Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions

 ,
,  and
and
Abstract
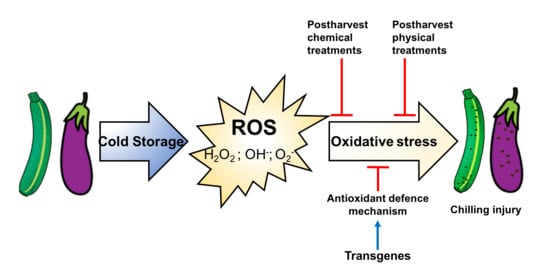
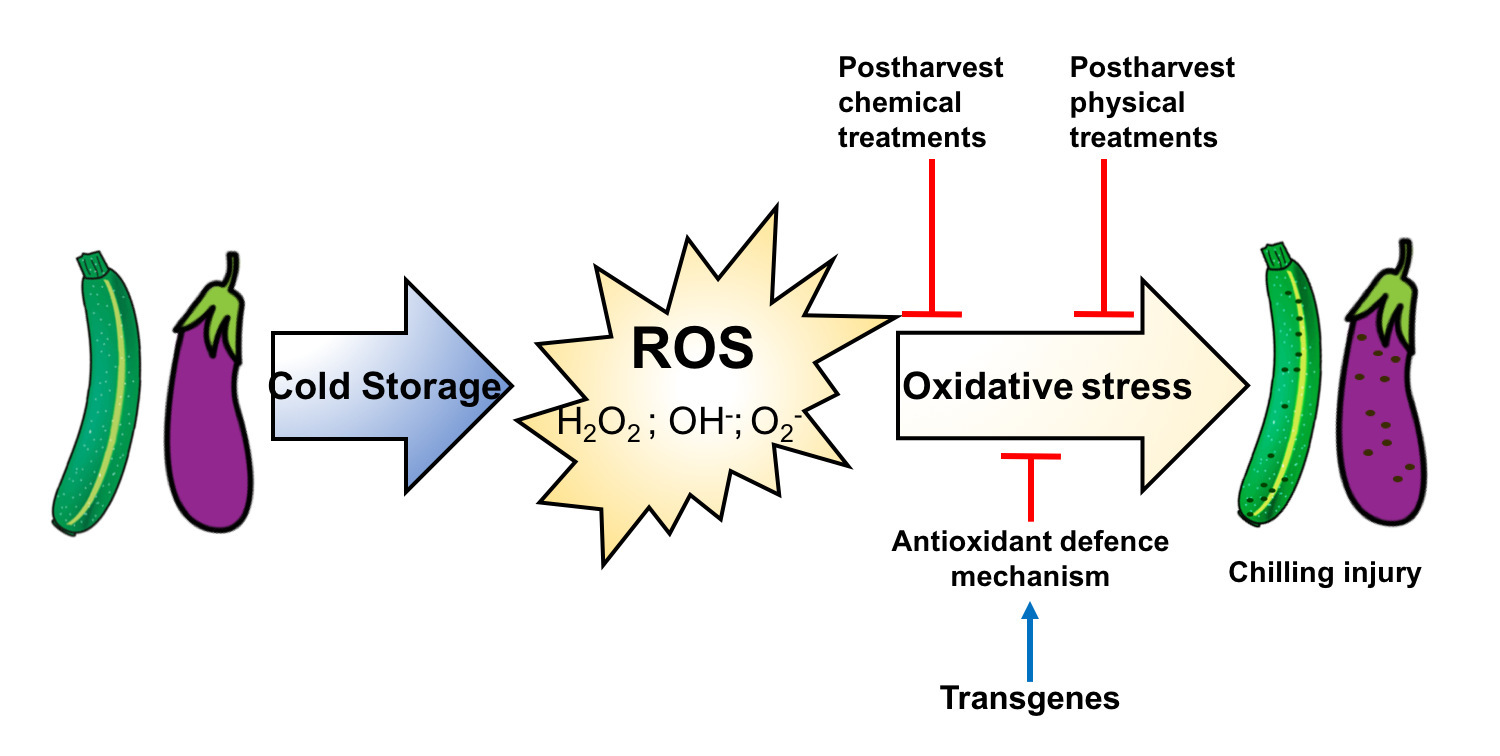
:
1. Postharvest Technology and Physiology of Immature Fruits
1.1. Immature Fruits
1.2. Harvest Time/Optimal Developmental Stage
1.3. Postharvest Physiology
2. Chilling Injury and Oxidative Stress
2.1. Chilling Injury (CI)
2.2. Oxidative Stress
2.2.1. Overview
2.2.2. Low Temperature and Oxidative Stress in Vegetable Fruit
3. Postharvest Treatments that Reduce Oxidative Stress and Chilling Injury
3.1. Physical Approaches
3.1.1. Temperature Treatments
3.1.2. The Use of Controlled and Modified Atmospheres
3.1.3. The Effects of Different Types of Wrapping
3.1.4. The Effects of Edible Coatings
3.2. Chemical Approaches
3.2.1. Ethylene (ET)
3.2.2. Brassinosteroids (BRs)
3.2.3. Abscisic Acid (ABA)
3.2.4. Salicylic Acid (SA) and Jasmonic Acid (JA)
3.2.5. Polyamines (PAs)
3.2.6. Nitric Oxide (NO)
3.2.7. Other Chemical Treatments
3.3. Breeding and Biotechnological Approaches
4. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Singh, D.P.; Sarma, B.K.; Upadhyay, G.; Singh, H.B. Polyphenolics from various extracts/fractions of red onion (Allium cepa) peel with potent antioxidant and antimutagenic activities. Food Chem. Toxicol. 2009, 47, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Díaz, M.T.; Font, R.; Martínez-Valdivieso, D.; del Río-Celestino, M. Diversity of natural pigments and phytochemical compounds from exocarp and mesocarp of 27 Cucurbita pepo accessions. Sci. Hortic. 2015, 197, 357–365. [Google Scholar] [CrossRef]
- Zaro, M.J.; Chaves, A.R.; Vicente, A.R.; Concellón, A. Distribution, stability and fate of phenolic compounds in white and purple eggplants (Solanum melongena L.). Postharvest Biol. Technol. 2014, 92, 70–78. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Nema, N.K.; Maity, N.; Sarkar, B.K. Phytochemical and therapeutic potential of cucumber. Fitoterapia 2013, 84, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Nema, N.K.; Maity, N.; Sarkar, B.; Mukherjee, P.K. Cucumis sativus fruit-potential antioxidant, anti-hyaluronidase, and anti-elastase agent. Arch. Dermatol. Res. 2011, 303, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Aburjai, T.; Natsheh, F.M. Plants used in cosmetics. Phyther. Res. 2003, 17, 987–1000. [Google Scholar] [CrossRef] [PubMed]
- Konopacka, D.; Seroczyńska, A.; Korzeniewska, A.; Jesionkowska, K.; Niemirowicz-Szczytt, K.; Płocharski, W. Studies on the usefulness of Cucurbita maxima for the production of ready-to-eat dried vegetable snacks with a high carotenoid content. LWT Food Sci. Technol. 2010, 43, 302–309. [Google Scholar] [CrossRef]
- Blanco-Díaz, M.T.; del Río-Celestino, M.; Martínez-Valdivieso, D.; Font, R. Use of visible and near-infrared spectroscopy for predicting antioxidant compounds in summer squash (Cucurbita pepo ssp pepo). Food Chem. 2014, 164, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.; Yadav, S. Pharmacological actions and potential uses of Momordica charantia: A review. J. Ethnopharmacol. 2004, 93, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-J.; Ng, L.-T. Antioxidant and free radical scavenging activities of wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) in Taiwan. LWT Food Sci. Technol. 2008, 41, 323–330. [Google Scholar] [CrossRef]
- Tan, S.P.; Parks, S.E.; Stathopoulos, C.E.; Roach, P.D. Greenhouse-grown bitter melon: Production and quality characteristics. J. Sci. Food Agric. 2014, 94, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Jini, D. Antidiabetic effects of Momordica charantia (bitter melon) and its medicinal potency. Asian Pac. J. Trop. Dis. 2013, 3, 93–102. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Liaw, C-C.; Hsiao, P-C.; Huang, H.-C.; Lin, Z.-H.; Hsu, F.-L.; Kuo, Y.-H. Cucurbitane-type glycosides from the fruits of Momordica charantia and their hypoglycaemic and cytotoxic activities. J. Funct. Foods 2014, 6, 564–574. [Google Scholar] [CrossRef]
- Alam, M.A.; Uddin, R.; Subhan, N.; Rahman, M.M.; Jain, P.; Reza, H.M. Beneficial role of bitter melon supplementation in obesity and related complications in metabolic syndrome. J. Lipids 2015, 2015, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jamuna, S.; Karthika, K.; Paulsami, S. Phytochemical and pharmacological properties of certain medicinally important species of Cucurbitaceae family—A review. J. Res. Biol. 2015, 5, 1835–1849. [Google Scholar]
- Du, Q.; Xu, Y.; Li, L.; Zhao, Y.; Jerz, G.; Winterhalter, P. Antioxidant constituents in the fruits of Luffa cylindrica (L.) roem. J. Agric. Food Chem. 2006, 54, 4186–4190. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; You, K.-R.; Lee, S.-Y.; Song, C.-H.; Kim, D.-G. Oxidative stress-mediated apoptosis the anticancer effect of the sesquiterpene lactone parthenolide. J. Biol. Chem. 2002, 277, 38954–38964. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, P.K.; Kumar, S.; Jena, P.K. Nutritional and medicinal values of selected wild cucurbits available in Simipla biosphere reserve forest, Odisha. Int. J. Pharm. Sci. Res. 2014, 5, 5430–5437. [Google Scholar]
- Toivonen, P.M.A. Postharvest Physiology of Fruits and Vegetables. In Postharvest Ripening Physiology of Crops, 1st ed.; Pareek, S., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 49–79. [Google Scholar]
- Kader, A.A. Postharvest Technology of Horticultural Crops; University of California, Agriculture and Natural Resources: Richmond, CA, USA, 2002. [Google Scholar]
- Pareek, S. Ripening physiology: An overview. In Postharvest Ripening Physiology of Crops, 1st ed.; Pareek, S., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1–48. [Google Scholar]
- Montero-Calderón, M.; Cerdas-Araya, M.M. Postharvest Physiology and Storage. In Tropical and Subtropical Fruits: Postharvest Physiology, Processing and Packaging; Siddiq, M., Ed.; Wiley-Blackwell: Oxford, UK, 2012; pp. 17–33. [Google Scholar] [CrossRef]
- Megías, Z.; Martínez, C.; Manzano, S.; Garcia, A.; del Mar, R.-F.M.; Garrido, D.; Valenzuela, J.L.; Jamilena, M. Individual shrink wrapping of zucchini fruit improves postharvest chilling tolerance associated with a reduction in ethylene production and oxidative stress metabolites. PLoS ONE 2015, 10, e0133058. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.C.; Wang, C.Y.; Salveit, M. The Commercial Storage of Fruits, Vegetables, and Florist and Nursery Stocks; Gross, K.C., Wang, C.Y., Salveit, M., Eds.; Agriculture Handbook Number 66; USDA Agricultural Research Service: Beltsville, MD, USA, 2016.
- Salveit, M.E. Cucumber. In The Commercial Storage of Fruits, Vegetables, and Florist and Nursery Stocks; Gross, K.C., Wang, C.Y., Salveit, M., Eds.; USDA Agricultural Research Service: Beltsville, MD, USA, 2016; pp. 302–304. [Google Scholar]
- Perkins-Veazie, P. Okra. In The Commercial Storage of Fruits, Vegetables, and Florist and Nursery Stocks; Gross, K.C., Wang, C.Y., Salveit, M., Eds.; USDA Agricultural Research Service: Beltsville, MD, USA, 2016; pp. 430–432. [Google Scholar]
- Toivonen, P.M. Postharvest physiology of vegetables. In Handbook of Vegetables and Vegetable Processing; Sinha, N., Hui, Y.H., Evranuz, E.Ö., Siddiq, M., Ahmed, J., Eds.; Wiley-Blackwell Publishing: Oxford, UK, 2010; pp. 199–220. ISBN 978-0-8138-1541-1. [Google Scholar]
- Nunes, M.D.N.; Emond, J.P. Storage temperature. In Postharvest Physiology and Pathology of Vegetables; Jerry, A., Bartz, J.K.B., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2003; pp. 209–228. [Google Scholar]
- Cantwell, M. Summary table of optimal handling conditions for fresh produce. In Postharvest Technology of Horticultural Crops; Kader, A.A., Ed.; Agriculture and Natural Resources Publication 3311; University of California: Davis, CA, USA, 2002; pp. 511–518. [Google Scholar]
- Toivonen, P.M.A.; Hodges, D. Abiotic stress in harvested fruits and vegetables. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A.K., Venkteswarlu, B., Eds.; Intech Open Access Publishers: Rijeka, Croatia, 2011; pp. 39–59. [Google Scholar]
- Kays, S.J.; Hayes, M.J. Induction of ripening in the fruits of Momordica charantia by ethylene. Trop. Agric. 1978, 55, 167–172. [Google Scholar]
- Zong, R.-J.; Morris, L.; Cantwell, M. Postharvest physiology and quality of bitter melon (Momordica charantia L.). Postharvest Biol. Technol. 1995, 6, 65–72. [Google Scholar] [CrossRef]
- Megías, Z.; Manzano, S.; Martínez, C.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. Ethylene production by fruits of zucchini cultivars differing in postharvest fruit quality and tolerance to chilling injury. Proccedings of the 10th EUCARPIA Meet Genet Breed Cucurbitaceae, Antalya, Turkey, 15–18 October 2012; pp. 638–642. [Google Scholar]
- Megías, Z.; Manzano, S.; Martínez, C.; García, A.; Aguado, E.; Garrido, D.; del Mar, R.M.; Valenzuela, J.L.; Jamilena, M. Postharvest cold tolerance in summer squash and its association with reduced cold-induced ethylene production. Euphytica 2017, 213, 9. [Google Scholar] [CrossRef]
- Carvajal, F.; Martinez, C.; Jamilena, M.; Garrido, D. Differential response of zucchini varieties to low storage temperature. Sci. Hortic. 2011, 130, 90–96. [Google Scholar] [CrossRef]
- Megías, Z.; Martínez, C.; Manzano, S.; Garcia, A.; del Mar, R.-F.M.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. Ethylene biosynthesis and signaling elements involved in chilling injury and other postharvest quality traits in the non-climacteric fruit of zucchini (Cucurbita pepo). Postharvest Biol. Technol. 2016, 113, 48–57. [Google Scholar] [CrossRef]
- Wang, C.Y.; Adams, D.O. Chilling-induced ethylene production in cucumbers (Cucumis sativus L.). Plant Physiol. 1982, 69, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Hyodo, H.; Fukasawa, R. Ethylene production in kiwifruit. J. Jpn. Soc. Hortic. Sci. 1985, 54, 209–215. [Google Scholar] [CrossRef]
- McCollum, T.G.; McDonald, R.E. Electrolyte leakage, respiration, and ethylene production as indices of chilling injury in grapefruit. HortScience 1991, 26, 1191–1192. [Google Scholar]
- Lafuente, M.T.; Zacarias, L.; Martínez-Téllez, M.A.; Sanchez-Ballesta, M.T.; Dupille, E. Phenylalanine ammonia-lyase as related to ethylene in the development of chilling symptoms during cold storage of citrus fruits. J. Agric. Food Chem 6020, 49, 6020–6025. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, Y.; Chen, X.; Peng, Y.; Hurr, B.M.; Mao, L. The Role of ethylene and calcium in programmed cell death of cold-stored cucumber fruit. J. Food Biochem. 2014, 38, 337–344. [Google Scholar] [CrossRef]
- Zacarías, L.; Lafuente, M.T.; Marcos, J.F.; Saladie, M.; Dupille, E. Regulation of ethylene biosynthesis during cold storage of the chilling-sensitive Fortune mandarin fruit. In Biology and Biotechnology of the Plant Hormone Ethylene III; Vendrell, E., Klee, H., Pech, J.C., Romojaro, F., Eds.; IOS Press: Amsterdam, The Netherlands, 2003; pp. 112–117. [Google Scholar]
- Sapitnitskaya, M.; Maul, P.; McCollum, G.T.; Guy, C.L.; Weiss, B.; Samach, A.; Porat, R. Postharvest heat and conditioning treatments activate different molecular responses and reduce chilling injuries in grapefruit. J. Exp. Bot. 2006, 57, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Maul, P.; McCollum, G.; Guy, C.L.; Porat, R. Temperature conditioning alters transcript abundance of genes related to chilling stress in “Marsh” grapefruit flavedo. Postharvest Biol. Technol. 2011, 60, 177–185. [Google Scholar] [CrossRef]
- Megias, Z.; Martinez, C.; Manzano, S.; Barrera, A.; Rosales, R.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. Cold-induced ethylene in relation to chilling injury and chilling sensitivity in the non-climacteric fruit of zucchini (Cucurbita pepo L.). LWT Food Sci. Technol. 2014, 57, 194–199. [Google Scholar] [CrossRef]
- Lado, J.; Rodrigo, M.J.; Cronje, P.; Zacarías, L. Involvement of lycopene in the induction of tolerance to chilling injury in grapefruit. Postharvest Biol. Technol. 2015, 100, 176–186. [Google Scholar] [CrossRef]
- Glenn, G.M.; Chiou, B.S.; Imam, S.H.; Wood, D.F.; Orts, W.J. Role of cuticles in produce quality and preservation. In Produce Degradation: Pathways and Prevention; Lamikanra, O., Imam, S., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 20–53. [Google Scholar]
- Kubo, Y.; Xue, Y.; Nakatsuka, A.; Mathooko, F.M.; Inaba, A.; Nakamura, R. Expression of a water stress-induced polygalacturonase gene in harvested cucumber fruit. J. Jpn. Soc. Hortic. Sci. 2000, 69, 273–279. [Google Scholar] [CrossRef]
- Lurie, S.; Shapiro, B.; Ben-Yehoshua, S. Effects of water stress and degree of ripeness on rate of senescence of harvested bell pepper fruit. J. Am. Soc. Hortic. Sci. 1986, 111, 880–885. [Google Scholar]
- Massolo, J.F.; Concellón, A.; Chaves, A.R.; Vicente, A.R. 1-Methylcyclopropene (1-MCP) delays senescence, maintains quality and reduces browning of non-climacteric eggplant (Solanum melongena L.) fruit. Postharvest Biol. Technol. 2011, 59, 10–15. [Google Scholar] [CrossRef]
- Rao, C.G. Engineering for Storage of Fruits and Vegetables: Cold Storage, Controlled Atmosphere Storage, Modified Atmosphere Storage; Rao, C.G., Ed.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Sevillano, L.; Sanchez-Ballesta, M.T.; Romojaro, F.; Flores, F.B. Physiological, hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. J. Sci. Food Agric. 2009, 89, 555–573. [Google Scholar] [CrossRef]
- Kratsch, H.A.; Wise, R.R. The ultrastructure of chilling stress. Plant Cell Environ. 2000, 23, 337–350. [Google Scholar] [CrossRef]
- Fernández-Trujillo, J.P.; Martínez, J.A. Ultrastructure of the onset of chilling injury in cucumber fruit. J. Appl. Bot. Food Qual. 2006, 80, 100–110. [Google Scholar]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Martinez-Romero, D.; Guillen, F.; Serrano, M.; Valero, D. Vapour treatments with methyl salicylate or methyl jasmonate alleviated chilling injury and enhanced antioxidant potential during postharvest storage of pomegranates. Food Chem. 2011, 124, 964–970. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, J.; Tao, X.; Zheng, X.; Mao, L. The possible role of BAX and BI-1 genes in chilling-induced cell death in cucumber fruit. Acta Physiol. Plant. 2014, 36, 1345–1351. [Google Scholar] [CrossRef]
- Tatsumi, Y.; Maeda, K.; Murata, T. Morphological changes in cucumber fruit surfaces associated with chilling injury. J. Jpn. Soc. Hortic. Sci. 1987, 56, 187–192. [Google Scholar] [CrossRef]
- Hakim, A.; Purvis, A.C.; Mullinix, B.G. Differences in chilling sensitivity of cucumber varieties depends on storage temperature and the physiological dysfunction evaluated. Postharvest Biol. Technol. 1999, 17, 97–104. [Google Scholar] [CrossRef]
- Carvajal, F.; Palma, F.; Jamilena, M.; Garrido, D. Cell wall metabolism and chilling injury during postharvest cold storage in zucchini fruit. Postharvest Biol. Technol. 2015, 108, 68–77. [Google Scholar] [CrossRef]
- Balandrán-Quintana, R.R.; Mendoza-Wilson, A.M.; Gardea-Béjar, A.A.; Vargas-Arispuro, I.; Martı́nez-Téllez, M. A Irreversibility of chilling injury in zucchini squash (Cucurbita pepo L.) could be a programmed event long before the visible symptoms are evident. Biochem. Biophys. Res. Commun. 2003, 307, 553–557. [Google Scholar] [CrossRef]
- Lim, C.S.; Kang, S.M.; Cho, J.L.; Gross, K.C. Antioxidizing enzyme activities in chilling-sensitive and chilling-tolerant pepper fruit as affected by stage of ripeness and storage temperature. J. Am. Soc. Hortic Sci. 2009, 134, 156–163. [Google Scholar]
- Lim, C.S.; Kang, S.M.; Cho, J.L.; Gross, K.C.; Woolf, A.B. Bell Pepper (Capsicum annuum L.) fruits are susceptible to chilling injury at the breaker stage of ripeness. HortScience 2007, 42, 1659–1664. [Google Scholar]
- Boonsiri, K.; Ketsa, S.; van Doorn, W.G. Seed browning of hot peppers during low temperature storage. Postharvest Biol. Technol. 2007, 45, 358–365. [Google Scholar] [CrossRef]
- Pérez-Gilbert, M.; García-Carmona, F. Characterization of catecholase and cresolase activities of eggplant polyphenol oxidase. J. Agric. Food Chem. 2000, 48, 695–700. [Google Scholar] [CrossRef]
- Concellón, A.; Añón, M.C.; Chaves, A.R. Effect of low temperature storage on physical and physiological characteristics of eggplant fruit (Solanum melongena L.). LWT Food Sci. Technol. 2007, 40, 389–396. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Concellón, A.; Añón, M.C.; Chaves, A.R. Characterization and changes in polyphenol oxidase from eggplant fruit (Solanum melongena L.) during storage at low temperature. Food Chem. 2004, 88, 17–24. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.M.; Collins, J.K. Cultivar, packaging, and storage temperature differences in postharvest shelf life of okra. HortTechnology 1992, 2, 350–352. [Google Scholar]
- Cantwell, M.; Suslow, T. Perishables Handling #107; Department of Plant Sciences, University of California, Davis: Davis, CA, USA, 2001. [Google Scholar]
- Boonyaritthongchai, P.; Srilaong, V.; Wongs-Aree, C.; Techavuthiporn, C. Effect of methyl jasmonate on reducing chilling injury symptom and maintaining postharvest quality of okra (Hibiscus esculentum L.). Acta Hortic. 2013, 1012, 1119–1124. [Google Scholar] [CrossRef]
- Zong, R.; Cantwell, M.; Morris, L.L. Postharvest handling of Asian specialty vegetables under study. Calif Agric. 1993, 47, 27–29. [Google Scholar]
- Han, C.; Zuo, J.H.; Wang, Q.; Dong, H.Z.; Gao, L.P. Salicylic acid alleviates postharvest chilling injury of sponge gourd (Luffa cylindrica). J. Integr. Agric. 2017, 16, 735–741. [Google Scholar] [CrossRef]
- Purvis, A.C. Diphenylamine reduces chilling injury of green bell pepper fruit. Postharvest Biol. Technol. 2002, 25, 41–48. [Google Scholar] [CrossRef]
- Babarinde, G.O.; Fabunmi, O.A. Effects of packaging materials and storage temperature on quality of fresh okra (Abelmoschus esculentus) fruit. Agric. Trop. Subtrop. 2009, 42, 151–156. [Google Scholar]
- Wang, L.; Li, Q.; Cao, J.; Cai, T.; Jiang, W. Keeping quality of fresh-cut bitter gourd (Momordica charantia L.) at low temperature of storage. J. Food Process. Preserv. 2007, 31, 571–582. [Google Scholar] [CrossRef]
- Del Río, L.A.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Parkin, K.L.; Kuo, S.J. Chilling-induced lipid degradation in cucumber (Cucumis sativa L. cv Hybrid C) fruit. Plant Physiol. 1989, 90, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2016, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C.G. Alternative oxidase: A mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int. J. Mol. Sci. 2013, 14, 6805–6847. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fait, A.; Bouchez, D.; Møller, S.G.; Fromm, H. Mitochondrial succinic-semialdehyde dehydrogenase of the γ-aminobutyrate shunt is required to restrict levels of reactive oxygen intermediates in plants. Proc. Natl. Acad. Sci. USA 2003, 100, 6843–6848. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; Lester, G.E.; Munro, K.D.; Toivonen, P.M.A. Oxidative stress: Importance for postharvest quality. HortScience 2004, 39, 924–929. [Google Scholar]
- Toivonen, P.M.A. Postharvest storage procedures and oxidative stress. HortScience 2004, 39, 938–942. [Google Scholar]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Chen, B.; Yang, H. 6-Benzylaminopurine alleviates chilling injury of postharvest cucumber fruit through modulating antioxidant system and energy status. J. Sci. Food. Agric. 2013, 93, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Hariyadi, P.; Parkin, K.L. Chilling-induced oxidative stress in cucumber fruits. Postharvest Biol. Technol. 1991, 1, 33–45. [Google Scholar] [CrossRef]
- Carvajal, F.; Palma, F.; Jamilena, M.; Garrido, D. Preconditioning treatment induces chilling tolerance in zucchini fruit improving different physiological mechanisms against cold injury. Ann. Appl. Biol. 2015, 166, 340–354. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Jamilena, M.; Garrido, D. Putrescine treatment increases the antioxidant response and carbohydrate content in zucchini fruit stored at low temperature. Postharvest Biol. Technol. 2016, 118, 68–70. [Google Scholar] [CrossRef]
- Wang, C.Y. Effect of temperature preconditioning on catalase, peroxidase, and superoxide dismutase in chilled zucchini squash. Postharvest Biol. Technol. 1995, 5, 67–76. [Google Scholar] [CrossRef]
- Zheng, Y.; Fung, R.W.M.; Wang, S.Y.; Wang, C.Y. Transcript levels of antioxidative genes and oxygen radical scavenging enzyme activities in chilled zucchini squash in response to superatmospheric oxygen. Postharvest Biol. Technol. 2008, 47, 151–158. [Google Scholar] [CrossRef]
- Yang, H.; Wu, F.; Cheng, J. Reduced chilling injury in cucumber by nitric oxide and the antioxidant response. Food Chem. 2011, 127, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; He, Z.; Zhao, Y.; Mi, H.; Chen, X.; Mao, L. Maturity-dependent chilling tolerance regulated by the antioxidative capacity in postharvest cucumber (Cucumis sativus L.) fruits. J. Sci. Food. Agric. 2013, 93, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Mi, H.; Zhao, Y.; He, Z.; Mao, L. Effect of maturity stage on the gene expression of antioxidative enzymes in cucumber (Cucumis sativus L.) fruits under chilling stress. J. Integr. Agric. 2013, 12, 1495–1500. [Google Scholar] [CrossRef]
- Martı́nez-Téllez, M.; Ramos-Clamont, M.; Gardea, A.; Vargas-Arispuro, I. Effect of infiltrated polyamines on polygalacturonase activity and chilling injury responses in zucchini squash (Cucurbita pepo L.). Biochem. Biophys. Res. Commun. 2002, 295, 98–101. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Lluch, C.; Jamilena, M.; Garrido, D. Changes in carbohydrate content in zucchini fruit (Cucurbita pepo L.) under low temperature stress. Plant Sci. 2014, 217, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Palma, F.; Carvajal, F.; Jamilena, M.; Garrido, D. Contribution of polyamines and other related metabolites to the maintenance of zucchini fruit quality during cold storage. Plant Physiol. Biochem. 2014, 82, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Buta, J.G. Methyl jasmonate reduces chilling injury in Cucurbita pepo through its regulation of abscisic acid and polyamine levels. Environ. Exp. Bot. 1994, 34, 427–432. [Google Scholar] [CrossRef]
- Gualanduzzi, S.; Baraldi, E.; Braschi, I.; Carnevali, F.; Gessa, C.E.; De Santis, A. Respiration, hydrogen peroxide levels and antioxidant enzyme activities during cold storage of zucchini squash fruit. Postharvest Biol. Technol. 2009, 52, 16–23. [Google Scholar] [CrossRef]
- Couee, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bel, P.; Egea, I.; Sánchez-Ballesta, M.T.; Martinez-Madrid, C.; Fernandez-Garcia, N.; Romojaro, F.; Olmos, E.; Estrella, E.; Bolarin, M.C.; Flores, F.B. Understanding the mechanisms of chilling injury in bell pepper fruits using the proteomic approach. J. Proteom. 2012, 75, 5463–5478. [Google Scholar] [CrossRef]
- Fung, R.W.M.; Wang, C.Y.; Smith, D.L.; Gross, K.C.; Tian, M. MeSA and MeJA increase steady-state transcript levels of alternative oxidase and resistance against chilling injury in sweet peppers (Capsicum annuum L.). Plant Sci. 2004, 166, 711–719. [Google Scholar] [CrossRef]
- Kaneyuki, T.; Noda, Y.; Traber, M.G.; Mori, A.; Packer, L. Superoxide anion and hydroxyl radical scavenging activities of vegetable extracts measured using electron spin resonance. IUBMB Life 1999, 47, 979–989. [Google Scholar] [CrossRef]
- Prohens, J.; Rodríguez-Burruezo, A.; Raigón, M.D.; Nuez, F. Total phenolic concentration and browning susceptibility in a collection of different varietal types and hybrids of eggplant: Implications for breeding for higher nutritional quality and reduced browning. J. Am. Soc. Hortic. Sci. 2007, 132, 638–646. [Google Scholar]
- Fan, L.; Shi, J.; Zuo, J.; Gao, L.; Lv, J.; Wang, Q. Methyl jasmonate delays postharvest ripening and senescence in the non-climacteric eggplant (Solanum melongena L.) fruit. Postharvest Biol. Technol. 2016, 120, 76–83. [Google Scholar] [CrossRef]
- Mao, L.; Pang, H.; Wang, G.; Zhu, C. Phospholipase D and lipoxygenase activity of cucumber fruit in response to chilling stress. Postharvest Biol. Technol. 2007, 44, 42–47. [Google Scholar] [CrossRef]
- Mao, L.-C.; Wang, G.-Z.; Zhu, C.-G.; Pang, H.-Q. Involvement of phospholipase D and lipoxygenase in response to chilling stress in postharvest cucumber fruits. Plant Sci. 2007, 172, 400–405. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Fortiz, J.; Cruz, R.; Baez, R.; Wang, C.Y. Methyl Jasmonate reduces chilling injury and maintains postharvest quality of mango fruit. J. Agric. Food Chem. 2000, 48, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Lurie, S. Postharvest heat treatments. Postharvest Biol. Technol. 1998, 14, 257–269. [Google Scholar] [CrossRef]
- Bar-Yosef, A.; Alkalai-Tuvia, S.; Perzelan, Y.; Aharon, Z.; Llic’, Z.; Lurie, S.; Fallik, E. Effect of shrink packaging in combination with rinsing and brushing treatment on chilling injury and decay of sweet pepper during storage. Adv. Hortic. Sci. 2009, 23, 225–230. [Google Scholar]
- Sato, Y.; Murakami, T.; Funatsuki, H.; Matsuba, S.; Saruyama, H.; Tanida, M. Heat shock-mediated APX gene expression and protection against chilling injury in rice seedlings. J. Exp. Bot. 2001, 52, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, M.; Yang, H. Postharvest chitosan-g-salicylic acid application alleviates chilling injury and preserves cucumber fruit quality during cold storage. Food Chem. 2015, 174, 558–563. [Google Scholar] [CrossRef] [PubMed]
- McCollum, T.G.; Doostdar, H.; Mayer, R.T.; McDonald, R.E. Immersion of cucumber fruit in heated water alters chilling-induced physiological changes. Postharvest Biol. Technol. 1995, 6, 55–64. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Trajković, R.; Pavlović, R.; Alkalai-Tuvia, S.; Perzelan, Y.; Fallik, E. Effect of heat treatment and individual shrink packaging on quality and nutritional value of bell pepper stored at suboptimal temperature. Int. J. Food Sci. Technol. 2012, 47, 83–90. [Google Scholar] [CrossRef]
- Rodriguez, S. del C.; López, B.; Chavesz, A.R. Effect of different treatments on the evolution of polyamines during refrigerated storage of eggplants. J. Agric. Food Chem. 2001, 49, 4700–4705. [Google Scholar] [CrossRef]
- Li, N.; Parsons, B.L.; Liu, D.; Mattoo, A.K. Accumulation of wound-inducible ACC synthase transcript in tomato fruit is inhibited by salicylic acid and polyamines. Plant Mol. Biol. 1992, 18, 477–487. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, W.; Rossouw, W.; Korsten, L. Comparison of safe alternative dipping treatment to maintain quality of zucchini. J. Food Qual. 2016, 39, 109–115. [Google Scholar] [CrossRef]
- Yi Wang, C. Combined treatment of heat shock and low temperature conditioning reduces chilling injury in zucchini squash. Postharvest Biol. Technol. 1994, 4, 65–73. [Google Scholar] [CrossRef]
- Wang, H.; Qian, Z.; Ma, S.; Zhou, Y.; Patrick, J.W.; Duan, X.; Jiang, Y.; Qu, H. Energy status of ripening and postharvest senescent fruit of litchi (Litchi chinensis Sonn.). BMC Plant Biol. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, S. Pre-storage cold acclimation maintained quality of cold-stored cucumber through differentially and orderly activating ROS scavengers. Postharvest Biol. Technol. 2017, 129, 1–8. [Google Scholar] [CrossRef]
- Kader, A.A. Biochemical and physiological basis for effects of controlled and modified atmospheres on fruits and vegetables. Food Technol. 1986, 40, 99–100. [Google Scholar]
- Kader, A.A. Atmosphere modification. In Postharvest Physiology and Pathology of Vegetables; Bartz, J.A., Brecht, J.K., Weichmann, J., Eds.; Marcel Dekker: New York, NY, USA, 2003; p. 733. [Google Scholar]
- Kader, A.A.; Saltveit, M.E. Respiración and gas exchange. In Postharvest Physiology and Pathology of Vegetables; Bartz, J.A., Brecht, J.K., Weichmann, J., Eds.; Marcel Dekker: New York, NY, USA, 2003; pp. 7–29. ISBN 0-8247-0687-0. [Google Scholar]
- Kader, A.A.; Zagory, D.; Kerbel, E.L.; Wang, C.Y. Modified atmosphere packaging of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 1989, 28, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Scully, A.D.; Horsham, M.A. Active packaging for fruits and vegetables. In Intelligent and Active Packaging for Fruits and Vegetables; Wilson, C.L., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 57–71, ISBN-13: 978-0849391668. [Google Scholar]
- Yahia, E.M. Introduction. In Modified and Controlled Atmospheres for the Storage, Transportation and Packaging of Horticultural Commodities; Yahia, E.M., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2009; pp. 1–16, ISBN-13: 978-1420069570. [Google Scholar]
- Mangaraj, S.G.T. Modified atmosphere packaging—An ideal food preservation technique. J. Food Sci. Technol. 2009, 46, 399–410. [Google Scholar]
- Kanellis, A.K.; Tonutti, P.P.P. Biochemical and molecular aspects of modified and controlled atmospheres. In Modified and Controlled Atmospheres for the Storage, Transportation and Packaging of Horticultural Commodities; Yahia, E.M., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2009; pp. 553–567, ISBN-13: 978-14200695702009. [Google Scholar]
- Mishra, B.; Khatkar, B.S.; Garg, M.K.; Wilson, L.A. Permeability of edible coatings. J. Food Sci. Technol. 2010, 47, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, K.; Nakano, K. Optimal design of modified atmosphere packaging for alleviating chilling injury in cucumber fruit. Environ. Control Biol. 2014, 52, 233–240. [Google Scholar] [CrossRef]
- Wang, C.Y.; Qi, L. Modified atmosphere packaging alleviates chilling injury in cucumbers. Postharvest Biol. Technol. 1997, 10, 195–200. [Google Scholar] [CrossRef]
- Serrano, M.; Pretel, M.T.; Martínez-Madrid, M.C.; Romojaro, F.; Riquelme, F. CO2 treatment of zucchini squash reduces chilling-induced physiological changes. J. Agric. Food Chem. 1998, 46, 2465–2468. [Google Scholar] [CrossRef]
- Wang, C.Y.; Ji, Z.L. Effect of low-oxygen storage on chilling injury and polyamines in zucchini squash. Sci. Hortic. 1989, 39, 1–7. [Google Scholar] [CrossRef]
- Zagory, D.; Kader, A.A. Modified atmosphere packaging of fresh produce. Food Technol. 1988, 42, 70–77. [Google Scholar]
- Serrano, M.; Martínez-Madrid, M.C.; Pretel, M.T.; Riquelme, F.; Romojaro, F. Modified Atmosphere Packaging minimizes increases in putrescine and abscisic acid levels caused by chilling injury in pepper fruit. J. Agric. Food Chem. 1997, 45, 1668–1672. [Google Scholar] [CrossRef]
- Sahoo, N.R.; Bal, L.M.; Pal, U.S.; Sahoo, D. A Comparative study on the effect of packaging material and storage environment on shelf life of fresh bell-pepper. J. Food Meas Charact. 2014, 8, 164–170. [Google Scholar] [CrossRef]
- Rao, D.S.; Rao, K.G.; Krishnamurthy, S. Extension of shelf life of cucumber by modified atmosphere packaging (MAP) and shrink wrapping. Indian Food Pack. 2000, 54, 65–71. [Google Scholar]
- Dhall, R.K.; Sharma, S.R.; Mahajan, B.V.C. Effect of shrink wrap packaging for maintaining quality of cucumber during storage. J. Food Sci. Technol. 2012, 49, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, B.V.C.; Dhillon, W.S.; Sidhu, M.K.; Jindal, S.K.; Dhaliwal, M.S.; Singh, S. Effect of packaging and storage environments on quality and shelf-life of bell pepper (Capsicum annuum). Indian J. Agric. Sci. 2016, 86, 738–742. [Google Scholar]
- Shi, J.Y.; Gao, L.P.; Zuo, J.H.; Wang, Q.; Wang, Q.; Fang, L. Effect of exogenous ceramide treatment on chilling tolerance in green bell pepper fruits. Mod. Food Sci. Technol. 2016, 32, 164–170. [Google Scholar]
- Ghaouth, A.; Arul, J.; Ponnampalam, R.; Boulet, M. Use of chitosan coating to reduce water loss and maintain quality of cucumber and bell pepper fruits. J. Food Process. Preserv. 1991, 15, 359–368. [Google Scholar] [CrossRef]
- Mohammadi, A.; Hashemi, M.; Hosseini, S.M. Postharvest treatment of nanochitosan-based coating loaded with Zataria multiflora essential oil improves antioxidant activity and extends shelf-life of cucumber. Innov. Food Sci. Emerg. Technol. 2016, 33, 580–588. [Google Scholar] [CrossRef]
- Abbasi, N.A.; Ashraf, S.; Ali, I.; Butt, S.J. Enhancing storage life of bell pepper by UV-C irradiation and edible coatings. Pak. J. Agric. Sci. 2015, 52, 403–411. [Google Scholar]
- Gholamipour, F.K.; Kamari, S.; Ghasemnezhad, M.; Ghazvini, R.F. Effect of chitosan coating on weight loss and postharvest quality of green pepper (Capsicum annum L.) Fruits. Acta Hortic. 2010, 877, 821–826. [Google Scholar] [CrossRef]
- Beaulieu, J.C.; Park, H.S.; Mims, A.G.B.; Kuk, M.S. Extension of green bell pepper shelf life using oilseed-derived lipid films from soapstock. Ind. Crops Prod. 2009, 30, 271–275. [Google Scholar] [CrossRef]
- Lerdthanangkul, S.; Krochta, J.M. Edible coating effects on postharvest quality of green bell peppers. J. Food Sci. 1996, 61, 176–179. [Google Scholar] [CrossRef]
- de Souza, P.A.; Aroucha, E.M.M.; de Souza, A.E.; da Costa, A.R.; de Ferreira, G.S.; Bezerra Neto, F. Conservação pós-colheita de berinjela com revestimentos de fécula de mandioca ou filme de PVC. Hortic. Bras. 2009, 27, 235–239. [Google Scholar] [CrossRef]
- Han, C.; Zuo, J.; Wang, Q.; Xu, L.; Zhai, B.; Wang, Z.; Dong, H.; Gao, L. Effects of chitosan coating on postharvest quality and shelf life of sponge gourd (Luffa cylindrica) during storage. Sci. Hortic. 2014, 166, 1–8. [Google Scholar] [CrossRef]
- Abeles, F.B.; Morgan, P.W.; Saltveit, M.E. The role of ethylene in agriculture. In Ethylene in Plant Biology, 2nd ed.; Abeles, F.B., Morgan, P.W., Saltveit, M.E., Eds.; Academic Press: Cambridge, MA, USA, 1992; pp. 264–296. [Google Scholar] [CrossRef]
- Barry, C.S.; Giovannoni, J.J. Ethylene and fruit ripening. J. Plant Growth Regul. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Li, Z.Q.; Qiao, Y.S.; Tong, Z.G.; Zhou, J.; Zhang, Z. Effect of ethylene and 1-MCP on post-harvest physiology and on expression of the ethylene receptor genes PpETR3 and PpERS2 in pear (Pyrus pyrifolia Nakai “Kikusui”) fruit. J. Hortic. Sci. Biotechnol. 2010, 85, 71–77. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Watkins, C.B. The use of 1-methylcyclopropene (1-MCP) on fruits and vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Yang, Z.; Zheng, Y. Effect of 1-methylcyclopene on senescence and quality maintenance of green bell pepper fruit during storage at 20 °C. Postharvest Biol. Technol. 2012, 70, 1–6. [Google Scholar] [CrossRef]
- Smith, A.W.J.; Poulston, S.; Rowsell, L.; Terry, L.A.; Anderson, J.A. A New Palladium-based ethylene scavenger to control ethylene-induced ripening of climacteric fruit. Platin. Met. Rev. 2009, 53, 112–122. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, T.; Gao, L.; Pang, J.; Yang, N. Effect of brassinolide on chilling injury of green bell pepper in storage. Sci. Hortic. 2012, 144, 195–200. [Google Scholar] [CrossRef]
- Gao, H.; Kang, L.; Liu, Q.; Cheng, N.; Wang, B.; Cao, W. Effect of 24-epibrassinolide treatment on the metabolism of eggplant fruits in relation to development of pulp browning under chilling stress. J. Food Sci. Technol. 2015, 52, 3394–3401. [Google Scholar] [CrossRef] [PubMed]
- Swamy, P.M.; Smith, B.N. Role of abscisic acid in plant stress tolerance. Curr. Sci. 1999, 76, 1220–1227. [Google Scholar]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.H.; Endo, A.; Zhou, L.; Penney, J.; Chen, H.-C.; Arroyo, A.; Leon, P.; Nambara, E.; Asami, T.; Seo, M. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 2002, 14, 2723–2743. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Hirai, N.; Matsumoto, C.; Ohigashi, H.; Ohta, D.; Sakata, K.; Mizutani, M. Arabidopsis CYP707As encode (+)- abscisic acid 8’-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid. Plant Physiol. 2004, 134, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Cross-talk between calcium and reactive oxygen species originated from NADPH oxidase in abscisic acid-induced antioxidant defence in leaves of maize seedlings. Plant Cell Environ. 2003, 26, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Kurkela, S.; Franck, M. Cloning and characterization of a cold-and ABA-inducible Arabidopsis gene. Plant Mol. Biol. 1990, 15, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Lang, V.; Palva, E.T. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Mol. Biol. 1992, 20, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Mantyla, E.; Lang, V.; Palva, E.T. Role of abscisic acid in drought-induced freezing tolerance, cold acclimation, and accumulation of LT178 and RAB18 proteins in Arabidopsis thaliana. Plant Physiol. 1995, 107, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Asghari, M.; Aghdam, M.S. Impact of salicylic acid on post-harvest physiology of horticultural crops. Trends Food Sci. Technol. 2010, 21, 502–509. [Google Scholar] [CrossRef]
- Hu, Q.; Wang, T.; Zhou, M.; Xue, J.; Luo, Y. In vitro antioxidant-activity evaluation of gallic-acid-grafted chitosan conjugate synthesized by free-radical-induced grafting method. J. Agric. Food Chem. 2016, 64, 5893–5900. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Ziosi, V.; Bonghi, C.; Bregoli, A.M.; Trainotti, L.; Biondi, S.; Sutthiwal, S.; Kondo, S.; Costa, G.; Torrigiani, P. Jasmonate-induced transcriptional changes suggest a negative interference with the ripening syndrome in peach fruit. J. Exp. Bot. 2008, 59, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of CBF expression-C-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell. 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Liu, H.; Xiong, L. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [PubMed]
- Carrera, R.; Zapata, S.; García, A.; Aguado, E.; Rebolloso, M.M.; Manzano, S.; Jamilena, M.; Valenzuela, J.L. Efectos de los tratamientos de Metil Jasmonato y Ácido Salicílico en la calidad poscosecha y daños por frío de frutos de berenjena. Actas Port. Hortic. 2016, 28, 239–244. [Google Scholar]
- Zapata, S.; Carrera, R.; Manzano, S.; García, A.; Aguado, E.; Garrido, D.; Rebolloso, M.M.; Manzano, S.; Valenzuela, J.L.; Valenzuela, J.L. Efectos de los tratamientos de Metil Jasmonato y Ácido Salicílico en la reducción del daño por frío en calabacín. Actas Port. Hortic. 2016, 28, 232–238. [Google Scholar]
- Purvis, A.C. Role of the alternative oxidase in limiting superoxide production by plant mitochondria. Physiol. Plant. 1997, 100, 165–170. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Zhu, S.; Wang, Y. Postharvest application of MeJA and NO reduced chilling injury in cucumber (Cucumis sativus) through inhibition of H2O2 accumulation. Postharvest Biol. Technol. 2016, 119, 77–83. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Ramos, J.M.; Jamilena, M.; Garrido, D. Effect of putrescine application on maintenance of zucchini fruit quality during cold storage: Contribution of GABA shunt and other related nitrogen metabolites. Postharvest Biol. Technol. 2015, 99, 131–140. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, B.; Li, W.; Song, H.; Yu, Y.; Chen, J. Polyamines enhance chilling tolerance of cucumber (Cucumis sativus L.) through modulating antioxidative system. Sci. Hortic. 2009, 122, 200–208. [Google Scholar] [CrossRef]
- Wink, D.A.; Mitchell, J.B. Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic. Biol. Med. 1998, 25, 434–456. [Google Scholar] [CrossRef]
- Soegiarto, L.; Wills, R.B.H. Short term fumigation with nitric oxide gas in air to extend the postharvest life of broccoli, green bean, and bok choy. Horttechnology 2004, 14, 538–540. [Google Scholar]
- Izumi, H.; Watada, A.E. Calcium treatment to maintain quality of zucchini squash slices. J. Food Sci. 1995, 60, 789–793. [Google Scholar] [CrossRef]
- Massolo, J.F.; Lemoine, M.L.; Chaves, A.R.; Concellón, A.; Vicente, A.R. Benzyl-aminopurine (BAP) treatments delay cell wall degradation and softening, improving quality maintenance of refrigerated summer squash. Postharvest Biol. Technol. 2014, 93, 122–129. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, T.; Zuo, J.; Gao, L.; Fan, L. Amelioration of postharvest chilling injury in sweet pepper by glycine betaine. Postharvest Biol. Technol. 2016, 112, 114–120. [Google Scholar] [CrossRef]
- Liu, L.; Wei, Y.; Shi, F.; Liu, C.; Liu, X.; Ji, S. Intermittent warming improves postharvest quality of bell peppers and reduces chilling injury. Postharvest Biol. Technol. 2015, 18, 18–25. [Google Scholar] [CrossRef]
- Wang, C.Y. Effect of abscisic acid on chilling injury of zucchini squash. J. Plant Growth Regul. 1991, 10, 101–105. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A.; Krishnamoorthi, S. Emerging techniques to decipher microRNAs (miRNAs) and their regulatory role in conferring abiotic stress tolerance of plants. Plant Biotechnol. Rep. 2016, 10, 185–205. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kwon, E.-J.; Kwon, S.-Y.; Jeong, Y.-J.; Lee, E.-M.; Jo, M.-H.; Kim, H.-S.; Woo, I.-S.; Shinmyo, A.; Yoshida, K. Transgenic cucumber fruits that produce elevated level of an anti-aging superoxide dismutase. Mol. Breed. 2003, 11, 213–220. [Google Scholar] [CrossRef]
- Chatzidimitriadou, K.; Nianiou-Obeidat, I.; Madesis, P.; Perl-Treves, R.; Tsaftaris, A. Expression of SOD transgene in pepper confer stress tolerance and improve shoot regeneration. Electron. J. Biotechnol. 2009, 12, 7–8. [Google Scholar] [CrossRef]
- Sanmartin, M.; Drogoudi, P.D.; Lyons, T.; Pateraki, I.; Barnes, J.; Kanellis, A.K. Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta 2003, 216, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.-M.; Chen, C.-C.; Chen, S.-P.; Lin, K.-H.; Chen, L.-R.; Su, Y.-H.; Yen, H.-C. Overexpression of the ascorbate peroxidase gene from eggplant and sponge gourd enhances flood tolerance in transgenic Arabidopsis. J. Plant Res. 2017, 130, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.M.; Chen, L.F.O.; Shih, S.W.; Lin, K.H. Expression of eggplant ascorbate peroxidase increases the tolerance of transgenic rice plants to flooding stress. J. Plant Biochem. Biotechnol. 2015, 24, 257–267. [Google Scholar] [CrossRef]
- Choi, H.W.; Hwang, B.K. The pepper extracellular peroxidase CaPO2 is required for salt, drought and oxidative stress tolerance as well as resistance to fungal pathogens. Planta 2012, 235, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene CaLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010, 152, 948–967. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Han, S.-W.; Hwang, I.S.; Kim, D.S.; Hwang, B.K.; Lee, S.C. The pepper lipoxygenase CaLOX1 plays a role in osmotic, drought and high salinity stress response. Plant Cell Physiol. 2015, 56, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.J.; Kim, S.J.; Kim, W.T.; Park, K.Y. Constitutive S-adenosylmethionine decarboxylase gene expression increases drought tolerance through inhibition of reactive oxygen species accumulation in Arabidopsis. Planta 2014, 239, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Rathore, M.; Goyary, D.; Khare, N.; Anandhan, S.; Pande, V.; Ahmed, Z. Marker-free transgenic cucumber expressing Arabidopsis CBF1 gene confers chilling stress tolerance. Biol. Plant. 2012, 56, 57–63. [Google Scholar] [CrossRef]
- Liu, L.; Duan, L.; Zhang, J.; Zhang, Z.; Mi, G.; Ren, H. Cucumber (Cucumis sativus L.) over-expressing cold-induced transcriptome regulator ICE1 exhibits changed morphological characters and enhances chilling tolerance. Sci. Hortic. 2010, 124, 29–33. [Google Scholar] [CrossRef]
- Zhang, Y.; Chu, G.; Hu, Z.; Gao, Q.; Cui, B.; Tian, S.; Wang, B.; Chen, G. Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Mol. Breed. 2016, 36, 54. [Google Scholar] [CrossRef]
- Seong, E.-S.; Choi, D.-I.; Cho, H.-S.; Lim, C.-K.; Cho, H.-J.; Wang, M.-H. Characterization of a stress-responsive ankyrin repeat-containing zinc finger protein of Capsicum annuum (CaKR1). BMB Rep. 2007, 40, 952–958. [Google Scholar] [CrossRef]
- Yi, S.Y.; Lee, D.J.; Yeom, S.-I.; Yoon, J.; Kim, Y.-H.; Kwon, S.-Y.; Choi, D. A novel pepper (Capsicum annuum) receptor-like kinase functions as a negative regulator of plant cell death via accumulation of superoxide anions. New Phytol. 2010, 185, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Sun, X.; Wang, X.; Shi, Q.; Yang, X.; Yang, F. Involvement of a cucumber MAPK gene (CsNMAPK) in positive regulation of ROS scavengence and osmotic adjustment under salt stress. Sci. Hortic. 2011, 127, 488–493. [Google Scholar] [CrossRef]
- Sarowar, S.; Kim, E.N.; Kim, Y.J.; Ok, S.H.; Kim, K.D.; Hwang, B.K.; Shin, J.S. Overexpression of a pepper ascorbate peroxidase-like 1 gene in tobacco plants enhances tolerance to oxidative stress and pathogens. Plant Sci. 2005, 169, 55–63. [Google Scholar] [CrossRef]

{kind=link}
| Immature Fruit | Symptoms | Threshold Temperature (°C) | References |
|---|---|---|---|
| Cucumber | Surface pitting, increased yellowing and disease susceptibility, water-soaked areas of the flesh | 10–12 | [26,38,55] |
| Eggplant | Surface pitting and scald, browning of the flesh and seeds | 8–12 | [51] |
| Bell pepper | Surface pitting, water-soaked areas, seed browning, and decay | [62,74] | |
| Okra | Discoloration; water-soaked areas; surface pitting; exuding lesions, and decay by mould or mildew, calyx discoloration | 7–10 | [27,75] |
| Zucchini | Surface pitting, large sunken areas, dehydration, discoloration | 7–10 | [24,36] |
| Bitter gourd | Pitting that coalesced to form large sunken dark brown pits; surface discoloration; internal tissue breakdown; decay | 8–10 | [76] |
| Postharvest Technologies | Technology | Species | Effects | References |
|---|---|---|---|---|
| Physical | Heat treatment | Cucumber | Reduced electrolyte leakage, chilling-induced ethylene, and ACS and ACO activity | [117] |
| Reduced electrolyte leakage and MDA, and enhanced PLD and LOX activity | [110,111] | |||
| Green bell pepper | Reduced CI, electrolyte leakage and LOX activity | [114] | ||
| Reduced weight loss, softening, decay and CI | [118] | |||
| Enhanced PA content and increased PAL and PPO activity | [112] | |||
| Reduced CI, maintained firmness, and delayed unsaturated fatty acid accumulation | [187] | |||
| Zucchini | Induced HSP genes, maintained flesh firmness | [113] | ||
| Eggplant | Retarded CI, reduced spermidine | [119] | ||
| Temperature preconditioning treatment | Zucchini | Alleviated CI and weight loss, reducedH2O2, MDA and ascorbic acid content, and induced activity of antioxidant enzymes | [46,91,93] | |
| Cucumber | Increased soluble solids, ascorbic acid, and MDA, O2− and H2O2, induced activity of antioxidant enzymes, and the scavengers AsA and glutathione | [124] | ||
| Controlled atmospheres. CO2 and O2 treatments | Cucumber | Alleviated chilling injury, weight loss and changed in peel colour, maintenance of electrolyte leakage and MDA | [134] | |
| Zucchini | Reduced CI, increased levels of spermidine, spermine and total phenolics, induced activities of alternative oxidase, SOD, APX and CAT | [94,136,137] | ||
| Controlled atmospheres. Use of plastic covers | Green bell pepper | Reduced CI and weight loss, maintained of ACC, Put, and ABA levels, reduced ascorbic acid content | [139,140] | |
| Okra | Reduced weight loss and ascorbic acid content, increased titratable acidity | [75] | ||
| Eggplant | Retarded chilling injury, decreased spermidine levels | [119] | ||
| Cucumber | Reduced weight loss, decay and fruit deformation, maintenance of freshness, colour and firmness | [141,142] | ||
| Green bell pepper | Reduced CI, weight loss, membrane leakage and LOX activity, induction of HSP from the HSP70 family | [114,118,140,143] | ||
| Zucchini | Reduced ethylene production and ethylene gene expression, reduced H2O2 and MDA | [24] | ||
| Ceramide coating | Green bell pepper | Maintenance of membrane integrity, reduced MDA, enhanced activity of POD, CAT, and APX | [144] | |
| Chitosan coating | Zucchini | Reduced CI, preservation of flesh firmness | [121] | |
| Cucumber | Reduced CI, electrolyte leakage and MDA accumulation, increased content of soluble solids, chlorophyll and ascorbic acid, SA, and induced activity of SOD, CAT, APX and GR | [116] | ||
| Sponge gourd | Delayed PPO, increased content of ascorbic acid and total phenolics | [152] | ||
| Chemical | 1-MCP | Green bell pepper | Delaying senescence associated with enhanced antioxidant enzyme activities | [158] |
| Zucchini | Reduced fruit weight loss, respiration rate and cold-induced ethylene, reduced expression of ethylene genes | [37] | ||
| Eggplant | Reduced weight loss and browning, and reduced of PAL, PPO and POD activity, and total phenolics | [51] | ||
| Brassinosteroids | Green bell pepper | Decreased electrolyte leakage and MDA content, enhanced CAT, APX, and GR activities | [160] | |
| Eggplant | Maintenance of membrane integrity and moisture and reduced flesh browning, reduced phenolic accumulation and repressed PAL, PPO, and POD activities | [161] | ||
| ABA | Zucchini | Delayed development of CI symptoms | [188] | |
| Salicyloyl chitosan coating | Cucumber | Higher total soluble solids, chlorophyll and ascorbic acid content, reduced electrolyte leakage and MDA, and induction of SOD, CAT, APX and GR | [116] | |
| SA and MeSA | Sponge gourd | Higher antioxidant activity reduce MDA, enhanced SOD, CAT, APX activities | [73] | |
| MeSA and MeJA | Green bell pepper | Increased expression of AOX gene | [106,178] | |
| Cucumber | Reduced H2O2 accumulation enhanced catalase activity | [179] | ||
| Eggplant | Reduced ethylene production | [176] | ||
| MeJA | Cucumber | Induced chilling tolerance by inhibiting H2O2 generation and CAT activity | [179] | |
| PAs | Green bell pepper | Reduction of ethylene production | [112] | |
| Zucchini | Induction of APX, CAT and GR activities, increased content of ascorbate, FRAP, glucose, fructose and raffinose, and reduced LOX activity | [98,99,122,180] | ||
| Nitric oxide | Cucumber | Reduced lipid peroxidation, O2− and H2O2 accumulation, and enhanced CAT, SOD, APX and POD activities | [95,176] | |
| Green beans | Shelf life extension | [183] | ||
| 6-BA | Cucumber | Increased chlorophyll, ascorbic acid, total phenolic contents, and antioxidant capacity, reduced O•−2, H2O2 and lipid peroxidation, increased activities of SOD, CAT, APX, GR and ATP | [89] | |
| CK | Zucchini | Slower deterioration and dehydration, phenolic compound accumulation, and decreased pectin and sugar solubility, delayed cell wall dismantling | [185] | |
| GB | Green bell pepper | Reduction in cellular leakage, MDA content, and lipid peroxidation increased activity and induced gene expression of POD, CAT, APX, and GR | [186] |
| Species | Transgene | Effect of Overexpression/Silencing | References |
|---|---|---|---|
| Cucumber | Cassava SOD | Anti-oxidative cucumber as a functional cosmetic material | [190] |
| Cucumber Nitric Oxide Synthase Associated 1 gene CsNOA1 | Chilling tolerance of cucumber seedling | [179] | |
| Arabidopsis CBF1 | Protection against chilling stress in cucumber leaves | [199] | |
| Cucumber Mitogen-activated protein kinase gene CsNMAPK | ROS scavenge and osmotic adjustment in cucumbers under salt stress | [204] | |
| Cucumber Inducer of CBF expression 1 (ICE1) | Enhanced chilling tolerance | [200] | |
| Cucumber APX | Alteration in ascorbate and glutathione redox states and increased sensitivity to ozone-induced oxidative stress in tobacco | [192] | |
| Eggplant | Eggplant APX (SmAPX) | Greater resistance to flooding and less oxidative injury in Arabidopsis and rice | [193,194] |
| Eggplant SmMYB1 gene encoding a R2R3 MYB transcription factor | Increasing anthocyanin content in different organs, including fruit peels, and enhanced tolerance to seedling freezing stress | [201] | |
| Bell pepper | Pepper LOX gene CaLOX1 | Reduction of lipid peroxidation and H2O2, associated with high salinity and drought stress tolerance, and defence against pathogens in Arabidopsis | [196,197] |
| Pepper PX gene CaPO2 | Drought and oxidative stress tolerance in Arabidopsis | [195] | |
| Pepper S-adenosylmethionine decarboxylase gene SAMDC | Reduced ROS production and drought tolerance in Arabidopsis | [198] | |
| Capsicum annuumRECEPTOR-LIKE KINASE 1 (CaRLK1) | Supress plant cell death by increasing concentration of superoxide anion | [203] | |
| Tomato Cu/Zn SOD | Increased resistance to oxidative damage and improved shoot regeneration in pepper | [191] | |
| Pepper ankyrin repeat domain zinc finger transcription factor gene CaKR1 | Reduced levels of free oxygen radicals. Enhanced tolerance to salinity and oxidative stress in tomato | [63] | |
| Pepper ascorbate peroxidase-like 1 gene CaPOA1 | Enhances tolerance to oxidative stress and pathogens in tobacco | [205] | |
| Sponge gourd | Luffa cylindrica ASCORBATE PEROXIDASE (LcAPX) | Enhanced flood tolerance in transgenic Arabidopsis plants by reducing H2O2 and MDA | [193] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, J.L.; Manzano, S.; Palma, F.; Carvajal, F.; Garrido, D.; Jamilena, M. Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions. Int. J. Mol. Sci. 2017, 18, 1467. https://doi.org/10.3390/ijms18071467
Valenzuela JL, Manzano S, Palma F, Carvajal F, Garrido D, Jamilena M. Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions. International Journal of Molecular Sciences. 2017; 18(7):1467. https://doi.org/10.3390/ijms18071467
Chicago/Turabian StyleValenzuela, Juan Luis, Susana Manzano, Francisco Palma, Fátima Carvajal, Dolores Garrido, and Manuel Jamilena. 2017. "Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions" International Journal of Molecular Sciences 18, no. 7: 1467. https://doi.org/10.3390/ijms18071467