Proteomic High Affinity Zn2+ Trafficking: Where Does Metallothionein Fit in?

Department of Chemistry and Biochemistry, University of Wisconsin–Milwaukee, Milwaukee, WI 53217, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(6), 1289; https://doi.org/10.3390/ijms18061289
Submission received: 5 May 2017
/
Revised: 7 June 2017
/
Accepted: 9 June 2017
/
Published: 17 June 2017
(This article belongs to the Special Issue Metallothioneins in Bioinorganic Chemistry: Recent Developments)
Abstract
:The cellular constitution of Zn-proteins and Zn-dependent signaling depend on the capacity of Zn2+ to find specific binding sites in the face of a plethora of other high affinity ligands. The most prominent of these is metallothionein (MT). It serves as a storage site for Zn2+ under various conditions, and has chemical properties that support a dynamic role for MT in zinc trafficking. Consistent with these characteristics, changing the availability of zinc for cells and tissues causes rapid alteration of zinc bound to MT. Nevertheless, zinc trafficking occurs in metallothionein-null animals and cells, hypothetically making use of proteomic binding sites to mediate the intracellular movements of zinc. Like metallothionein, the proteome contains a large concentration of proteins that strongly coordinate zinc. In this environment, free Zn2+ may be of little significance. Instead, this review sets forth the basis for the hypothesis that components of the proteome and MT jointly provide the platform for zinc trafficking.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Overview of Zinc Trafficking
Zinc is a required nutrient for all forms of life. In mammalian organisms, as many as 3000 proteins may require Zn2+ to support their three-dimensional structures and/or biochemical functions [1,2]. Such proteins represent the endpoint of complex, only partially understood trafficking pathways that convey Zn2+ from external nutrient sources into organisms, then into cells through the agency of membrane transporters, and finally into contact with apo-Zn-proteins where metal ion binding occurs to constitute native Zn-proteins [3].
Studies over the past two decades have identified two families of Zn-transport proteins, ZIP and Zn-T, that escort Zn2+ into and out of mammalian cells and subcellular compartments [4,5,6] (Figure 1). Numerous reports document their key role in determining the intracellular availability of Zn2+ and the deleterious impact of mutations in Zn-transporter genes upon zinc-dependent processes [7]. Moreover, it is increasingly evident that the expression of at least some of these genes is under regulatory control, in part by the concentration of Zn2+ to which cells are exposed [8].
The trafficking of intracellular Zn2+ to its final sites of binding in Zn-proteins has remained relatively unexplored. Using the example of the constitution of Cu-proteins, it is hypothesized that upon transport into a cell, Zn2+ encounters metal-binding ligands (Lm) and undergoes one or more ligand substitution reactions that culminate in the formation of functional Zn-proteins (Zn-Pn) [9]:
In the case of copper trafficking in yeast and mammalian cells, L1–Lm represent a limited number of specific chaperone proteins that conduct Cu from its plasma membrane transporter to a small set of Cu-proteins by associative ligand substitution processes. Clearly, individualized ligand substitution pathways for the formation of the large array of zinc proteins cannot be built with this same level of selectivity, as that would require as many as 3000 or more chaperone proteins targeting different zinc protein structures. This paper considers alternative hypotheses to explain the trafficking of Zn2+ based on past and current studies.
The difficulty of this task lies first of all in the complexity of intracellular Zn2+ metabolism that involves the delivery of zinc to thousands of metalloproteins. Second, in comparison with the understanding about the widespread use of Zn2+ as a co-factor in protein structure and function, there is a paucity of information about Zn-binding proteins that may serve as participants in Zn2+ trafficking [10]. Third, the analytical tools needed to track the cellular movement of Zn2+ remain underdeveloped [11]. Thus, portrayals of zinc trafficking acknowledge the important role of ZIP and ZnT transporters, but have little to add inside the cell beyond free zinc (Zn2+) and a protein that researchers have been preoccupied with for the past 60 years, metallothionein (MT) (Figure 1) [4,5,6,7,8,12,13,14].
MT pops into view in many different cells and tissues with its concentration or complement of Zn2+ changing significantly depending on the physiological or pathological context [15,16,17,18]. It may be a transient storage site for Zn2+ or a Zn2+ chaperone protein (Reaction (1)), but it cannot be the only structure that supports the flexible trafficking of Zn2+ to, from, and between the multitude of Zn-proteins.
Where does MT fit into cellular Zn2+ metabolism? The hypothesis will be developed that Zn2+ trafficking occurs through the general operation of Reaction (1). It involves the joint, interactive participation of high affinity Zn2+ binding sites provided by the proteome and metallothionein.
2. Chemical Properties of Metallothionein Related to Zn2+ Trafficking
The background for considering the role of Metallothionein I and II in the mammalian intracellular trafficking of zinc begins with an understanding of its chemical properties. MT is a small, sulfhydryl-rich protein containing 20 cysteinyl residues that can bind multiple metal ions, including nutritionally essential Zn2+, Cu1+, and a variety of non-physiological metal ions such as the toxic metal ions Cd2+, Hg2+, and metals with pharmaceutical properties, like Pt2+ and Bi3+ [19,20]. Because it coordinates conspicuous amounts of a variety of metal ions that cells encounter, attention has been focused on its metal coordination, metal exchange, and ligand substitution properties. In addition, in some circumstances the concentration of MT is large enough that its aggregate of thiol ligands is comparable in size to the proteomic and low molecular weight glutathione (GSH) pools of sulfhydryl groups [21]. In such instances, MT has the potential to contribute quantitatively to the cell’s redox chemistry [22].
MT that is saturated with divalent metal ions (M) segregates them between two domains (α and β), in which either a M4S11 or M3S9 metal-thiolate cluster, respectively, occupies the interior of one of the folded domains [19,23]. In early studies, the titration of apo-MT with Zn2+ in the presence of a non-specific protease resulted in stable products that were either Zn4S11-α or Zn7-MT [24].
These findings, together with the knowledge of the structure of the clusters and their multiple sulfhydryl groups bridging between and connecting bound metal ions, supported the hypothesis that the protein structure is stabilized when one or both metal-thiolate clusters are formed.
The kinetics of formation of Zn7- and Cd7-MT have been characterized using stopped flow instrumentation [25]. Apo-MT and metal ions were mixed and allowed to react for increasing times before EDTA was added to quench the reaction. Three-sevenths of the overall reaction was too rapid to measure. The rest quickly formed an observable species that was thought to be M4S11-α, a structure slowly degraded by EDTA. Thus, in the presence of excess metal ions, stable domain structures fully occupied with metal ions are favored.
M7-MT undergoes facile ligand substitution- and sulfhydryl-based reactions. In a variety of reactions involving M7-MT, the kinetics are biphasic with each cluster thought to be responsible for one of the kinetic steps [26,27,28,29]. This implies that the rate-limiting reactions involve an attack on intact clusters. Once these structures are compromised, the remaining, bound metals or thiol groups react more rapidly.
This property has been used to examine the stability constants of Zn2+ in Zn7-MT [30,31]. Assigning each cluster to a kinetic step in a ligand substitution reaction and using the extent of reaction of each one to calculate equilibrium constants, the stability constant per Zn2+ at pH 7 in each cluster centered closely on 1011.2 M−1:
Zn3-β + 3L ⥂ 3Zn-L + β (faster phase)
Zn4-α + 4L ⥂ 4Zn-L + α (faster phase)
Recent analysis of the binding affinity of metals for MT has demonstrated that Znn-MT (n ≠ 4 or 7) can exist not just protein with fully occupied clusters [32]. Accordingly, the titration of MT by Zn2+ is characterized by seven stability constants, which at pH 7 range from 1011.8 to 1012.5.
The magnitude of these equilibrium constants indicates that on a thermodynamic basis apo-MT should be able to compete for Zn2+ with at least some native Zn-proteins [10,33]. Conversely, apo-proteins with strong affinity for Zn2+ may be able to sequester it from Znn-MT (n = 1–7), as in Reaction (1). For example, multiple experiments have revealed that apo-carbonic anhydrase (stability constant at pH 7 = 1011.4) successfully competes for Zn2+ bound to Zn7-MT [26,32,34,35,36]. The reconstitution of the zinc-finger protein, Zn-tramtrack from apo-tramtrak and Zn7-MT has also been shown [37].
The findings and conclusions above have been challenged by experiments showing that Zn7-MT can restore Zn2+ and activity to several proteins that display only moderate binding strength with Zn2+ [35,38,39]. In some of these cases, MT donated only one zinc to the acceptor protein [38,39]. Later, it was found that the protein preparation used in these studies contained one Zn2+ characterized by a stability constant at pH 7 of 107.8, as well as six others with stability constants centered on 1011 [40]. Apparently, protein activation by Zn2+ related to this low affinity site. Recently, it was discovered that a step in the preparation of the protein involving its incubation at pH 2 converted native MT with seven strong binding sites to this modified form [31]. Thus far, there is no evidence that cellular conditions can produce this altered species. Instead, MT apparently serves as a high affinity ligand for all of its bound Zn2+ ions.
The behavior of apo-MT in its titration with Co2+ contrasts with Reaction (2) [25,41]. Acting as a surrogate for Zn2+, the reaction of Co2+ with apo-MT was monitored first by electron paramagnetic resonance spectroscopy and later by the differential reactivity of metal-free and bound sulfhydryl groups. It occurs in step-wise fashion, not in a concerted all-or-nothing process that generates only fully formed clusters:
Co2+ + apo-MT ⥂ Co-MT ⥂ Co2-MT ⥂ Co3-MT ⥂ Co4,S11-MT…
An early titration of apo-MT with Zn2+ monitored with electrospray mass spectrometry (ESI) suggested that cluster unsaturated species of Znn-MT (n = 1–3, 5, 6) form along with clusters fully occupied with Zn2+ [42]. Subsequently, an elegant series of experiments by Stillman and his students, also using ESI, strongly supported the stepwise formation of Zn7-MT from Zn2+ and apo-MT [32,43,44]. In particular, when apo-MT, apo-CA, and various concentrations of Zn2+ were mixed, the metal ion distributed itself among Zn-CA and Znn-MT (n = 1–7) such that equilibrium constants for the binding of each Zn2+ to MT could be determined as described above [32]. They ranged between 1012.5 to 1011.8 for the first and seventh zinc ion, respectively.
The large thermodynamic stability of Zn7-MT supports its capacity to bind extra Zn2+ that enters cells and suggests that it may compete for intracellular Zn2+ (Figure 1). In contrast, its unusual kinetic reactivity and demonstrated reaction with apo-zinc proteins raises the possibility that Znn-MT might participate in zinc trafficking as in Reaction (1) [45]. How these properties reveal themselves in the cell is considered below.
3. Cellular Zn2+ and Its Milieu
Typical mammalian cells contain concentrations of zinc in the range of 100–500 µM [46,47]. Virtually all of it is thought to represent the collection of native Zn-proteins that populate the cell. Conditional stability constants for a few of them have been measured at pH 7 [10,33]. They range between 109 and 1012 M−1. As discussed below, the pressure of zinc deficiency exerts little if any measurable effect on the concentration of Zn2+ in a number of tissues and cells, exclusive of that bound to metallothionein.
The large concentration of Zn2+ bound to Zn-proteins is mobile on some time scale because in the dynamics of synthesis and degradation of intracellular proteins, Zn2+ must be continually delivered to newly synthesized apo-Zn-proteins as it is released from proteins undergoing hydrolytic break-down (Figure 2). Potentially, this is a cyclic process with respect to Zn2+ and zinc binding ligands L1-m (Reaction (1)), but may also involve cellular uptake and release of Zn2+ via transporters. In this diagram, it is assumed that L1-m represent proteins or possibly small metal coordinating ligands such as glutathione (GSH). A role for MT is not specified; but it has been suggested that MT serves as an intermediate in the trafficking of zinc to apo-zinc proteins [26,45].
The functional groups of a number of amino acid side chains can serve as ligands for metal ions (amine, carboxyl, imidazole, thiol) [48]. It is not surprising, therefore, that proteins generally display affinity for a variety of metal ions. Maret and Krezel used the term zinc buffering to describe non-specific binding of Zn2+ by the cell’s complement of proteins and small molecules. To reveal this property, cytosol from human colon cancer (HT-29) cells was titrated with Zn2+ in the presence of a colorimetric sensor, Zincon (ZI), which has a modest stability constant for Zn2+ at pH 7 of 104.9 M−1 [46]. On a cellular basis, 30 µM Zn2+ or about 10% of the size of the Zn-proteome reacted with the proteome before Zn-ZI began to form.
A similar experiment employed proteome from the sonicated supernatant of pig kidney LLC-PK1 cells [49]. In the presence of the fluorescent sensor, Newport Green (NPG) (KZn-NPG = 105 M−1, pH 7), titration of the proteome with Zn2+ revealed a much larger complement of high affinity coordination sites that bind Zn2+ exclusively in the presence of NPG and an even greater concentration of low affinity sites with which NPG directly competes (Reactions (6) and (7)).
Proteomehigh affinity•Zn + S (ZI, NPG) ⥂ no reaction
Proteomelower affinity•Zn + S ⥂ Zn-S + Proteomelow affinity
Titration of LLC-PK1 proteome with Zn2+ in the presence of another fluorescent probe, Fluozin-3 (FZ-3), which has a higher conditional stability constant for Zn2+ of 108.1 M−1 at pH 7, resulted in the same general picture, except that FZ-3 competes with Proteomehigh affinity for Zn2+ (Karim and Petering, unpublished information). Because of this, a conditional stability constant of Proteomehigh affinity for Zn2+ could be estimated at 1010 M−1. The LLC-PK1 cell proteome contains a multitude of such binding sites for Zn2+, about 1.5 times more than are present in native Zn-proteins. Repetition of the experiments with ZI, NPG, and FZ-3 after treating the proteome with the thiol reactive agent N-ethylmaleimide quantitatively abolished high affinity sites, leaving only unperturbed, low affinity binding of Zn2+ by the proteome. Dithiodipyridine and nitric oxide also react with sulhydryl groups and have a similar qualitative impact on adventitious zinc binding by the proteome [50,51]. Apparently, strong binding sites for Zn2+ involve thiolate ligands. Not known, however, are the numbers, identities, and concentrations of the proteins that interact with Zn2+.
The Zn2+ binding strength and large size of the collection of proteins in the proteomehigh affinity make clear that glutathione (stability constant for Zn2+ at pH 7 of 104.2) does not play an independent role as an intermediate in Zn2+ trafficking (Reaction (1)) [52]. Instead, the model presented in Figure 2 portrays trafficking proteins (L1-m). Consistent with this emphasis, Zn2+ added to cells or cell lysates or mobilized from native Zn-proteins by displacement with Cd2+ associates with the proteome fraction not with glutathione or other low molecular weight molecules [53]. It is within this context that a role for metallothionein in Zn2+ trafficking must be considered and begins to make sense.
The findings above bring into question the significance of free zinc (Zn2+). The pool of Zn2+ in Figure 1 and Figure 2 has been invoked as the direct source of zinc for the synthesis of Zn-proteins, a participant in zinc signaling processes (see below), and as the agent of zinc toxicity [40,54]. Thus, the measurement of its concentration under various conditions using fluorescent zinc sensors (S) has received substantial attention [55,56]. Sensors are thought to react with free or available Zn2+ according to Reactions (8)–(10):
in which L represents non-specific binding sites and Zn-Proteins are specific sites. Depending on the conditional stability constant of Zn-S (K), and the kinetics of its reaction with zinc-ligand complexes, S may be able to sequester Zn2+ from Zn-L or Zn-Protein sites as well as react with Zn2+. Thus, whether measurements with zinc fluorescent probes reflect the presence of Zn2+ or the accessibility of other pools of Zn2+ remains in doubt. Nevertheless, in unperturbed cells, the most that fluorescent probes detect is low nM to low pM zinc [57,58,59]. At such miniscule concentrations, and in comparison with the total Zn2+ in cells, it seems unlikely, if not impossible on a kinetic basis, that steady state concentrations of free Zn2+ can supply the metal ion needed to constitute new Zn-proteins in proliferative cells [3]. Therefore, Reaction (1) remains as the viable, hypothetical pathway that leads to the formation of Zn-proteins. More generally, the focus on free Zn2+ seems misplaced. In the equilibrium or steady state reactions that connect free and bound Zn2+, it is the structures with which mobile Zn2+ interacts that give rise to functional outcomes, and not free, unbound Zn2+.
Zn2+ + S ⥂ Zn-S (K)
Zn-L + S ⥂ Zn-S + L (K′)
Zn-Protein + S ⥂ Zn-S + Protein (K′′)
4. Metallothionein and Zinc Trafficking—The Biological Context
Metallothionein was discovered as a cadmium-binding protein in horse kidney [60]. Within a decade it was clear that the protein was rapidly inducible and that copious amounts of hepatic Zn-MT could be accumulated upon elevated dietary intake of Zn2+, and lost rapidly upon imposition of zinc deficiency [61]. Soon, it was realized that high concentrations of metallothionein were present under normal physiological conditions. For example, fetal and neonatal bovine and human liver contains large amounts of Zn,Cu-MT, thought to serve as a mobilizable store of these metals for the developing organism [62,63]. Likewise, juvenile and adult rat kidney contains substantial pools of Zn,Cu-MT, for reasons that remain unclear [15]. Moreover, rats exposed to a variety of stresses, such as cold, pathogens, burns, chemicals, etc., quickly accumulate Zn2+ from plasma in hepatic MT as part of the general organismic stress response [16]. Such results focused attention on MT’s participation in Zn2+ trafficking as a dynamic storage location that might chemically exchange metal ions with other sites.
The constitutive presence or induction of MT protein also potentially introduces a large pool of sulfhydryl groups into the cell alongside those of glutathione and the proteome. As a consequence, MT may play an important role in the cell’s response to stress imposed by electrophilic reagents, such as oxidants. Numerous chemical and cellular studies document the reactivity of thiolate groups in Zn- and apo-MT with agents such as H2O2 vs. NO [21,22,64,65]. As anticipated, Zn2+ retards the reaction of sulfhydryl groups with electrophiles [25,64]. Conversely, oxidative modification of MT’s sulfhydryl groups renders bound Zn2+ available for trafficking reactions [66]. From this point of view, the involvement of MT in Zn2+ trafficking and in cellular redox chemistry become intertwined.
5. Zinc Trafficking in Metallothionein-Null Organisms and Cells
Inquiries into the functions and importance of metallothionein must recognize that gene knock-outs of MT I and II do not prevent the birth and survival of MT-null mice [67]. Clearly, MT is not required for zinc trafficking; cells can manage zinc metabolism without it. In this context, investigations with MT-null animals have revealed the protein’s significance in protecting cells faced with a host of stress conditions, including exposure to heavy metals such as Cd2+ and methylmercury, challenge by toxic oxidants and chemotherapeutic drugs with electrophilic properties, [68,69,70,71].
Laboratory animals live in hygienic conditions and consume super-optimal diets, commonly receiving, for instance, multiple times their daily requirement of Zn2+. Reproduction and survival of MT-null mice occurs under such conditions. In contrast, when pregnant MT-null mice were subjected to mild zinc deficiency that caused little harm to control fetuses, about 30% died or showed frank malformations [72]. Moreover, MT-null mice experienced reduced body weight gain and bone structure development with the effects intensifying as dietary zinc declined from normal to modestly deficient [73]. Evidently, the absence of MT compromised the handling of zinc within the mother and fetus and then in the growing offspring. These results signal a need to examine how MT plays a significant, but subtle, role in zinc trafficking.
Other reports also document the deleterious impact of the absence of MT I and II on the physiology of mice. The MT null phenotype alters neurological function and causes alterations in energy metabolism leading to modest obesity [74,75]. Transient down-regulation of MT inhibits cell cycle progression in proliferative breast cancer cells and causes apoptosis of ovarian and prostate tumor cell lines [76,77,78].
All of these results infer that the MT protein has important roles to play in a variety of cellular activities, a number of which are associated with zinc. However, a general short-coming of these reports is that the observations of physiological derangements in MT-null mice stand alone. Parallel studies have not been undertaken to understand the underlying chemistry of MT and zinc in control cells and their MT-null counterparts. As a result, mechanisms by which MT exerts its effects remain speculative.
6. Metallothionein, Cell Proliferation, and Zinc Deficiency
Normal fetal and neonatal/juvenile growth and development are highly reliant on the presence of an adequate supply of nutrient Zn2+ [79]. For example, in juvenile rats, plasma zinc concentration plummets within 24 h upon imposition of a zinc deficient diet [16]. Increase in body weight terminates in this same time frame. Kidney, which contains a readily measurable concentration of Zn,Cu-MT, also loses its complement of MT-bound Zn2+ during this period. In contrast, upon imposition of zinc deficiency for at least 30 days, the concentration of proteomic zinc other than what is bound to MT does not change in kidney or other tissues, such as liver [16]. After restoration of zinc to the diet, plasma and MT regain Zn2+ with the same kinetics, and the rats begin growing again. Thus, at this gross level of analysis, MT appears unique in its responsiveness to nutrient zinc status. Such observations sharpen the focus on metallothionein in relation to Zn2+ trafficking.
Cancer tumor growth is also keenly sensitive to dietary zinc status [80]. In an Ehrlich ascites carcinoma tumor model, cancer cells injected into the mouse peritoneum stimulate ascites fluid production that provides the cells with their nutrition [81,82]. The concentration of zinc in the fluid paralleled levels of dietary zinc; at low values, cell proliferation was inhibited. In contrast, over the range of normal to deficient concentrations of dietary zinc, the amount intracellular zinc did not measurably vary.
The involvement of metallothionein in supporting cell proliferation has also been established in a series of MT-knockdown and knockout studies [76,77,83,84]. With this association between MT, zinc, and cell proliferation established, studies have inquired into the mechanistic role of metallothionein in supporting zinc-contingent growth of organisms and cell populations.
Ehrlich ascites cells contain a steady-state pool of Zn-MT that accounts for about 15% of the cellular Zn2+ [85]. When placed in a zinc-deficient medium in vitro, MT lost Zn2+ rapidly with a first order half-time of 1 h, whereas the rest of the proteomic zinc declined slowly with an estimated t1/2 of 43 h [86]. The half-times for MT and proteome protein degradation were measured as ca. 5 and 11 h, respectively. The results demonstrated that the shift of Zn2+ out of MT was not rate limited by its biodegradation. In addition, the rate of loss of proteomic zinc was much slower than the turnover rate of the proteome’s complement of proteins. Not determined in this work was the fate of the zinc that was transferred out of MT. This experiment also revealed that proteomic Zn2+ was maintained despite the turnover of protein. Importantly, it supported the hypothesis that Zn-MT participates in cellular Zn2+ trafficking and that it does so through a chemical mechanism not passive protein turnover.
The rate limiting chemical reaction might involve dissociation of Zn2+ from MT or ligand substitution with competing binding sites for Zn2+:
Because of the large conditional stability constants for the coordination of Zn2+ by MT, the kinetics of dissociation of Zn2+ from MT (Reaction (11)) must be very slow and cannot account for the swift depletion of MT-bound Zn2+ in this experiment [31,32]. Thus, Reaction (12) remains, and is well supported by chemical studies, showing the ligand substitution reactivity of MT [26,28,45]. This experiment distinguished the behavior of Zn2+ bound to MT from that associated with the proteome. The former behaved like a trafficking intermediate or labile store of Zn2+, and the latter as its steady state destination.
Zn-MT ⥂ MT + Zn2+
Zn-MT + Lm ⥂ MT + Zn-Lm
In vivo experiments examined the behavior of Ehrlich ascites cells in mouse peritoneum after transition of animals to a zinc-deficient diet. Ordinarily, the Ehrlich ascites tumor is lethal to mice about 17 days after injection of cells. In contrast, under zinc-limiting conditions, there was no gross evidence of tumor after 24 days, consistent with previous observations that zinc deficiency inhibits both normal tissue and tumor growth [81]. Dormant tumor cells contained 80% of the control level of proteomic zinc, essentially missing Zn2+ associated with MT, in agreement with the view that MT-bound Zn2+ is particularly labile in the face of changing external concentrations of zinc [87].
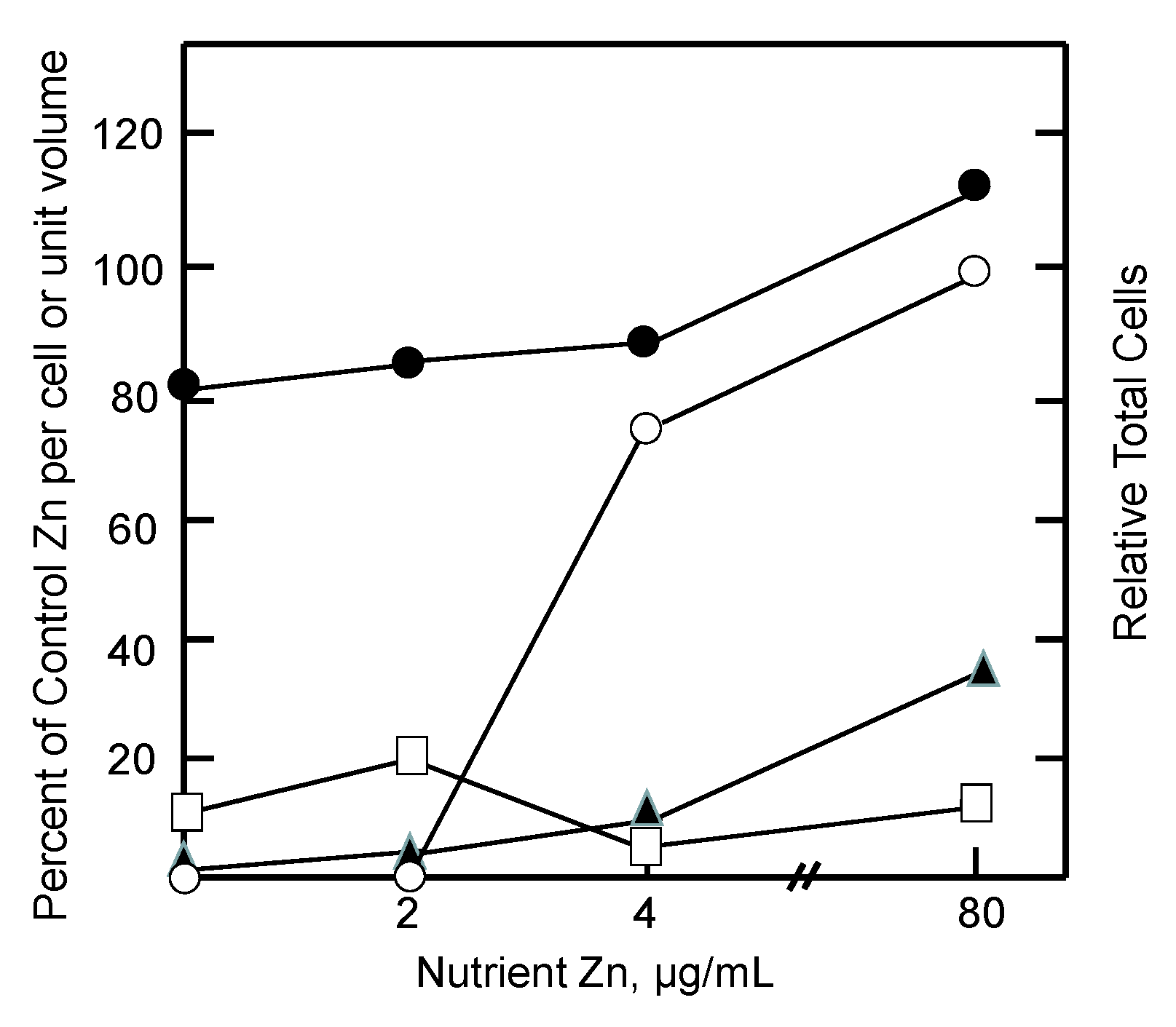
During the reactivation of tumor growth by zinc, the extent of reentry of zinc into (i) ascites fluid, the immediate source of Zn2+ for the cells; (ii) the growing cell population; and (iii) intracellular proteomic and MT pools were assessed. As illustrated in Figure 3, measurements during days 6–9 after addition of zinc to the diet showed that tumor growth was restored by increasing concentrations of nutrient zinc. Evidently, the tumor acquired the requisite amount of zinc to support cell division. But the availability of zinc did not drastically alter the per cell proteomic zinc concentration. In contrast, even at the super optimal level of 80 µg/mL of dietary zinc, ascites fluid zinc remained depressed while zinc associated with MT was only partially restored to control levels. It was clear that if cells are to divide, they must acquire their total, functional complement of proteomic zinc. That was not the case for ascites fluid and MT. Instead, as in the in vitro cellular experiments described above, metallothionein behaved like an intermediate along the pathway of zinc from diet to the proteome as did ascites fluid. As such, the results are consistent with the zinc trafficking sequence,
Diet → Ascites fluid → MT → Proteome
The behavior of metallothionein during the stress of zinc deficiency has been the subject of other studies, as well. O’Halloran and others discovered that MT mRNA increased markedly in fibroblast cells (LZA-LTK-) and animals upon their transfer into a zinc-deficient growth medium, suggesting a role for the protein in the response to the stress of limiting available Zn2+ to support proliferation [73,88]. These studies documented the inhibition of cell proliferation by the zinc deficit, but were unable to show that the MT protein concentration was correspondingly elevated.
Recent experiments addressed this question with three cell lines, LLC-PK1 (pig kidney proximal tubule), TE-671 (human rhabdomyosarcoma), and U-87 (human glioma) (Rana and Petering, unpublished information). The first two contain little MT. The third displays a significant, constitutive concentration of zinc-unsaturated MT. When each was shifted into a growth medium that was depleted of Zn2+, elevated concentrations of apo-MT appeared within 24 h that ranged between 1.6 and 4 times control levels. Coincidently, the proliferation of each cell line was halted without net loss of zinc from the cells.
These results are rationalized in relation to Figure 4. In the absence of nutrient zinc, the plasma membrane exporter of Zn2+, ZnT1, is down-regulated to prevent the loss of intracellular Zn2+, which becomes transiently mobile during the biodegradation of Zn-proteins [89]. In addition, in this model, the pool of intracellular zinc is sustained by up-regulation of MT concentration which both sequesters Zn2+ released by Zn-protein degradation and potentially donates it back to apo-Zn-proteins.
7. Metal-Unsaturated Metallothionein and the Trafficking of Zinc
The 3-dimensional structures of Cd5,Zn2-MT and Cd7-MT are determined by the interior metal-thiolate clusters, which constrain the folding of the polypeptide chain around them [19,90]. The conventional view has held that in the absence of metals, MT is unstructured and should be swiftly degraded in cells. Moreover, its destruction is necessary to prevent deleterious competition for Zn2+ between metal-unsaturated or apo-MT and native Zn-proteins. Nevertheless, it was discovered that numerous proliferative cell types contain readily measurable and sometimes large concentrations of constitutive apo- or unsaturated Zn-MT under normal growth conditions [91]. Subsequently, this finding was extended as well to non-proliferative tissues [92]. Moreover, a variety of agents that induce MT synthesis such as the stress hormone dexamethasone result in metal-unsaturated MT, as do conditions that cause the loss of metal ions from MT [93]. Detection of unsaturated Zn-MT is fully consistent with recent chemical studies showing that MT accommodates 1–7 Zn2+ ions [32,43].
Apo-MT and partially zinc-saturated protein display a similar overall shape to M7-MT, migrating during gel filtration chromatography like an ellipsoid (10 kDa) rather than a globular (6 kDa) protein [91,94]. That the apo-protein adopts a secondary structure is supported by experiments showing that the reactivity of cysteinyl sulfhydryl groups differs in native and denatured apo-MT, and that the rate of binding of Cd2+ to the protein also varies between these two states [95,96].
Considering the large stability constants that characterize the interaction of Zn2+ with MT (1011–13), it is important to understand how cellular apo-MT can co-exist with the extant Zn-proteome, and with the zinc-trafficking mechanism that provides zinc for Zn-proteins being synthesized during cell proliferation [31,32]. Remarkably, on a quantitative basis, apo-MT competes poorly, if at all, with isolated Zn-proteome from LLC-PK1 cells, even though a number of organic chelating agents with conditional stability constants ranging from 1015 to 109 at pH 7 are able to extract up to 30% of proteomic zinc under similar conditions (Reaction (14)) [97].
Zn-Proteome + L ⥂ Zn-L + Proteome
Evidently, apo-MT is kinetically inert to ligand substitution in reactions of this sort. Possibly, intermediate L-Zn-protein adducts are crucial for the mechanism of substitution. Whereas smaller multidentate ligands make such intermediates achievable, the steric bulkiness of apo-MT inhibits this stage of the reaction. In support of this view, it has been demonstrated that the partial MT sequence 51–60, including four cysteinyl thiolates that normally bind to a single Zn2+ in Zn7-MT, sequesters Cd2+ from Cd-carbonic anhydrase much more rapidly than the more sterically hindered apo-MT [98].
Apo-MT does react with and inactivate the zinc-finger protein, Zn3-Sp1, which utilizes three tandem cys23his2 Zn-fingers to bind to its cognate DNA [97].
Presumably, the relatively accessible zinc coordination sites of Zn3-Sp1 favor ligand substitution, as discussed above. Comparison of this reaction in the presence and absence of a GC-rich DNA binding sequence for human Zn3-Sp1 revealed that DNA protected the protein against loss of Zn2+ to MT [97]. This observation is consistent with the finding that specific DNA binding also protects Zn3-Sp1 against reaction with powerful chelating agents such as EDTA [99].
Zn3-Sp1 + apo-MT ⥂ Zn3-MT + apo-Sp1
These results show that apo-MT, a strong themodynamic sink for Zn2+, neither sequesters much zinc in competitive reactions with the native Zn-proteome, nor pulls Zn2+ into cells via ZIP transporters to saturate its binding sites. Nevertheless, when cells were exposed to elevated extracellular Zn2+, increasing concentrations of MT were synthesized that accommodated extra Zn2+ entering the cells [61]. In this instance, the production of apo-MT served to protect cell viability. What does the existence of cellular apo-MT signify for the mechanism of zinc trafficking under conditions of normal zinc flux into cells?
It is evident that the presence of a substantial concentration of this powerful ligand for Zn2+ does not compete with and inhibit the normal trafficking pathway that conducts Zn2+ from the plasma membrane to apo-proteins that require Zn2+ for their function. A hypothesis that rationalizes this observation is based on Figure 2, and posits that MT acts as an intermediate in the trafficking of Zn2+ to Zn-proteins (Figure 4). As it delivers Zn2+ to apo-proteins, MT undergoes a cycle, moving between Zn-MT and apo- or unsaturated-MT. The concentration of Zn2+ bound to MT, Znn-MT (n = 1–7), depends on the relative velocities of the reactions that shift Zn2+ into apo-protein (V2) and restore Zn2+ to apo-MT (V1). Differences in V1 and V2 in various cells would explain the observed variation in saturation of MT with Zn2+. Furthermore, V1 is expected to be dependent on the activity of ZIP and ZnT transporters that facilitate the transport of Zn2+ between the external medium and the cell interior. Thus, the relative saturation of the MT pool with zinc is likely to be an integrative outcome of the relative rates of the various zinc trafficking reactions shown in Figure 4 including zinc transport.
In order to test this hypothesis, the degree of Zn2+-saturation of MT was investigated as a function of the proliferative state of TE671 cells [94]. Normally, a steady-state pool of apo-MT exists in the cells, which double about every 24 h. Exposure to thymidine suppressed cell proliferation. In the process apo-MT was converted to Zn-MT. As the requirement for Zn2+ declined, V2 decreased. Assuming V1 remained approximately unchanged, Znn-MT would become favored over apo-MT, as was observed. In a second test of this hypothesis, Ehrlich cells contain a steady state pool of Zn7-MT (V1 > V2) [87]. Determination of the rate of V2 under zinc-restricted conditions in which V1 = 0 showed that the magnitude of V2 is sufficient to supply most or all of the Zn2+ needed to support the rate of doubling by control Ehrlich cells [86].
8. Oxidized Metallothionein and the Trafficking of Zn2+
Mammalian MT contains 20 cysteinyl amino acids that supply sulfhydryl group ligands that bind Zn2+ and other metal ions [19]. Besides its participation in zinc trafficking, the sulfhydryl groups provide MT with the potential to serve as an important cellular anti-oxidant. Numerous reports demonstrate this role under a variety of conditions of oxidative stress [21,22,65,66,70]. Most of these studies showed that the presence or lack of MT correlated with a reduction in, or enhancement of, oxidant damage, respectively. Others found that Znn-MT undergoes thiol oxidation in the presence of oxidants [21]. For MT to play a significant role in intercepting reactive oxygen species, it needs to be able to cycle between reduced and oxidized forms. Glutathione (GSH) readily reduces disulfide bonds in oxidized MT and, thereby, provides the means to maintain a pool of reduced MT [100].
A complete analysis of the importance of MT in protection against oxidant damage needs to compare its concentration and activity with the other sources of sulfhydryl groups. Glutathione constitutes a major pool of 1–10 mM thiol [101]. The proteome, which is a target of oxidant damage, contains a similar concentration of sulfhydryl groups [21,64]. To the extent that cysteinyl residues in the proteome become oxidized, its zinc buffering capacity may also be compromised, as discussed above. When H2O2 was the oxidant, Zn-MT was especially reactive among these classes of thiol groups [21]. In contrast, the other pools were preferentially reactive with NO [64].
Zinc-bound sulfhydryl groups that participate in redox reactions release Zn2+ as the protein is oxidized. Considering the interest in Znn-MT as a donor of Zn2+ to apo-Zn-proteins, and the concern that its large affinity for Zn2+ might restrict such reactions, the hypothesis has gained popularity that the availability of zinc bound to MT for such reactions is dependent on the oxidation of zinc-metallothionein by the oxidized form of GSH, GSSG [38].
Zn7,S20-MT + 10GSSG ⥂ MT-(S-S)10 + 7Zn2+ + 20GS−
Zn2+ + Proteomehigh affinity ⥂ Proteomehigh affinity•Zn
Proteomehigh affinity•Zn + apo-Protein ⥂ Zn-Protein + Proteomehigh affinity
9. Metallothionein Gene Expression
The facile induction of MT synthesis by Zn2+ and Cd2+ offers an example of trafficking that can be understood on the basis of a ligand substitution mechanism (Figure 1). Characterization of the determinants of metallothionein gene expression helped to usher in the modern era of eukaryotic molecular biology [103]. Of particular interest was the mechanism of induction of MT protein synthesis by metal ions including Zn2+. First, metal response elements (MREs) in the MT promoter region were discovered that conferred inducibility by zinc on the MT gene [104]. Then, a metal-response element-binding transcription factor, MTF-1, was identified [105]. It contains a MRE binding region comprised of six tandem Zn-fingers that utilize the common cys2his2 ligand set to bind Zn2+. Further, MTF-1’s DNA binding function was shown to be upregulated by Zn2+.
A simple model for MTF-1 activation was invoked, in which the protein gained the ability to bind to MRE DNA sequences upon conversion of MTF-1 into Znn-MTF-1:
Experiments using lysates as a source of MTF-1 required µM additions of Zn2+ to stimulate MTF-1 binding to MREs [106]. This suggested that the conditional stability constants of the zinc-finger contingent of MTF-1 were modest and on the order of 106 M−1 at pH 7.
nZn2+ + MTF-1 ⥂ Znn-MTF-1
The picture changed when direct measurements indicated that the conditional stability constants of component zinc-fingers of MTF-1 cluster about 1011 M−1 at pH 7 and it was recognized that the lysate used above was populated by strong coordination sites that competed with MTF-1 for Zn2+ [3,107]. As a robust Zn2+ binding ligand, MTF-1 could possibly react with pM-nM concentrations of free Zn2+ and might also be able to interact with Zn2+ associated with the proteome (KZn-proteome ≈ 1010) to undergo activation. Once the extracellular Zn2+ concentration returned to normal, the reversal of Reaction (19) seemed unlikely as the mechanism of deactivation, because the Zn2+ dissociation rate constants would be too small to support kinetically favorable loss of Zn2+ from the protein. Thus, sequestration of Zn2+ from Znn-MTF-1 (n = 1–6) must involve ligand substitution, in which the product of Znn-MTF-1 stimulated gene expression, apo-MT, or components of the proteome compete for Zn2+ to inactivate it (Reactions (20) and (21)):
Both apo-MT and Proteomehigh affinity bind Zn2+ with sufficiently large stability constants and/or concentrations to compete for zinc bound to Znn-MTF-1.
Znn-MTF-1 + apo-MT ⥂ MTF-1 + Znn-MT (n = 1–6)
Znn-MTF-1 + nProteomehigh affinity ⥂ MTF-1 + nProteomehigh affinity•Zn
Supporting the involvement of apo-MT is the observation that the concentration of MT in the nucleus, the preferential location of zinc-activated MTF-1, increases following its induction by extracellular Zn2+ [108,109]. A possible complication for this reaction lies in the finding with another zinc-finger protein, Zn3-Sp1, that association with DNA markedly decreased its reactivity in model ligand substitution reactions [97]. If that is the case with Znn-MTF-1, then the rate of dissociation of Znn-MTF-1•MRE may act as the rate limiting step in the deactivation of this protein.
MTF-1 activation also occurs in cells exposed to cadmium ions. Originally, it was thought that Cd2+ like Zn2+ might activate the transcription factor directly through reaction with the apo-zinc finger domains of the protein. However, in vitro DNA binding assays showed that Cd2+ inhibited the MTF-1 interaction with metal response element DNA [110]. Moreover, according to structures of model peptides, Zn- and Cd-finger structures are subtly different such that the DNA recognition helix of the Cd-finger cannot make effective contact with cognate DNA [111]. Thus, it was proposed that Cd2+ reacts with Znn-MT, causing the displacement of Zn2+ and making it available for reaction with MTF-1 [112]:
Missing from this hypothesis was the recognition that the concentration of Znn-MT may not be significant in some cells; yet induction of MT biosynthesis occurs upon their exposure to Cd2+. In these, and perhaps in most, cells, Zn2+ may be labilized for reaction with MTF-1 through metal ion exchange reactions between Cd2+ and Zn-proteins (Zn-proteome). Proteomic studies have shown that much of the native Zn-proteome can exchange with Cd2+ resulting in the liberation of Zn2+ from native zinc-binding sites and its subsequent adventitious association with high affinity, adventitious coordination sites in the proteome [53]:
In this mechanism, Proteome•Zn serves as the donor of Zn2+ to MTF-1 (Reaction (25)). Based on this example, it seems evident that viable models of signaling processes that utilize MTF-1 or other proteins to detect variable concentrations of Zn2+ will need to envision mechanisms that work within the strong Zn2+ binding environment provided by the proteome and apo- or unsaturated-MT.
nCd2+ + Znn-MT ⥂ Cdn-MT + nZn2+
Cd2+ + Zn-Proteome ⥂ Cd-Proteome + Zn2+
Zn2+ + Proteome ⥂ Proteome•Zn
Proteome•Zn + MTF-1 ⥂ Zn-MTF-1 + Proteome
10. Interaction of Metallothionein and the Proteome during Zinc Trafficking
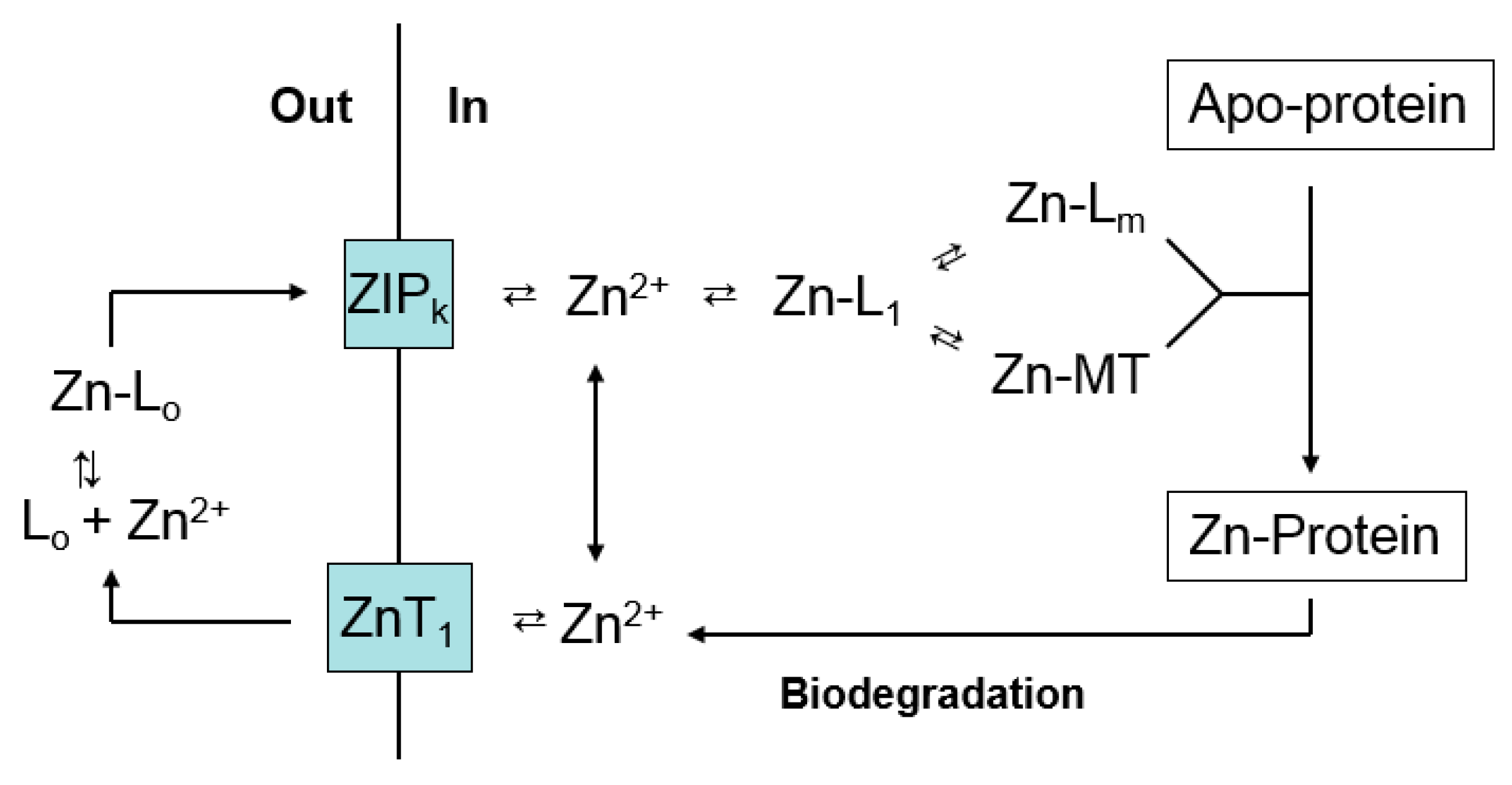
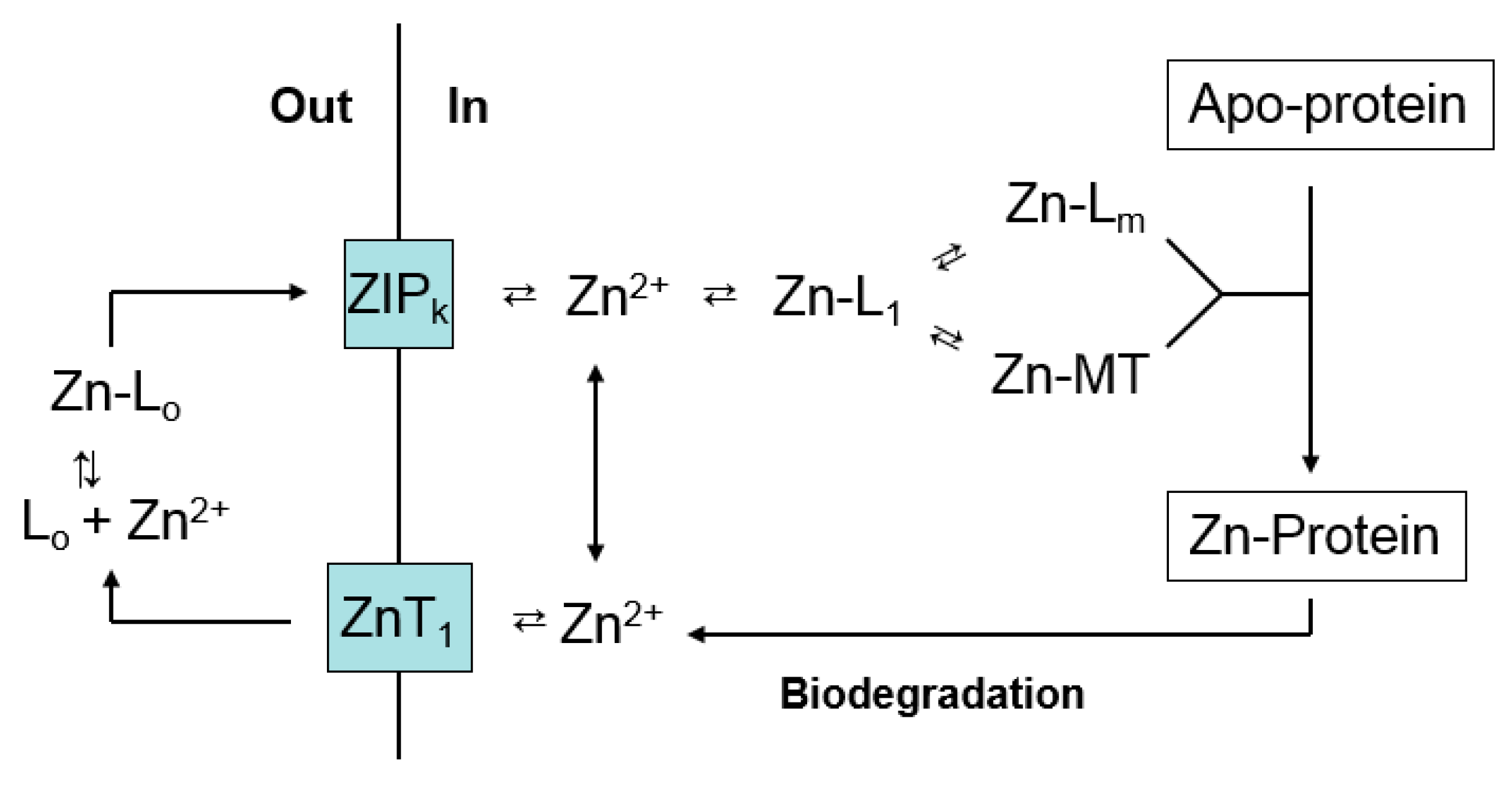
A number of studies demonstrate that upon entering cells, Zn2+ faces a complex, potent ligand environment. This array of binding sites has been described as a buffer for free Zn2+ and a muffler to further reduce the availability of mobile Zn2+ [57]. Within this maze, pathways by which Zn2+ reaches apo-zinc proteins must also be present (Figure 2 and Figure 4). Although emphasis has been placed on metallothionein as a donor of Zn2+ to such structures, the survival of MT-null cells and organisms firmly indicates that other general routes of trafficking exist. With the additional recognition that both proteome and MT bind Zn2+ strongly, the involvement of proteomic-based Zn2+ trafficking must be considered. Nevertheless, the clear involvement of MT in the metabolism of zinc under a variety of circumstances, for example under conditions of cell proliferation or zinc deficiency as described above, points to the need to integrate both proteome and MT into a model of zinc trafficking. Figure 5 merges Figure 2 and Figure 4 to offer a hypothetical view of zinc trafficking that incorporates both proteome (L1-m) and MT as binding sites for Zn2+ as it traverses the cell from plasma membrane to apo-zinc protein.
The model considers MT as part of the proteomic manifold of ligands for Zn2+ and makes several assertions: (i) the pathway that utilizes L1-m can operate without metallothionein; (ii) L1-m and MT interact by ligand substitution during the handling of Zn2+; and (iii) Zn-MT has some special roles to play in the distribution and movement of intracellular Zn2+ that merit singling it out in the model. Examining each in turn, MT-null cells distribute Zn2+ satisfactorily; thus, proteomic pathways involving L1-m exist to mediate the intracellular movements of Zn2+. How might they be revealed and studied?
A zinc sensor has been devised that employs plasmid expressed apo-carbonic anhydrase (apo-CA) as the target binding site for Zn2+ and a sensitive fluorescent probe that selectively forms a ternary complex with Zn-CA [59]. Using apo-CA instead as a model apo-zinc protein, its reaction with Zn2+ and the fluorophore, dansylamide (DNSA), has been investigated in the presence of supernatant from LLC-PK1 cells (Mahim and Petering, unpublished):
Like apo-MT, apo-CA poorly competes with the native Zn-proteome for Zn2+ despite its favorable conditional stability constant of 1011.4 relative to at least some Zn-proteins [10,33,59]. Reaction (26) occurred under conditions in which the high affinity proteomic binding sites were in substantial excess of apo-CA, and virtually all of the added Zn2+ was initially bound to the proteome. Evidently, a kinetic pathway existed for tightly bound proteomic zinc to react with the target apo-protein.
Apo-CA + Proteomehigh affinity•Zn + DNSA ⥂ DNSA-Zn-CA + Proteomehigh affinity
Zn7-MT also donates Zn2+ to apo-CA and may be the preferable source when it is present in the cell [26,32]. Because proteome and MT both represent strong binding sites for Zn2+ that are kinetically reactive in ligand substitution reactions, the possibility must be considered that trafficking Zn2+ is distributed among these two pools. In a test of this idea, LLC-PK1 cells were incubated for 24 h with elevated Zn2+ (30 µM) to stimulate metallothionein synthesis without compromising proliferation (Mahim and Petering, unpublished). Then, the distribution of Zn2+ within cell supernatant was determined. As expected, MT-bound zinc was observed. In addition, the amount of Zn2+ in the proteome fraction increased and constituted 25% of the extra metal ion that was transported into the cells. At least in this circumstance, proteome as well as MT participates in the trafficking of zinc. Whether they represent independent pools or are coupled through metal exchange remains to be determined.
11. Emerging Technology to Address Zinc Trafficking
A metallothionein-proteome axis seems to be emerging as the basis for zinc trafficking. To achieve a more refined understanding of the roles of each, studies at the level of individual proteins will be needed. A promising analytical approach links laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) with high-resolution separation of metalloproteins by polyacrylamide gel electrophoresis (PAGE) [11]. The methodology has foundered because of the lack of an adequate, native method of separation. One- or two-dimensional sodium dodecylsulfate PAGE (SDS-PAGE) methods provide excellent separation of proteins but do so at the cost of their denaturation. Recently, the formulation of native SDS-PAGE (NSDS-PAGE) demonstrated that high-resolution protein separation can be maintained together with retention of protein-bound metals [113]. This has raised hopes that LA-ICP-MS can be used as a highly sensitive, isotope specific monitor of protein-bound Zn2+.
At the same time, a means of probing Zn2+ bound adventitiously to the proteome has emerged from observations that the zinc fluorescent sensors, TSQ and Zinquin (ZQ), primarily detect Zn-proteins in cells through the formation of ternary adducts [114,115,116].
Zn-Protein + ZQ/TSQ ⥂ TSQ/ZQ-Zn-Protein
Moreover, when extra Zn2+ was added to the proteome, both TSQ and ZQ formed adducts with non-specifically bound zinc [51,117]. Using this information, an affinity bead has been fashioned that employs ZQ as the ligand [118]. With this tool, proteomic samples highly enriched in Zn-proteins have been isolated. Extending this method to the isolation of protein•Zn complexes should provide new options for studying zinc trafficking and the interaction of members of the proteome with metallothionein.
Acknowledgments
The authors acknowledge the support of NIH grant ES-024509 and a Research Growth Initiative grant from the University of Wisconsin–Milwaukee.
Author Contributions
David H. Petering conceived of and wrote the article. Afsana Mahim contributed important unpublished information to the key section, “Interaction of Metallothionein and the Proteome during Zinc Trafficking”, that provided the framework for that section.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the three domains of life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.; Karim, M.; Petering, D. Zinc proteomics. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Scott, R., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Tepaamorndech, S. The SLC30 family of zinc transporters—A review of current understanding of their biological and pathophysiological roles. Mol. Asp. Med. 2013, 34, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kambe, T. The functions of metallothionein and ZIP and ZnT transporters: An overview and perspective. Int. J. Mol. Sci. 2016, 17, 336. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Kambe, T. Molecular and genetic features of zinc transporters in physiology and pathogenesis. Metallomics 2011, 3, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiologic regulation. Annu. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, T.; Ohrvik, H.; Thiele, D.J. Charting the travels of copper in eukaryotes from yeast to mammals. Biochim. Biophys. Acta 2012, 1823, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Li, Y. Coordination dynamics of zinc in proteins. Chem. Rev. 2009, 109, 4682–4707. [Google Scholar] [CrossRef] [PubMed]
- Sussulini, A.; Becker, J.S. Combination of PAGE and LA-ICP-MS as an analytical workflow in metallomics: State of the art, new quantification strategies, advantages and limitations. Metallomics 2011, 3, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Sigel, A.; Sigel, H.; Sigel, R.O. Metallothioneins and related chelators. In Metal Ions in Life Sciences; RSC Publishing: Cambridge, UK, 2009; Volume 5. [Google Scholar]
- Grzywacz, A.; Gdula-Argasińska, J.; Muszyńska, B.; Tyszka-Czochara, M.; Librowski, T.; Opoka, W. Metal responsive transcription factor 1 (MTF-1) regulates zinc dependent cellular processes at the molecular level. Acta Biochim. Pol. 2015, 62, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Aras, M.A.; Aizenman, E. Redox regulation of intracellular zinc: Molecular signaling in the life and death of neurons. Antioxid. Redox Signal. 2011, 15, 2249–2263. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Deagen, J.T.; Whanger, P.D.; Weswig, P.H. Biological Function of Metallothionein. V. Its induction in rats by various stresses. Am. J. Physiol. 1978, 234, E282–E285. [Google Scholar] [PubMed]
- Petering, D.H.; Loftsgaarden, J.; Schneider, J.; Fowler, B. Metabolism of cadmium, zinc and copper in the rat kidney: The role of metallothionein and other binding sites. Environ. Health Perspect. 1984, 54, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K.; Lee, D.K.; Ravindra, R.; Lichtlen, P.; Sirito, M.; Sawadogo, M.; Schaffner, W. The transcription factors MTF-1 and USF1 cooperate to regulate mouse metallothionein-I expression in response to the essential metal zinc in visceral endoderm cells during early development. EMBO J. 2001, 20, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.Ø.; Larsen, A.; Stoltenberg, M.; Penkowa, M. The Role of metallothionein in oncogenesis and cancer prognosis. Prog. Histochem. Cytochem. 2009, 44, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Robbins, A.H.; McRee, D.E.; Williamson, M.; Collett, S.A.; Xuong, N.H.; Furey, W.F.; Wang, B.C.; Stout, C.D. Refined crystal structure of Cd, Zn metallothionein at 2.0 A resolution. J. Mol. Biol. 1991, 221, 1269–1293. [Google Scholar] [PubMed]
- Petering, D.H.; Krezoski, S.; Tabatabai, N.M. Metallothionein toxicology: Metal ion trafficking and Cellular protection. In Metallothioneins and Related Chelators: Metal Ions in Life Sciences; Sigel, A., Sigel, H., Sigel, R.O., Eds.; RSC Publishing: Cambridge, UK, 2009; Volume 5, pp. 353–397. [Google Scholar]
- Quesada, A.R.; Byrnes, R.W.; Krezoski, S.O.; Petering, D.H. Direct reaction of H2O2 with sulfhydryl groups in HL-60 cells: Zinc-metallothionein and other sites. Arch. Biochem. Biophys. 1996, 334, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Lazo, J.S.; Kuo, S.M.; Woo, E.S.; Pitt, B.R. The protein thiol metallothionein as an antioxidant and protectant against antineoplastic drugs. Chem. Biol. Interact. 1998, 111, 255–262. [Google Scholar] [CrossRef]
- Otvos, J.D.; Armitage, I.M. Structure of the metal clusters in rabbit liver metallothionein. Proc. Natl. Acad. Sci. USA 1980, 77, 7094–7098. [Google Scholar] [CrossRef] [PubMed]
- Nielson, K.B.; Winge, D.R. Order of metal binding in metallothionein. J. Biol. Chem. 1983, 258, 13063–13069. [Google Scholar] [PubMed]
- Ejnik, J.; Robinson, J.; Zhu, J.; Försterling, H.; Shaw, C.F.; Petering, D.H. Folding pathway of Apo-metallothionein induced by Zn2+, Cd2+ and Co2+. J. Inorg. Biochem. 2002, 88, 144–152. [Google Scholar] [CrossRef]
- Li, T.Y.; Kraker, A.J.; Shaw, C.F., III; Petering, D.H. Ligand substitution reactions of metallothioneins with EDTA and Apo-carbonic anhydrase. Proc. Natl. Acad. Sci. USA 1980, 77, 6334–6338. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Y.; Minkel, D.T.; Shaw, C.F., III; Petering, D.H. On the reactivity of metallothioneins with 5,5′-dithiobis-(2-nitrobenzoic acid). Biochem. J. 1981, 193, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.F., III; Savas, M.M.; Petering, D.H. Ligand substitution and sulfhydryl reactivity of metallothionein. Methods Enzymol. 1991, 205, 401–414. [Google Scholar] [PubMed]
- Gan, T.; Munoz, A.; Shaw, C.F., III; Petering, D.H. Reaction of 111Cd7-metallothionein with EDTA. A Reappraisal. J. Biol. Chem. 1995, 270, 5339–5345. [Google Scholar] [CrossRef] [PubMed]
- Petering, D.H.; Shaw, C.F., III. Stability constants and related equilibrium properties of metallothioneins. Methods Enzymol. 1991, 205, 475–484. [Google Scholar] [PubMed]
- Namdarghanbari, M.A.; Meeusen, J.; Bachowski, G.; Giebel, N.; Johnson, J.; Petering, D.H. Reaction of the zinc sensor FluoZin-3 with Zn7-metallothionein: Inquiry into the existence of a proposed weak binding site. J. Inorg. Biochem. 2010, 104, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Pinter, T.B.; Stillman, M.J. The zinc balance: Competitive zinc metalation of carbonic anhydrase and metallothionein 1A. Biochemistry 2014, 53, 6276–6285. [Google Scholar] [CrossRef] [PubMed]
- Kochańczyk, T.; Drozd, A.; Krężel, A. Relationship between the architecture of zinc coordination and zinc binding affinity in proteins—Insights into zinc regulation. Metallomics 2015, 7, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, L.L.; Krebs, J.F.; Paterno, S.A.; Fierke, C.A. Engineering a cysteine ligand into the zinc binding site of human carbonic anhydrase II. Biochemistry 1993, 32, 9896–9900. [Google Scholar] [CrossRef] [PubMed]
- Udom, A.O.; Brady, F.O. Reactivation in vitro of zinc-requiring Apo-enzymes by rat liver zinc-thionein. Biochem. J. 1980, 187, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Zaia, J.; Fabris, D.; Wei, D.; Karpel, R.L.; Fenselau, C. Monitoring metal ion flux in reactions of metallothionein and drug-modified metallothionein by electrospray mass spectrometry. Protein Sci. 1998, 7, 2398–2404. [Google Scholar] [CrossRef] [PubMed]
- Roesijadi, G.; Bogumil, R.; Vasák, M.; Kägi, J.H. Modulation of DNA binding of a tramtrack zinc finger peptide by the metallothionein-thionein conjugate pair. J. Biol. Chem. 1998, 273, 17425–17432. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Maret, W.; Vallee, B.L. Control of zinc transfer between thionein, metallothionein, and zinc proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 3489–3494. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.J.; Maret, W.; Vallee, B.L. The glutathione redox couple modulates zinc transfer from metallothionein to zinc-depleted sorbitol dehydrogenase. Proc. Natl. Acad. Sci. USA 1998, 95, 3483–3488. [Google Scholar] [CrossRef] [PubMed]
- Krezel, A.; Maret, W. Dual nanomolar and picomolar Zn(II) binding properties of metallothionein. J. Am. Chem. Soc. 2007, 129, 10911–10921. [Google Scholar] [CrossRef] [PubMed]
- Vasák, M.; Kägi, J.H. Metal thiolate clusters in Cobalt(II)-metallothionein. Proc. Natl. Acad. Sci. USA 1981, 78, 6709–6713. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, P.M.; You, C.; Dallinger, R.; Gruber, C.; Brouwer, M.; Kägi, J.H.; Hunziker, P.E. Electrospray ionization mass spectrometry of zinc, cadmium, and copper metallothioneins: Evidence for metal-binding cooperativity. Protein Sci. 2000, 9, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.E.; Summers, K.L.; Stillman, M.J. Noncooperative metalation of metallothionein 1A and its isolated domains with zinc. Biochemistry 2012, 51, 6690–6700. [Google Scholar] [CrossRef] [PubMed]
- Pinter, T.B.; Stillman, M.J. Putting the pieces into place: Properties of intact zinc metallothionein 1A determined from interaction of its isolated domains with carbonic anhydrase. Biochem. J. 2015, 471, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Otvos, J.D.; Petering, D.H.; Shaw, C.F. Structure-reactivity relationships of metallothionein, a unique metal binding protein. Comments Inorg. Chem. 1989, 9, 1–35. [Google Scholar] [CrossRef]
- Krezel, A.; Maret, W. Zinc-buffering capacity of a eukaryotic cell at physiological pZn. J. Biol. Inorg. Chem. 2006, 11, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Zinc Transporters and the cellular trafficking of zinc. Biochim. Biophys. Acta 2006, 1763, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Petering, D.H.; Kothinti, R.; Meeusen, J.; Rana, U. Cellular inorganic chemistry concepts and examples. In Cellular and Molecular Biology of Metals; Zalups, R.K., Koropatnick, J., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2010; Chapter 1; pp. 1–35. [Google Scholar]
- Karim, M.R.; Petering, D.H. Newport green, a fluorescent sensor of weakly bound cellular Zn2+: Competition with proteome for Zn2+. Metallomics 2016, 8, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Krezel, A.; Hao, Q.; Maret, W. The zinc/thiolate redox biochemistry of metallothionein and the control of zinc ion fluctuations in cell signaling. Arch. Biochem. Biophys. 2007, 463, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.R.; Petering, D.H. Detection of Zn2+ release in nitric oxide treated cells and proteome: Dependence on fluorescent sensor and proteomic sulfhydryl groups. Metallomics 2016, 9, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.J.; Ahner, B.A. Determination of stability constants of Cu(I), Cd(II) & Zn(II) complexes with thiols using fluorescent probes. J. Inorg. Biochem. 2013, 128, 12–23. [Google Scholar]
- Namdarghanbari, M.A.; Bertling, J.; Krezoski, S.; Petering, D.H. Toxic metal proteomics: Reaction of the mammalian zinc proteome with Cd2+. J. Inorg. Biochem. 2014, 136, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Bozym, R.A.; Chimienti, F.; Giblin, L.J.; Gross, G.W.; Korichneva, I.; Li, Y.; Libert, S.; Maret, W.; Parviz, M.; Frederickson, C.J.; et al. Free zinc ions outside a narrow concentration range are toxic to a variety of Cells in vitro. Exp. Biol. Med. 2010, 235, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Pluth, M.D.; Tomat, E.; Lippard, S.J. Biochemistry of mobile zinc and nitric oxide revealed by fluorescent sensors. Annu. Rev. Biochem. 2011, 80, 333–355. [Google Scholar] [CrossRef] [PubMed]
- Carter, K.P.; Young, A.M.; Palmer, A.E. Fluorescent sensors for measuring metal ions in living systems. Chem. Rev. 2014, 114, 4564–4601. [Google Scholar] [CrossRef] [PubMed]
- Colvin, R.A.; Holmes, W.R.; Fontaine, C.P.; Maret, W. Cytosolic zinc buffering and muffling: Their role in intracellular zinc homeostasis. Metallomics 2010, 2, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Simons, T.J. Intracellular free zinc and zinc buffering in human red blood cells. J. Membr. Biol. 1991, 123, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Bozym, R.A.; Thompson, R.B.; Stoddard, A.K.; Fierke, C.A. Intracellular exchangeable zinc in PC-12 cells using a ratiometric fluorescence biosensor. ACS Chem. Biol. 2006, 1, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Kagi, J.H.; Vallee, B.L. Metallothionein: A cadmium- and zinc-containing protein from equine renal cortex. J. Biol. Chem. 1960, 235, 3460–3465. [Google Scholar] [PubMed]
- Chen, R.W.; Vasey, E.J.; Whanger, P.D. Accumulation and depletion of zinc in rat liver and kidney metallothioneins. J. Nutr. 1977, 107, 805–813. [Google Scholar] [PubMed]
- Klein, D.; Scholz, P.; Drasch, G.A.; Müller-Höcker, J.; Summer, K.H. Metallothionein, copper and zinc in fetal and neonatal human liver: Changes during development. Toxicol. Lett. 1991, 56, 61–67. [Google Scholar] [CrossRef]
- Munger, K.; Germann, U.A.; Beltramini, M.; Niedermann, D.; Baitella-Eberlej, G.; Kagi, J.H.R.; Lerch, K. (Cu,Zn)-metallothioneins from fetal bovine liver: chemical and spectroscopic properties. J. Biol. Chem. 1985, 260, 10032–10038. [Google Scholar] [PubMed]
- Zhu, J.; Meeusen, J.; Krezoski, S.; Petering, D.H. Reactivity of Zn-, Cd-, and Apo-metallothionein with nitric oxide compounds: In vitro and cellular comparison. Chem. Res. Toxicol. 2010, 23, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Redox biochemistry of mammalian metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.E.; Choo, K.H.A. Targeting and Germ-line transmission of a null mutation at the metallothionein I and II loci in mouse. Proc. Natl. Acad. Sci. USA 1993, 90, 8088–8092. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Liu, J.; Choo, K.H.; Michalska, A.E.; Klaassen, C.D. Metallothionein-I and -II knock-out mice are sensitive to cadmium-induced liver mRNA expression of c-Jun and p53. Toxicol. Appl. Pharmacol. 1996, 136, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Eddins, D.; Petro, A.; Pollard, N.; Freedman, J.H.; Levin, E.D. Mercury-induced cognitive impairment in metallothionein-1/2 null mice. Neurotoxicol. Teratol. 2008, 30, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Lazo, J.S.; Kondo, Y.; Dellapiazza, D.; Michalska, A.E.; Choo, K.H.; Pitt, B.R. Enhanced sensitivity to oxidative stress in cultured embryonic cells from transgenic mice deficient in metallothionein I and II genes. J. Biol. Chem. 1995, 270, 5506–5510. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Woo, E.S.; Michalska, A.E.; Choo, K.H.; Lazo, J.S. Metallothionein null cells have increased sensitivity to anticancer drugs. Cancer Res. 1995, 55, 2021–2023. [Google Scholar] [PubMed]
- Andrews, G.K.; Geiser, J. Expression of the mouse metallothionein-I and -II genes provides a reproductive advantage during maternal dietary zinc deficiency. J. Nutr. 1999, 129, 1643–1648. [Google Scholar] [PubMed]
- Fong, L.; Tan, K.; Tran, C.; Cool, J.; Scherer, M.A.; Elovaris, R.; Coyle, P.; Foster, B.K.; Rofe, A.M.; Xian, C.J. Interaction of dietary zinc and intracellular binding protein metallothionein in postnatal bone growth. Bone 2009, 44, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Perraut, C.; Pollard, N.; Freedman, J.H. Metallothionein expression and neurocognitive function in mice. Physiol. Behav. 2006, 87, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.H.; Wood, A.M.; Newman, A.M.; Bremner, I.; Choo, K.H.A.; Michalska, A.E.; Duncan, S.; Trayhurn, P. Obesity and hyperleptinemia in metallothionein (-I and -II) null mice. Proc. Natl. Acad. Sci. USA 1998, 95, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mageed, A.B.; Agrawal, K.C. Antisense down-regulation of metallothoinein induces growth arrest and apoptosis in human breast carcinoma cells. Cancer Gene Ther. 1997, 4, 199–207. [Google Scholar] [PubMed]
- Lim, D.; Jocelyn, K.M.; Yip, G.W.; Bay, B.H. Silencing the metallothionein-2A gene inhibits cell cycle progression from G1- to S-phase involving ATM and cdc25A signaling in breast cancer cells. Cancer Lett. 2009, 276, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Tekur, S.; Ho, S.M. Ribozyme-mediated downregulation of human metallothionein II(a) Induces apoptosis in human prostate and ovarian cancer cell lines. Mol. Carcinog. 2002, 33, 44–55. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.S. The role of zinc in growth and cell proliferation. J. Nutr. 2000, 130, 1500S–1508S. [Google Scholar] [PubMed]
- Petering, H.G.; Buskirk, H.H.; Crim, J.A. The effect of dietary mineral supplements of the rat on the antitumor activity of 3-ethoxy-2-oxobutyraldehyde bis(thiosemicarbazone). Cancer Res. 1967, 27, 1115–1121. [Google Scholar] [PubMed]
- Minkel, D.T.; Dolhun, P.J.; Calhoun, B.L.; Saryan, L.A.; Petering, D.H. Zinc deficiency and growth of ehrlich ascites tumor. Cancer Res. 1979, 39, 2451–2456. [Google Scholar] [PubMed]
- Saryan, L.A.; Minkel, D.T.; Dolhun, P.J.; Calhoun, B.L.; Wielgus, S.; Schaller, M.; Petering, D.H. Effects of zinc deficiency on cellular processes and morphology in ehrlich ascites tumor cells. Cancer Res. 1979, 39, 2457–2465. [Google Scholar] [PubMed]
- Mita, M.; Imura, N.; Kumazawa, Y.; Himeno, S. Suppressed proliferative response of spleen T cells from metallothionein null mice. Microbiol. Immunol. 2002, 46, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.R.; Mara, T.W.; Cherian, M.G. Impaired hepatic regeneration in metallothionein-I/II knockout mice after partial hepatectomy. Exp. Biol. Med. 2005, 230, 61–67. [Google Scholar]
- Kraker, A.J.; Petering, D.H. Tumor-host zinc metabolism: The central role of metallothionein. Biol. Trace Elem. Res. 1983, 5, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Krezoski, S.K.; Villalobos, J.; Shaw, C.F., III; Petering, D.H. Kinetic lability of zinc bound to metallothionein in ehrlich cells. Biochem. J. 1988, 255, 483–491. [Google Scholar] [PubMed]
- Kraker, A.J.; Krakower, G.; Shaw, C.F., III; Petering, D.H.; Garvey, J.S. Zinc metabolism in ehrlich cells: Properties of a metallothionein-like zinc-binding protein. Cancer Res. 1988, 48, 3381–3388. [Google Scholar] [PubMed]
- Suhy, D.A.; Simon, K.D.; Linzer, D.I.; O’Halloran, T.V. Metallothionein is part of a zinc-scavenging mechanism for cell survival under conditions of extreme zinc deprivation. J. Biol. Chem. 1999, 274, 9183–9192. [Google Scholar] [CrossRef] [PubMed]
- Langmade, S.J.; Ravindra, R.; Daniels, P.J.; Andrews, G.K. The transcription factor MTF-1 mediates metal regulation of the mouse ZnT1 gene. J. Biol. Chem. 2000, 275, 34803–34809. [Google Scholar] [CrossRef] [PubMed]
- Braun, W.; Vasák, M.; Robbins, A.H.; Stout, C.D.; Wagner, G.; Kägi, J.H.; Wüthrich, K. Comparison of the NMR solution structure and the X-ray crystal structure of Rat metallothionein-2. Proc. Natl. Acad. Sci. USA 1992, 89, 10124–10128. [Google Scholar] [CrossRef] [PubMed]
- Pattanaik, A.; Shaw, C.F., III; Petering, D.H.; Garvey, J.; Kraker, A.J. Basal metallothionein in tumors: widespread presence of apoprotein. J. Inorg. Biochem. 1994, 54, 91–105. [Google Scholar] [CrossRef]
- Yang, Y.; Maret, W.; Vallee, B.L. Differential fluorescence labeling of cysteinyl clusters uncovers high tissue levels of thionein. Proc. Natl. Acad. Sci. USA 2001, 98, 5556–5559. [Google Scholar] [CrossRef] [PubMed]
- Petering, D.H.; Zhu, J.; Krezoski, S.; Meeusen, J.; Kiekenbush, C.; Krull, S.; Specher, T.; Dughish, M. Apo-metallothionein emerging as a major player in the cellular activities of metallothionein. Exp. Biol. Med. 2006, 231, 1528–1534. [Google Scholar] [CrossRef]
- Petering, D.H.; Quesada, A.; Dughish, M.; Eisch, S.; Gan, T.; Lemkuil, D.; Pattanaik, A.; Byrnes, R.W.; Savas, M.; Whelan, H.; et al. Metallothionein in tumor and host: intersections of zinc metabolism, the stress response, and tumor therapy. In Metallothionein III, Biological Roles and Medical Implications; Suzuki, K., Imura, N., Kimura, M., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1993; pp. 329–346. [Google Scholar]
- Irvine, G.W.; Duncan, K.E.; Gullons, M.; Stillman, M.J. Metalation kinetics of the human α-metallothionein 1A fragment is dependent on the fluxional structure of the Apo-protein. Chemistry 2015, 21, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Irvine, G.W.; Santolini, M.; Stillman, M.J. Selective cysteine modification of metal-free human metallothionein 1A and its isolated domain fragments: Solution structural properties revealed via ESI-MS. Protein Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rana, U.; Kothinti, R.; Meeusen, J.; Tabatabai, N.M.; Krezoski, S.; Petering, D.H. Zinc binding ligands and cellular zinc trafficking: Apo-metallothionein, glutathione, TPEN, proteomic zinc, and Zn-Sp1. J. Inorg. Biochem. 2008, 102, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Ejnik, J.; Muñoz, A.; Gan, T.; Shaw, C.F., III; Petering, D.H. Interprotein metal ion exchange between cadmium-carbonic anhydrase and Apo- or zinc-metallothionein. J. Biol. Inorg. Chem. 1999, 4, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Kothinti, R.; Tabatabai, N.M.; Petering, D.H. Electrophoretic mobility shift assay of zinc finger proteins: competition for Zn2+ bound to Sp1 in protocols including EDTA. J. Inorg. Biochem. 2011, 105, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Maret, W. Catalytic selenols couple the redox cycles of metallothionein and glutathione. Eur. J. Biochem. 2001, 268, 3346–3353. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Tachibana, C.; Rahr Winther, J.; Appenzeller-Herzog, C. Intracellular glutathione pools are heterogeneously concentrated. Redox Biol. 2013, 1, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Savas, M.M.; Shaw, C.F., III; Petering, D.H. The oxidation of rabbit liver metallothionein-II by 5,5′-dithiobis(2-nitrobenzoic acid) and glutathione disulfide. J. Inorg. Biochem. 1993, 52, 235–249. [Google Scholar] [CrossRef]
- Hamer, D.H. Metallothionein. Annu. Rev. Biochem. 1986, 55, 913–951. [Google Scholar] [CrossRef] [PubMed]
- Stuart, G.W.; Searle, P.F.; Palmiter, R.D. Identification of multiple metal regulatory elements in mouse metallothionein-I promoter by assaying synthetic sequences. Nature 1985, 317, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Radtke, F.; Heuchel, R.; Georgiev, O.; Hergersberg, M.; Gariglio, M.; Dembic, Z.; Schaffner, W. Cloned Transcription factor MTF-1 activates the mouse metallothionein I promoter. EMBO. J. 1993, 12, 1355–1362. [Google Scholar] [PubMed]
- Dalton, T.P.; Bittel, D.; Andrews, G.K. Reversible activation of mouse metal response element-binding transcription factor 1 DNA binding involves zinc interaction with the zinc finger domain. Mol. Cell. Biol. 1997, 17, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Guerrerio, A.L.; Berg, J.M. Metal ion affinities of the zinc finger domains of the metal responsive element-binding transcription factor-1 (MTF1). Biochemistry 2004, 43, 5437–5444. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, I.V.; Bittel, D.C.; Ravindra, R.; Jiang, H.; Andrews, G.K. Zinc and cadmium can promote rapid nuclear translocation of metal response element-binding transcription factor-1. J. Biol. Chem. 2000, 275, 9377–9784. [Google Scholar] [CrossRef] [PubMed]
- Woo, E.S.; Lazo, J.S. Nucleocytoplasmic functionality of metallothionein. Cancer Res. 1997, 57, 4236–4241. [Google Scholar] [PubMed]
- Bittel, D.; Dalton, T.; Samson, S.L.; Gedamu, L.; Andrews, G.K. The DNA binding activity of metal response element-binding transcription factor-1 is activated in vivo and in vitro by zinc, but not by other transition metals. J. Biol. Chem. 1998, 273, 7127–7133. [Google Scholar] [CrossRef] [PubMed]
- Krepkiy, D.; Försterling, F.H.; Petering, D.H. Interaction of Cd2+ with Zn finger 3 of transcription factor IIIA: Structures and binding to cognate DNA. Chem. Res. Toxicol. 2004, 17, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Georgiev, O.; Hagmann, M.; Günes, C.; Cramer, M.; Faller, P.; Vasák, M.; Schaffner, W. Activity of metal-responsive transcription factor 1 by toxic heavy metals and H2O2 in vitro is modulated by metallothionein. Mol. Cell. Biol. 2003, 23, 8471–8485. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.B.; Wobig, W.J.; Petering, D.H. Native SDS-PAGE: High resolution electrophoretic separation of proteins with retention of native properties including bound metal ions. Metallomics 2014, 6, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, J.W.; Tomasiewicz, H.; Nowakowski, A.; Petering, D.H. TSQ (6-Methoxy-8-p-toluenesulfonamido-quinoline), a common fluorescent sensor for cellular zinc, images zinc proteins. Inorg. Chem. 2011, 50, 7563–7573. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.B.; Petering, D.H. Reactions of the fluorescent sensor, zinquin, with the zinc-proteome: adduct formation and ligand substitution. Inorg. Chem. 2011, 50, 10124–10133. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, J.W.; Nowakowski, A.; Petering, D.H. Reaction of metal-binding ligands with the zinc proteome: Zinc sensors and N,N,N′,N′-tetrakis(2-pyridylmethyl) ethylenediamine. Inorg. Chem. 2012, 51, 3625–3632. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.B.; Meeusen, J.W.; Menden, H.; Tomasiewicz, H.; Petering, D.H. Chemical-biological properties of zinc sensors TSQ and zinquin: Formation of sensor-Zn-protein adducts versus Zn(sensor)2 complexes. Inorg. Chem. 2015, 54, 11637–11647. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A. Zinc Chemical Biology: The Pursuit of the Intracellular Targets of Zinquin; UWM Digital Commons: Milwaukee, WI, USA; University of Wisconsin Milwaukee: Milwaukee, WI, USA, 2013. [Google Scholar]
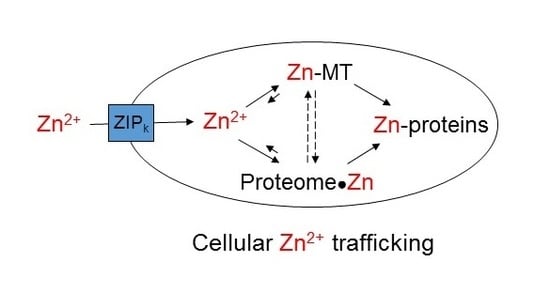
Figure 1.
Contemporary view of cellular zinc trafficking [6,13,14]. Transporters move Zn2+ into cells and between compartments. Metallothionein (MT) controls the concentration of free Zn2+ that supplies zinc to proteins. Zn-MT may also contribute to trafficking of Zn2+ to Zn-proteins.
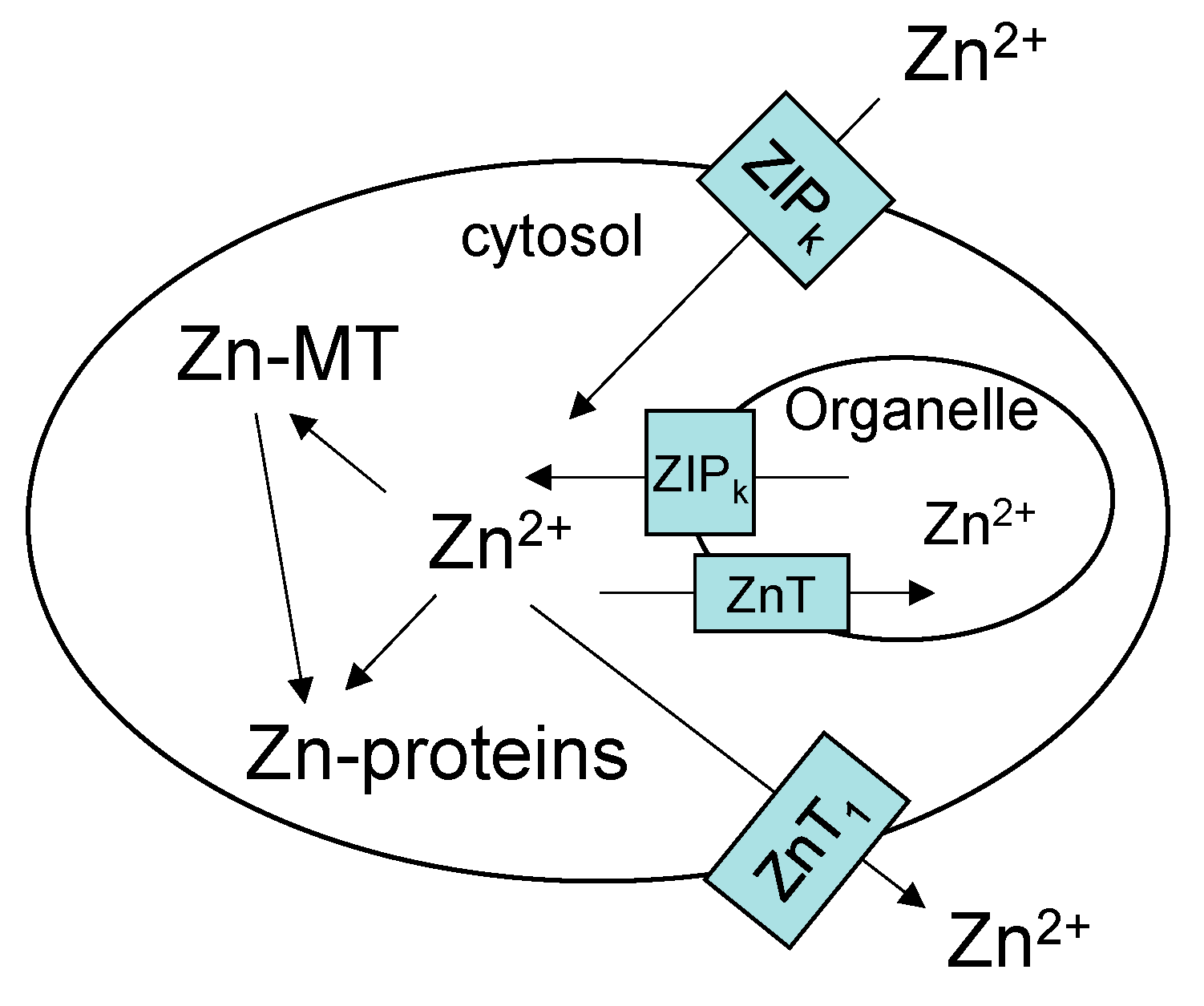
Figure 2.
Hypothetical Zn2+ trafficking pathway based on Figure 1.
Figure 2.
Hypothetical Zn2+ trafficking pathway based on Figure 1.
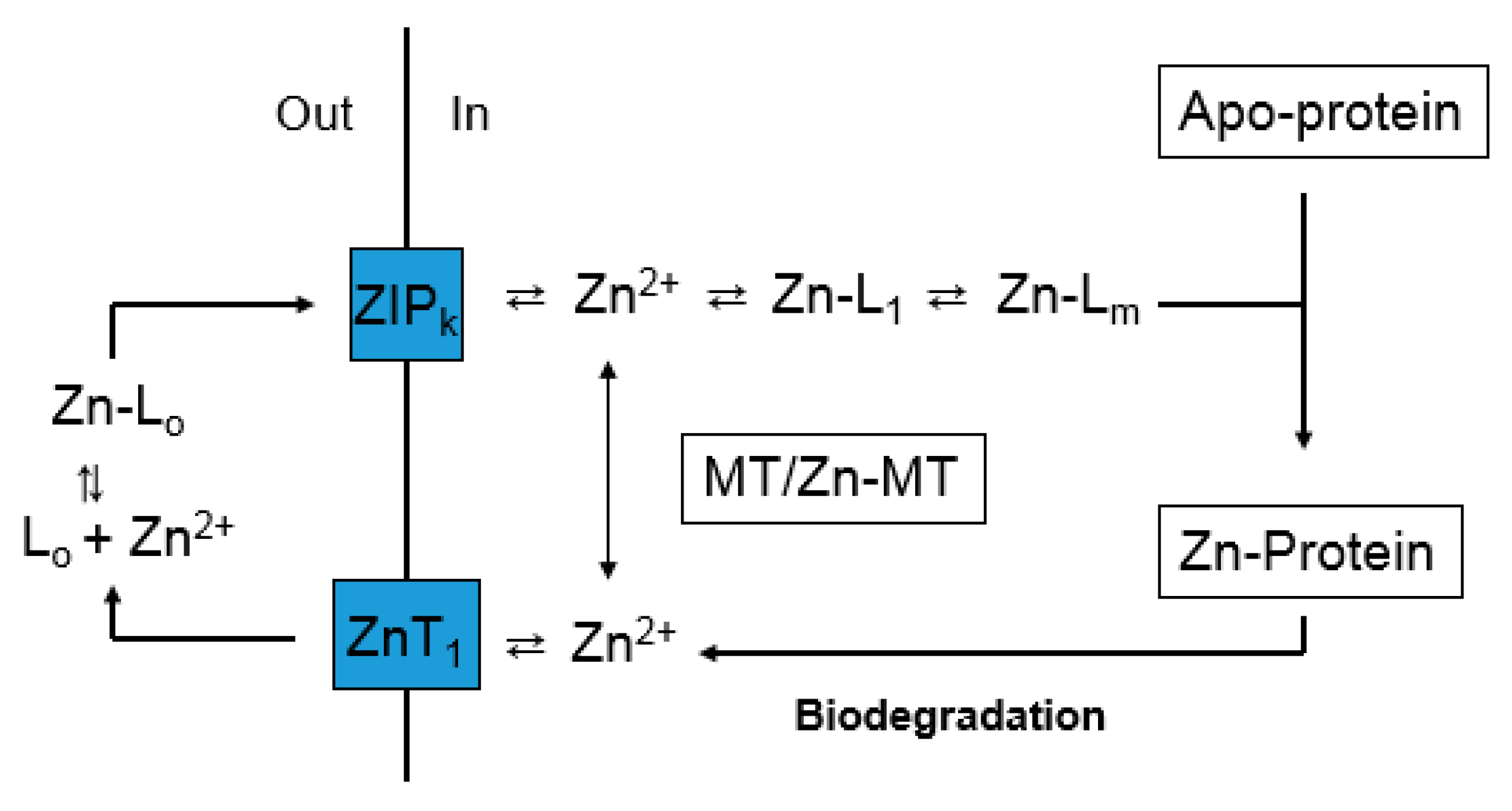
Figure 3.
Response of tumor and host pools of zinc to graded restoration of nutrient Zn (adapted from [87]). Mice were held on a Zn-deficient diet for 2 weeks prior to injection of tumor cells. Measurements were made 6–9 days after reintroduction of nutrient Zn. Relative total cells (○) and relative proteomic Zn (●), MT Zn (▲), and ascites fluid Zn (□) per cell or unit volume.
Figure 3.
Response of tumor and host pools of zinc to graded restoration of nutrient Zn (adapted from [87]). Mice were held on a Zn-deficient diet for 2 weeks prior to injection of tumor cells. Measurements were made 6–9 days after reintroduction of nutrient Zn. Relative total cells (○) and relative proteomic Zn (●), MT Zn (▲), and ascites fluid Zn (□) per cell or unit volume.
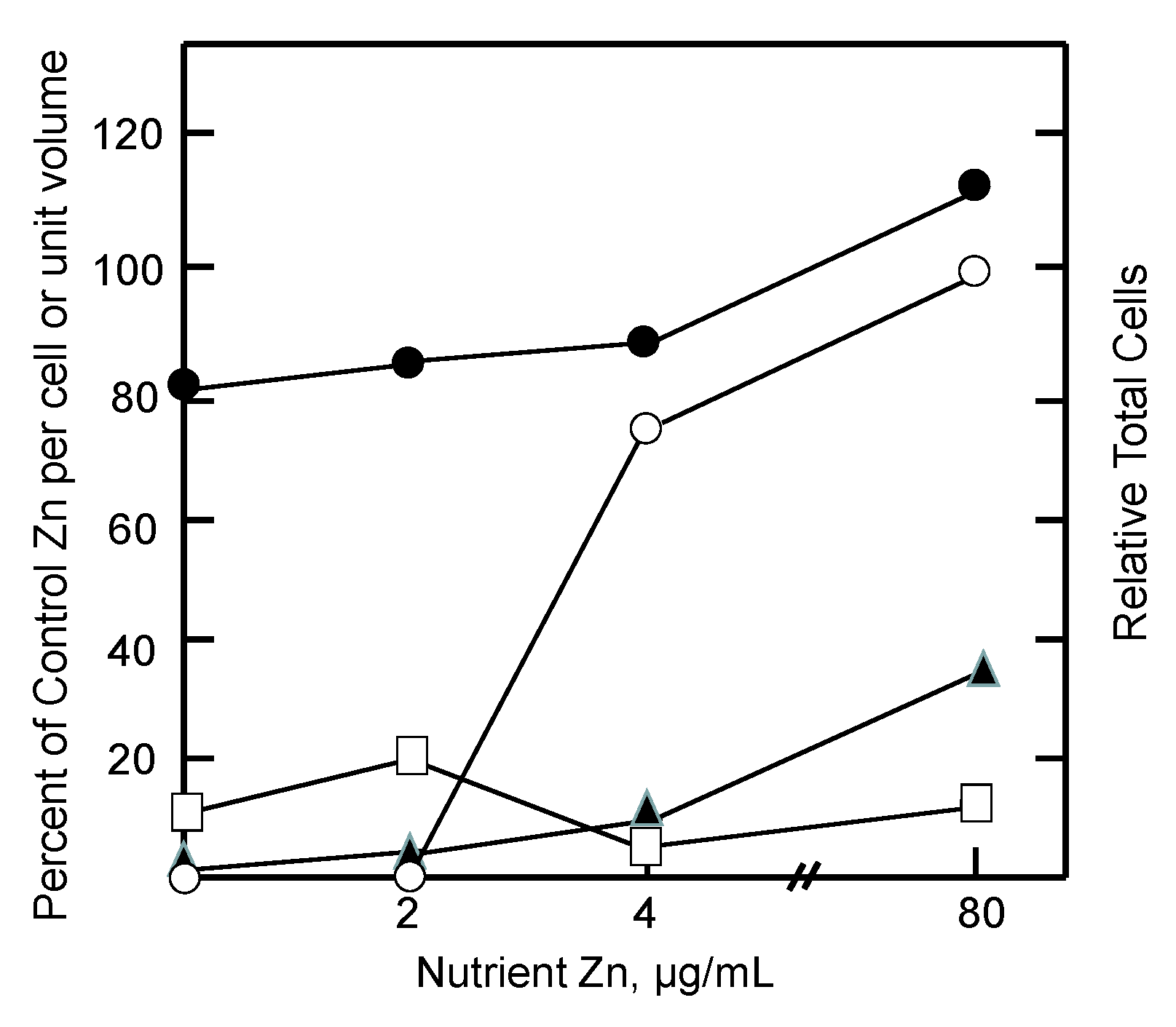
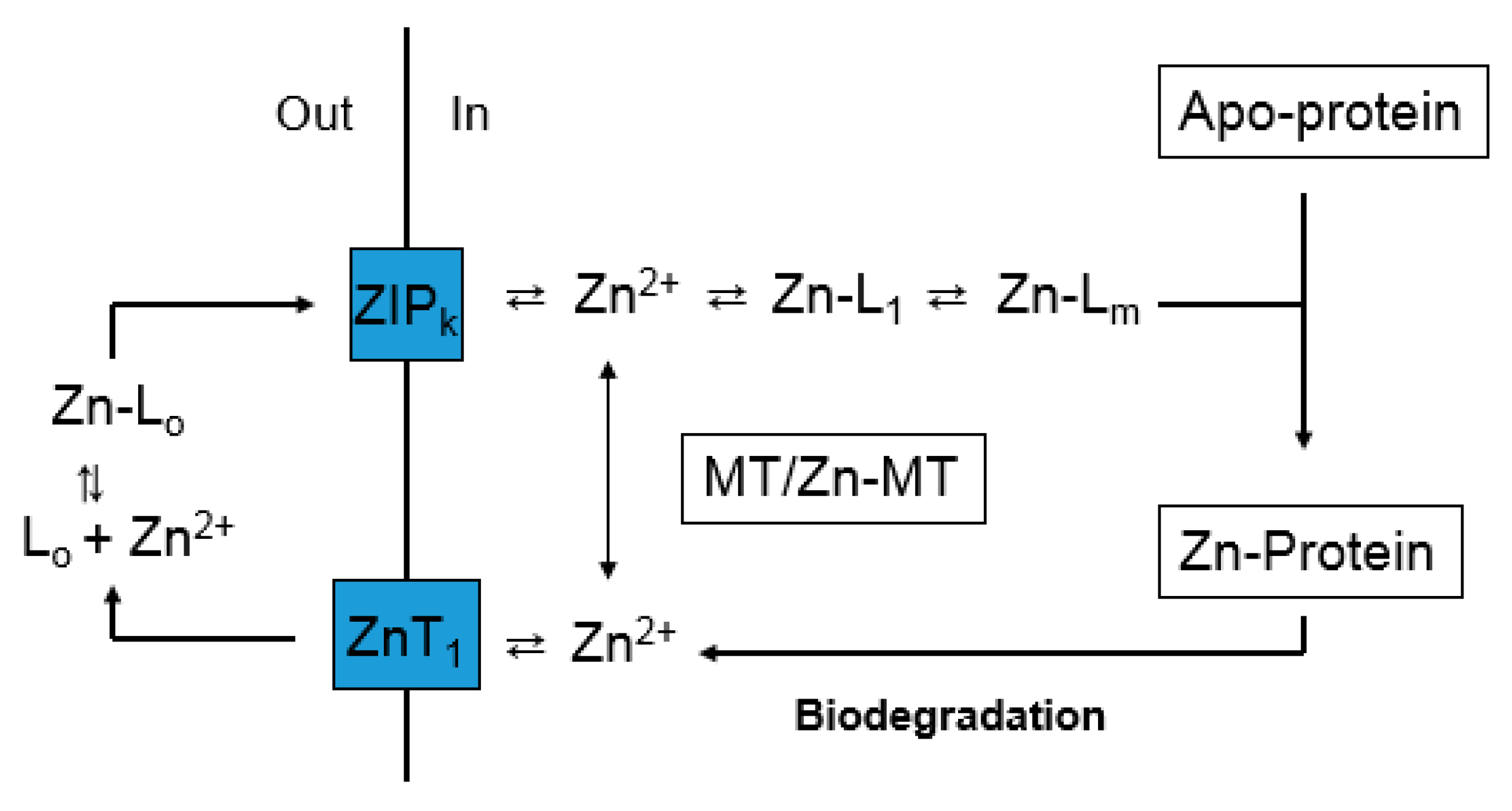
Figure 4.
Cellular Zn2+ trafficking involving metallothionein. Based on Figure 3, the pathway emphasizes Zn-MT, but does not exclude other Proteomic intermediates (L1-m). V1 and V2 represent, respectively, the rates of formation of Zn-MT from apo- or undersaturated MT and the donation of Zn2+ to apo-proteins by Zn-MT.
Figure 4.
Cellular Zn2+ trafficking involving metallothionein. Based on Figure 3, the pathway emphasizes Zn-MT, but does not exclude other Proteomic intermediates (L1-m). V1 and V2 represent, respectively, the rates of formation of Zn-MT from apo- or undersaturated MT and the donation of Zn2+ to apo-proteins by Zn-MT.
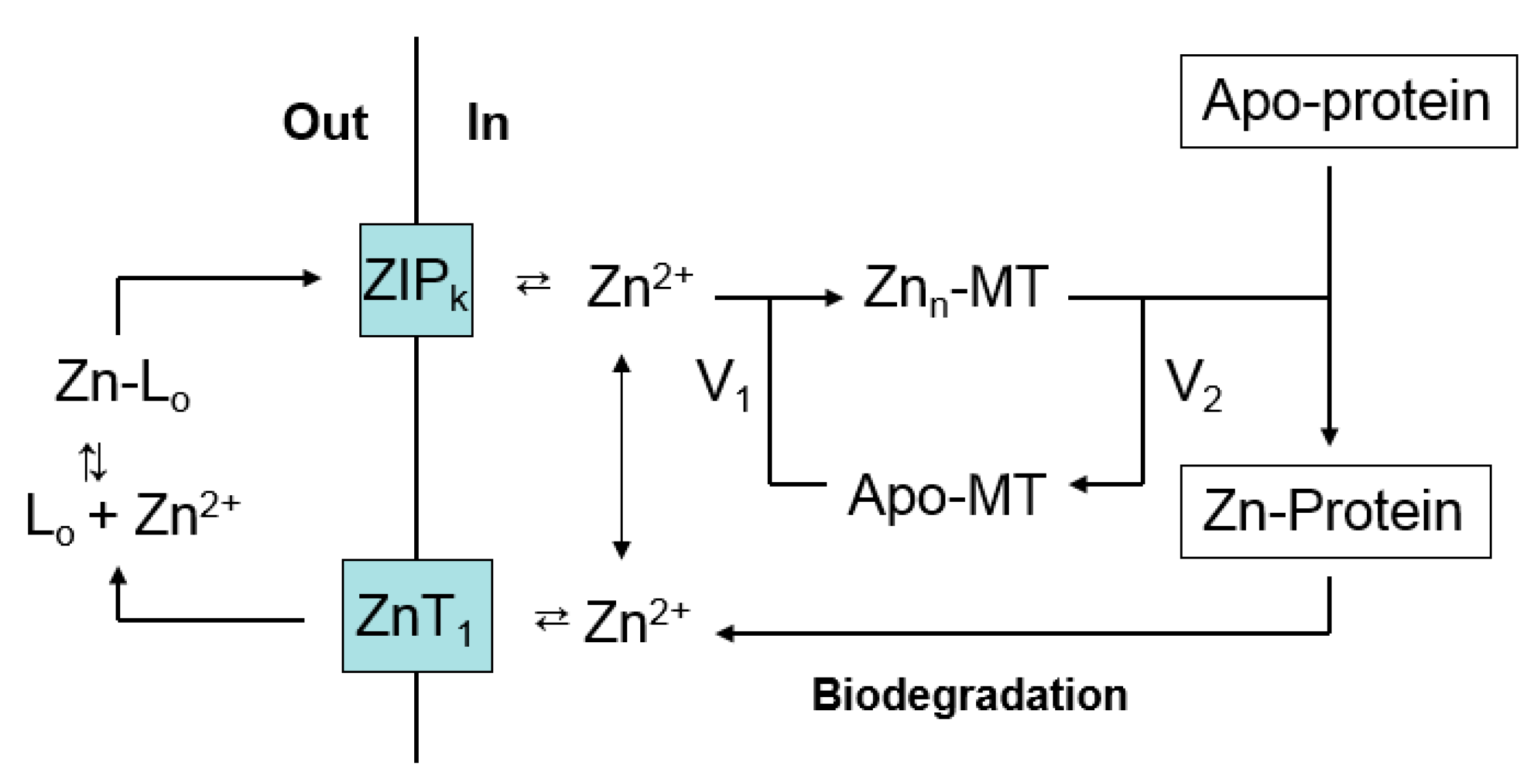
Figure 5.
Zinc trafficking linking proteomic and MT binding sites for Zn2+.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Petering, D.H.; Mahim, A. Proteomic High Affinity Zn2+ Trafficking: Where Does Metallothionein Fit in? Int. J. Mol. Sci. 2017, 18, 1289. https://doi.org/10.3390/ijms18061289
AMA Style
Petering DH, Mahim A. Proteomic High Affinity Zn2+ Trafficking: Where Does Metallothionein Fit in? International Journal of Molecular Sciences. 2017; 18(6):1289. https://doi.org/10.3390/ijms18061289
Chicago/Turabian StylePetering, David H., and Afsana Mahim. 2017. "Proteomic High Affinity Zn2+ Trafficking: Where Does Metallothionein Fit in?" International Journal of Molecular Sciences 18, no. 6: 1289. https://doi.org/10.3390/ijms18061289
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.