
Estrogen Modulates Specific Life and Death Signals Induced by LH and hCG in Human Primary Granulosa Cells In Vitro

 ,
,
Abstract
:1. Introduction
1.1. Gonadotropin Receptors-Mediated Signals
1.2. Differences between Luteinizing Hormone (LH) and Human Chorionic Gonadotropin (hCG) at the Molecular Levels
1.3. Role of Estrogens in the Ovary
1.4. Aim of the Study
2. Results
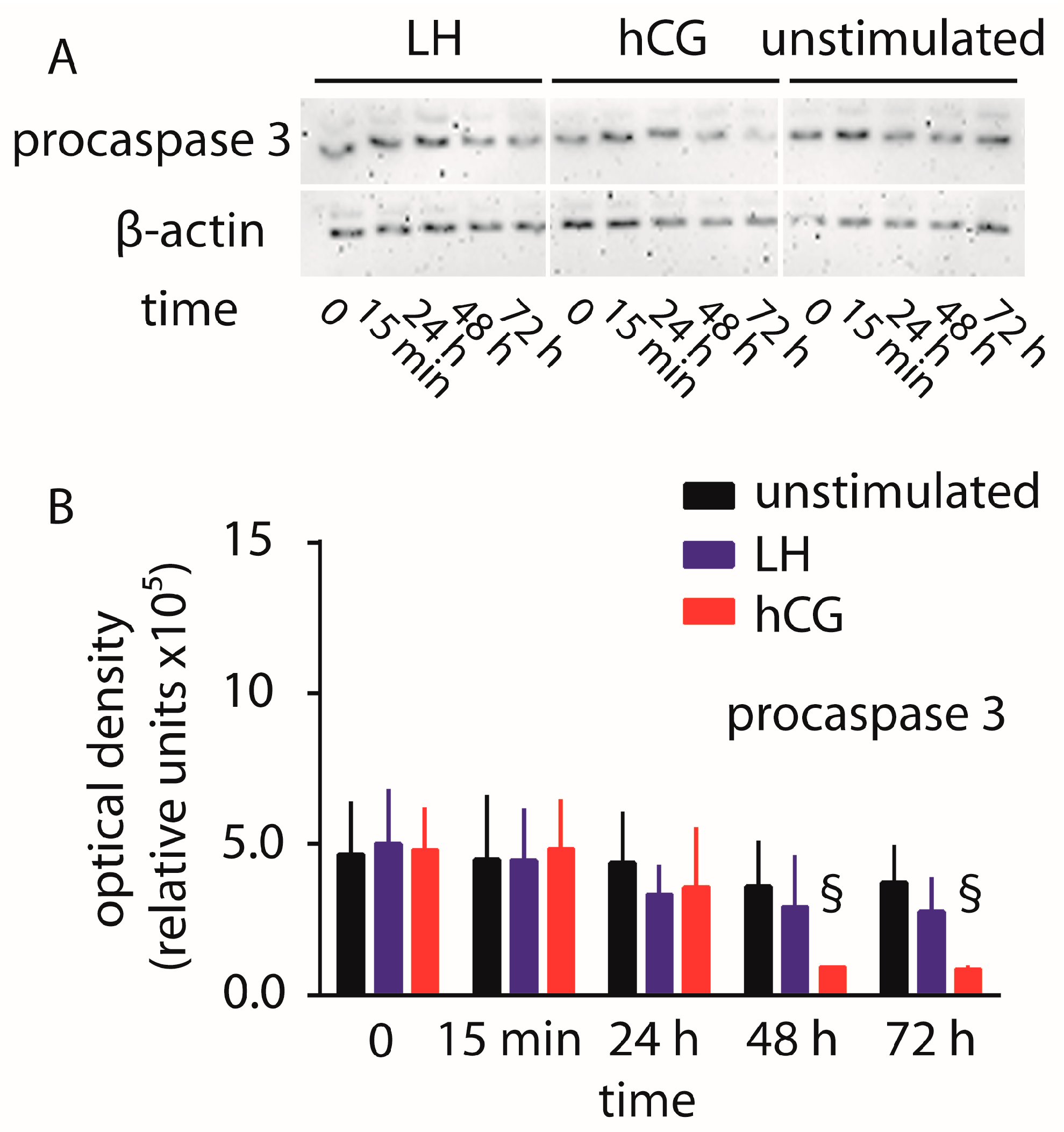
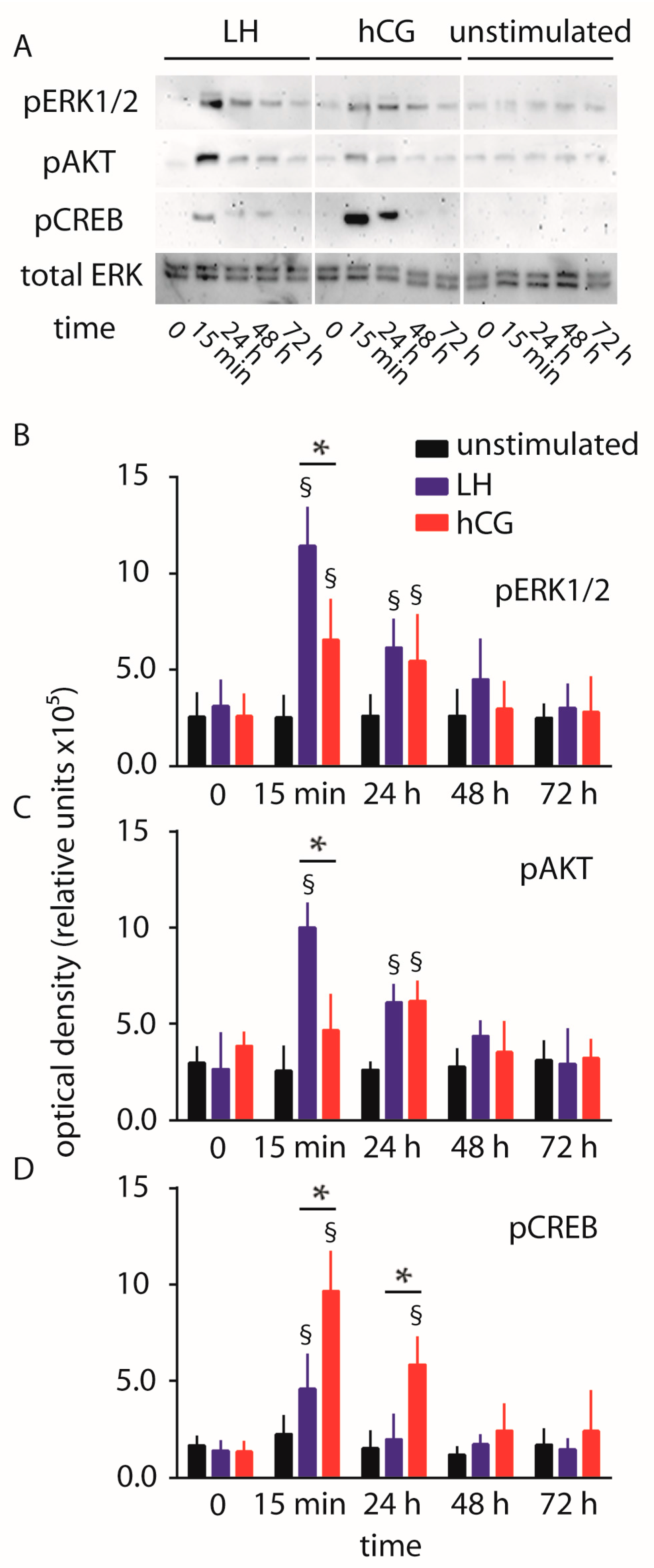
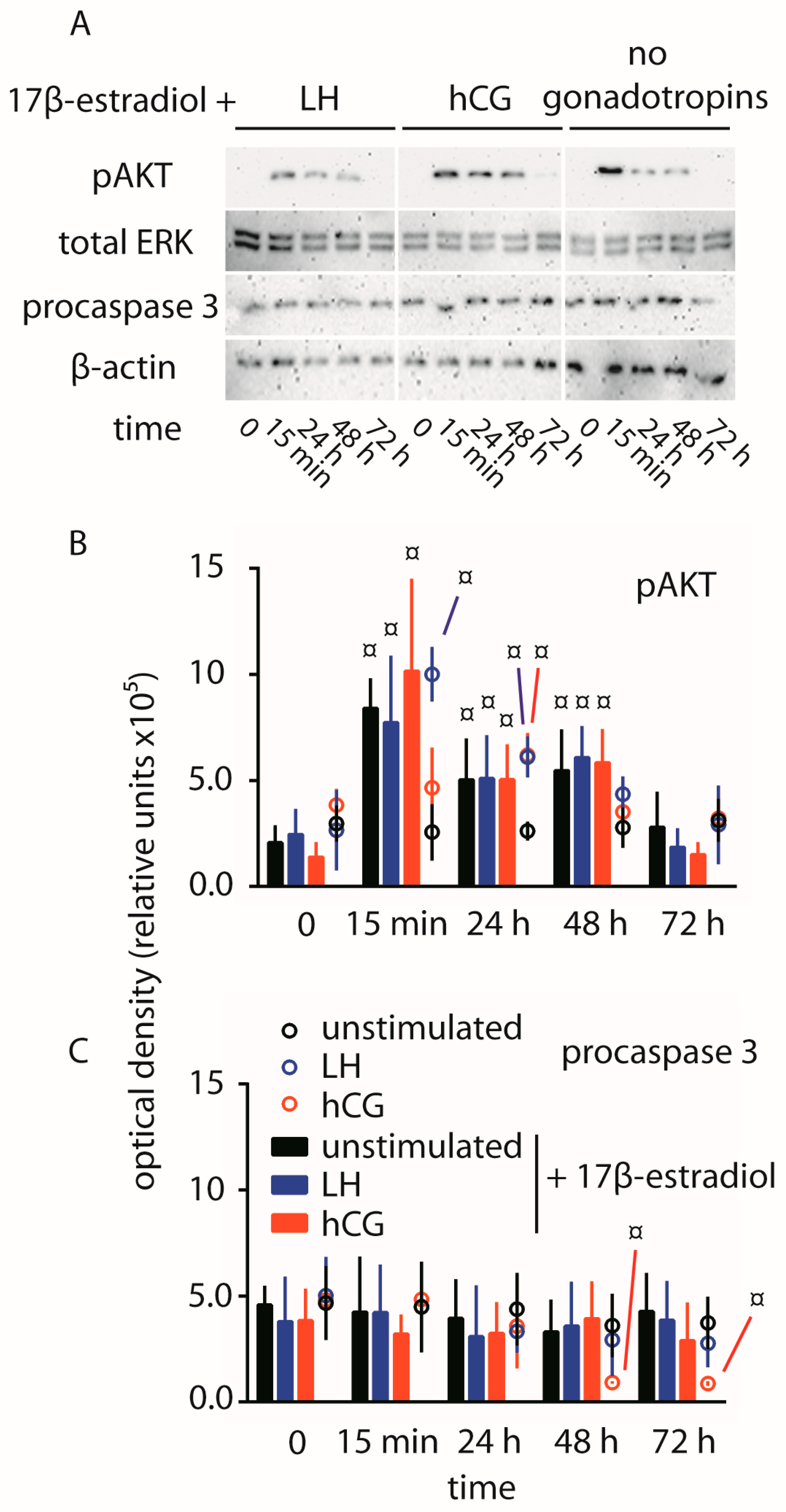
2.1. Cell Signaling Analysis and Procaspase 3 Cleavage
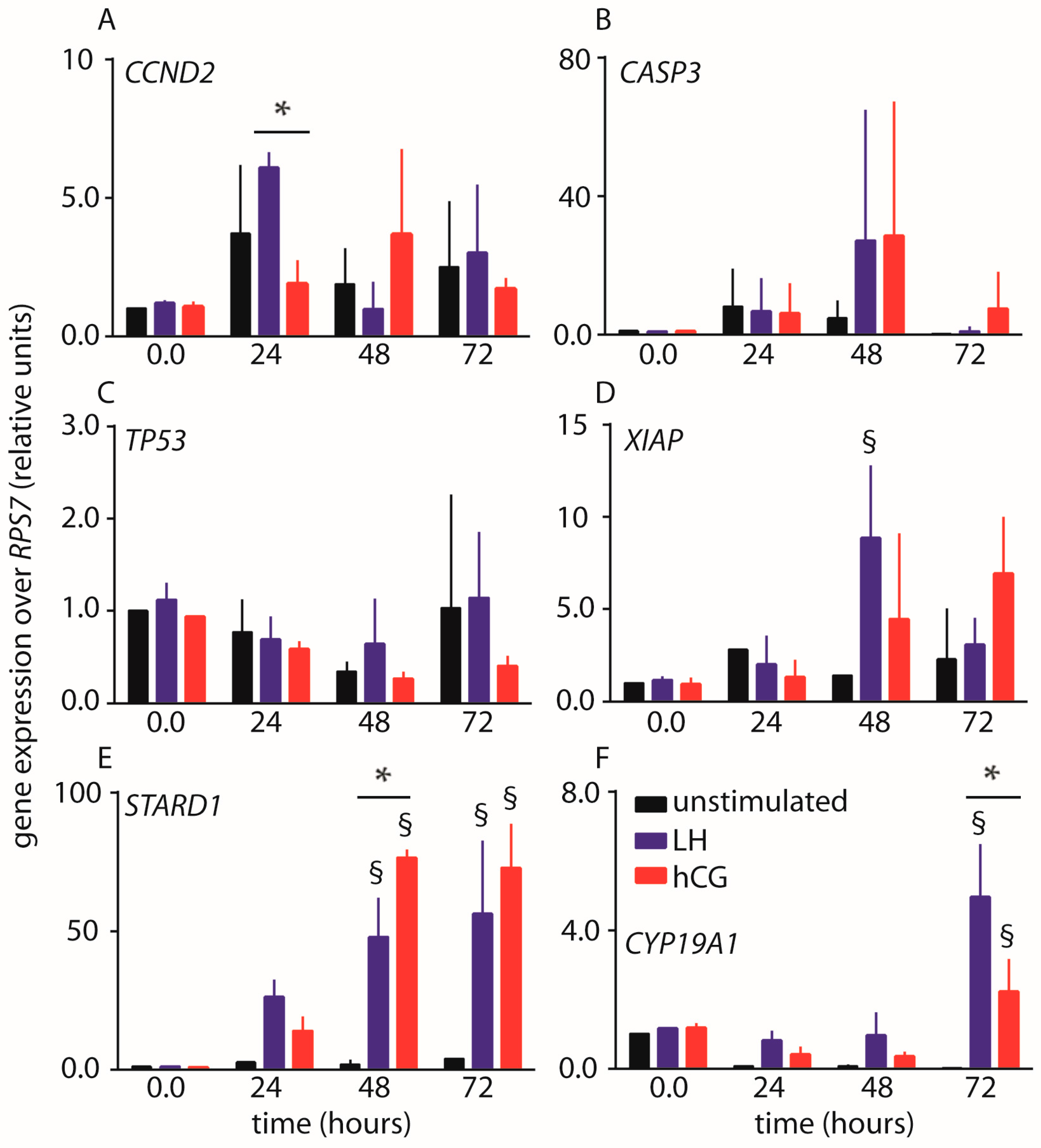
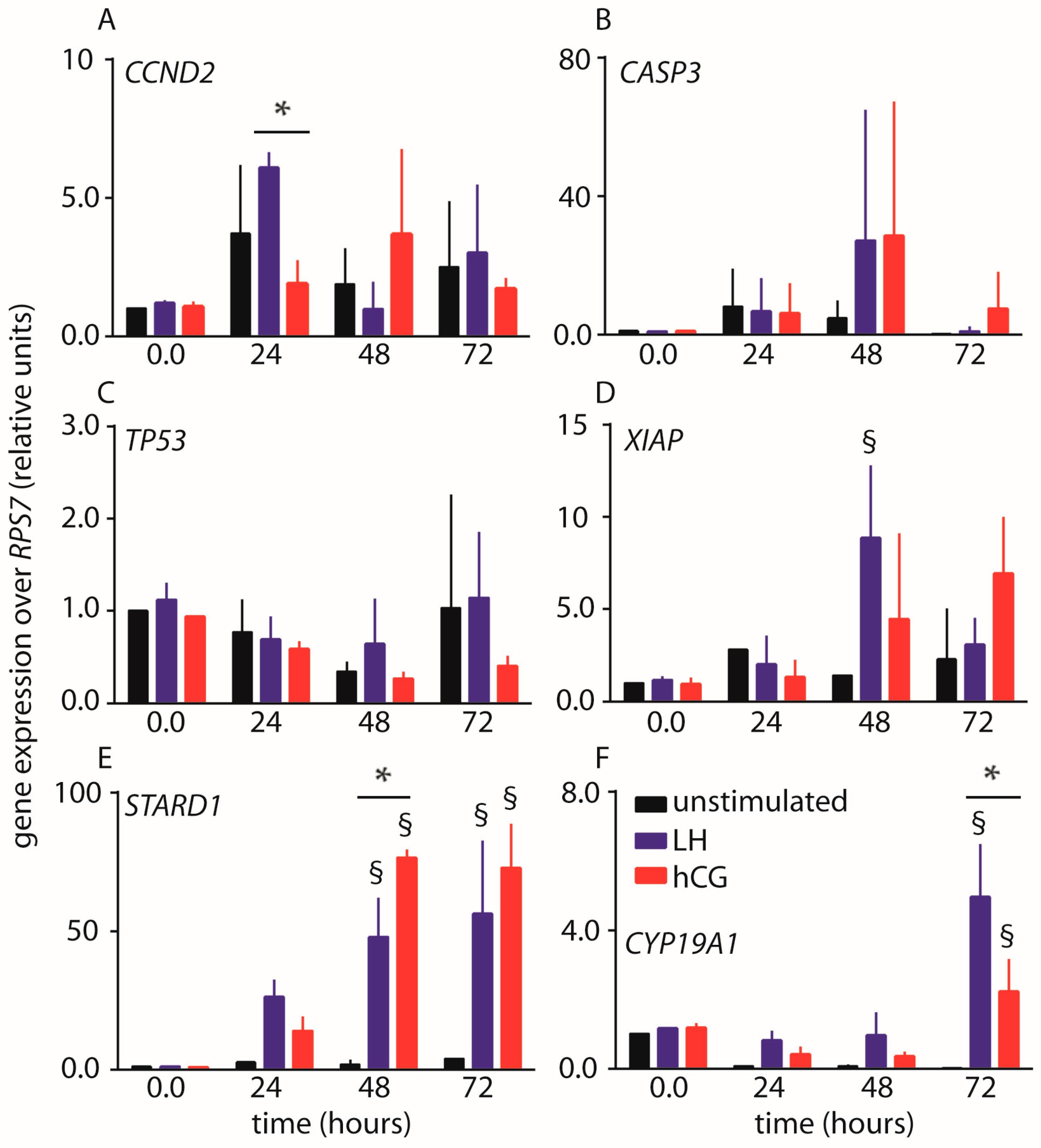
2.2. Gene Expression Analysis
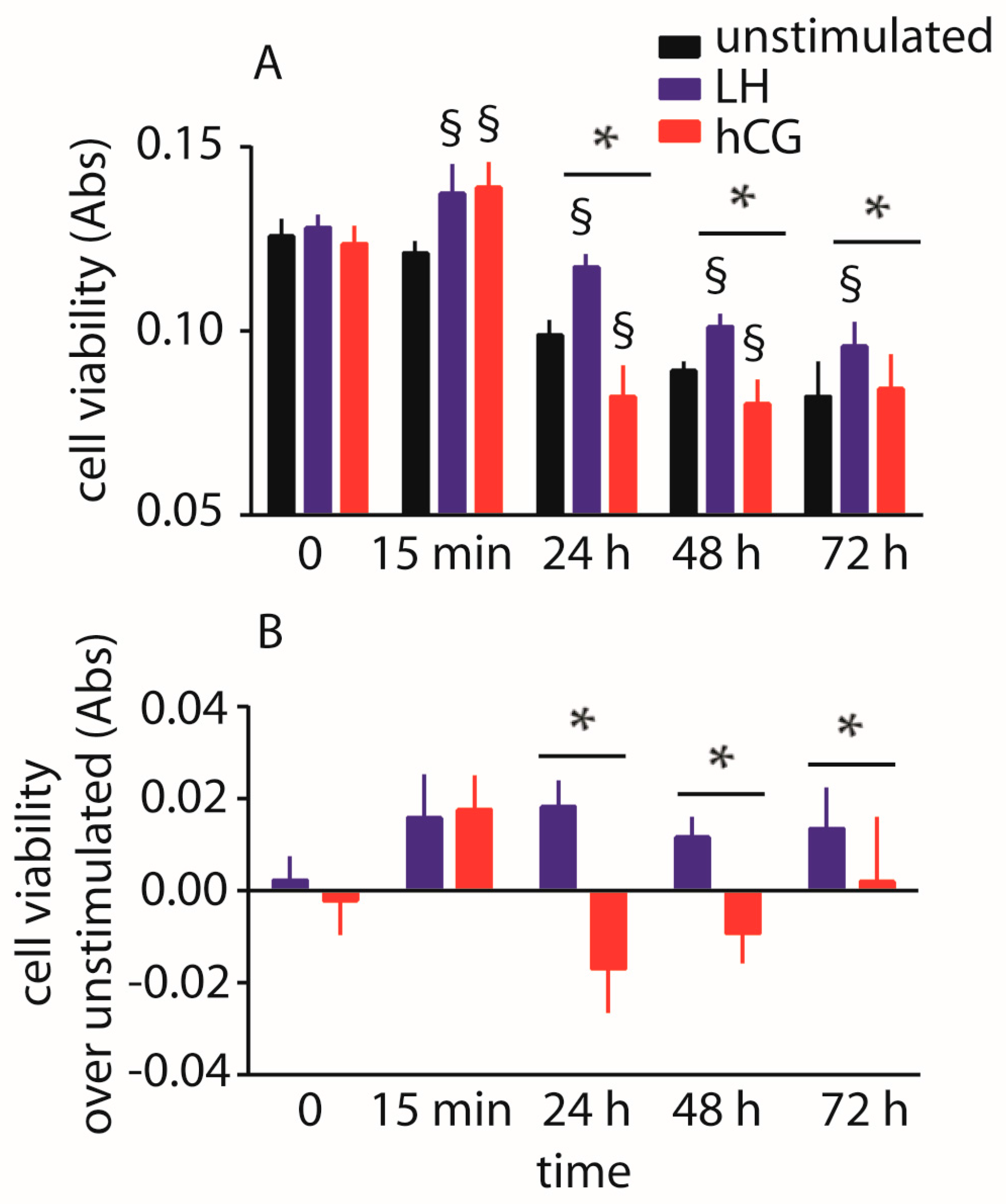
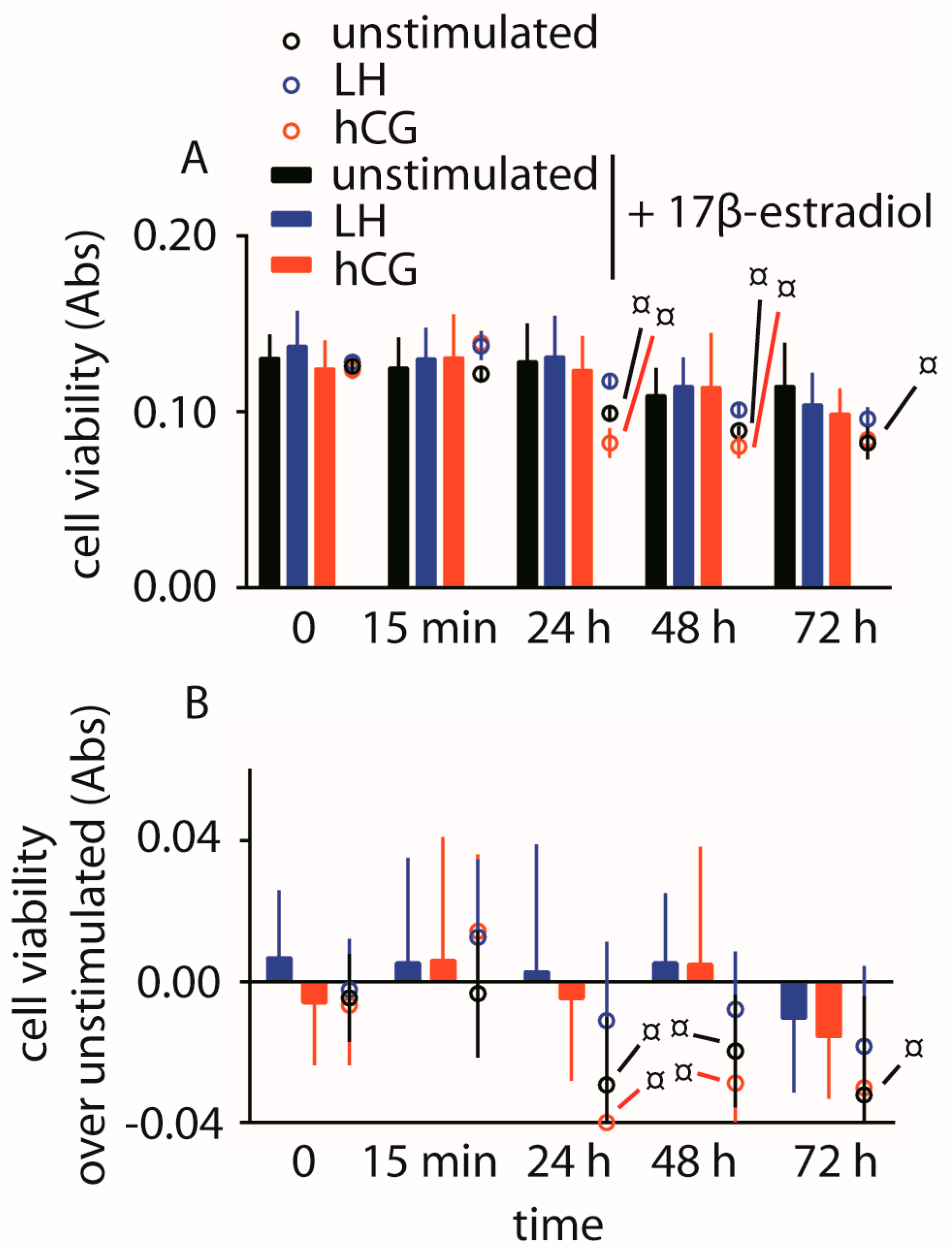
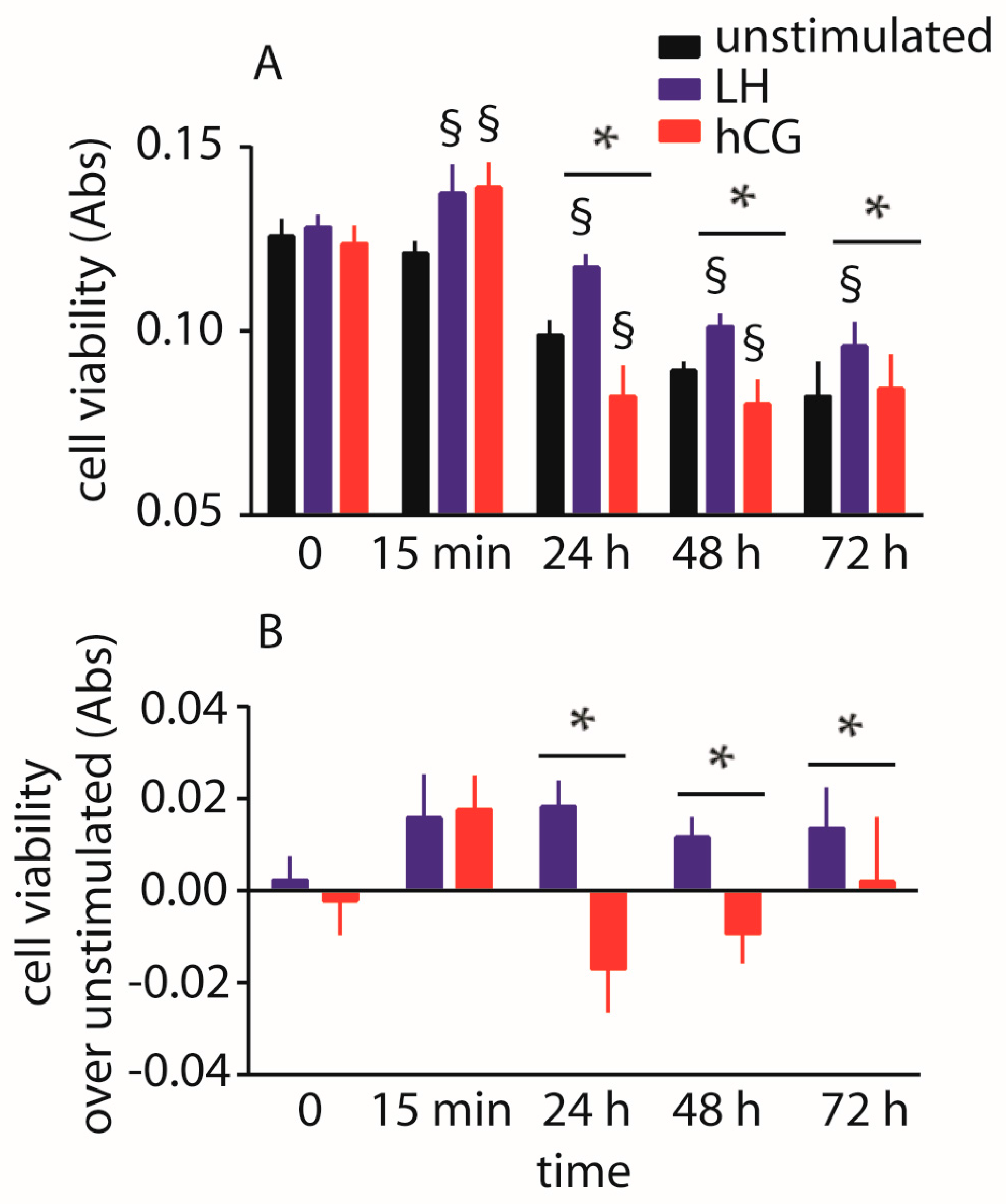
2.3. Evaluation of Cell Viability
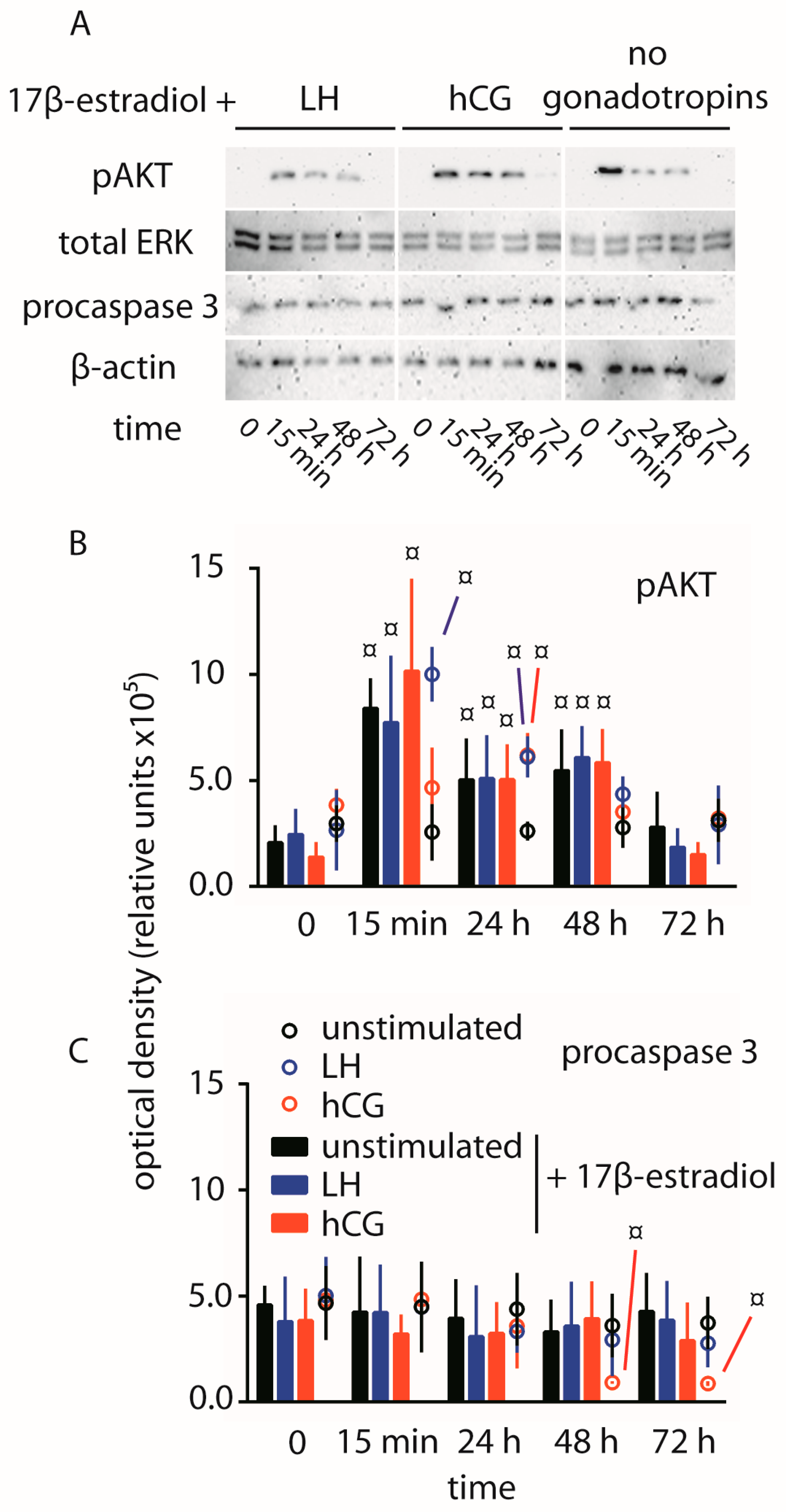
2.4. Abolition of Gonadotropin-Mediated Pro-Apoptotic Signals by Estrogens
3. Discussion
3.1. Gonadotropin-Mediated Intracellular Signaling In Vitro
3.2. Cross-Talk between Life and Death Signals
3.3. Estradiol as a Mediator of Proliferative and Anti-Apoptotic Effects
4. Materials and Methods
4.1. Recombinant Gonadotropins
4.2. Patients’ Selection
4.3. Granulosa Cells Isolation and Culture
4.4. Cell Line
4.5. Western Blot Analysis and Antibodies
4.6. Stimulation and Evaluation of Gene Expression
4.7. Analysis of Cell Viability
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Simoni, M.; Gromoll, J.; Nieschlag, E. The follicle-stimulating hormone receptor: Biochemistry, molecular biology, physiology, and pathophysiology. Endocr. Rev. 1997, 18, 739–773. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, M.; Fanelli, F.; Segaloff, D.L. The lutropin/choriogonadotropin receptor, a 2002 perspective. Endocr. Rev. 2002, 23, 141–174. [Google Scholar] [CrossRef] [PubMed]
- Troppmann, B.; Kleinau, G.; Krause, G.; Gromoll, J. Structural and functional plasticity of the luteinizing hormone/choriogonadotrophin receptor. Hum. Reprod. Update 2013, 19, 583–602. [Google Scholar] [CrossRef] [PubMed]
- Kossack, N.; Simoni, M.; Richter-Unruh, A.; Themmen, A.P.N.; Gromoll, J. Mutations in a novel, cryptic exon of the luteinizing hormone/chorionic gonadotropin receptor gene cause male pseudohermaphroditism. PLoS Med. 2008, 5, e88. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Lefkowitz, R.J. GRKs and β-arrestins: Roles in receptor silencing, trafficking and signaling. Trends Endocrinol. Metab. 2006, 17, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, J.A.; Tesmer, J.J.; Freeman, J.L.; Capel, W.D.; Stone, W.C.; Lefkowitz, R.J. Feedback inhibition of G protein-coupled receptor kinase 2 (GRK2) activity by extracellular signal-regulated kinases. J. Biol. Chem. 1999, 274, 34531–34534. [Google Scholar] [CrossRef] [PubMed]
- Strungs, E.G.; Luttrell, L.M. Arrestin-dependent activation of ERK and Src family kinases. Handb. Exp. Pharmacol. 2014, 219, 225–257. [Google Scholar] [PubMed]
- Casarini, L.; Reiter, E.; Simoni, M. β-arrestins regulate gonadotropin receptor-mediated cell proliferation and apoptosis by controlling different FSHR or LHCGR intracellular signaling in the hGL5 cell line. Mol. Cell. Endocrinol. 2016, 437, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, Z.; Keren-Tal, I.; Deutsch, M.; Dantes, A.; Aharoni, D.; Weinerb, A.; Tirosh, R.; Amsterdam, A. Fourier analysis of differential light scattering for the quantitation of FSH response associated with structural changes in immortalized granulosa cells. Mol. Cell. Endocrinol. 1996, 118, 145–153. [Google Scholar] [CrossRef]
- Zwain, I.H.; Amato, P. cAMP-induced apoptosis in granulosa cells is associated with up-regulation of P53 and bax and down-regulation of clusterin. Endocr. Res. 2001, 27, 233–349. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Gold, R.; Hosokawa, K.; Yoshida, Y.; Sasson, R.; Jung, Y.; Kotsuji, F. Crosstalk among multiple signaling pathways controlling ovarian cell death. Trends Endocrinol. Metab. 1999, 10, 255–262. [Google Scholar] [CrossRef]
- Amsterdam, A.; Keren-Tal, I.; Aharoni, D. Cross-talk between cAMP and p53-generated signals in induction of differentiation and apoptosis in steroidogenic granulosa cells. Steroids 1996, 61, 252–256. [Google Scholar] [CrossRef]
- Casarini, L.; Lispi, M.; Longobardi, S.; Milosa, F.; la Marca, A.; Tagliasacchi, D.; Pignatti, E.; Simoni, M. LH and hCG action on the same receptor results in quantitatively and qualitatively different intracellular signalling. PLoS ONE 2012, 7, e46682. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.R.O.; Carvalheira, J.B.C.; Lima, M.H.M.; Zimmerman, S.F.; Caperuto, L.C.; Amanso, A.; Gasparetti, A.L.; Meneghetti, V.; Zimmerman, L.F.; Velloso, L.A.; et al. Novel signal transduction pathway for luteinizing hormone and its interaction with insulin: activation of janus kinase/signal transducer and activator of transcription and phosphoinositol 3-kinase/Akt pathways. Endocrinology 2003, 144, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Casarini, L.; Riccetti, L.; de Pascali, F.; Nicoli, A.; Tagliavini, S.; Trenti, T.; la Sala, G.B.; Simoni, M. Follicle-stimulating hormone potentiates the steroidogenic activity of chorionic gonadotropin and the anti-apoptotic activity of luteinizing hormone in human granulosa-lutein cells in vitro. Mol. Cell. Endocrinol. 2016, 422, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Chapekar, T.; Chhabra, Y.; Singh, P.; Sinha, S.; Luthra, K. Differential response to sustained stimulation by hCG & LH on goat ovarian granulosa cells. Indian J. Med. Res. 2012, 135, 331–340. [Google Scholar] [PubMed]
- Riccetti, L.; de Pascali, F.; Gilioli, L.; Potì, F.; Giva, L.B.; Marino, M.; Tagliavini, S.; Trenti, T.; Fanelli, F.; Mezzullo, M.; et al. Human LH and hCG stimulate differently the early signalling pathways but result in equal testosterone synthesis in mouse Leydig cells in vitro. Reprod. Biol. Endocrinol. 2017, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Terranova, P.F.; Rice, V.M. Review: Cytokine involvement in ovarian processes. Am. J. Reprod. Immunol. 1997, 37, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami, I.; Armon, L.; Freimann, S.; Strassburger, D.; Ron-El, R.; Amsterdam, A. EGF-like growth factors as LH mediators in the human corpus luteum. Hum. Reprod. 2008, 24, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Britt, K.L.; Findlay, J.K. Estrogen actions in the ovary revisited. J. Endocrinol. 2002, 175, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Palter, S.F.; Tavares, A.B.; Hourvitz, A.; Veldhuis, J.D.; Adashi, E.Y. Are estrogens of import to primate/human ovarian folliculogenesis? Endocr. Rev. 2001, 22, 389–424. [Google Scholar] [PubMed]
- Pavlik, R.; Wypior, G.; Hecht, S.; Papadopoulos, P.; Kupka, M.; Thaler, C.; Wiest, I.; Pestka, A.; Friese, K.; Jeschke, U. Induction of G protein-coupled estrogen receptor (GPER) and nuclear steroid hormone receptors by gonadotropins in human granulosa cells. Histochem. Cell Biol. 2011, 136, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Heublein, S.; Mayr, D.; Friese, K.; Jarrin-Franco, M.; Lenhard, M.; Mayerhofer, A.; Jeschke, U. The G-protein-coupled estrogen receptor (GPER/GPR30) in ovarian granulosa cell tumors. Int. J. Mol. Sci. 2014, 15, 15161–15172. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Liu, M.; Luo, H.; Wu, C.; Tang, X.; Tang, S.; Hu, P.; Yan, Y.; Wang, Z.; Tu, G. GPER mediates enhanced cell viability and motility via non-genomic signaling induced by 17β-estradiol in triple-negative breast cancer cells. J. Steroid Biochem. Mol. 2014, 143, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Zavatti, M.; Guida, M.; Maraldi, T.; Beretti, F.; Bertoni, L.; la Sala, G.B.; de Pol, A. Estrogen receptor signaling in the ferutinin-induced osteoblastic differentiation of human amniotic fluid stem cells. Life Sci. 2016, 164, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Liao, L.-Y.; Zhao, T.-T.; Mo, X.-M.; Chen, G.G.; Liu, Z.-M. GPER/ERK&AKT/NF-κB pathway is involved in cadmium-induced proliferation, invasion and migration of GPER-positive thyroid cancer cells. Mol. Cell. Endocrinol. 2017, 442, 68–80. [Google Scholar] [PubMed]
- Casarini, L.; Moriondo, V.; Marino, M.; Adversi, F.; Capodanno, F.; Grisolia, C.; la Marca, A.; la Sala, G.B.; Simoni, M. FSHR polymorphism p.N680S mediates different responses to FSH in vitro. Mol. Cell. Endocrinol. 2014, 393, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.E.; Findlay, J.K. The role of estrogen in folliculogenesis. Mol. Cell. Endocrinol. 1999, 151, 57–64. [Google Scholar] [CrossRef]
- Grzesik, P.; Kreuchwig, A.; Rutz, C.; Furkert, J.; Wiesner, B.; Schuelein, R.; Kleinau, G.; Gromoll, J.; Krause, G. Differences in signal activation by LH and hCG are mediated by the LH/CG Receptor’s extracellular hinge region. Front. Endocrinol. 2015, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- Huhtaniemi, I.T.; Catt, K.J. Differential binding affinities of rat testis luteinizing hormone (LH) receptors for human chorionic gonadotropin, human LH, and ovine LH. Endocrinology 1981, 108, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Gromoll, J.; Eiholzer, U.; Nieschlag, E.; Simoni, M. Male hypogonadism caused by homozygous deletion of exon 10 of the luteinizing hormone (LH) receptor: Differential action of human chorionic gonadotropin and LH. J. Clin. Endocrinol. Metab. 2000, 85, 2281–2286. [Google Scholar] [CrossRef] [PubMed]
- Gromoll, J.; Wistuba, J.; Terwort, N.; Godmann, M.; Müller, T.; Simoni, M. A new subclass of the luteinizing hormone/chorionic gonadotropin receptor lacking exon 10 messenger RNA in the New World Monkey (Platyrrhini) lineage. Biol. Reprod. 2003, 69, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Dantes, A.; Hosokawa, K.; Schere-Levy, C.P.; Kotsuji, F.; Aharoni, D. Steroid regulation during apoptosis of ovarian follicular cells. Steroids 1998, 63, 314–318. [Google Scholar] [CrossRef]
- Amsterdam, A.; Keren-Tal, I.; Aharoni, D.; Dantes, A.; Land-Bracha, A.; Rimon, E.; Sasson, R.; Hirsh, L. Steroidogenesis and apoptosis in the mammalian ovary. Steroids 2003, 68, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-Y.; Cheung, C.K.M.; Wang, Y.; Tsang, B.K. Regulation of cell death and cell survival gene expression during ovarian follicular development and atresia. Front. Biosci. 2003, 8, d222–d237. [Google Scholar] [PubMed]
- Tajima, K.; Hosokawa, K.; Yoshida, Y.; Dantes, A.; Sasson, R.; Kotsuji, F.; Amsterdam, A. Establishment of FSH-responsive cell lines by transfection of pre-ovulatory human granulosa cells with mutated p53 (p53val135) and Ha-ras genes. Mol. Hum. Reprod. 2002, 8, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Hunzicker-Dunn, M.E.; Lopez-Biladeau, B.; Law, N.C.; Fiedler, S.E.; Carr, D.W.; Maizels, E.T. PKA and GAB2 play central roles in the FSH signaling pathway to PI3K and AKT in ovarian granulosa cells. Proc. Natl. Acad. Sci. USA 2012, 109, E2979–E2988. [Google Scholar] [CrossRef] [PubMed]
- Muniz, L.C.; Yehia, G.; Mémin, E.; Ratnakar, P.V.A.L.; Molina, C.A. Transcriptional Regulation of Cyclin D2 by the PKA Pathway and Inducible cAMP Early Repressor in Granulosa Cells. Biol. Reprod. 2006, 75, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kayampilly, P.P.; Menon, K.M.J. Inhibition of extracellular signal-regulated protein kinase-2 phosphorylation by dihydrotestosterone reduces follicle-stimulating hormone-mediated cyclin D2 messenger ribonucleic acid expression in rat granulosa cells. Endocrinology 2004, 145, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hao, X.; Yang, J.; Li, J.; Zhang, M. CREB activity is required for luteinizing hormone-induced the expression of EGF-like factors. Mol. Reprod. Dev. 2016, 83, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Hanoch, T.; Dantes, A.; Tajima, K.; Strauss, J.F.; Seger, R. Mechanisms of gonadotropin desensitization. Mol. Cell. Endocrinol. 2002, 187, 69–74. [Google Scholar] [CrossRef]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.A.; Kliman, H.J.; Strauss, J.F.; Paavola, L.G. Gonadotropins and cyclic adenosine 3′,5′-monophosphate (cAMP) alter the morphology of cultured human granulosa cells. Biol. Reprod. 1986, 34, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Billig, H.; Chun, S.Y.; Eisenhauer, K.; Hsueh, A.J. Gonadal cell apoptosis: Hormone-regulated cell demise. Hum. Reprod. Update 1996, 2, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Quirk, S.M.; Cowan, R.G.; Harman, R.M. The susceptibility of granulosa cells to apoptosis is influenced by oestradiol and the cell cycle. J. Endocr. 2006, 189, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; LaRocca, J.; Pietruska, J.; Ota, M.; Anderson, L.; Duncan Smith, S.; Weston, P.; Rasoulpour, T.; Hixon, M.L. Subfertility caused by altered follicular development and oocyte growth in female mice lacking PKBα/Akt11. Biol. Reprod. 2010, 82, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Feng, X.; Chen, J.; Qi, X.; Zhou, L.; Shi, S.; KC, K.; Stanley, J.L.; Baker, P.N.; Zhang, H. G protein-coupled receptor 30 regulates trophoblast invasion and its deficiency is associated with preeclampsia. J. Hypertens. 2016, 34, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, Y.; Wang, S.; Liu, Y.; Zhang, J.; Xu, Y.; Zhang, Z.; Bao, W.; Wu, S. LSD1 sustains estrogen-driven endometrial carcinoma cell proliferation through the PI3K/AKT pathway via di-demethylating H3K9 of cyclin D1. Int. J. Oncol. 2017, 50, 942–952. [Google Scholar] [PubMed]
- Nordhoff, V.; Sonntag, B.; Von Tils, D.; Götte, M.; Schüring, A.N.; Gromoll, J.; Redmann, K.; Casarini, L.; Simoni, M. Effects of the FSH receptor gene polymorphism p.N680S on cAMP and steroid production in cultured primary human granulosa cells. Reprod. Biomed. Online 2011, 23, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Rainey, W.H.; Sawetawan, C.; Shay, J.W.; Michael, M.D.; Mathis, J.M.; Kutteh, W.; Byrd, W.; Carr, B.R. Transformation of human granulosa cells with the E6 and E7 regions of human papillomavirus. J. Clin. Endocrinol. Metab. 1994, 78, 705–710. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene and Primer Type | Primer Sequence from 5′ to 3′ | NCBI or GenBank Accession Number |
|---|---|---|
| CCND2 forward | CTGGCCTCCAAACTCAAAGA | NM_001759.3 |
| CCND2 reverse | TTCCACTTCAACTTCCCCA | |
| CASP3 forward | TGTTTGTGTGCTTCTGAGCC | AY219866.1 |
| CASP3 reverse | TCTACAACGATCCCCTCTGAA | |
| TP53 forward | GAGAACACCGCTTGGAACTA | NG_017013.2 |
| TP53 reverse | AGCCCACTTACAGCCTTTCA | |
| XIAP forward | TTGAAAATAGTGCCACGCAG | NM_001167.3 |
| XIAP reverse | TGTGTCTCAGATGGCCTGTC | |
| STARD1 forward | AAGAGGGCTGGAAGAAGGAG | NM_000349.2 |
| STARD1 reverse | TCTCCTTGACATTGGGGTTC | |
| CYP19A1 forward | CCCTTCTGCGTCGTGTCAT | NM_000103.3 |
| CYP19A1 reverse | GATTTTAACCACGATAGCACTTTCG | |
| RPS7 forward | AATCTTTGTTCCCGTTCCTCA | NM_001011.3 |
| RPS7 reverse | TTCTGCCTAAGCCAACTCG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casarini, L.; Riccetti, L.; De Pascali, F.; Gilioli, L.; Marino, M.; Vecchi, E.; Morini, D.; Nicoli, A.; La Sala, G.B.; Simoni, M. Estrogen Modulates Specific Life and Death Signals Induced by LH and hCG in Human Primary Granulosa Cells In Vitro. Int. J. Mol. Sci. 2017, 18, 926. https://doi.org/10.3390/ijms18050926
Casarini L, Riccetti L, De Pascali F, Gilioli L, Marino M, Vecchi E, Morini D, Nicoli A, La Sala GB, Simoni M. Estrogen Modulates Specific Life and Death Signals Induced by LH and hCG in Human Primary Granulosa Cells In Vitro. International Journal of Molecular Sciences. 2017; 18(5):926. https://doi.org/10.3390/ijms18050926
Chicago/Turabian StyleCasarini, Livio, Laura Riccetti, Francesco De Pascali, Lisa Gilioli, Marco Marino, Eugenia Vecchi, Daria Morini, Alessia Nicoli, Giovanni Battista La Sala, and Manuela Simoni. 2017. "Estrogen Modulates Specific Life and Death Signals Induced by LH and hCG in Human Primary Granulosa Cells In Vitro" International Journal of Molecular Sciences 18, no. 5: 926. https://doi.org/10.3390/ijms18050926