
Mammalian Metallothionein-3: New Functional and Structural Insights

1
Department of Chemistry B, University of Zürich, Winterthurerstrasse 190, CH-8057 Zürich, Switzerland
2
Department of Chemistry and Biochemistry, University of Texas at Dallas, 800 W Campbell Road, Richardson, TX 75080-3021, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(6), 1117; https://doi.org/10.3390/ijms18061117
Submission received: 15 April 2017
/
Revised: 12 May 2017
/
Accepted: 15 May 2017
/
Published: 24 May 2017
(This article belongs to the Special Issue Metallothioneins in Bioinorganic Chemistry: Recent Developments)
Abstract
:Metallothionein-3 (MT-3), a member of the mammalian metallothionein (MT) family, is mainly expressed in the central nervous system (CNS). MT-3 possesses a unique neuronal growth inhibitory activity, and the levels of this intra- and extracellularly occurring metalloprotein are markedly diminished in the brain of patients affected by a number of metal-linked neurodegenerative disorders, including Alzheimer’s disease (AD). In these pathologies, the redox cycling of copper, accompanied by the production of reactive oxygen species (ROS), plays a key role in the neuronal toxicity. Although MT-3 shares the metal-thiolate clusters with the well-characterized MT-1 and MT-2, it shows distinct biological, structural and chemical properties. Owing to its anti-oxidant properties and modulator function not only for Zn, but also for Cu in the extra- and intracellular space, MT-3, but not MT-1/MT-2, protects neuronal cells from the toxicity of various Cu(II)-bound amyloids. In recent years, the roles of zinc dynamics and MT-3 function in neurodegeneration are slowly emerging. This short review focuses on the recent developments regarding the chemistry and biology of MT-3.

1. Introduction
Aging, a major risk factor for neurodegenerative disorders, is accompanied with structural, chemical, functional, neuropsychological, and genetic changes, with increased susceptibility to diseases and cognitive impairments [1]. In the normal brain, a high concentration of essential transition metal ions such as zinc, copper, and iron is present. The homeostasis of these transition metals is tightly regulated and essential for brain physiology [2,3]. In physiological conditions, micromolar concentrations of zinc and copper are actively released from neurons during neurotransmission processes into pre- and postsynaptic clefts. However, in dysregulated metal metabolism, present in neurodegenerative disorders, the ill-binding of copper and zinc to the disease specific amyloidogenic peptides or proteins correlates with deposition of amyloid fibrils. Among the most prominent examples are the Aβ peptides in Alzheimer (AD), α-synuclein in Parkinson (PD), prion protein in prion deposits typical of Creutzfeldt–Jakob disease (CJD), and superoxide dismutase-1 (SOD-1) in amyotrophic lateral sclerosis (ALS). In these diseases, copper, due to its reactivity with molecular oxygen (O2), generates neurotoxic reactive oxygen species (ROS) such as superoxide, hydrogen peroxide, and hydroxyl radicals [2,3].
In the central nervous system (CNS), the natural metal chelator metallothionein-3 (MT-3) represents one of the major players in copper and zinc homeostasis. This protein occurs intra- and extracellularly and was found down-regulated in AD [4]. However, in contrast to the almost exclusive expression of MT-3 in the CNS [5], the expression of MT-1/-2 isoforms are found in almost all organs. Besides the differences in the expression pattern, the control of MT-3 expression also differs from the MT-1/-2 isoforms. Thus, in marked contrast to the induction of MT-1/-2 biosynthesis by a range of factors including glucocorticoids, cytokines, reactive oxygen species, and metal ions [6], the expression of MT-3 is unresponsive to these inducers. However, the MT-3 gene was identified as hypoxia-inducible in several human tissues. Here the induction of MT-3 in cultured human astrocytes by hypoxia suggested that the protein may protect the brain from hypoxic damage [7]. Furthermore, a similar hypoxic induction of the MT-3 gene in human adipocytes implied that the protein may protect adipocytes from hypoxic damage [8]. In other studies, the question of whether the MT-3 and MT-1 isoforms possess different biological properties has been addressed using cultured kidney cells that allowed constitutive expression of both isoforms. The studies revealed that under zinc deficient conditions MT-3, but not MT-1 inhibited the cell growth [9]. The neuronal growth inhibitory activity of MT-3 appears to be the most prominent biological property of this protein. In this regard, an extracellular addition of MT-3, but not MT-1/-2 counteract the ability of AD brain extract to stimulate survival and neuritic sprouting of cultured neurons [10,11]. Both the growth inhibitory activity and the protective effect of solely MT-3 from the toxic effect of amyloid peptide Aβ1–40 [10] have been linked to its possible role in the pathogenesis of AD. However, the bioactivity and the protective role of MT-3 in AD are functionally unrelated. An increased interest in the physiological and pathophysiological processes in the human brain brought about an increased research into the understanding of the structure and biology of MT-3. In recent years, the list of MT-3 functions suggesting its role not only in the CNS but also outside of this organ, is steadily increasing. At present, there are a few reviews on record dealing with the MT-1 through MT-3 isoforms [12,13,14,15,16,17,18,19,20,21,22]. This review focuses on the recent advances in our knowledge regarding the structural/chemical and functional properties of MT-3 in metal related biological processes.
2. Structural and Chemical Properties of Metallothionein-3
As recent publications cover the protective role of MT-3 in AD, and its possible role in lead detoxification in the human brain, we wish to briefly summarize the current knowledge about the structure and reactivity of the protein required to discuss these new reports. Since the bioactivity of MT-3 in neuronal assays has been established for the native protein, containing both copper and zinc ions, and for the recombinant human Zn7MT-3 [4,11,23], the structural features of both metalloforms have been investigated (Table 1). The structural studies on recombinant MII7MT-3 revealed that, like the MII7MT-1/-2 isoforms, the seven divalent metal ions are tetrahedrally coordinated through the array of 20 conserved cysteines forming two metal-thiolate clusters, i.e., MII3(CysS)9 and MII4(CysS)11. A MII3(CysS)9 metal-cluster is located in the N-terminal β-domain (residues 1–31) and a MII4(CysS)11 cluster is located in the C-terminal α-domain (residues 32–68) of MII7MT-3 [24,25]. In these clusters, unprecedented dynamic processes exist. A fast exchange between conformational cluster substates and very slow exchange processes between configurational cluster substates in the β-domain differentiate MT-3 from other isoforms [26]. As a result, only the 3D structure of the C-terminal α-domain of mouse and human 113Cd7MT-3, containing an adamantane-like Cd4Cys11 cluster, could be determined by NMR measurements [27,28]. The structure of this domain reveals a peptide fold and cluster organization very similar to that found in mammalian Cd7MT-1/-2 (Figure 1A).
Native MT-3, as isolated from human [4] and bovine [31] brains, contains four Cu(I) and between three and four Zn(II) ions. The isolated Cu(I)4Zn3–4MT-3 is a monomeric protein that is stable in air. Extended X-ray absorption fine structure (EXAFS) studies on the native protein revealed the presence of two homometallic clusters, a Cu(I)4–thiolate cluster and a Zn3–4–thiolate cluster [31]. In contrast to tetrahedrally coordinated Zn(II) ions, Cu(I) ions are diagonally and/or trigonally coordinated by two or three cysteine ligands [32]. The localization of both clusters in the protein structure was established by the spectroscopic and immunochemical studies of Cu(I)4,Zn4MT-3. Thus, the Cu(I)4–thiolate cluster is located in the N-terminal β-domain and the Zn4–thiolate cluster in the C-terminal α-domain [22,33]. Based on the spectroscopic and luminescence properties of air-stable Cu(I)4,Zn4MT-3 (Table 1), generated in the aerobic reaction between Zn7MT-3 and free Cu(II), the presence of a Cu(I)4S5−x cluster containing five reduced thiolates and two disulfide bonds has been postulated (Figure 1B) [29,30]. It may be noted that the adamantane-like {M4S6} polyhedron is the most frequently observed species in copper-thiolate chemistry. Therefore, by analogy with inorganic model complexes, besides the reduced cysteine thiolates the participation of a disulfide bridge sulfur (indicated by x) in the Cu(I) coordination cannot be excluded.
A fundamental question about all metalloproteins is what determinants control which metals they bind in vivo. In some cases, metals are delivered to the metalloproteins by specialized metallochaperones. However, for most metalloproteins, a critical factor is the availability of the appropriate metal species in the buffered pools in the cell. In a number of studies, the Escherichia coli expression system was used to address the question regarding the in vivo metal-binding abilities of a number of MTs [34]. In this regard, the particular MT was first recombinantly expressed in E. coli cultures enriched with Zn(II), Cd(II), or Cu(II). Subsequently, ZnMT preparations were then titrated in vitro with Cd(II) or Cu(I) ions and the generated metalloforms analyzed. A comparison of the E. coli generated metalloforms with those formed in vitro was taken to indicate the metal binding preference of the particular MT. Based on such studies carried out on MT-3, it has been concluded that this MT isoform exhibits a marked Cu–thionein character, with a high tendency to coordinate Cu(I) ions [35]. In all these studies, a high degree of metal selectivity is discussed by the authors in terms of inherent properties of MTs. A critical assessment of this approach in assigning the metal selectivity of MTs in vivo has been given [35]. In this work, it has been put forward that all metals in the cell are present buffered to given concentrations, due to their binding to other proteins and low molecular weight compounds. The metal binding preference of a particular MT in E. coli cultures implies that the metal selectivity reflects its structural properties. However, the E. coli cytosol still contains other proteins and low molecular weight compounds contributing to the metal buffering. Consequently, the exact maintenance of the ratios of buffered metal ions will dictate the correct metal incorporation into a protein. Overall, it would appear that the binding of the correct metal into the protein structure might be regulated by adjusting the ratios of buffered metal ions to different location in various cell types [36]. A similar conclusion has been drawn from computational studies in which, in the absence of metallochaperons, the local metal concentrations were found critical for the metal specificity of proteins [37]. Although the enhanced copper-thionein character of MT-3 over MT-1/-2 may still, in part, reflect subtle differences between the two sequences and consequently their structural properties [38], it would appear that the reported metal binding preference of various MTs in E. coli cultures may not exactly reflect the buffered metal concentrations present in different mammalian cell types where the MT biosynthesis occurs.
3. Metallothionein-3 in Metal-Linked Neurodegenerative Disorders
The protective role of MT-3 from AD pathology has been implicated based on the finding that the protein (a) is down-regulated in AD, (b) shows extracellular growth inhibitory activity and antagonizes the ability of AD brain extract to stimulate survival and neuritic sprouting, and (c) protects, by an unknown mechanism, the neuronal cells from the toxic effect of amyloid peptide Aβ1–40 [4,5,10,11,39]. In an AD brain, Aβ forms amyloid plaques in specific regions in the brain, which have been considered a key element in its pathogenesis [40,41,42]. Furthermore, a large body of evidence connects the metal-induced aggregation of Aβ by transition metal ions copper and zinc and accompanied oxidative stress with the disease progression [43]. In contrast to redox-inert zinc, redox-active copper aberrantly bound to amyloidogenic Aβ peptides can activate molecular oxygen, resulting in the production of ROS through Fenton and Haber–Weiss reactions [44]. In view of the observation that extracellular Zn7MT-3 protects the neuronal cells from the toxic effect of amyloid peptide Aβ1–40 [10,45], an interaction of free Cu(II) and that bound in Cu(II)–Aβ1–40 with Zn7MT-3 has been found responsible for this protective effect [30]. The spectroscopic and mass spectrometric studies on the interaction between free Cu(II) and human Zn7MT-3 revealed that concomitant with the Cu(II) reduction to Cu(I) by the thiolate ligands the monovalent copper is bound to the protein. As a result, an air-stable Cu(I)4Zn4MT-3 species is formed cooperatively in the β-domain together with two intramolecular disulfide bonds and the release of three originally bound Zn(II) ions. Furthermore, the Cu(II)-mediated hydroxyl radical production was fully abolished, indicating that the protein, through the formation of the redox-inert Cu(I)4Zn4MT-3 structure, efficiently scavenges the free Cu(II) ions [30]. In the follow up studies, the question as to the reported protective effect of human Zn7MT-3 against Aβ1–40 toxicity in neuronal cell culture [10] has been addressed. The toxicity of Aβ1–40 stems mainly from the formed Cu(II)–Aβ1–40 complex in the extracellular space and its interaction with molecular oxygen, whereby the ROS are generated. In vitro studies of the interaction between Cu(II)–Aβ1–40 and Zn7MT-3 established that the protein can efficiently remove Cu(II) from both the soluble and insoluble aggregates of Cu(II)–Aβ1–40. The products of this reaction are the already discussed air-stable Cu(I)4Zn4MT-3 species and the redox-inert Zn(II)–Aβ1–40 complex. In addition, the metal-swap between Zn7MT-3 and Cu(II)–Aβ1–40 fully abolishes the Cu(II)-mediated ROS production. This protective effect against the ROS toxicity has been demonstrated not only by in vitro experiments but also in human neuroblastoma cell cultures [46]. The mechanism of exchange of metal ions between Zn7MT-3 and Cu(II)–Aβ1–40 was also investigated by spectroscopy and transmission electron microscopy [47]. The studies revealed that metal ion exchange occurs via free Cu(II) on a time scale of seconds to minutes and that metal ion exchange induces time-dependent amyloidogenic structural and morphological changes in Aβ1–40 on a time scale of hours. The morphological changes were primarily due to the binding of Zn(II) to Aβ1–40 aggregates [48]. Taken together, these studies corroborate a possible beneficial role of endogenous MT-3 in protecting from the AD etiology. In vivo studies, investigating the function of MT-3 in double transgenic mice presenting AD pathology (Tg2576 mouse AD model), support this role [49]. In a mouse AD model, APP increases death rate and affects behavioral phenotype. The absence of MT-3 partially reversed the APP-induced mortality and changes in the behavior of females. Furthermore, the amyloid plaques formation and/or APP expression in the cortex and hippocampus of MT-3 deficient females were decreased. Interestingly, MT-3 absence, on the one hand, substantially decreased APP in brain homogenates from female mice and, on the other hand, shows a tendency to increase copper levels in both genders [49].
In the later studies, the protective effect of the human Zn7MT-2A isoform against Cu(II)–Aβ1–40 toxicity was also investigated and compared with that of Zn7MT-3 [50]. The studies showed that MT-2A can protect against copper-induced Aβ aggregation and neurotoxicity. However, in contrast to the original studies employing Zn7MT-3, in which the formation of the air stable Cu(I)4Zn4MT-3 species was found crucial for the protective effect [46], the underlying reaction between Cu(II)–Aβ1–40 and Zn7MT-2A has been interpreted using the chemical characteristics of the fully reduced Cu(I)10MT-2A and Cu(I)12MT-3 species. However, as the reduction of Cu(II) in Cu(II)–Aβ1–40 to Cu(I) is accomplished by the thiolate ligands of Zn7MTs, the formation of fully reduced Cu(I)10MT-2A and Cu(I)12MT-3 in the oxidizing extracellular environment is unlikely. This conclusion is supported by the mass spectrometric demonstration that the cysteine thiolates in Zn7MT-3 can reduce up to 8 Cu(II) [30] concomitant with the formation of intramolecular disulfides. Thus, while the stepwise reduction of the first 4 Cu(II) equivalents confirmed the cooperative formation of the air-stable Cu(I)4Zn4MT-3 species, the reductions of the following Cu(II) equivalents resulted in the simultaneous presence of a number of air-sensitive Cu(I)MT-3 species [30]. These studies signify the importance of Cu(I)4Zn4MT-3 for its protective effect in AD. By contrast, in the studies of the interaction between Cu(II)–Aβ1–40 and Zn7MT-2A, the direct metal swap between Zn(II), coordinated by thiolate ligands in Zn7MT-2A and Cu(II), coordinated by nitrogen and oxygen ligands in Cu(II)–Aβ1–40, has been implicated and the importance of metal affinities for the protective effect stressed [50]. However, the assumed direct metal swap between Cu(II)–Aβ1–40 and Zn7MT-2A cannot occur without the preceding Cu(II) reduction to Cu(I) by cysteine ligands. Accordingly, the differences in affinities are not essential for the protective effect, but rather the long-term stability of partially oxidized Cu(I),ZnMT-2A formed upon Cu(II) reduction and its removal from Cu(II)–Aβ1–40 in the oxidizing extracellular space. Thus, to gain an insight into the protective effect of Zn7MT-2A the redox stability of different Cu(I)x, ZnyMT-2A species, generated upon aerobic Cu(II) reduction, should be addressed.
The amyloid hypothesis in AD considers the Aβ production and deposition as the trigger of this neurodegenerative disorder [51]. So far, the protective effect of Zn7MT-3 from the toxicity of the Cu(II) containing Aβ1–40 and Aβ1–42 peptides has been described [46]. However, the proteolytic cleavage of APP by β- and γ-secretase yields several other isoforms of the Aβ peptides. β-Secretase, the β-site APP-cleaving enzyme 1 (BACE1), is an aspartyl protease [52,53,54], while γ-secretase is a multiprotein complex consisting of at least four essential components (presenilin, nicastrin, Aph-1, and Pen-2) necessary for full enzymatic activity [55]. The APP cleavage by β- and γ-secretase gives rise to Aβ1–42 and Aβ isoforms ranging from Aβ1–40 down to Aβ1–17 [56]. In addition, β-secretase cleavage together with α-secretase cleavage yield several short Aβ peptides [55]. In this context, the increased concentrations of Aβ1–16 have been found in sporadic AD (SAD) and familial AD (FAD) compared to non-demented controls.
Apart from Aβ with aspartate (Asp1), as the first amino acid in Aβ1−x, several N-truncated and modified Aβ peptides have been found in the brains of AD patients. However, to extract and identify the exact levels of the various N-truncated Aβ variants in post-mortem brains is difficult as many factors—such as antibody specificities, extraction protocols and brain areas studied—can influence an analysis [57]. Nevertheless, there is general agreement that plaque-born peptides harbor high amounts of N-truncated Aβ, especially that starting with Phe4. As revealed by detailed analyses of Aβ isoform pattern in the cerebellum, cortex, and hippocampus of FAD subjects, SAD subjects and non-demented controls, the dominating Aβ isoforms in AD brains are Aβ4–42, Aβ1–40, and Aβ1–42 [58,59]. The first three amino acids (Phe-Arg-His) of N-truncated Aβ4–42 give rise to the H2N-Xaa–Yaa–His– motif which enables a high affinity Cu(II) binding [60], already reported in human serum albumin (HAS) [61]. Recently, the question regarding the Aβ4–42 affinity toward Cu(II) ions has been addressed [62]. By using the Aβ4–16 peptide as a model to fully characterize the Cu(II) binding properties, it was demonstrated that the Phe-Arg-His sequence of Aβ4–16 stoichiometrically binds Cu(II) with a conditional Kd value of 30 fM at pH 7.4 (a conditional Kd is defined as the Kd at pH 7.4). The determined affinity is more than three orders of magnitude higher than that reported for Aβ1−x peptides (conditional Kd ∼1 × 10−10 M) [63]. Consequently, Aβ4–16 can compete successfully with Aβ1−x peptides for Cu(II) ions. Furthermore, Aβ4–40/42 forms fibers twice as fast as Aβ1–42 with a very different morphology, forming bundles of very short amyloid rods. The generation of ROS by Cu(II)–Aβ4–16, monitoring the hydroxyl radical production, revealed that the N-truncated Cu(II)–Aβ4–16 and Cu(II)–Aβ4–42 species exhibit low levels of hydroxyl radical production compared to Cu(II)–Aβ1–42. The results are in agreement with voltammetric experiments in which low level of redox activity of the Cu(II)–Aβ4–16 complex was determined. Thus, Cu(II) in Cu(II)–Aβ4–16 could be oxidized irreversibly to Cu(III), but could not be reduced to Cu(I) [62].
In view of the latter studies, the question has been addressed whether the N-terminal Cu(II)–Aβ4–16 complex is fully resistant to the copper/zinc swap with Zn7MT-3 [64]. In marked contrast to Cu(II)–Aβ1–16, no copper can be extracted from the N-truncated Cu(II)–Aβ4–16 complex by Zn7MT-3. A reverse swap experiment, where apo–Aβ4–16 was added to the preformed Cu(I)/Zn(II)MT-3 complex, was also conducted. Here the absence of Cu(I) transfer from MT-3 to the peptide reflects different affinities of thiolate vs. oxygen and nitrogen ligands for Cu(I) ions. Conversely, addition of Cu(II) ions to a mixture of Zn7MT-3 and apo–Aβ4–16 resulted in the partition of copper between these biomolecules. From these studies, it has been concluded that Aβ4–42 and Zn7MT-3 may play parallel roles in synaptic copper clearance in different synaptic settings [64].
Other naturally N-terminally truncated Aβ possessing the copper binding motif H2N-Xaa–Yaa–His– is the Aβ11–40/42 peptide which starts with the residue Glu11. This peptide is found in the cerebrospinal fluid and has a similar abundance to Aβ1–42, constituting one-fifth of the plaque load [65]. A recent publication reported for Cu(II)–Aβ11–42 Kd of a 34 fM at pH 7.4 [66], which is similar to 30 fM reported for Cu(II)–Aβ4–42. In this work, no studies regarding the redox stability of the Cu(II)–Aβ11–42 complex and the peptide reactivity with Cu(II)–Aβ1–x or Zn7MT-3 have been conducted [66]. Nevertheless, the N-truncated Aβ11–42 peptide may show a similar reactivity to that reported for to the Aβ4–42 peptide (see above). However, although Aβ11–42 species with an intact N terminus could be detected, the majority of peptides truncated at the N-terminal residue Glu11 are cyclized to pyroglutamate to generate pGlu-Aβ11–42 [67,68]. This peptide modification blocks the Cu(II) coordination site.
Apart from the discovered protective effect of MT-3 in AD, its role in other metal-linked neurodegenerative disorders like PD and prion disease has also been investigated. Similarly to AD, the aberrant binding of Cu(II) to the amylogenic protein α-synuclein (α-Syn) in PD [69,70,71] and the prion protein in CJD [2] has deleterious effects in these diseases. These include the Cu(II)-mediated ROS production and contribution to the formation of oligomers and fibrils. PD is characterized by the loss of dopaminergic neurons and the formation of intracellular inclusions known as Lewy bodies, composed mainly of aggregated α-Syn in neurons of substantia nigra pars compacta [72]. The formation of Cu(II)–α-Syn contributes to the progression of PD. In the pathogenesis of prion diseases, the structural conversion of a natively folded prion protein (PrPC) [73], into a misfolded (PrPSc) is the key molecular event [2,74]. While PrPC is mainly a helical monomeric protein, PrPSc is oligomeric and rich in β-sheet structure [2]. The binding of up to six Cu(II) to the histidine residues present in the disordered part of PrPC structure plays an important role in its conversion to PrPSc and thus the disease progression. The studies of the interaction between Zn7MT-3 and Cu(II)-containing α-Syn and prion protein in vitro revealed that Zn7MT-3, through copper subtraction and the formation of air-stable Cu(I)4Zn4MT-3, plays a protective role [75,76]. Taken together, as the nature of the Cu(II) binding sites in Aβ, α-Syn, and prion proteins substantially differ, the observed protective effect of Zn7MT-3 against Cu(II) toxicity suggests that the protein may play a general protective role in metal-linked neurodegenerative disorders (Figure 2).
4. Toxicological and Neuropathological Aspects of Lead (Pb) Toxicity in Relation to Mammalian Metallothioneins
The biosynthesis of most predominant mammalian isoforms MT-1/-2, but not of MT-3 is upregulated in response to different stimuli, especially heavy metals, but also oxidative stress or stress hormones (glucocorticoids). High cysteine content of MT-1/-2 (ca. 30%) confers them with a high capacity to bind essential (Zn(II) and Cu(I)) and toxic (Cd(II), Ag(I), and Hg(II)) heavy metal ions in vivo and in vitro. The exposure of humans or experimental animals to these metal ions lead to an induction of MT-1/-2 biosynthesis and the formation of MT-1/-2 metalloforms in which the corresponding metal ion is bound. The mechanism of the metal-mediated induction of MT-1/-2 biosynthesis is discussed below. The role of MT-1/-2 in the heavy metal detoxification has been established first for Cd(II) [78,79,80,81,82]. In recent years, there is an increased interest in understanding of the toxicity of lead (Pb), a ubiquitous naturally occurring toxic metal. Pb-induced toxic effects can manifest in several organs but the brain and kidney are primary targets. Compelling evidence is emerging that Pb exposure, particularly in early-life, may cause neurodegeneration in later life stages. The sporadic nature of AD argues for an environmental link that may drive AD pathogenesis. In rodents, exposure to Pb during brain development predetermined the expression and regulation of the APP, its amyloidogenic Aβ product and other AD-related genes in old age [83]. Life time whole-body occupational Pb exposure has been recently shown to be a risk factor for PD [84]. Since the formation of Lewy bodies in PD required MT-1/-2, the interaction between α-Syn and MT-1/-2 has been established using MT-null and wild-type cell lines [85]. A direct interaction between α-Syn and MT-1/-2 was confirmed by antibody pull-down assay [86]. However, a similar study using the MT-3 isoform has not been undertaken.
By analogy to the MT-1/-2 induction by Cd(II), the formation of mammalian Pb(II)MT-1/-2 in a similar detoxification process has been anticipated. However, so far no Pb(II)-containing MTs have been isolated and characterized from natural sources. Nevertheless, the in vitro characterization of the Pb(II)-containing MT-1/-2 and MT-3 isoforms has been reported [87,88]. Prior to the discussion of the structural features of these Pb(II)-containing MTs, the current knowledge regarding the nature of low molecular weight Pb(II)-binding proteins, obtained upon the Pb(II) administration to experimental animals, will be briefly discussed. Apart from these proteins, the heme pathway enzyme δ-aminolevulinic acid dehydratase (ALAD) isoforms (240–280 kDa proteins), have attracted the greatest attention as the enzymes are both inhibited and induced by lead (reviewed in [89]).
From a number of studies, the nature of low molecular weight proteins appears to be uncertain as the occurrence of either low molecular weight lead-binding proteins or MT-like proteins have been reported [89]. At this point, it should be noted that in all instances, no full characterization of these proteins has been carried out. Lead-binding proteins are a series of low molecular weight proteins (10 to 12 kDa) which have the capacity to bind and sequester lead in a nontoxic form in several organs (kidney, brain, lung, liver, and erythrocyte). These proteins are rich in aspartate and glutamate amino acids and possess a dissociation constant Kd for Pb(II) on the order of 10−8 M and appear to normally bind zinc [90]. The occurrence of low molecular weight Pb(II)-binding protein(s) with a large percentage of cysteine and a greater UV absorbance at 254 than at 280 nm, features diagnostic of a MT, have also been reported [91]. In this regard, it has been shown that depending on the conditions of Pb(II) administration to experimental animals the gel filtration fractions containing only ZnMT, both PbMT and ZnMT metalloforms, or the metal mixed Pb,ZnMT form have been identified [89]. These observations can be explain, in part, by the underlying mechanism of the detoxification process described for Cd(II) [92]. A hallmark of the promoters/enhancers of most MT genes are short DNA sequence motifs termed metal response elements (MREs); many also harbor antioxidant response elements (AREs) and glucocorticoid response elements (GREs) [93]. Transcriptional induction of MT-1/-2 genes is mediated by the metal-regulatory transcription factor 1 (MTF-1) [92,94]. This protein is essential not only for zinc-induced transcription of MT-1/-2 genes, but also for their induction by other heavy metals like cadmium and copper and/or oxidative stress [92]. In the induction process, MTF-1, upon activation with zinc displaced from zinc-saturated Zn7MT-1/-2 in the cytosol by cadmium or copper binding, translocate from the cytosol to the nucleus where it binds to promoter proximal MREs [94,95]. Lead, similarly to cadmium and copper, will also displace Zn(II) from Zn7MT-1/-2 and activate MTF-1. Consequently, depending on the experimental conditions the newly synthetized thionein (the metal-free form of MT-1/-2) may then bind with different metal ions reflecting the concentration of buffered metals available at folding (see above). It may be noted that although similar MREs and other DNA sequence motifs were also identified in the MT-3 gene, the MT-3 biosynthesis was unresponsive to these inducers [39].
The spectroscopic/biochemical studies on the in vitro interaction of Pb(II) with MT-2 and MT-3 have been carried out and the Pb(II)-MT forms characterized [87,88]. In the structure of MII7MTs, the divalent metal ions Zn(II) and/or Cd(II) are tetrahedrally coordinated by four cysteine sulfurs. Recent structural studies aimed at the understanding of Pb(II) binding to thiol-rich structural proteins containing zinc-binding sites. The studies showed that although zinc is bound tetrahedrally, Pb(II) shows a clear preference for a three-coordinate Pb(II)-S3 mode. In the latter structure, Pb(II) is coordinated by three sulfurs in a trigonal pyramidal geometry with the Pb 6s2 lone-pair electrons occupying the axial position [96]. It has been concluded, moreover, that Pb(II) assumes a preferred coordination mode of Pb-S3 even when an additional thiolate ligand is available for binding. The interaction of Pb(II) with MT-2 has been studied by combination of spectroscopic and computational methods [87]. By contrast to Zn(II) and Cd(II), the binding of Pb(II) results in the formation of two different Pb7MT-2 complexes depending on the pH employed. The first complex, designated Pb7MT-2(I), is formed under neutral conditions and the second, Pb7MT-2(II), under weakly acidic (pH 4.5) conditions, The characterization of Pb7MT-2(I) revealed that all seven Pb(II) ions are trigonally coordinated by three cysteine sulfurs (Pb–S3 mode) and that no metal-ligand ring in the α- or β-domain is present. The marked structural differences to the cluster structure in Zn(II)- and Cd(II)-containing MII7MTs results from the Pb 6s2 lone-pair electrons, which disrupt the tetrahedral coordination by occupying the axial position. In the case of Pb7MT-2(II), completely different binding modes from Pb7MT-2(I) exist with all cysteine ligands coordinating in the terminal form. These binding modes include the trigonal pyramidal Pb–S3 mode, the distorted trigonal pyramidal Pb–S2O1 mode in the α-domain, and the distorted quadrilateral pyramidal Pb–S3O1 in the β-domain.
The binding of Pb(II) and Zn(II) to MT-3 has been studied by isothermal titration calorimetry (ITC), a technique capable of measuring the thermodynamics of metal binding [88]. Anaerobic ITC measurements of Pb(II) displacing Zn(II) from Zn7MT-3 and experiments in which EDTA was used to chelate metal ions from both Pb7MT-3 and Zn7MT-3 revealed that (i) MT-3 binds Pb(II) with a higher affinity than it does Zn(II) and that Pb(II) displaces bound Zn(II) in Zn7MT-3 and that (ii) EDTA extracted both metal ions from Zn7MT-3 and Pb7MT-3 in a tri-phasic process, indicating that they bind to the protein in three populations with different binding thermodynamics. These data indicate that Pb(II) binding to both apoMT-3 and Zn7MT-3 is thermodynamically favorable [88]. Overall, despite the number of toxicological studies over many years, the in vivo interaction of MTs with Pb(II) is not entirely clear.
5. Concluding Remarks
Since the initial discovery of MT-3 in normal human brains by Uchida et al. 1991 [4], the functional role of this metalloprotein has been investigated intensively. MT-3 is involved in homeostasis of essential transition metals zinc and copper in the brain. The amyloid-forming diseases such as AD, PD, ALS, and prion (CJD) represent a group of neurodegenerative disorders that affect both animals and humans. In these diseases, the role of metal ions, especially copper and zinc, is the subject of intense research. Their involvement in protein misfolding, aggregation and in the generation of reactive oxygen species have been shown. The structural and biological studies also show that these metal ions not only bind to the disease specific amyloidogenic peptides or proteins with high affinity, but also modify their biochemical properties, making them important players in the progression of the disease. The protective effect of MT-3 from the copper toxicity has been demonstrated for AD, PD, and prion diseases (see above). The neuroprotective effect of MT-3 in the pathogenesis of ALS has also been established. In this work, MT-3 prevents the loss of motor neurons of ALS model mice and prolongs the life span, even when the administration is started at the time of onset [97]. Furthermore, recent biological data reveal that MT-3 contributes to neuronal and astrocytic cell death through zinc release [98] and that its interaction with actin modulates c-Abl signaling in astrocytes; c-Abl is a member of the nonreceptor tyrosine kinase family [99]. In addition, the observed developed obesity in male MT3-null mice is due likely to abnormal leptin signaling in the hypothalamus [100]. Apart from the CNS, the MT-3 expression is also observed in a few non-CNS areas, including the pancreas [101], prostate, testis, and tongue [102]. In this regard, more recent studies on the zinc-rich insulin producing islet β-cells suggest a role for MT-3 in streptozotocin (STZ)-induced islet cell death and consequent hyperglycemia [103]. Thus, it has been demonstrated that MT-3 may through PDE3a (phosphodiesterase 3A) play a key role in zinc dyshomeostasis and cell death in STZ-treated islets. Overall, the list of MT-3 functions, suggesting its role not only in the CNS, but also outside of this organ, is steadily increasing.
Acknowledgments
Gabriele Meloni is supported by start-up funds from the University of Texas at Dallas and by the Robert A. Welch Foundation.
Author Contributions
Milan Vašák and Gabriele Meloni wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Aβ | Amyloid-beta |
| AD | Alzheimer’s Disease |
| ALAD | δ-aminolevulinic acid dehydratase |
| ALS | Amyotrophic lateral sclerosis |
| APP | Amyloid precursor protein |
| AREs | Antioxidant response elements |
| BACE | Beta-secretase |
| CJD | Creutzfeldt–Jakob disease |
| CNS | Central nervous system |
| EXAFS | Extended X-ray absorption fine structure |
| FAD | Familiar Alzheimer’s Disease |
| GREs | Glucocorticoid response elements |
| ITC | Isothermal Titration Calorimetry |
| MRE | Metal response elements |
| MT | Metallothionein |
| MTF-1 | Metal-regulatory transcription factor 1 |
| PD | Parkinson’s Disease |
| PrP | Prion Protein |
| PSEN1 | Presenilin 1 |
| PSEN2 | Presenilin 1 |
| SAD | Sporadic Alzheimer’s Disease |
| SOD | Superoxide Dismutase |
| α-Syn | Alpha-Synuclein |
References
- Mocchegiani, E.; Malavolta, M.; Costarelli, L.; Giacconi, R.; Cipriano, C.; Piacenza, F.; Tesei, S.; Basso, A.; Pierpaoli, S.; Lattanzio, F. Zinc, metallothioneins and immunosenescence. Proc. Nutr. Soc. 2010, 69, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Gaggelli, E.; Kozlowski, H.; Valensin, D.; Valensin, G. Copper homeostasis and neurodegenerative disorders (Alzheimer’s, prion, and Parkinson’s diseases and amyotrophic lateral sclerosis). Chem. Rev. 2006, 106, 1995–2044. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug. Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Takio, K.; Titani, K.; Ihara, Y.; Tomonaga, M. The growth inhibitory factor that is deficient in the Alzheimer’s disease brain is a 68 amino acid metallothionein-like protein. Neuron 1991, 7, 337–347. [Google Scholar] [CrossRef]
- Masters, B.A.; Quaife, C.J.; Erickson, J.C.; Kelly, E.J.; Froelick, G.J.; Zambrowicz, B.P.; Brinster, R.L.; Palmiter, R.D. Metallothionein III is expressed in neurons that sequester zinc in synaptic vesicles. J. Neurosci. 1994, 14, 5844–5857. [Google Scholar] [PubMed]
- Miles, A.T.; Hawksworth, G.M.; Beattie, J.H.; Rodilla, V. Induction, regulation, degradation, and biological significance of mammalian metallothioneins. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 35–70. [Google Scholar] [CrossRef] [PubMed]
- Tanji, K.; Irie, Y.; Uchida, Y.; Mori, F.; Satoh, K.; Mizushima, Y.; Wakabayashi, K. Expression of metallothionein-III induced by hypoxia attenuates hypoxia-induced cell death in vitro. Brain Res. 2003, 976, 125–129. [Google Scholar] [CrossRef]
- Wang, B.; Wood, I.S.; Trayhurn, P. PCR arrays identify metallothionein-3 as a highly hypoxia-inducible gene in human adipocytes. Biochem. Biophys. Res. Commun. 2008, 368, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D. Constitutive expression of metallothionein-III (MT-III), but not MT-I, inhibits growth when cells become zinc deficient. Toxicol. Appl. Pharmacol. 1995, 135, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Irie, Y.; Keung, W.M. Anti-amyloid beta activity of metallothionein-III is different from its neuronal growth inhibitory activity: Structure-activity studies. Brain Res. 2003, 960, 228–234. [Google Scholar] [CrossRef]
- Sewell, A.K.; Jensen, L.T.; Erickson, J.C.; Palmiter, R.D.; Winge, D.R. Bioactivity of metallothionein-3 correlates with its novel beta domain sequence rather than metal binding properties. Biochemistry 1995, 34, 4740–4747. [Google Scholar] [CrossRef] [PubMed]
- Atrian, S.; Capdevila, M. Metallothionein-protein interactions. Biomol. Concepts 2013, 4, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Ngu, T.T.; Chan, J.; Salgado, M.T.; Merrifield, M.E.; Stillman, M.J. Peptide folding, metal-binding mechanisms, and binding site structures in metallothioneins. Exp. Biol. Med. 2006, 231, 1488–1499. [Google Scholar] [CrossRef]
- Hidalgo, J.; Aschner, M.; Zatta, P.; Vašák, M. Roles of the metallothionein family of proteins in the central nervous system. Brain Res. Bull. 2001, 55, 133–145. [Google Scholar] [CrossRef]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system: Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Krezel, A.; Hao, Q.; Maret, W. The zinc/thiolate redox biochemistry of metallothionein and the control of zinc ion fluctuations in cell signaling. Arch. Biochem. Biophys. 2007, 463, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Henkel, G.; Krebs, B. Metallothioneins: Zinc, cadmium, mercury, and copper thiolates and selenolates mimicking protein active site features—Structural aspects and biological implications. Chem. Rev. 2004, 104, 801–824. [Google Scholar] [CrossRef] [PubMed]
- Vašák, M.; Hasler, D.W. Metallothioneins: New functional and structural insights. Curr. Opin. Chem. Biol. 2000, 4, 177–183. [Google Scholar] [CrossRef]
- Vašák, M.; Romero-Isart, N. Metallothioneins. In Encyclopedia of Inorganic Chemistry, 2nd ed.; King, R.B., Ed.; J. Wiley & Sons Ltd.: New York, NY, USA, 2005; pp. 3208–3221. [Google Scholar]
- Faller, P. Neuronal growth-inhibitory factor (metallothionein-3): Reactivity and structure of metal-thiolate clusters. FEBS J. 2010, 277, 2921–2930. [Google Scholar] [CrossRef] [PubMed]
- Vašák, M.; Meloni, G. Chemistry and biology of mammalian metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Bruinink, A.; Faller, P.; Sidler, C.; Bogumil, R.; Vašák, M. Growth inhibitory factor and zinc affect neural cell cultures in a tissue specific manner. Chem. Biol. Interact. 1998, 115, 167–174. [Google Scholar] [CrossRef]
- Faller, P.; Vašák, M. Distinct metal-thiolate clusters in the N-terminal domain of neuronal growth inhibitory factor. Biochemistry 1997, 36, 13341–13348. [Google Scholar] [CrossRef] [PubMed]
- Hasler, D.W.; Faller, P.; Vašák, M. Metal-thiolate clusters in the C-terminal domain of human neuronal growth inhibitory factor (GIF). Biochemistry 1998, 37, 14966–14973. [Google Scholar] [CrossRef] [PubMed]
- Faller, P.; Hasler, D.W.; Zerbe, O.; Klauser, S.; Winge, D.R.; Vašák, M. Evidence for a dynamic structure of human neuronal growth inhibitory factor and for major rearrangements of its metal-thiolate clusters. Biochemistry 1999, 38, 10158–10167. [Google Scholar] [CrossRef] [PubMed]
- Oz, G.; Zangger, K.; Armitage, I.M. Three-dimensional structure and dynamics of a brain specific growth inhibitory factor: Metallothionein-3. Biochemistry 2001, 40, 11433–11441. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Q.; Cai, B.; Li, H.; Sze, K.H.; Huang, Z.X.; Wu, H.M.; Sun, H. Solution structure and dynamics of human metallothionein-3 (MT-3). FEBS Lett. 2006, 580, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Roschitzki, B.; Vašák, M. A distinct Cu4-thiolate cluster of human metallothionein-3 is located in the N-terminal domain. J. Biol. Inorg. Chem. 2002, 7, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Meloni, G.; Faller, P.; Vasak, M. Redox silencing of copper in metal-linked neurodegenerative disorders: Reaction of Zn7metallothionein-3 with Cu2+ ions. J. Biol. Chem. 2007, 282, 16068–16078. [Google Scholar] [CrossRef] [PubMed]
- Bogumil, R.; Faller, P.; Pountney, D.L.; Vašák, M. Evidence for Cu(I) clusters and Zn(II) clusters in neuronal growth-inhibitory factor isolated from bovine brain. Eur. J. Biochem. 1996, 238, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Bogumil, R.; Faller, P.; Binz, P.A.; Vašák, M.; Charnock, J.M.; Garner, C.D. Structural characterization of Cu(I) and Zn(II) sites in neuronal-growth-inhibitory factor by extended X-ray absorption fine structure (EXAFS). Eur. J. Biochem. 1998, 255, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Roschitzki, B.; Vašák, M. Redox labile site in a Zn4 cluster of Cu4,Zn4-metallothionein-3. Biochemistry 2003, 42, 9822–9828. [Google Scholar] [CrossRef] [PubMed]
- Palacios, O.; Atrian, S.; Capdevila, M. Zn- and Cu-thioneins: A functional classification for metallothioneins? J. Biol. Inorg. Chem. 2011, 16, 991–1009. [Google Scholar] [CrossRef] [PubMed]
- Artells, E.; Palacios, O.; Capdevila, M.; Atrian, S. In vivo-folded metal-metallothionein 3 complexes reveal the Cu-thionein rather than Zn-thionein character of this brain-specific mammalian metallothionein. FEBS J. 2014, 281, 1659–1678. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.W.; Osman, D.; Robinson, N.J. Metal preferences and metallation. J. Biol. Chem. 2014, 289, 28095–28103. [Google Scholar] [CrossRef] [PubMed]
- Dudev, T.; Lim, C. Metal binding affinity and selectivity in metalloproteins: Insights from computational studies. Annu. Rev. Biophys. 2008, 37, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Jung, H.; Meloni, G. Copper metallothioneins. IUBMB Life 2017, 69, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, C.A.; Asanuma, M.; Sogawa, N.; Miyazaki, I.; Nakanishi, T.; Furuta, H.; Ogawa, N. Localization, regulation, and function of metallothionein-III/growth inhibitory factor in the brain. Acta Med. Okayama 2001, 55, 1–9. [Google Scholar] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Dickson, D.W. The pathogenesis of senile plaques. J. Neuropathol. Exp. Neurol. 1997, 56, 321–339. [Google Scholar] [CrossRef] [PubMed]
- Glenner, G.G.; Wong, C.W. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–990. [Google Scholar] [CrossRef]
- Faller, P.; Hureau, C.; La Penna, G. Metal ions and intrinsically disordered proteins and peptides: From Cu/Zn amyloid-β to general principles. Acc. Chem. Res. 2014, 47, 2252–2259. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Irie, Y.; Keung, W.M. Metallothionein-III antagonizes the neurotoxic and neurotrophic effects of amyloid beta peptides. Biochem. Biophys. Res. Commun. 2001, 282, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Meloni, G.; Sonois, V.; Delaine, T.; Guilloreau, L.; Gillet, A.; Teissie, J.; Faller, P.; Vašák, M. Metal swap between Zn7-metallothionein-3 and amyloid-β-Cu protects against amyloid-β toxicity. Nat. Chem. Biol. 2008, 4, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, J.T.; Hureau, C.; Hemmingsen, L.; Heegaard, N.H.; Ostergaard, J.; Vašák, M.; Faller, P. Rapid exchange of metal between Zn7-metallothionein-3 and amyloid-β peptide promotes amyloid-related structural changes. Biochemistry 2012, 51, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.; Meloni, G.; Talmard, C.; Vašák, M.; Faller, P. Zinc release of Zn7-metallothionein-3 induces fibrillar type amyloid-beta aggregates. Metallomics 2010, 2, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Manso, Y.; Carrasco, J.; Comes, G.; Meloni, G.; Adlard, P.A.; Bush, A.I.; Vašák, M.; Hidalgo, J. Characterization of the role of metallothionein-3 in an animal model of Alzheimer’s disease. Cell. Mol. Life Sci. 2012, 69, 3683–3700. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.S.; Howells, C.; Eaton, E.D.; Shabala, L.; Zovo, K.; Palumaa, P.; Sillard, R.; Woodhouse, A.; Bennett, W.R.; Ray, S.; et al. The native copper- and zinc-binding protein metallothionein blocks copper-mediated Abeta aggregation and toxicity in rat cortical neurons. PLoS ONE 2010, 5, e12030. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Powell, D.; Howlett, D.R.; Tew, D.G.; Meek, T.D.; Chapman, C.; Gloger, I.S.; Murphy, K.E.; Southan, C.D.; Ryan, D.M.; et al. Identification of a novel aspartic protease (Asp 2) as β-secretase. Mol. Cell. Neurosci. 1999, 14, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Anderson, J.P.; Barbour, R.; Basi, G.S.; Caccavello, R.; Davis, D.; Doan, M.; Dovey, H.F.; Frigon, N.; Hong, J.; et al. Purification and cloning of amyloid precursor protein β-secretase from human brain. Nature 1999, 402, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Bienkowski, M.J.; Shuck, M.E.; Miao, H.; Tory, M.C.; Pauley, A.M.; Brashier, J.R.; Stratman, N.C.; Mathews, W.R.; Buhl, A.E.; et al. Membrane-anchored aspartyl protease with Alzheimer’s disease β-secretase activity. Nature 1999, 402, 533–537. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, B. Aph-1, Pen-2, and Nicastrin with Presenilin generate an active γ-Secretase complex. Neuron 2003, 38, 9–12. [Google Scholar] [CrossRef]
- Andreasson, U.; Portelius, E.; Andersson, M.E.; Blennow, K.; Zetterberg, H. Aspects of β-amyloid as a biomarker for Alzheimer’s disease. Biomark. Med. 2007, 1, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Takio, K.; Ogawara, M.; Selkoe, D.J. Mass spectrometry of purified amyloid β protein in Alzheimer’s disease. J. Biol. Chem. 1992, 267, 17082–17086. [Google Scholar] [PubMed]
- Moore, B.D.; Chakrabarty, P.; Levites, Y.; Kukar, T.L.; Baine, A.M.; Moroni, T.; Ladd, T.B.; Das, P.; Dickson, D.W.; Golde, T.E. Overlapping profiles of Aβ peptides in the Alzheimer’s disease and pathological aging brains. Alzheimers Res. Ther. 2012, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Portelius, E.; Bogdanovic, N.; Gustavsson, M.K.; Volkmann, I.; Brinkmalm, G.; Zetterberg, H.; Winblad, B.; Blennow, K. Mass spectrometric characterization of brain amyloid β isoform signatures in familial and sporadic Alzheimer’s disease. Acta Neuropathol. 2010, 120, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Drew, S.C.; Noble, C.J.; Masters, C.L.; Hanson, G.R.; Barnham, K.J. Pleomorphic copper coordination by Alzheimer’s disease amyloid-β peptide. J. Am. Chem. Soc. 2009, 131, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Rozga, M.; Sokolowska, M.; Protas, A.M.; Bal, W. Human serum albumin coordinates Cu(II) at its N-terminal binding site with 1 pM affinity. J. Biol. Inorg. Chem. 2007, 12, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Mital, M.; Wezynfeld, N.E.; Fraczyk, T.; Wiloch, M.Z.; Wawrzyniak, U.E.; Bonna, A.; Tumpach, C.; Barnham, K.J.; Haigh, C.L.; Bal, W.; et al. A Functional Role for Abeta in Metal Homeostasis? N-Truncation and High-Affinity Copper Binding. Angew. Chem. Int. Ed. Engl. 2015, 54, 10460–10464. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.N.; Nicklin, H.G. A possible role for the species peroxonitrite in nitrification. Biochim. Biophys. Acta 1970, 222, 660–661. [Google Scholar] [CrossRef]
- Wezynfeld, N.E.; Stefaniak, E.; Stachucy, K.; Drozd, A.; Plonka, D.; Drew, S.C.; Krezel, A.; Bal, W. Resistance of Cu(Aβ4–16) to Copper Capture by Metallothionein-3 Supports a Function for the Aβ4–42 Peptide as a Synaptic Cu(II) Scavenger. Angew. Chem. Int. Ed. Engl. 2016, 55, 8235–8238. [Google Scholar] [CrossRef] [PubMed]
- Huse, J.T.; Liu, K.; Pijak, D.S.; Carlin, D.; Lee, V.M.; Doms, R.W. β-secretase processing in the trans-Golgi network preferentially generates truncated amyloid species that accumulate in Alzheimer’s disease brain. J. Biol. Chem. 2002, 277, 16278–16284. [Google Scholar] [CrossRef] [PubMed]
- Barritt, J.D.; Viles, J.H. Truncated Amyloid-β(11–40/42) from Alzheimer Disease Binds Cu2+ with a Femtomolar Affinity and Influences Fiber Assembly. J. Biol. Chem. 2015, 290, 27791–27802. [Google Scholar] [PubMed]
- Gunn, A.P.; Masters, C.L.; Cherny, R.A. Pyroglutamate-Abeta: Role in the natural history of Alzheimer’s disease. Int. J. Biochem. Cell Biol. 2010, 42, 1915–1918. [Google Scholar] [CrossRef] [PubMed]
- Naslund, J.; Schierhorn, A.; Hellman, U.; Lannfelt, L.; Roses, A.D.; Tjernberg, L.O.; Silberring, J.; Gandy, S.E.; Winblad, B.; Greengard, P.; et al. Relative abundance of Alzheimer Aβ amyloid peptide variants in Alzheimer disease and normal aging. Proc. Natl. Acad. Sci. USA 1994, 91, 8378–8382. [Google Scholar] [CrossRef] [PubMed]
- Binolfi, A.; Lamberto, G.R.; Duran, R.; Quintanar, L.; Bertoncini, C.W.; Souza, J.M.; Cervenansky, C.; Zweckstetter, M.; Griesinger, C.; Fernandez, C.O. Site-specific interactions of Cu(II) with α and β-synuclein: Bridging the molecular gap between metal binding and aggregation. J. Am. Chem. Soc. 2008, 130, 11801–11812. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.R.; Shin, H.J.; Lee, J.H. Metal-catalyzed oxidation of α-synuclein in the presence of Copper(II) and hydrogen peroxide. Arch. Biochem. Biophys. 2000, 378, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Drew, S.C.; Leong, S.L.; Pham, C.L.; Tew, D.J.; Masters, C.L.; Miles, L.A.; Cappai, R.; Barnham, K.J. Cu2+ binding modes of recombinant α-synuclein-insights from EPR spectroscopy. J. Am. Chem. Soc. 2008, 130, 7766–7773. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.M.; Dawson, V.L. Molecular pathways of neurodegeneration in Parkinson’s disease. Science 2003, 302, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Aguzzi, A.; Heikenwalder, M. Pathogenesis of prion diseases: Current status and future outlook. Nat. Rev. Microbiol. 2006, 4, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S.B. Molecular biology of prion diseases. Science 1991, 252, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Meloni, G.; Vašák, M. Redox activity of α-synuclein-Cu is silenced by Zn7-metallothionein-3. Free Radic. Biol. Med. 2011, 50, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Meloni, G.; Crameri, A.; Fritz, G.; Davies, P.; Brown, D.R.; Kroneck, P.M.; Vašák, M. The catalytic redox activity of prion protein-Cu(II) is controlled by metal exchange with the Zn(II)-thiolate clusters of Zn7metallothionein-3. ChemBioChem 2012, 13, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Braun, W.; Vašák, M.; Robbins, A.H.; Stout, C.D.; Wagner, G.; Kägi, J.H.; Wüthrich, K. Comparison of the NMR solution structure and the X-ray crystal structure of rat metallothionein-2. Proc. Natl. Acad. Sci. USA 1992, 89, 10124–10128. [Google Scholar] [CrossRef] [PubMed]
- Piscator, M. On Cadmium in Normal Human Kidneys with a Report on the Isolation of Metallothioneine from Cadmium-Exposed Rabbit Livers. Nord. Hyg. Tidskr. 1964, 45, 76–82. [Google Scholar] [PubMed]
- Michalska, A.E.; Choo, K.H. Targeting and germ-line transmission of a null mutation at the metallothionein I and II loci in mouse. Proc. Natl. Acad. Sci. USA 1993, 90, 8088–8092. [Google Scholar] [CrossRef] [PubMed]
- Masters, B.A.; Kelly, E.J.; Quaife, C.J.; Brinster, R.L.; Palmiter, R.D. Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proc. Natl. Acad. Sci. USA 1994, 91, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, M.; Kojima, Y. Metallothionein and Other Low Molecular Weight Metal-Binding Proteins. In Metallothionein; Kägi, J.H.R., Nordberg, M., Eds.; Birkhäuser: Basel, Switzerland, 1979; pp. 41–124. [Google Scholar]
- Liu, Y.; Liu, J.; Iszard, M.B.; Andrews, G.K.; Palmiter, R.D.; Klaassen, C.D. Transgenic mice that overexpress metallothionein-I are protected from cadmium lethality and hepatotoxicity. Toxicol. Appl. Pharmacol. 1995, 135, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Basha, M.R.; Brock, B.; Cox, D.P.; Cardozo-Pelaez, F.; McPherson, C.A.; Harry, J.; Rice, D.C.; Maloney, B.; Chen, D.; et al. Alzheimer’s disease (AD)-like pathology in aged monkeys after infantile exposure to environmental metal lead (Pb): Evidence for a developmental origin and environmental link for AD. J. Neurosci. 2008, 28, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Coon, S.; Stark, A.; Peterson, E.; Gloi, A.; Kortsha, G.; Pounds, J.; Chettle, D.; Gorell, J. Whole-body lifetime occupational lead exposure and risk of Parkinson’s disease. Environ. Health Perspect. 2006, 114, 1872–1876. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, M.P.; Liu, J.; Goyer, R.A.; Diwan, B.A. Metallothionein-I/II double knockout mice are hypersensitive to lead-induced kidney carcinogenesis: Role of inclusion body formation. Cancer Res. 2004, 64, 7766–7772. [Google Scholar] [CrossRef] [PubMed]
- Zuo, P.; Qu, W.; Cooper, R.N.; Goyer, R.A.; Diwan, B.A.; Waalkes, M.P. Potential role of alpha-synuclein and metallothionein in lead-induced inclusion body formation. Toxicol. Sci. 2009, 111, 100–108. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, M.; Darabedian, N.; Liang, Y.; Wu, D.; Xiang, J.; Zhou, F. pH-dependent coordination of Pb2+ to metallothionein2: Structures and insight into lead detoxification. Inorg. Chem. 2014, 53, 2822–2830. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.C.; Shami Shah, A.; DeSilva, S.; Gleaton, A.; Su, A.; Goundie, B.; Croteau, M.L.; Stevenson, M.J.; Wilcox, D.E.; Austin, R.N. Thermodynamics of Pb(II) and Zn(II) binding to MT-3, a neurologically important metallothionein. Metallomics 2016, 8, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Gonick, H.C. Lead-binding proteins: A review. J. Toxicol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Fowler, B.A. Roles of lead-binding proteins in mediating lead bioavailability. Environ. Health Perspect. 1998, 106 (Suppl. S6), 1585–1587. [Google Scholar] [CrossRef] [PubMed]
- Church, H.J.; Day, J.P.; Braithwaite, R.A.; Brown, S.S. Binding of lead to a metallothionein-like protein in human erythrocytes. J. Inorg. Biochem. 1993, 49, 55–68. [Google Scholar] [CrossRef]
- Heuchel, R.; Radtke, F.; Georgiev, O.; Stark, G.; Aguet, M.; Schaffner, W. The transcription factor MTF-1 is essential for basal and heavy metal-induced metallothionein gene expression. Embo J. 1994, 13, 2870–2875. [Google Scholar] [PubMed]
- Dalton, T.; Palmiter, R.D.; Andrews, G.K. Transcriptional induction of the mouse metallothionein-I gene in hydrogen peroxide-treated Hepa cells involves a composite major late transcription factor/antioxidant response element and metal response promoter elements. Nucleic Acids Res. 1994, 22, 5016–5023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Georgiev, O.; Hagmann, M.; Gunes, C.; Cramer, M.; Faller, P.; Vašák, M.; Schaffner, W. Activity of metal-responsive transcription factor 1 by toxic heavy metals and H2O2 in vitro is modulated by metallothionein. Mol. Cell. Biol. 2003, 23, 8471–8485. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Andrews, G.K. Understanding the mechanisms of zinc-sensing by metal-response element binding transcription factor-1 (MTF-1). Arch. Biochem. Biophys. 2007, 463, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Magyar, J.S.; Weng, T.C.; Stern, C.M.; Dye, D.F.; Rous, B.W.; Payne, J.C.; Bridgewater, B.M.; Mijovilovich, A.; Parkin, G.; Zaleski, J.M.; et al. Reexamination of lead(II) coordination preferences in sulfur-rich sites: Implications for a critical mechanism of lead poisoning. J. Am. Chem. Soc. 2005, 127, 9495–9505. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Hayashi, Y.; Watabe, K.; Inuzuka, T.; Hozumi, I. Metallothionein-III prevents neuronal death and prolongs life span in amyotrophic lateral sclerosis model mice. Neuroscience 2011, 189, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, K.S.; Kim, H.N.; Kim, H.J.; Koh, J.Y. Role of zinc metallothionein-3 (ZnMT3) in epidermal growth factor (EGF)-induced c-Abl protein activation and actin polymerization in cultured astrocytes. J. Biol. Chem. 2011, 286, 40847–40856. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Kolbe, H.V.; Lepage, P.; Faure, T.; Sauerwald, R.; de la Salle, H.; Muller, C.; Bihoreau, N.; Paolantonacci, P.; Roitsch, C.; et al. Thrombin cleavage analysis of a novel antihaemophilic factor variant, factor VIII delta II. Eur. J. Biochem. 1991, 195, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.R.; Kim, D.K.; Koh, J.Y. Obesity and downregulated hypothalamic leptin receptors in male metallothionein-3-null mice. Neurobiol. Dis. 2011, 44, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Clifford, K.S.; MacDonald, M.J. Survey of mRNAs encoding zinc transporters and other metal complexing proteins in pancreatic islets of rats from birth to adulthood: Similar patterns in the Sprague-Dawley and Wistar BB strains. Diabetes Res. Clin. Pract. 2000, 49, 77–85. [Google Scholar] [CrossRef]
- Hozumi, I.; Suzuki, J.S.; Kanazawa, H.; Hara, A.; Saio, M.; Inuzuka, T.; Miyairi, S.; Naganuma, A.; Tohyama, C. Metallothionein-3 is expressed in the brain and various peripheral organs of the rat. Neurosci. Lett. 2008, 438, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.R.; Choi, J.A.; Koh, J.Y. The role of metallothionein-3 in streptozotocin-induced β-islet cell death and diabetes in mice. Metallomics 2014, 6, 1748–1757. [Google Scholar] [CrossRef] [PubMed]
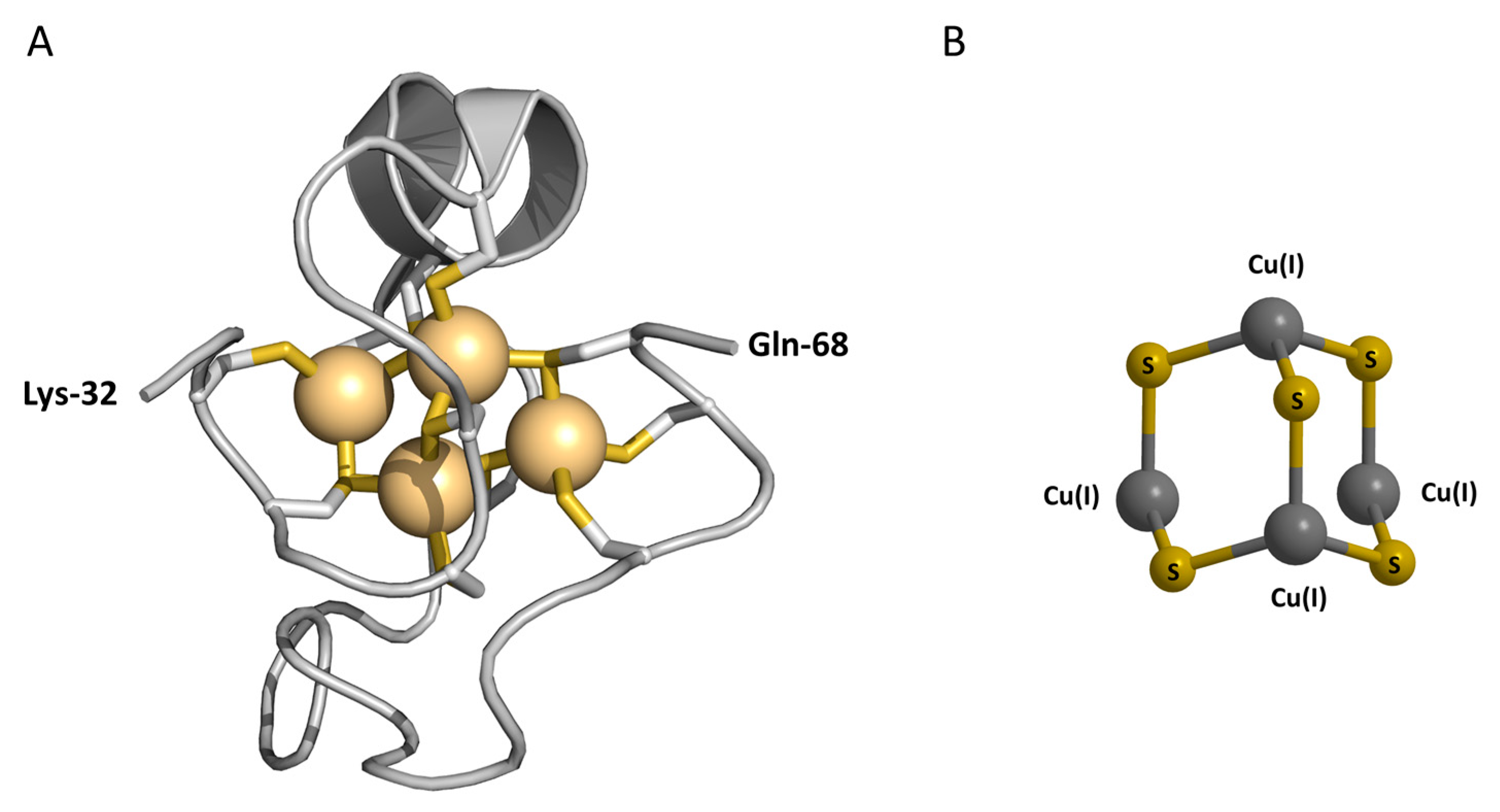
Figure 1.
Metal-thiolate clusters in metallothionein-3. (A) The NMR solution structure of the Cd4-α-domain of human Cd7MT-3. The Cd(II) ions are shown as light-orange spheres connected to the protein backbone by cysteine thiolate ligands. The model was generated with PyMOL using the Protein Data Bank coordinates of 2FJ5 [28]; (B) model of the Cu(I)4CysS5 cluster, derived from the spectroscopic characterization of Cu(I)4,Zn4MT-3 [29,30]. Metal ions are shown as dark gray spheres connected to cysteine sulfur ligands (yellow).
Figure 1.
Metal-thiolate clusters in metallothionein-3. (A) The NMR solution structure of the Cd4-α-domain of human Cd7MT-3. The Cd(II) ions are shown as light-orange spheres connected to the protein backbone by cysteine thiolate ligands. The model was generated with PyMOL using the Protein Data Bank coordinates of 2FJ5 [28]; (B) model of the Cu(I)4CysS5 cluster, derived from the spectroscopic characterization of Cu(I)4,Zn4MT-3 [29,30]. Metal ions are shown as dark gray spheres connected to cysteine sulfur ligands (yellow).

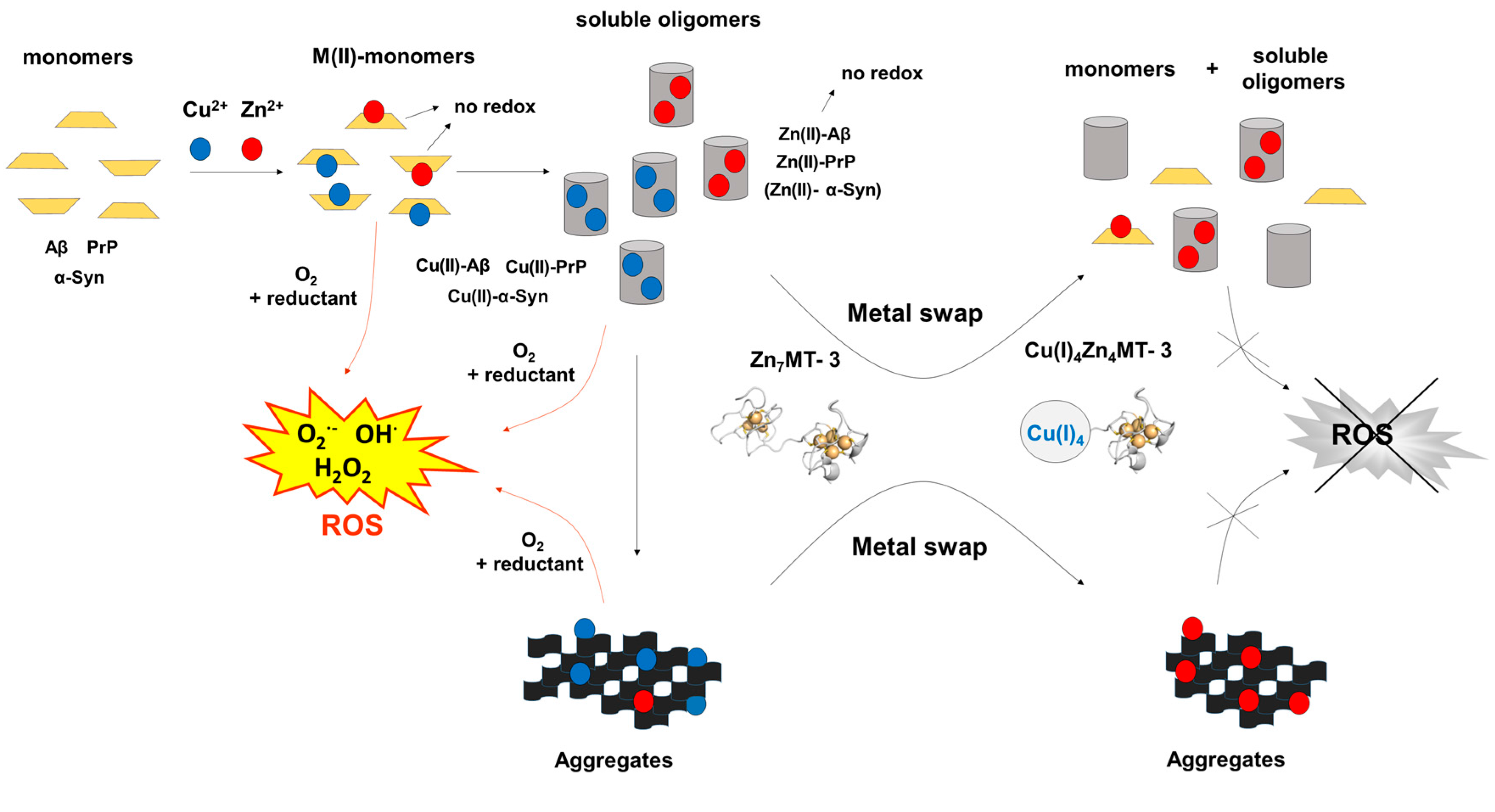
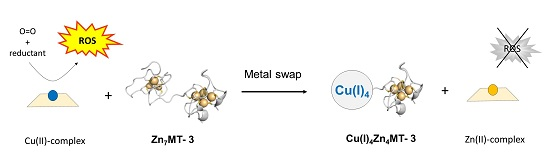
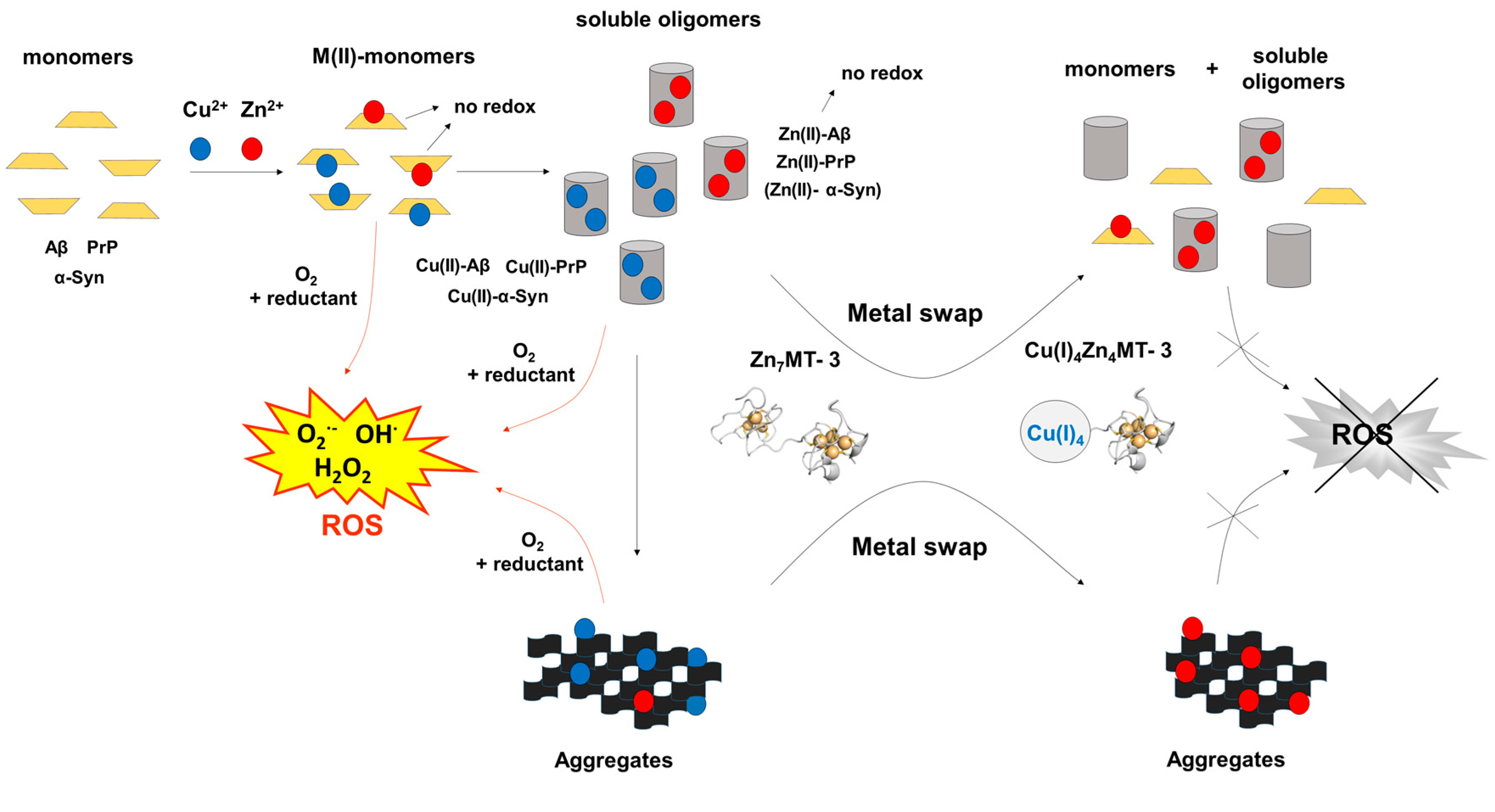
Figure 2.
Metal-swap reactions and Cu(II) redox-silencing by Zn7MT-3 in neurodegenerative disorders. The protective effect of human Zn7MT-3 (with the structure of β-domain modeled by that of rat Zn2Cd5MT-2 [77]) from the copper-mediated toxicity in AD, PD and prion diseases is summarized. The metal swap between Zn7MT-3 and the disease specific amyloidogenic Cu(II)–Aβ1–40 peptide [46], the Cu(II)–α-Syn [75] and Cu(II)–PrP proteins [76] abolishes the ROS production and the related cellular toxicity. In this process, Cu(II) is reduced by the protein thiolates concomitant with its binding into the N-terminal β-domain, forming the Cu(I)4Zn4MT-3 species and the non-redox-active Zn(II)–Aβ1–40, Zn(II)–α-Syn and Zn(II)–PrP. In Cu(I)4Zn4MT-3 an air-stable Cu(I)4-thiolate cluster and two disulfide bonds are present the N-terminal β-domain.
Figure 2.
Metal-swap reactions and Cu(II) redox-silencing by Zn7MT-3 in neurodegenerative disorders. The protective effect of human Zn7MT-3 (with the structure of β-domain modeled by that of rat Zn2Cd5MT-2 [77]) from the copper-mediated toxicity in AD, PD and prion diseases is summarized. The metal swap between Zn7MT-3 and the disease specific amyloidogenic Cu(II)–Aβ1–40 peptide [46], the Cu(II)–α-Syn [75] and Cu(II)–PrP proteins [76] abolishes the ROS production and the related cellular toxicity. In this process, Cu(II) is reduced by the protein thiolates concomitant with its binding into the N-terminal β-domain, forming the Cu(I)4Zn4MT-3 species and the non-redox-active Zn(II)–Aβ1–40, Zn(II)–α-Syn and Zn(II)–PrP. In Cu(I)4Zn4MT-3 an air-stable Cu(I)4-thiolate cluster and two disulfide bonds are present the N-terminal β-domain.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Electronic absorption and luminescence properties of Zn7MT-3, Cd7MT-3, and Cu(I)4Zn4MT-3 [26,29,30]. a ligand-to-metal-charge-transfer; b lifetime.
| MT-3 Form | Absorption | Luminescence (77 K) | |||||
|---|---|---|---|---|---|---|---|
| High-Energy Band | Low-Energy Band | ||||||
| 1st LMCT a Band (nm) | λem (nm) | τ b (μs) | λmax Excitation Spectra (nm) | λem (nm) | τ (μs) | λmax Excitation Spectra (nm) | |
| Zn7MT-3 | ~230 | - | - | - | - | - | - |
| Cd7MT-3 | ~250 | - | - | - | - | - | - |
| Cu(I)4Zn4MT-3 | ~265 | 425 | ~40 | envelope < 350 | 565 | ~135 | ~265; ~300 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vašák, M.; Meloni, G. Mammalian Metallothionein-3: New Functional and Structural Insights. Int. J. Mol. Sci. 2017, 18, 1117. https://doi.org/10.3390/ijms18061117
AMA Style
Vašák M, Meloni G. Mammalian Metallothionein-3: New Functional and Structural Insights. International Journal of Molecular Sciences. 2017; 18(6):1117. https://doi.org/10.3390/ijms18061117
Chicago/Turabian StyleVašák, Milan, and Gabriele Meloni. 2017. "Mammalian Metallothionein-3: New Functional and Structural Insights" International Journal of Molecular Sciences 18, no. 6: 1117. https://doi.org/10.3390/ijms18061117
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.