Plantago asiatica L. Seed Extract Improves Lipid Accumulation and Hyperglycemia in High-Fat Diet-Induced Obese Mice

Abstract
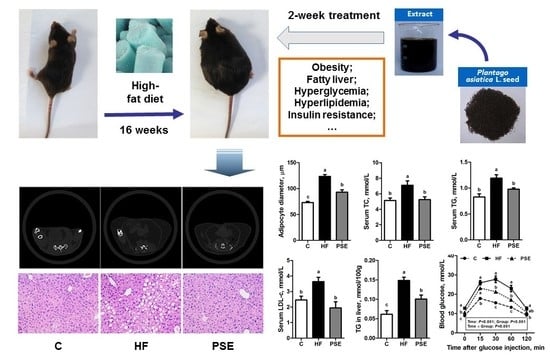
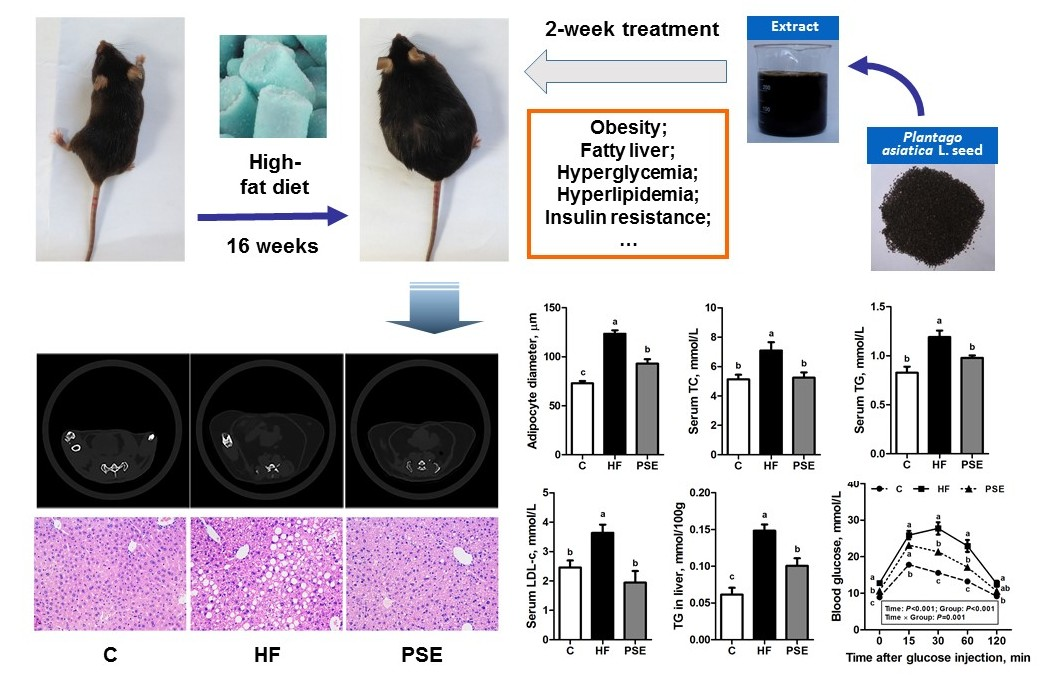
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Chemical Constituents of the Plantago asiatica L. Seed Extract (PSE)
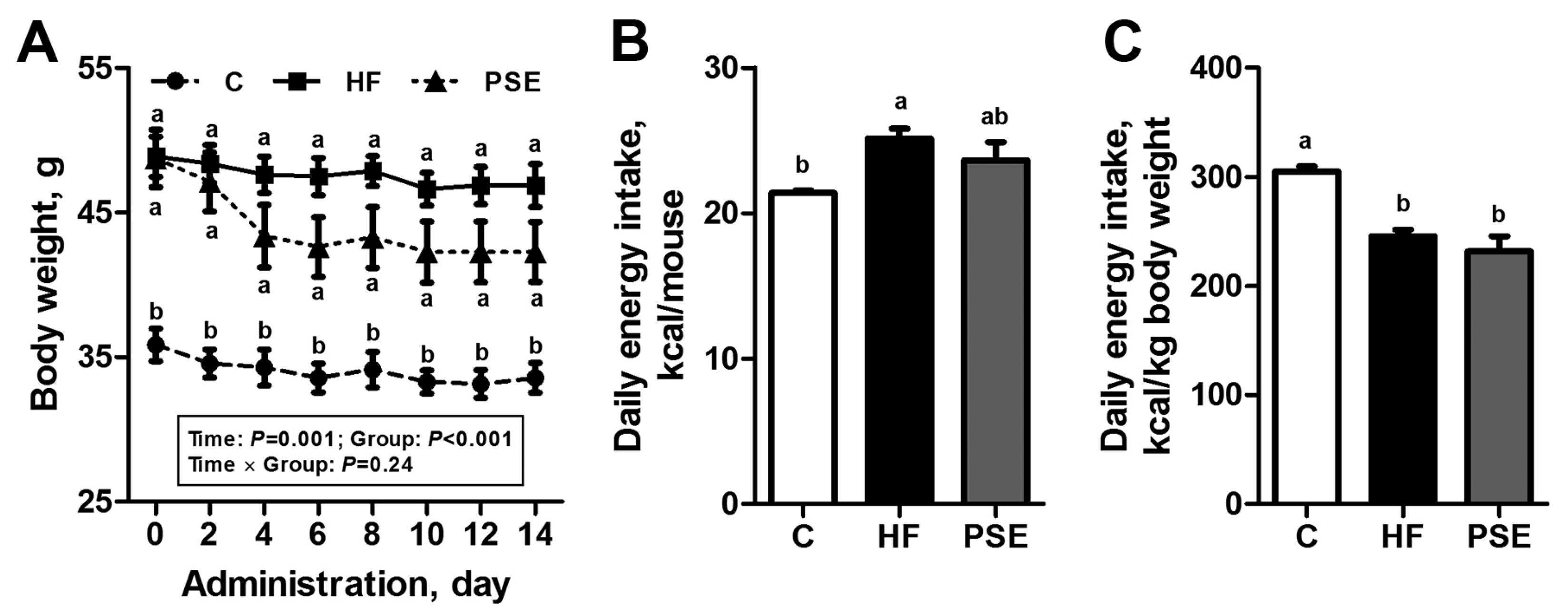
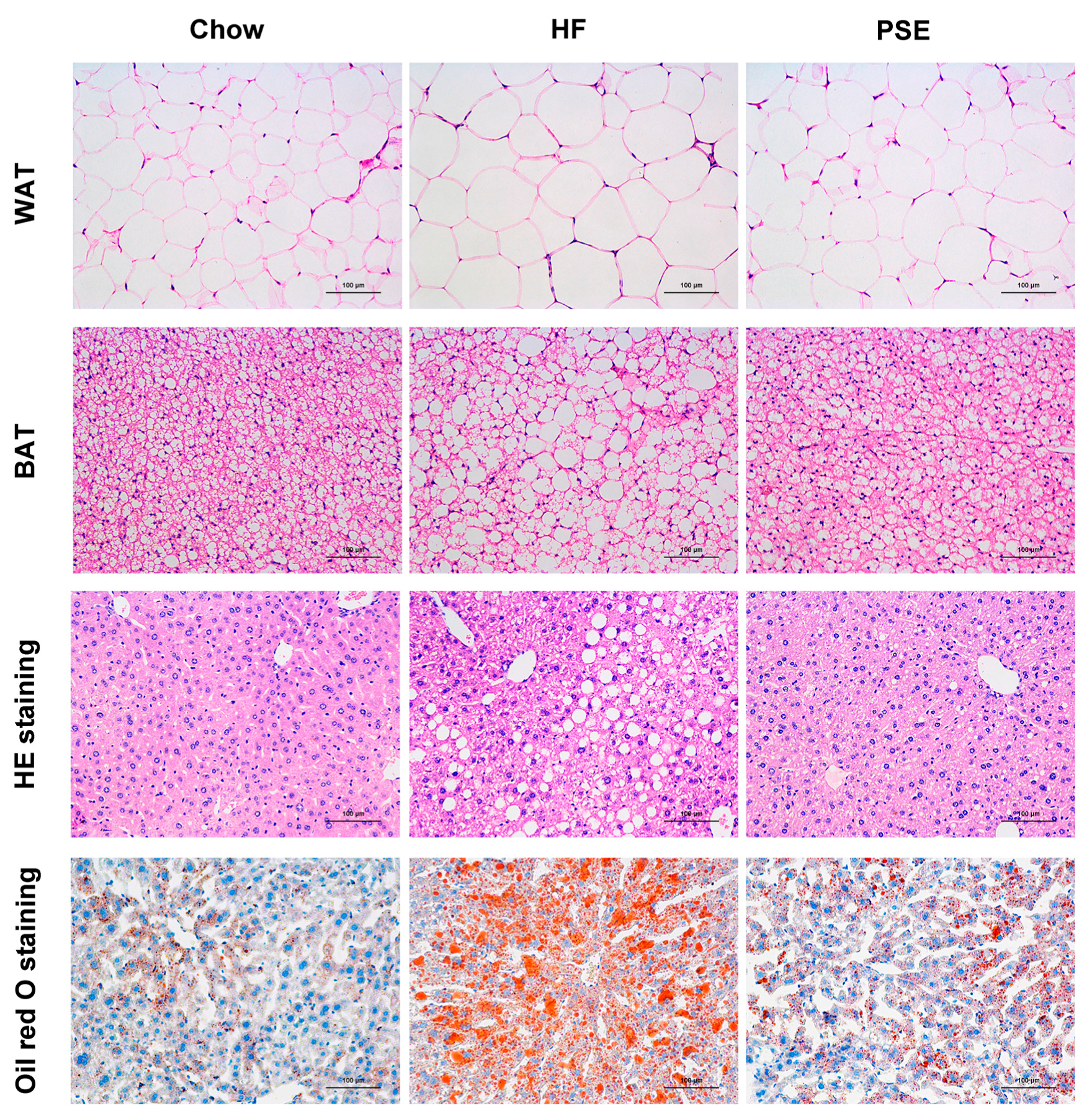
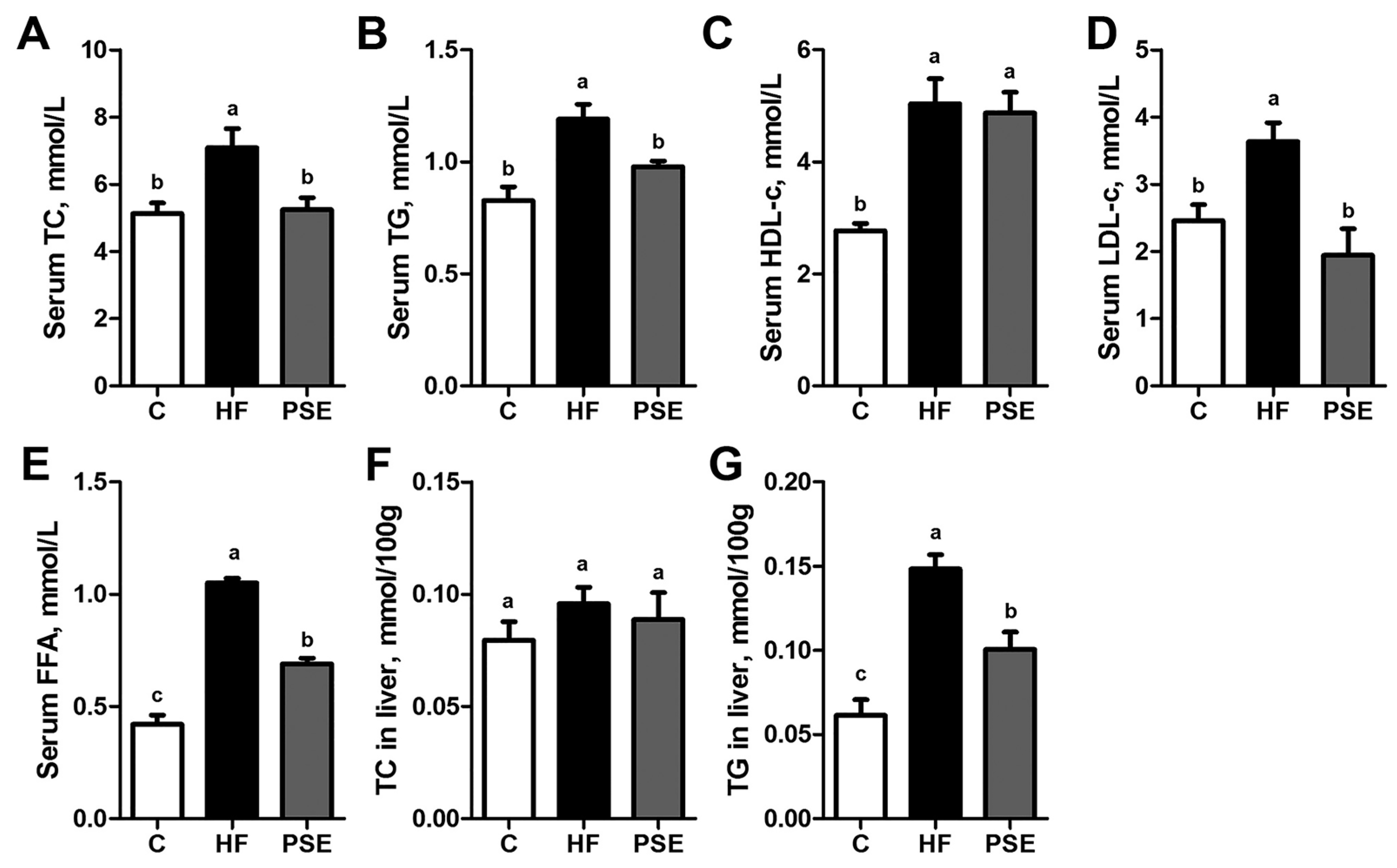
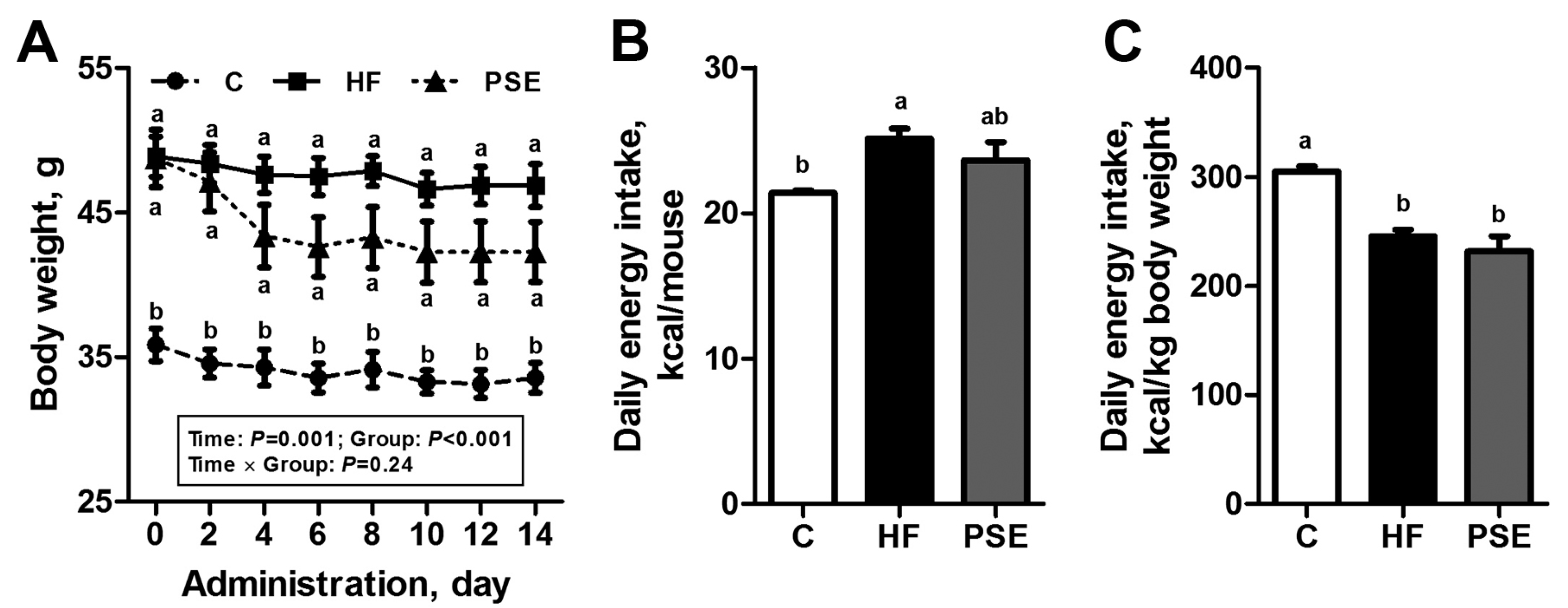
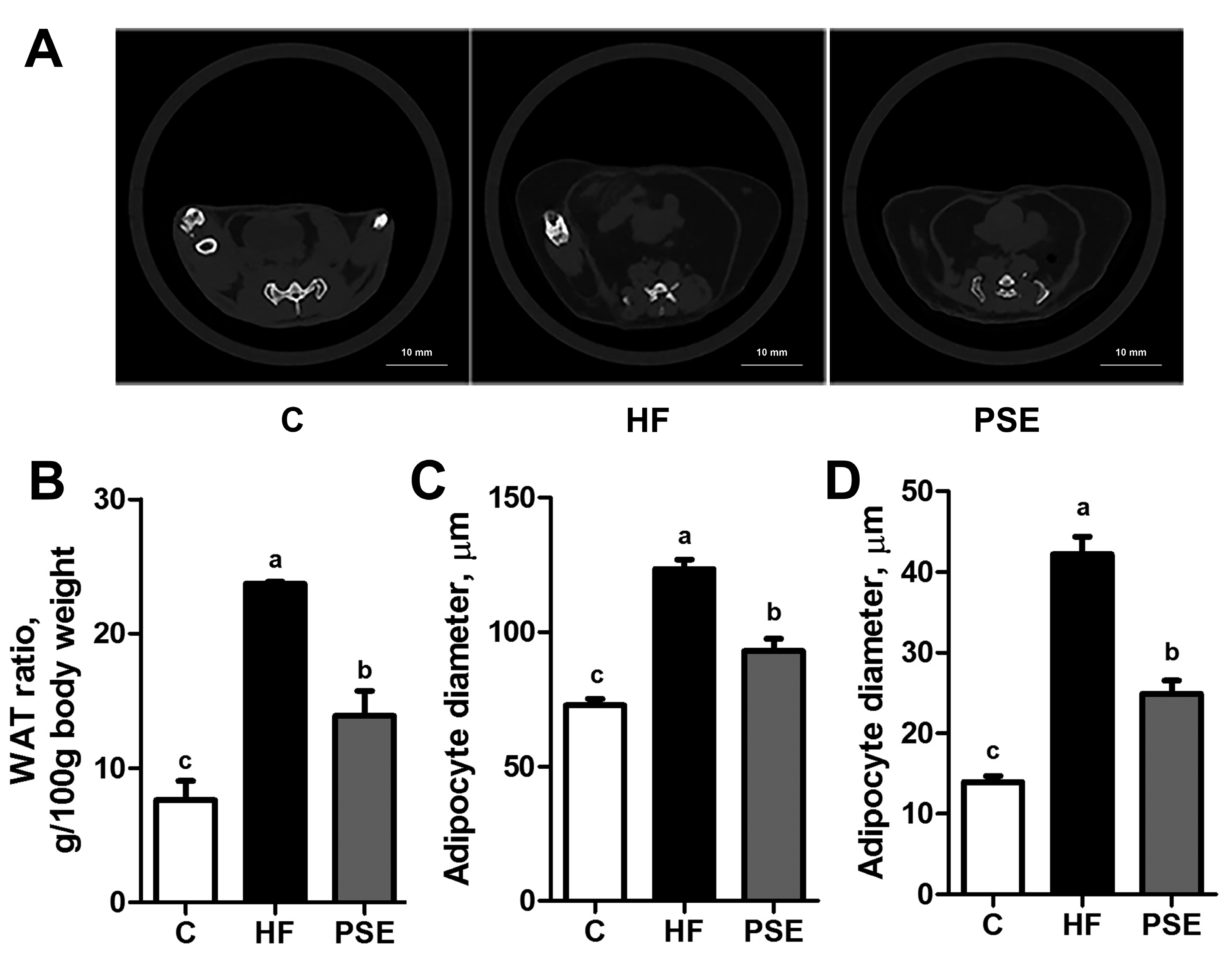
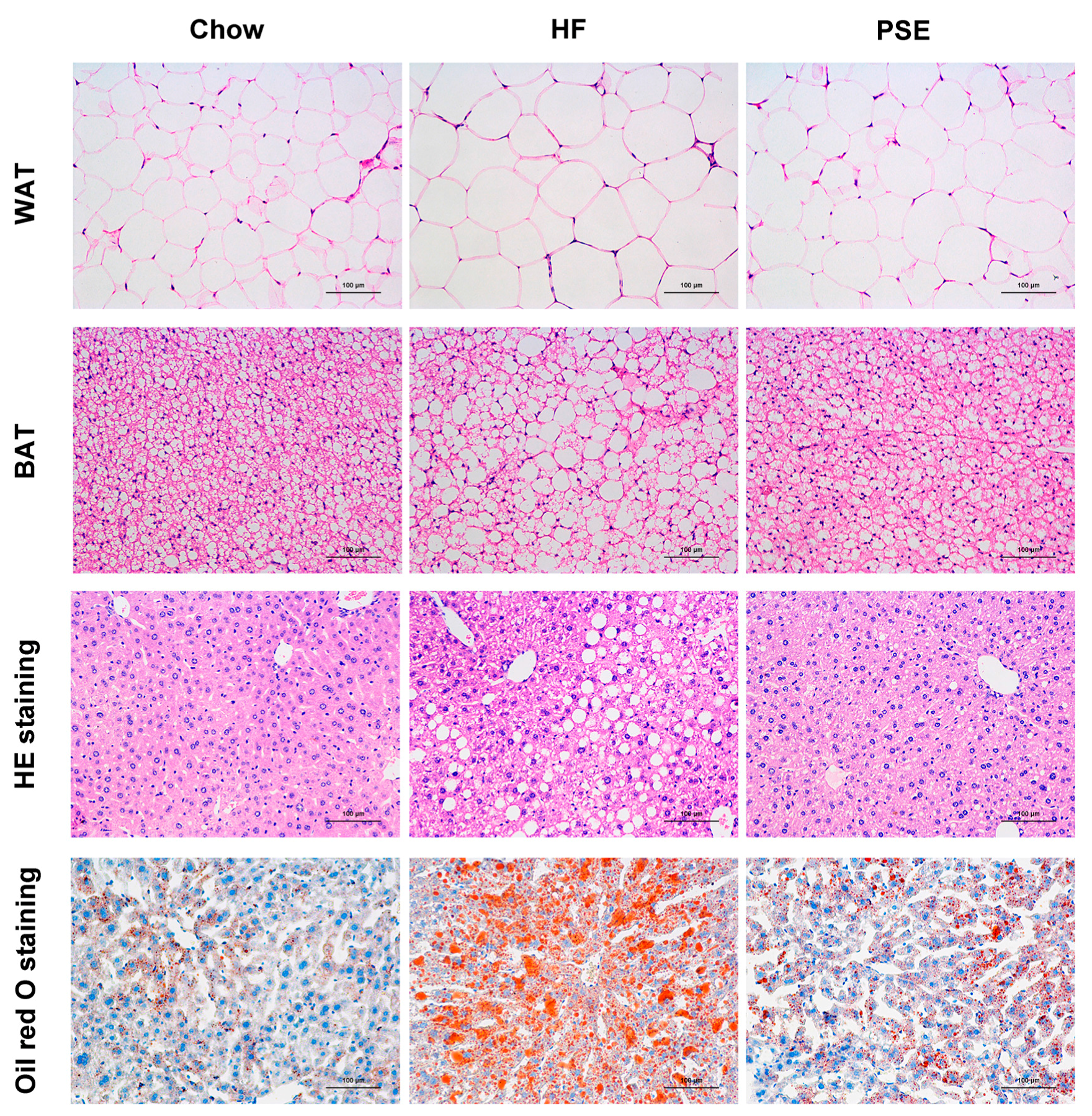
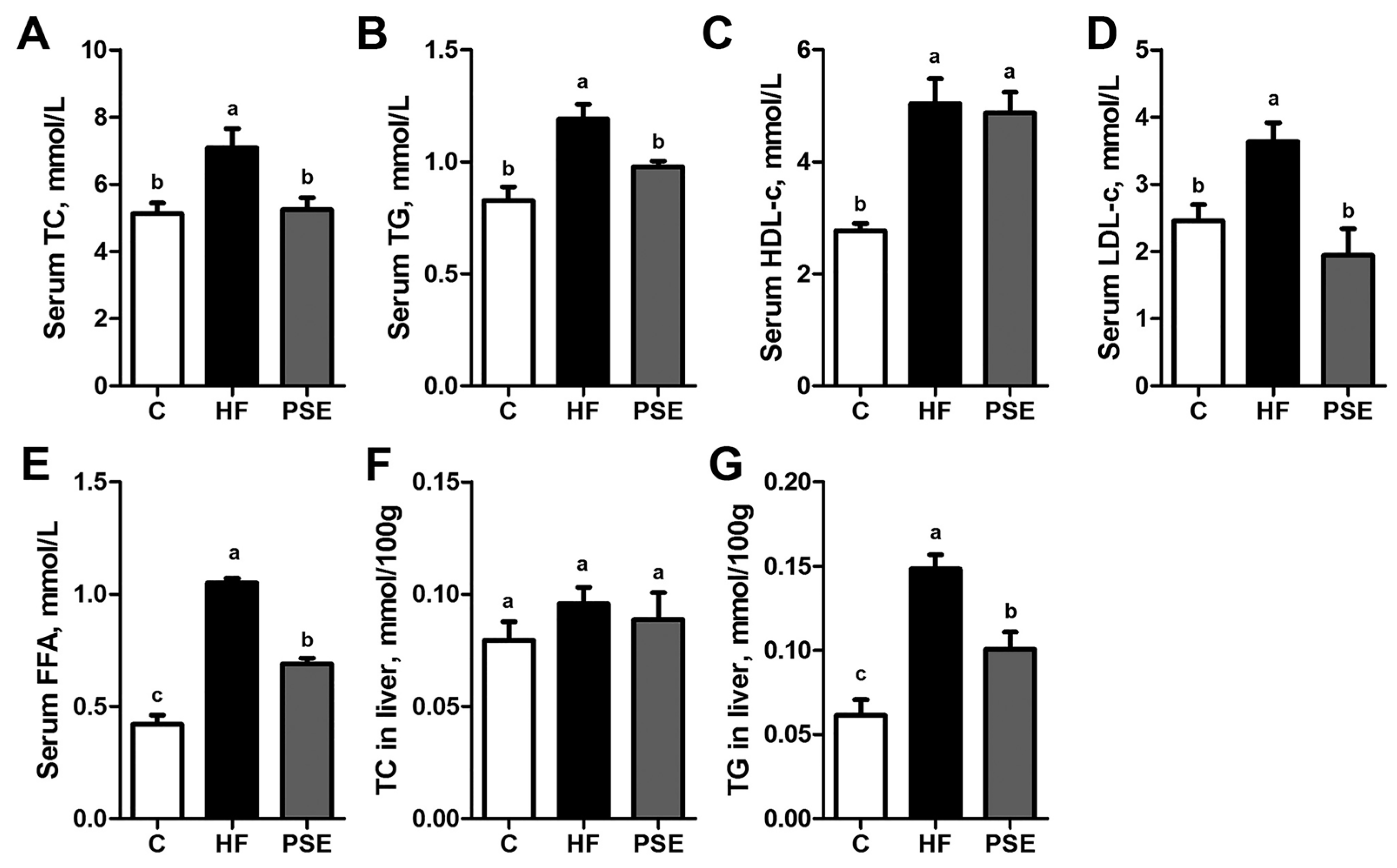
2.2. The Effects of PSE on the Body Weight, Food Intake and Lipid Accumulation of High-Fat (HF) Diet-Induced Obese C57BL/6 Mice
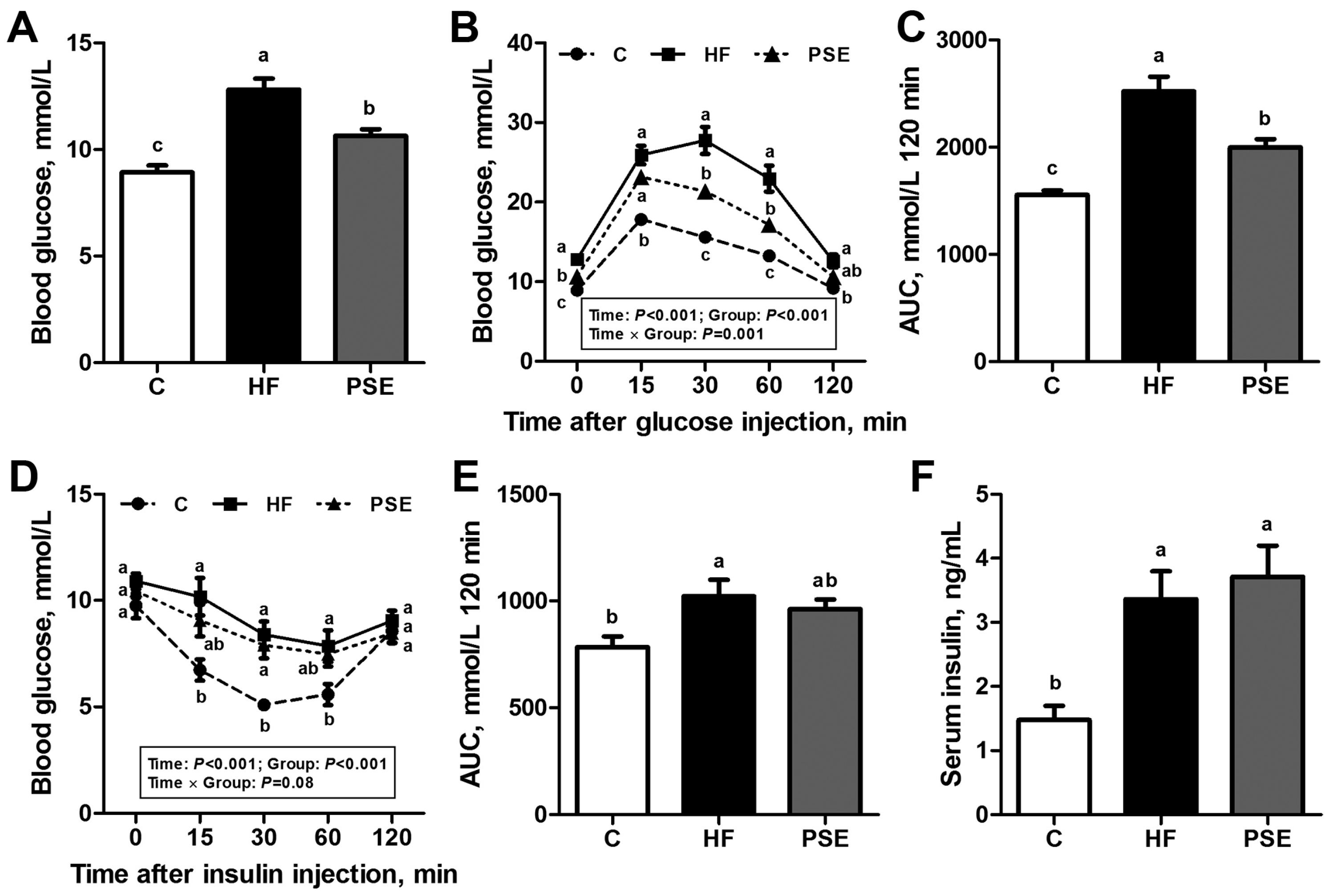
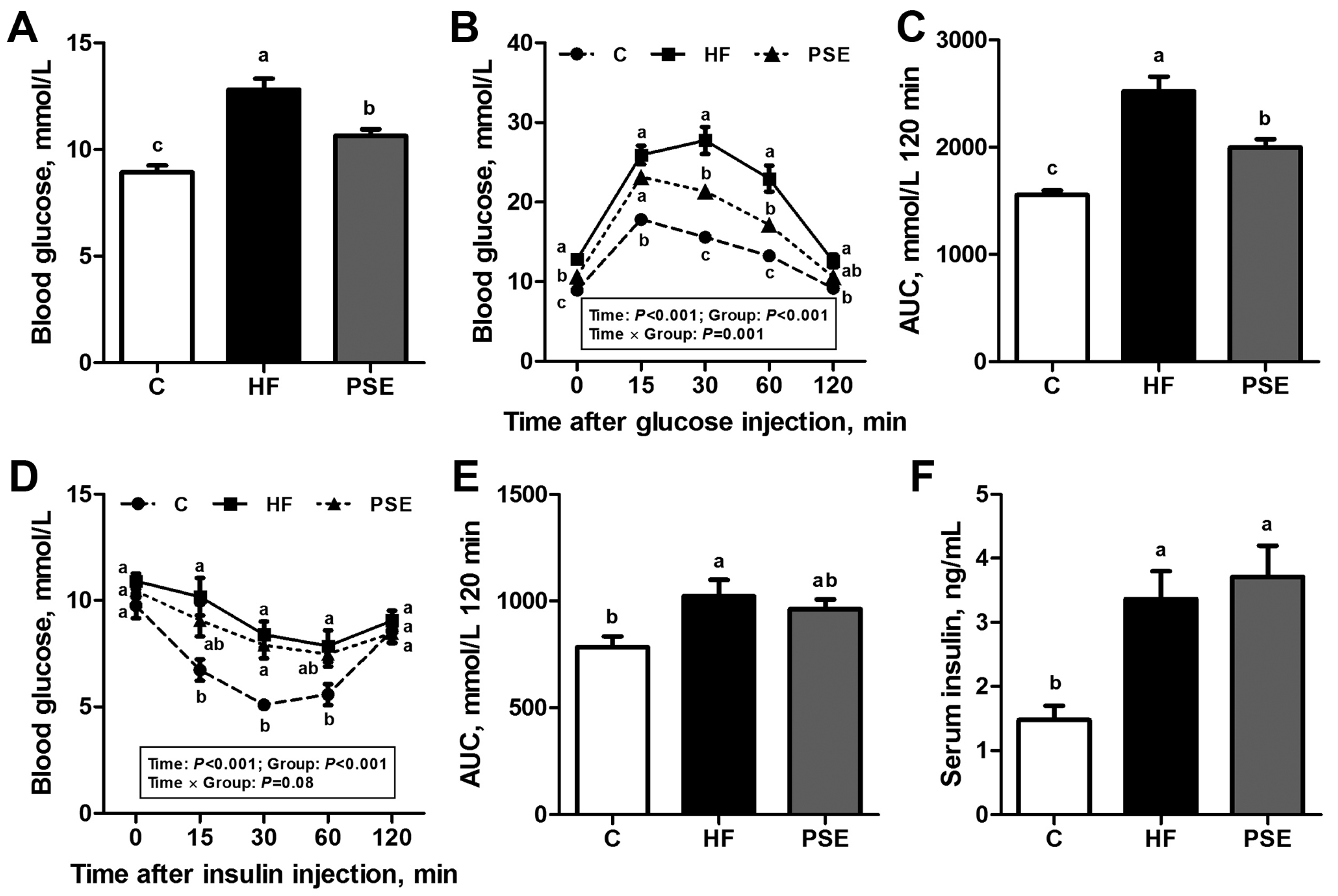
2.3. The Effects of PSE on Glucose Homeostasis of HF Diet-Induced Obese C57BL/6 Mice
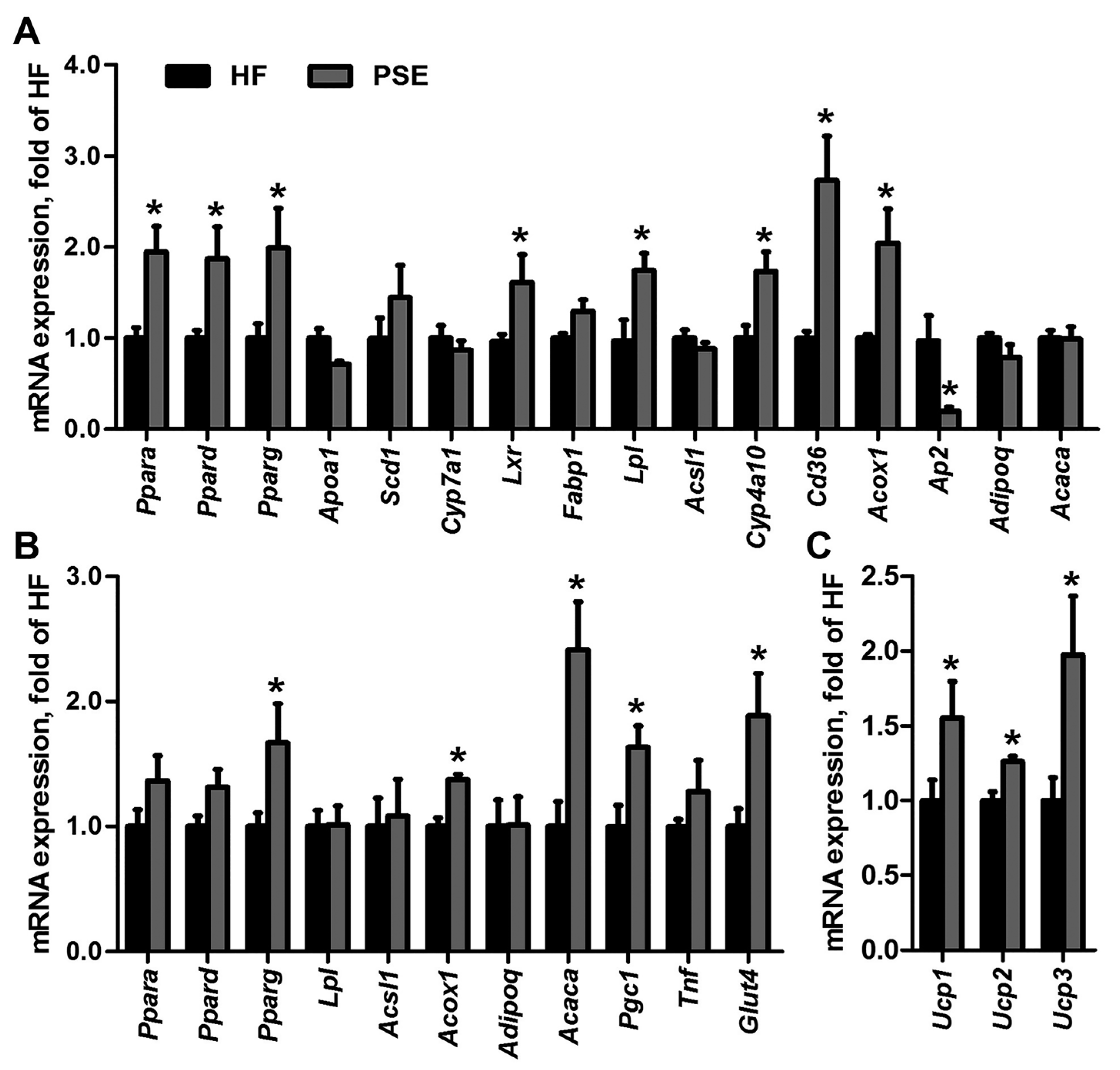
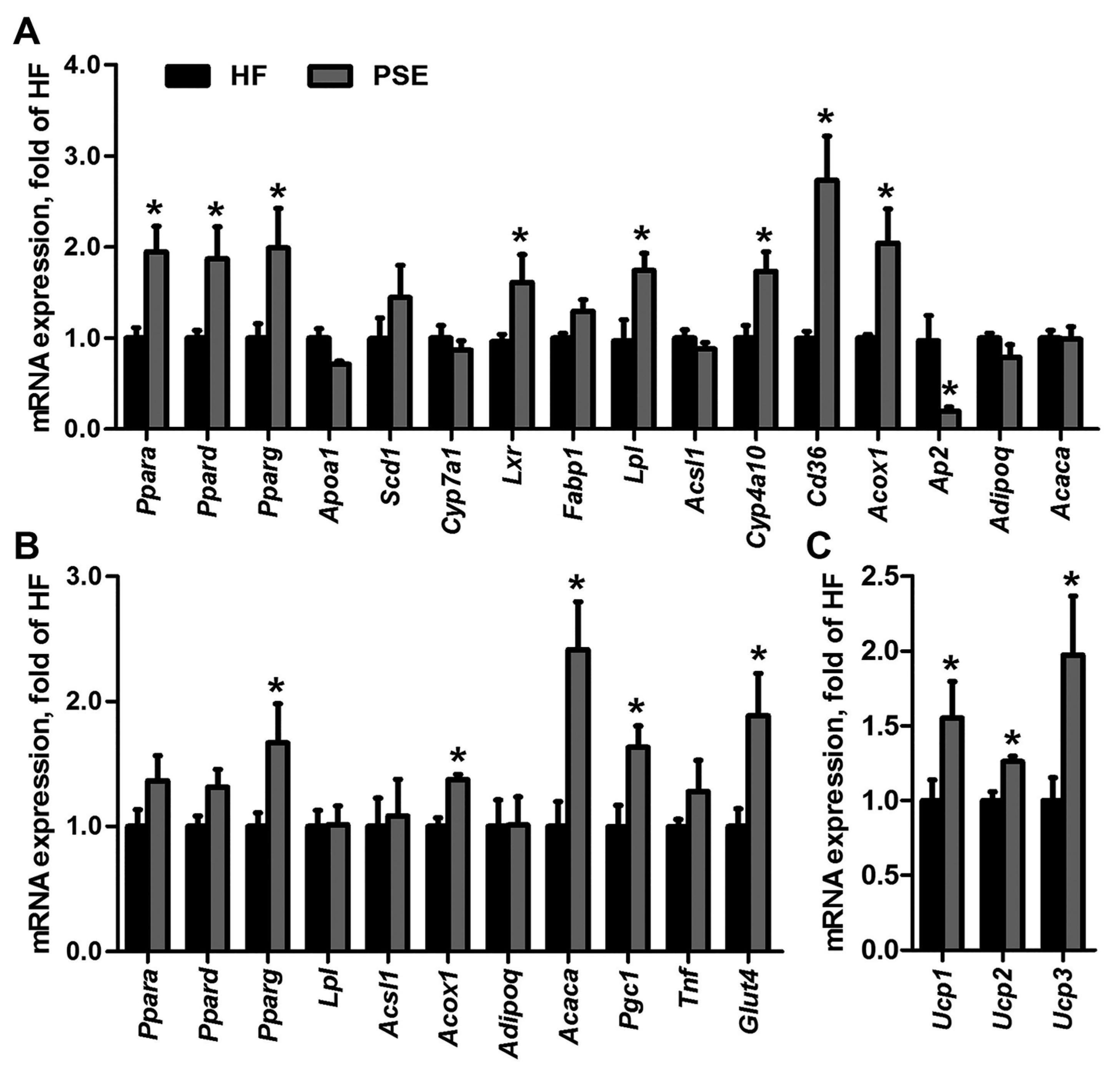
2.4. PSE Regulates Lipid and Glucose Metabolism Related Gene Expressions
3. Discussion
4. Materials and Methods
4.1. Extract Preparation and UPLC-MS Analysis
4.2. Experimental Animals and Diet
4.3. Computed Tomography Scan Analysis
4.4. Intraperitoneal Glucose and Insulin Tolerance Tests
4.5. Serum and Hepatic Lipid Concentrations
4.6. Histological Analysis
4.7. Microarray Hybridization
4.8. Transfection and Reporter Gene Assays
4.9. qRT-PCR Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Acaca | acetyl-Coenzyme A carboxylase α |
| Acox1 | acyl-Coenzyme A oxidase 1 |
| Acsl1 | acyl-CoA synthetase long-chain family member 1 |
| Adipoq | adiponectin |
| Ap2 | fatty acid binding protein 4 |
| Apoa1 | apolipoprotein A-I |
| AUC | area under the curve |
| BAT | brown adipose tissue |
| C | control |
| Cd36 | cluster of differentiation 36 |
| ChREBP | carbohydrate-responsive element-binding protein |
| Cyp | cytochrome P450 |
| Fabp1 | fatty acid binding protein 1 |
| FFA | free fatty acid |
| Glut4 | glucose transporter type 4 |
| GTT | glucose tolerance test |
| HDL-c | high density lipoprotein cholesterol |
| H&E | hematoxylin and eosin |
| HF | high-fat |
| HOMA-IS | HOMA-insulin sensitivity |
| HOMA-β | HOMA-beta cell function |
| ITT | insulin tolerance test |
| LDL-c | low density lipoprotein cholesterol |
| Lpl | lipoprotein lipase |
| Lxr | liver X receptor α |
| MAPK | mitogen-activated protein kinase |
| Pgc1 | PPAR gamma coactivator 1 α; |
| PPAR | peroxisome proliferator activated receptor |
| PSE | Plantago asiatica L. seed extract |
| Scd1 | stearoyl-Coenzyme A desaturase 1 |
| SREBP | sterol regulatory element-binding protein |
| TC | total cholesterol |
| TG | triglyceride |
| Tnf | tumor necrosis factor α |
| Ucp | uncoupling protein |
| UPLC-MS | ultra-performance liquid chromatography-mass spectrometry |
| WAT | white adipose tissue |
References
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox, and impact of weight loss. J. Am. Coll Cardiol. 2009, 53, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Battelino, T.; Shalitin, S. Obesity, Metabolic Syndrome and Nutrition. World Rev. Nutr. Diet 2014, 109, 1–22. [Google Scholar] [PubMed]
- Kennett, G.A.; Clifton, P.G. New approaches to the pharmacological treatment of obesity: Can they break through the efficacy barrier? Pharmacol. Biochem. Behav. 2010, 97, 63–83. [Google Scholar] [CrossRef] [PubMed]
- Thounaojam, M.C.; Nammi, S.; Jadeja, R. Natural products for the treatment of obesity, metabolic syndrome, and type 2 diabetes. Evid. Based Complement. Altern. Med. 2013, 2013, 871018. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kauter, K.; Brown, L. Naringin improves diet-induced cardiovascular dysfunction and obesity in high carbohydrate, high fat diet-fed rats. Nutrients 2013, 5, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Vasudeva, N.; Yadav, N.; Sharma, S.K. Natural products: A safest approach for obesity. Chin. J. Integr. Med. 2012, 18, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Liu, L.T.; Chiang, L.C.; Lin, C.C. In vitro anti-hepatoma activity of fifteen natural medicines from Canada. Phytother. Res. 2002, 16, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.O.; Hong, S.T.; Lee, Y.H.; Lee, K.W. Protective effects of Plantago asiatica L. extract against ferric-nitrilotriacetate (Fe-NTA) induced liver oxidative stress in Wistar rats. FASEB J. 2012, 26, 692–716. [Google Scholar]
- Zhou, Q.; Lu, W.; Niu, Y.; Liu, J.; Zhang, X.; Gao, B.; Akoh, C.C.; Shi, H.; Yu, L.L. Identification and quantification of phytochemical composition and anti-inflammatory, cellular antioxidant, and radical scavenging activities of 12 Plantago species. J. Agric. Food Chem. 2013, 61, 6693–6702. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2010; Volume 1, pp. 63–74. [Google Scholar]
- Geng, F.; Sun, Q.; Yang, L.; Wang, Z.T. Effects of the seeds and herbs of Plantago asiatica L. on diuresis. SH J. TCM 2009, 21, 150–156. [Google Scholar]
- Galisteo, M.; Sánchez, M.; Vera, R.; González Anguera, A.; Duarte, J.; Zarzuelo, A. A diet supplemented with husks of Plantago ovata reduces the development of endothelial dysfunction, hypertension, and obesity by affecting adiponectin and TNF-α in obese Zucker rats. J. Nutr. 2005, 135, 2399–2404. [Google Scholar] [PubMed]
- Hannan, J.M.; Ali, L.; Khaleque, J.; Akhter, M.; Flatt, P.R.; Abdel-Wahab, Y.H. Aqueous extracts of husks of Plantago ovata reduce hyperglycaemia in type 1 and type 2 diabetes by inhibition of intestinal glucose absorption. Br. J. Nutr. 2006, 96, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.; Lee, K.W.; Kim, J.Y.; Kwon, O. A beverage of Asiatic plantain extracts alleviated postprandial oxidative stress in overweight hyperlipidemic subjects challenged with a high-fat meal: A preliminary study. Nutr. Res. 2013, 33, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Jung, S.H.; Lee, H.S.; Park, K.W.; Yun, B.S.; Lee, K.W. Glycation inhibitory activity and the identification of an active compound in P. asiatica extract. Phytother. Res. 2008, 22, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Nie, S.P.; Li, C.; Xie, M.Y. In vitro effects of a novel polysaccharide from the seeds of Plantago asiatica L. on intestinal function. Int. J. Biol. Macromol. 2013, 54, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Nie, S.P.; Wu, Q.M.; Li, C.; Fu, Z.H.; Gong, J.; Cui, S.W.; Xie, M.Y. Polysaccharide from seeds of Plantago asiatica L. affects lipid metabolism and colon microbiota of mouse. J. Agric. Food Chem. 2014, 62, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Park, K.W.; Kim, K.H.; Kim, C.T.; Baek, J.P.; Bang, K.H.; Choi, Y.M.; Lee, S.J. Asian plantain (Plantago asiatica) essential oils suppress 3-hydroxy-3-methyl-glutaryl-co-enzyme A reductase expression in vitro and in vivo and show hypocholesterolaemic properties in mice. Br. J. Nutr. 2008, 99, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Lambert, J.D.; Ju, J.; Reuhl, K.R.; Shapses, S.A.; Yang, C.S. The major green tea polyphenol, (−)-epigallocatechin-3-gallate, inhibits obesity, metabolic syndrome, and fatty liver disease in high-fat-fed mice. J. Nutr. 2008, 138, 1677–1683. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, D.; Rao, S.S.C. Efficacy and safety of traditional medical therapies for chronic constipation: Systematic review. Am. J. Gastroenterol. 2005, 100, 936–971. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Yang, L.; Chou, G.X.; Wang, Z.T. Bioguided isolation of angiotensin-converting enzyme inhibitors from the seeds of Plantago asiatica L. Phytother. Res. 2010, 24, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.F.; Xie, M.Y.; Yin, J.Y.; Nie, S.P.; Tang, Y.F.; Xie, X.M.; Zhou, C. Immunomodulatory activity of the seeds of Plantago asiatica L. J. Ethnopharmacol. 2009, 124, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.L.; Hu, W.L.; Dai, D.H. Extraction of polysaccharides and the antioxidant activity from the seeds of Plantago asiatica L. Int. J. Biol. Macromol. 2011, 49, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.F.; Nie, S.P.; Jiang, L.M.; Xie, M.Y. A novel polysaccharide from the seeds of Plantago asiatica L. induces dendritic cells maturation through toll-like receptor 4. Int. Immunopharmacol. 2014, 18, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Xiong, A.; Geng, F.; Yang, L.; Wang, Z. A novel strategy for target profiling analysis of bioactive phenylethanoid glycosides in Plantago medicinal plants using ultra-performance liquid chromatography coupled with tandem quadrupole mass spectrometry. J. Sep. Sci. 2012, 12, 1470–1478. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qi, M.; Yang, Q.; Tong, R.; Wang, R.; Bligh, S.W.; Yang, L.; Wang, Z. Comprehensive metabolite profiling of Plantaginis Semen using ultra high performance liquid chromatography with electrospray ionization quadrupole time-of-flight tandem mass spectrometry coupled with elevated energy technique. J. Sep. Sci. 2016, 39, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, K.M.; Park, J.; Kwak, J.H.; Kim, Y.S.; Lee, S.M. Geniposidic acid protects against d-galactosamine and lipopolysaccharide-induced hepatic failure in mice. J. Ethnopharmacol. 2013, 146, 271–277. [Google Scholar]
- Luo, L.F.; Wu, W.H.; Zhou, Y.J.; Yan, J.; Yang, G.P.; Ouyang, D.S. Antihypertensive effect of Eucommia ulmoides Oliv. extracts in spontaneously hypertensive rats. J. Ethnopharmacol. 2010, 2, 238–243. [Google Scholar]
- Yamabe, N.; Noh, J.S.; Park, C.H.; Kang, K.S.; Shibahara, N.; Tanaka, T.; Yokozawa, T. Evaluation of loganin, iridoid glycoside from Corni fructus, on hepatic and renal glucolipotoxicity and inflammation in type 2 diabetic db/db mice. Eur. J. Pharmacol. 2010, 648, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Hirata, T.; Hosoo, S.; Nakajima, K.; Wada, A.; Yurugi, Y.; Soya, H.; Matsui, T.; Yamaguchi, A.; Ogata, M.; et al. Asperuloside stimulates metabolic function in rats across several organs under high-fat diet conditions, acting like the major ingredient of Eucommia leaves with anti-obesity activity. J. Nutr. Sci. 2012, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Qi, J.; Yu, B.Y. Iridoid and phenylpropanoid glycosides from Scrophularia ningpoensis Hemsl. and their α-glucosidase inhibitory activities. Fitoterapia 2014, 93, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; He, K.; Ibarra, A.; Bily, A.; Roller, M.; Chen, X.; Rühl, R. Iridoids from Fraxinus excelsior with adipocyte differentiation-inhibitory and PPARα activation activity. J. Nat. Prod. 2010, 73, 2–6. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Q.; Ma, G.Y.; Peng, J.N.; Ma, Z.Y.; Hamann, M.T. Liver X receptor and peroxisome proliferator-activated receptor agonist from Cornus alternifolia. Biochim. Biophys. Acta 2012, 1820, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Woo, E.R.; Choi, C.Y.; Shin, D.W.; Lee, D.G.; You, H.J.; Jeong, H.G. Protective effect of acteoside on carbon tetrachloride-induced hepatotoxicity. Life Sci. 2004, 74, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Hu, H.J.; Li, P.F.; Yang, Y.B.; Wu, L.H.; Chou, G.X.; Wang, Z.T. Diterpenoids and phenylethanoid glycosides from the roots of Clerodendrum bungei and their inhibitory effects against angiotensin converting enzyme and α-glucosidase. Phytochemistry 2014, 103, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.M.; Liu, F.; He, Z.D.; Ji, M.; Chu, X.X.; Kang, Z.Y.; Cai, D.Y.; Gao, N.N. Anti-obesity effect of total phenylpropanoid glycosides from Ligustrum robustum Blume in fatty diet-fed mice via up-regulating leptin. J. Ethnopharmacol. 2015, 169, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Rikimaru, K.; Sakai, M.; Nishibe, S.; Fujikawa, T.; Tamura, Y. Plantago lanceolata L. leaves prevent obesity in C57BL/6 J. mice fed a high-fat diet. Nat. Prod. Res. 2013, 27, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Goda, Y.; Kawahara, N.; Kiuchi, F.; Hirakura, K.; Kikuchi, Y.; Nishimura, H.; Takao, M.; Marumoto, M.; Kitazaki, H.A. Guanidine derivative from seeds of Plantago asiatica. J. Nat. Med. 2009, 63, 58–60. [Google Scholar]
- Wang, Z.T.; Geng, F.; Sun, Q.; Yang, L.; Qi, M.; Liao, L.P.; Ji, G.; Dow, W.; Zheng, X.M. Use of Guanidine Derivatives and Preparation, Drug Combination and Preparation of Medicine for Treating Metabolic Syndrome. Chinese Patent No. CN 200910201771.6, 9 November 2009. [Google Scholar]
- Tabata, A.; Taniguchi, M.; Shibano, M.; Ecliptamines, A.D. four new guanidine alkaloids from Eclipta prostrata L. Phytochem. Lett. 2015, 11, 224–228. [Google Scholar] [CrossRef]
- Zheng, X.M.; Meng, F.W.; Geng, F.; Qi, M.; Luo, C.; Yang, L.; Wang, Z.T. Plantadeprate A, a tricyclic monoterpene zwitterionic guanidium, and related derivatives from the seeds of Plantago depressa. J. Nat. Prod. 2015, 78, 2822–2826. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.J.; Guo, L.; Zhang, Y.; Sun, QH.; Yang, B.C.; Huang, C. Okra polysaccharide improves metabolic disorders in high-fat diet-induced obese C57BL/6 mice. Mol. Nutr. Food Res. 2013, 57, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Li, M.X.; Ding, X.B.; Fan, S.J.; Guo, L.; Gu, M.; Zhang, Y.; Feng, L.; Jiang, D.; Li, Y.M.; et al. Effects of Fortunella margarita fruit extract on metabolic disorders in high-fat diet-induced obese C57BL/6 mice. PLoS ONE 2014, 9, e93510. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X. PPARs: Diverse regulators in energy metabolism and metabolic diseases. Cell Res. 2010, 20, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Kim, J.Y.; Jun, H.J.; Kim, S.J.; Lee, J.H.; Hoang, M.H.; Hwang, K.Y.; Um, S.J.; Chang, H.I.; Lee, S.J. The natural carotenoid astaxanthin, a PPAR-α agonist and PPAR-γ antagonist, reduces hepatic lipid accumulation by rewiring the transcriptome in lipid-loaded hepatocytes. Mol. Nutr. Food Res. 2012, 6, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Burn, P. Lipid metabolic enzymes: Emerging drug targets for the treatment of obesity. Nat. Rev. Drug Discov. 2004, 8, 695–710. [Google Scholar]
- Furuhashi, M.; Hotamisligil, G.S. Fatty acid-binding proteins: Role in metabolic diseases and potential as drug targets. Nat. Rev. Drug Discov. 2008, 6, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Qian, Z.; Shi, Y.; Yang, L.; Xi, L.; Zhao, B.; Xu, X.; Ji, H. Crocetin improves the insulin resistance induced by high-fat diet in rats. Br. J. Pharmacol. 2008, 5, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.R.; Shin, D.H.; Kwon, D.Y.; Cha, Y.S. Effect of Cheonggukjang supplementation upon hepatic acyl-CoA synthase, carnitine palmitoyltransferase I.; acyl-CoA oxidase and uncoupling protein 2 mRNA levels in C57BL/6J. mice fed with high fat diet. Genes Nutr. 2008, 4, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Handschin, C.; Spiegelman, BM. Peroxisome proliferator-activated receptor gamma coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr. Rev. 2006, 7, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Armoni, M.; Harel, C.; Karnieli, E. Transcriptional regulation of the GLUT4 gene: From PPAR-gamma and FOXO1 to FFA and inflammation. Trends Endocrinol. Metab. 2007, 3, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Li, J.; Song, B.; Xiao, X.; Huang, W.; Zhang, B.; Tang, X.; Qi, M.; Yang, Q.; Yang, Q.; et al. Andrographolide prevents high-fat diet-induced obesity in C57BL/6 mice by suppressing the sterol regulatory element-binding protein pathway. J. Pharmacol. Exp. Ther. 2014, 351, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Zhang, Y.; Fan, S.J.; Ding, X.B.; Ji, G.; Huang, C. Extracts of Rhizoma Polygonati Odorati prevent high-fat diet-induced metabolic disorders in C57BL/6 Mice. PLoS ONE 2013, 8, e81724. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Guo, L.; Zhang, Y.; Fan, S.; Gu, M.; Lu, Y.; Jiang, D.; Li, Y.; Huang, C.; Zhou, Z. Extracts of pomelo peels prevent high-fat diet-induced metabolic disorders in c57bl/6 mice through activating the PPARα and GLUT4 pathway. PLoS ONE 2013, 8, e77915. [Google Scholar] [CrossRef] [PubMed]
- Zang, M.; Xu, S.; Maitland-Toolan, K.A.; Zuccollo, A.; Hou, X.; Jiang, B.; Wierzbicki, M.; Verbeuren, T.J.; Cohen, R.A. Polyphenols stimulate AMP-activated protein kinase, lower lipids, and inhibit accelerated atherosclerosis in diabetic LDL receptor–deficient mice. Diabetes 2006, 55, 2180–2191. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qu, X.; Chen, Y.; Liao, L.; Cheng, K.; Shao, C.; Zenke, M.; Keating, A.; Zhao, R.C. Mesenchymal stem/stromal cells induce the generation of novel IL-10-dependent regulatory dendritic cells by SOCS3 activation. J. Immunol. 2012, 189, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Zhang, Y.; Fan, S.; Gu, M.; Guan, Y.; Lu, X.; Huang, C.; Zhou, Z. Preventive and ameliorating effects of citrus d-limonene on dyslipidemia and hyperglycemia in mice with high-fat diet-induced obesity. Eur. J. Pharmacol. 2013, 715, 46–55. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Qi, M.; Tong, R.; Wang, D.; Ding, L.; Li, Z.; Huang, C.; Wang, Z.; Yang, L. Plantago asiatica L. Seed Extract Improves Lipid Accumulation and Hyperglycemia in High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2017, 18, 1393. https://doi.org/10.3390/ijms18071393
Yang Q, Qi M, Tong R, Wang D, Ding L, Li Z, Huang C, Wang Z, Yang L. Plantago asiatica L. Seed Extract Improves Lipid Accumulation and Hyperglycemia in High-Fat Diet-Induced Obese Mice. International Journal of Molecular Sciences. 2017; 18(7):1393. https://doi.org/10.3390/ijms18071393
Chicago/Turabian StyleYang, Qiming, Meng Qi, Renchao Tong, Dandan Wang, Lili Ding, Zeyun Li, Cheng Huang, Zhengtao Wang, and Li Yang. 2017. "Plantago asiatica L. Seed Extract Improves Lipid Accumulation and Hyperglycemia in High-Fat Diet-Induced Obese Mice" International Journal of Molecular Sciences 18, no. 7: 1393. https://doi.org/10.3390/ijms18071393