
| KY767841 (2C) | |||||
| V14-8151546 | 2014 | HK | B3 | This study | KY767822 |
| V14-8157864 | 2014 | HK | B3 # | This study | KY767835 (5′UTR), |
| KY767842 (2C) |
References
- Schieble, J.H.; Fox, V.L.; Lennette, E.H. A probable new human picornavirus associated with respiratory diseases. Am. J. Epidemiol. 1967, 85, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, S.; Savolainen, C.; Raman, L.; Roivainen, M.; Hovi, T. Human rhinovirus 87 and enterovirus 68 represent a unique serotype with rhinovirus and enterovirus features. J. Clin. Microbiol. 2002, 40, 4218–4223. [Google Scholar] [CrossRef] [PubMed]
- Khetsuriani, N.; Lamonte-Fowlkes, A.; Oberst, S.; Pallansch, M.A. Centers for Disease Control and Prevention. Enterovirus surveillance—United States, 1970–2005. MMWR Surveill. Summ. 2006, 55, 1–20. [Google Scholar] [PubMed]
- Ikeda, T.; Mizuta, K.; Abiko, C.; Aoki, Y.; Itagaki, T.; Katsushima, F.; Katsushima, Y.; Matsuzaki, Y.; Fuji, N.; Imamura, T.; et al. Acute respiratory infections due to enterovirus 68 in Yamagata, Japan between 2005 and 2010. Microbiol. Immunol. 2012, 56, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Fuji, N.; Suzuki, A.; Tamaki, R.; Saito, M.; Aniceto, R.; Galang, H.; Sombrero, L.; Lupisan, S.; Oshitani, H. Enterovirus 68 among children with severe acute respiratory infection, the Philippines. Emerg. Infect. Dis. 2011, 17, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Linsuwanon, P.; Puenpa, J.; Suwannakarn, K.; Auksornkitti, V.; Vichiwattana, P.; Korkong, S.; Theamboonlers, A.; Poovorawan, Y. Molecular epidemiology and evolution of human enterovirus serotype 68 in Thailand, 2006–2011. PLoS ONE 2012, 7, e35190. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.B.; Wo, Y.; Wang, H.Y.; Wei, M.T.; Zhang, L.; Yang, H.; Liu, E.M.; Li, T.Y.; Zhao, Z.T.; Liu, W.; et al. Detection of enterovirus 68 as one of the commonest types of enterovirus found in patients with acute respiratory tract infection in China. J. Med. Microbiol. 2014, 63, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.; van der Sanden, S.; Snijders, B.E.; Jaramillo-Gutierrez, G.; Bont, L.; van der Ent, C.K.; Overduin, P.; Jenny, S.L.; Jusic, E.; van der Avoort, H.G.; et al. Emergence and epidemic occurrence of enterovirus 68 respiratory infections in The Netherlands in 2010. Virology 2012, 423, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Opanda, S.M.; Wamunyokoli, F.; Khamadi, S.; Coldren, R.; Bulimo, W.D. Genetic diversity of human enterovirus 68 strains isolated in Kenya using the hypervariable 3′ end of VP1 gene. PLoS ONE 2014, 9, e102866. [Google Scholar] [CrossRef] [PubMed]
- Piralla, A.; Girello, A.; Grignani, M.; Gozalo-Marguello, M.; Marchi, A.; Marseglia, G.; Baldanti, F. Phylogenetic characterization of enterovirus 68 strains in patients with respiratory syndromes in Italy. J. Med. Virol. 2014, 86, 1590–1593. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Firth, C.; Madhi, S.A.; Howie, S.R.; Wu, W.; Sall, A.A.; Haq, S.; Briese, T.; Lipkin, W.I. Worldwide emergence of multiple clades of enterovirus 68. J. Gen. Virol. 2012, 93, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Enterovirus D68. Available online: https://www.cdc.gov/non-polio-enterovirus/about/ev-d68.html (accessed on 13 March 2017).
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012–14): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Lau, S.K.; Yip, C.C.; Zhao, P.S.; Chow, W.N.; To, K.K.; Wu, A.K.; Yuen, K.Y.; Woo, P.C. Enterovirus D68 infections associated with severe respiratory illness in elderly patients and emergence of a novel clade in Hong Kong. Sci. Rep. 2016, 6, 25147. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.W. Rates of spontaneous mutation among RNA viruses. Proc. Natl. Acad. Sci. USA 1993, 90, 4171–4175. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, T.C.; Kirkegaard, K. Poliovirus RNA recombination: Mechanistic studies in the absence of selection. EMBO J. 1992, 11, 3135–3145. [Google Scholar] [PubMed]
- Lukashev, A.N.; Lashkevich, V.A.; Ivanova, O.E.; Koroleva, G.A.; Hinkkanen, A.E.; Ilonen, J. Recombination in circulating enteroviruses. J. Virol. 2003, 77, 10423–10431. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.C.; Lau, S.K.; Lo, J.Y.; Chan, K.H.; Woo, P.C.; Yuen, K.Y. Genetic characterization of EV71 isolates from 2004 to 2010 reveals predominance and persistent circulation of the newly proposed genotype D and recent emergence of a distinct lineage of subgenotype C2 in Hong Kong. Virol. J. 2013, 10, 222. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Hassan, F.; Schuster, J.E.; Simenauer, A.; Selvarangan, R.; Halpin, R.A.; Lin, X.; Fedorova, N.; Stockwell, T.B.; Lam, T.T.; et al. Molecular Evolution and Intraclade Recombination of Enterovirus D68 during the 2014 Outbreak in the United States. J. Virol. 2015, 90, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, X.; Zhang, H.; Xu, X.; Li, W.; Zhu, D.; Wang, M. Antimicrobial susceptibility of Mycoplasma pneumoniae isolates and molecular analysis of macrolide-resistant strains from Shanghai, China. Antimicrob. Agents Chemother. 2009, 53, 2160–2162. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zheng, B.; Zheng, W.; Li, P.; Kang, J.; Hou, J.; Markham, R.; Zhao, K.; Yu, X.F. Analysis of Enterovirus 68 strains from the 2014 north american outbreak reveals a new clade, indicating viral evolution. PLoS ONE 2015, 10, e0144208. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.N.; Yang, S.L.; Shih, S.R.; Huang, Y.C.; Chang, P.Y.; Huang, C.G.; Kao, K.C.; Hu, H.C.; Liu, Y.C.; Tsao, K.C. Molecular evolution and the global reemergence of enterovirus D68 by genome-wide analysis. Medicine (Baltimore) 2016, 95, e4416. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Wong, S.; Drews, S.J.; Tipples, G.; Tellier, R. Full genome analysis of enterovirus D-68 strains circulating in Alberta, Canada. J. Med. Virol. 2016, 88, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, G.; Zhuge, J.; Nolan, S.M.; Dimitrova, N.; Fallon, J.T. Whole-genome sequence analysis reveals the enterovirus D68 isolates during the United States 2014 outbreak mainly belong to a novel clade. Sci. Rep. 2015, 5, 15223. [Google Scholar] [CrossRef] [PubMed]
- Aliabadi, N.; Messacar, K.; Pastula, D.M.; Robinson, C.C.; Leshem, E.; Sejvar, J.J.; Nix, W.A.; Oberste, M.S.; Feikin, D.R.; Dominguez, S.R. Enterovirus D68 infection in children with acute flaccid myelitis, Colorado, USA, 2014. Emerg. Infect. Dis. 2016, 22, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Schreiner, T.L.; van Haren, K.; Yang, M.; Glaser, C.A.; Tyler, K.L.; Dominguez, S.R. Acute flaccid myelitis: A clinical review of US cases 2012–2015. Ann. Neurol. 2016, 80, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J.; Lopez, A.S.; Cortese, M.M.; Leshem, E.; Pastula, D.M.; Miller, L.; Glaser, C.; Kambhampati, A.; Shioda, K.; Aliabadi, N.; et al. Acute flaccid myelitis in the United States, August–December 2014: Results of nationwide surveillance. Clin. Infect. Dis. 2016, 63, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Grard, G.; Drexler, J.F.; Lekana-Douki, S.; Caron, M.; Lukashev, A.; Nkoghe, D.; Gonzalez, J.P.; Drosten, C.; Leroy, E. Type 1 wild poliovirus and putative enterovirus 109 in an outbreak of acute flaccid paralysis in Congo, October–November 2010. Euro Surveill. 2010, 15, pii:19723. [Google Scholar]
- Ooi, M.H.; Wong, S.C.; Lewthwaite, P.; Cardosa, M.J.; Solomon, T. Clinical features, diagnosis, and management of enterovirus 71. Lancet Neurol. 2010, 9, 1097–1105. [Google Scholar] [CrossRef]
- Perez-Velez, C.M.; Anderson, M.S.; Robinson, C.C.; McFarland, E.J.; Nix, W.A.; Pallansch, M.A.; Oberste, M.S.; Glodé, M.P. Outbreak of neurologic enterovirus type 71 disease: A diagnostic challenge. Clin. Infect. Dis. 2007, 45, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Teoh, H.L.; Mohammad, S.S.; Britton, P.N.; Kandula, T.; Lorentzos, M.S.; Booy, R.; Jones, C.A.; Rawlinson, W.; Ramachandran, V.; Rodriguez, M.L.; et al. Clinical characteristics and functional motor outcomes of enterovirus 71 neurological disease in children. JAMA Neurol. 2016, 73, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, J.D.; Barnes, A.; McCarthy, J.E.; Schwartzman, J.D.; Oberste, M.S.; Rhodes, C.H.; Modlin, J.F.; Wright, P.F. A fatal central nervous system enterovirus 68 infection. Arch. Pathol. Lab. Med. 2011, 135, 793–796. [Google Scholar] [PubMed]
- Levy, A.; Roberts, J.; Lang, J.; Tempone, S.; Kesson, A.; Dofai, A.; Daley, A.J.; Thorley, B.; Speers, D.J. Enterovirus D68 disease and molecular epidemiology in Australia. J. Clin. Virol. 2015, 69, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Hixon, A.M.; Yu, G.; Leser, J.S.; Yagi, S.; Clarke, P.; Chiu, C.Y.; Tyler, K.L. A mouse model of paralytic myelitis caused by enterovirus D68. PLoS Pathog. 2017, 13, e1006199. [Google Scholar] [CrossRef] [PubMed]
- Dyrdak, R.; Grabbe, M.; Hammas, B.; Ekwall, J.; Hansson, K.E.; Luthander, J.; Naucler, P.; Reinius, H.; Rotzén-Östlund, M.; Albert, J. Outbreak of enterovirus D68 of the new B3 lineage in Stockholm, Sweden, August to September 2016. Euro Surveill. 2016, 21, pii:30403. [Google Scholar] [CrossRef] [PubMed]
- Knoester, M.; Schölvinck, E.H.; Poelman, R.; Smit, S.; Vermont, C.L.; Niesters, H.G.; van Leer-Buter, C.C. Upsurge of enterovirus D68, the Netherlands, 2016. Emerg. Infect. Dis. 2017, 23, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Chidini, G.; Cinnante, C.; Napolitano, L.; Giannini, A.; Terranova, L.; Niesters, H.; Principi, N.; Calderini, E. Acute flaccid myelitis associated with enterovirus-D68 infection in an otherwise healthy child. Virol. J. 2017, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.F.; Sam, I.C.; AbuBakar, S. Phylogenetic designation of enterovirus 71 genotypes and subgenotypes using complete genome sequences. Infect. Genet. Evol. 2010, 10, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Oberste, M.S.; Maher, K.; Kilpatrick, D.R.; Flemister, M.R.; Brown, B.A.; Pallansch, M.A. Typing of human enteroviruses by partial sequencing of VP1. J. Clin. Microbiol. 1999, 37, 1288–1293. [Google Scholar] [PubMed]
- Oberste, M.S.; Maher, K.; Kilpatrick, D.R.; Pallansch, M.A. Molecular evolution of the human enteroviruses: Correlation of serotype with VP1 sequence and application to picornavirus classification. J. Virol. 1999, 73, 1941–1948. [Google Scholar] [PubMed]
- Brown, B.A.; Nix, W.A.; Sheth, M.; Frace, M.; Oberste, M.S. Seven strains of enterovirus D68 Detected in the United States during the 2014 severe respiratory disease outbreak. Genome Announc. 2014, 2, pii:e01201-14. [Google Scholar] [CrossRef] [PubMed]
- Andersson, P.; Edman, K.; Lindberg, A.M. Molecular analysis of the echovirus 18 prototype: Evidence of interserotypic recombination with echovirus 9. Virus Res. 2002, 85, 71–83. [Google Scholar] [CrossRef]
- Lindberg, A.M.; Andersson, P.; Savolainen, C.; Mulders, M.N.; Hovi, T. Evolution of the genome of Human enterovirus B: Incongruence between phylogenies of the VP1 and 3CD regions indicates frequent recombination within the species. J. Gen. Virol. 2003, 84, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Lukashev, A.N.; Lashkevich, V.A.; Koroleva, G.A.; Ilonen, J.; Hinkkanen, A.E. Recombination in uveitis-causing enterovirus strains. J. Gen. Virol. 2004, 85, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, S.; Savolainen-Kopra, C.; Paananen, A.; El Bassioni, L.; El Maamoon Nasr, E.M.; Firstova, L.; Zamiatina, N.; Kutateladze, T.; Roivainen, M. Recurrent isolation of poliovirus 3 strains with chimeric capsid protein Vp1 suggests a recombination hot-spot site in Vp1. Virus Res. 2010, 151, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; Samoilovich, E.; Dunn, G.; Lackenby, A.; Feldman, E.; Heath, A.; Svirchevskaya, E.; Cooper, G.; Yermalovich, M.; Minor, P.D. Isolation of an intertypic poliovirus capsid recombinant from a child with vaccine-associated paralytic poliomyelitis. J. Virol. 2002, 76, 10921–10928. [Google Scholar] [CrossRef] [PubMed]
- Phuektes, P.; Chua, B.H.; Sanders, S.; Bek, E.J.; Kok, C.C.; McMinn, P.C. Mapping genetic determinants of the cell-culture growth phenotype of enterovirus 71. J. Gen. Virol. 2011, 92, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Holloway, B.; Dare, R.K.; Kuypers, J.; Yagi, S.; Williams, J.V.; Hall, C.B.; Erdman, D.D. Real-time reverse transcription-PCR assay for comprehensive detection of human rhinoviruses. J. Clin. Microbiol. 2008, 46, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Campanella, J.J.; Bitincka, L.; Smalley, J. MatGAT: An application that generates similarity/identity matrices using protein or DNA sequences. BMC Bioinform. 2003, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.C.; Lau, S.K.; Woo, P.C.; Wong, S.S.; Tsang, T.H.; Lo, J.Y.; Lam, W.K.; Tsang, C.C.; Chan, K.H.; Yuen, K.Y. Recombinant coxsackievirus A2 and deaths of children, Hong Kong, 2012. Emerg. Infect. Dis. 2013, 19, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]


| Case | Strain | Specimen Type | Collection Date | Gender | Age | Underlying Diseases | Diagnosis | Clade |
|---|---|---|---|---|---|---|---|---|
| 1 | V14-8157864 | NPA | 16 August 2014 | M | 10 | α-thalassemia trait | Encephalitis, Mycoplasma pneumoniae co-infection | B3 # |
| 2 | V12-2240471 | NPS | 7 May 2012 | M | 5 | Allergic rhinitis | URTI, asthma | B1 |
| 3 | V12-2268728 * | NPA | 20 July 2012 | M | 2 | None | URTI, asthma | B1 |
| 4 | V13-2245157 * | NPS | 21 May 2013 | F | 94 | HT, CVA, gout, dementia | Pneumonia | D1 |
| 5 | V14-8133616 | NPA | 18 May 2014 | M | 4 | Allergic rhinitis, asthma | URTI, asthma | B3 |
| 6 | V14-8135150 | NPA | 23 May 2014 | F | 4 | MRSA skin infection | URTI, RSV co-infection | B3 |
| 7 | V14-8143594 | NPS | 21 June 2014 | F | 4 | Eczema | Wheezy bronchitis | B3 |
| 8 | V14-8143833 | NPS | 24 June 2014 | F | 4 | None | URTI, asthma | B3 |
| 9 | V14-8151518 | NPS | 23 July 2014 | M | 5 | α-thalassemia trait | URTI, Henoch Schonlein purpura | B3 |
| 10 | V14-8151546 * | NPS | 23 July 2014 | M | 5 | None | Febrile wheeze | B3 |
| Strains * | Pairwise Identity (%) | Average Nucleotide Divergence (%) [Range] |
|---|---|---|
| Interclade | 10.8 [6.5–14.8] | |
| Prototype vs. A | 87–87.9 | |
| Prototype vs. B | 85.5–86.6 | |
| Prototype vs. C | 87.8–88 | |
| Prototype vs. D | 85.2–86.7 | |
| A vs. B | 86.5–90.4 | |
| A vs. C | 88.8–91.4 | |
| A vs. D | 90.2–93.5 | |
| B vs. C | 89.8–93.1 | |
| B vs. D | 86.4–89.7 | |
| C vs. D | 87.9–90.5 | |
| A vs. D1 | 90.2–91.7 | |
| A vs. D2 | 91.8–93.5 | |
| Intersubclade | 4.7 [2.6–7.1] | |
| B1 vs. B2 | 94–95.2 | |
| B1 vs. B3 | 94.7–97.4 | |
| B2 vs. B3 | 92.9–94.2 | |
| D1 vs. D2 | 94.1–95.6 | |
| Intrasubclade | 1.4 [0–3.5] | |
| B1 | 96.5–99.8 | |
| B2 | 98.9–100 | |
| B3 | 97.3–99.9 | |
| D1 | 97.5–99.7 | |
| D2 | 98.1–99.6 |
| Strains | Positions | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 6 | 92 | 103 | 131 | 140 | 142 | 147 | 177 | 186 | 279 | C-terminus | |
| Clade A | |||||||||||||
| CA/RESP/10-786 | D | D | A | A | F | V | S | S | T | A | I | R | VTT |
| NZ-2010-541 | D | D | A | A | F | V | S | S | T | A | I | R | VTT |
| NYC403 | D | D | A | A | F | V | S | S | T | A | I | R | VTT |
| TW-02378_2010 | D | D | A | A | F | V | S | S | T | A | I | R | |
| ITA/19391/12 | D | H | A | A | F | V | S | S | T | A | I | R | |
| HK2/2010 | D | D | A | A | F | V | S | S | T | A | I | R | VTT |
| Clade D | |||||||||||||
| Subclade D1: | |||||||||||||
| TW-02722_2013 | E | T | I | G | N | M | G | V | RLVNT | ||||
| US/KY/14-18953 | E | T | I | G | N | M | G | V | RLVNT | ||||
| 2011-21286 | E | T | I | G | N | M | G | V | RLVNT | ||||
| 2012-12225 | E | T | I | G | N | M | G | V | RLVNT | ||||
| 2013-1017-26 | E | T | I | G | N | M | G | V | RLVNT | ||||
| HK5/2011 | E | T | I | G | N | M | G | V | RLVNT | ||||
| HK13/2012 | E | T | I | G | N | M | G | V | RLVNT | ||||
| HK21/2013 | E | T | I | G | N | M | G | V | RLVNT | ||||
| V13-2245157 | E | T | I | G | N | M | G | V | RLVNT | ||||
| Subclade D2: | |||||||||||||
| BCH895A | E | D | A | T | F | I | G | N | M | G | V | R | RLVNT |
| BJ24 | E | D | A | T | F | I | G | N | M | G | V | R | RLVNT |
| TW-01166_2007 | E | D | A | T | F | I | G | N | M | G | V | R | |
| TW-11134_2009 | E | D | A | T | F | I | G | N | M | G | V | R | |
| 1737-Yamagata-2008 | E | D | A | T | F | I | G | N | M | G | V | R | RLVNT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yip, C.C.Y.; Lo, J.Y.C.; Sridhar, S.; Lung, D.C.; Luk, S.; Chan, K.-H.; Chan, J.F.W.; Cheng, V.C.C.; Woo, P.C.Y.; Yuen, K.-Y.; et al. First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis. Int. J. Mol. Sci. 2017, 18, 1065. https://doi.org/10.3390/ijms18051065
Yip CCY, Lo JYC, Sridhar S, Lung DC, Luk S, Chan K-H, Chan JFW, Cheng VCC, Woo PCY, Yuen K-Y, et al. First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis. International Journal of Molecular Sciences. 2017; 18(5):1065. https://doi.org/10.3390/ijms18051065
Chicago/Turabian StyleYip, Cyril C. Y., Janice Y. C. Lo, Siddharth Sridhar, David C. Lung, Shik Luk, Kwok-Hung Chan, Jasper F. W. Chan, Vincent C. C. Cheng, Patrick C. Y. Woo, Kwok-Yung Yuen, and et al. 2017. "First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis" International Journal of Molecular Sciences 18, no. 5: 1065. https://doi.org/10.3390/ijms18051065