Impact of Novel Prebiotic Galacto-Oligosaccharides on Various Biomarkers of Colorectal Cancer in Wister Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Body Weight Changes
2.2. Aberrant Crypt Foci (ACF)
2.3. Short Chain Fatty Acids (SCFAs)
2.4. Bacterial Enzyme
2.5. Bacterial Populations
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals, Treatments and Diets
4.3. Preparation of GalOS
4.4. Measurement of Body Mass
4.5. Aberrant Crypt Foci (ACF) Analysis
4.6. Samples Collection and Enzyme Analysis
4.7. Short Chain Fatty Acids (SCFAs)
4.8. Microbial Analysis
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CRC | Colorectal cancer |
| ACF | Aberrant crypt foci |
| GalOS | Galacto-oligosaccharides |
| DMH | 1,2-Dimethylhydrazine dihydrochloride |
| SCFAs | Short chain fatty acids |
| ITF | Inulin-type fructans |
| DP | Degree of polymerization |
| FOS | Fructo-oligosaccharides |
| oNPG | Ortho-nitrophenyl-β-galactopyranoside |
| HED | Human equivalent dose |
| EDTA | Ethylenediaminetetraacetic acid |
| β-Gal | β-Galactosidase |
| HPAEC-PAD | High performance anion exchange chromatography with pulsed amperometric detection |
References
- Medicine, I.O. Opportunities in the Nutrition and Food Sciences: Research Challenges and the Next Generation of Investigators; The National Academies Press: Washington, DC, USA, 1994; p. 328. [Google Scholar]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Ebert, M.N.; Beyer-Sehlmeyer, G.; Liegibel, U.M.; Kautenburger, T.; Becker, T.W.; Pool-Zobel, B.L. Butyrate induces glutathione S-transferase in human colon cells and protects from genetic damage by 4-hydroxy-2-nonenal. Nutr. Cancer 2001, 41, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Petzelbauer, I.; Reiter, A.; Splechtna, B.; Kosma, P.; Nidetzky, B. Transgalactosylation by thermostable β-glycosidases from Pyrococcus furiosus and Sulfolobus solfataricus. Binding interactions of nucleophiles with the galactosylated enzyme intermediate make major contributions to the formation of new β-glycosides during lactose conversion. Eur. J. Biochem. 2000, 267, 5055–5066. [Google Scholar] [PubMed]
- De Roode, B.M.; Franssen, M.C.; van der Padt, A.; Boom, R.M. Perspectives for the industrial enzymatic production of glycosides. Biotechnol. Prog. 2003, 19, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Rao, C.V. Molecular markers and targets for colorectal cancer prevention. Acta Pharmacol. Sin. 2008, 29, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Colorectal carcinogenesis: Review of human and experimental animal studies. J. Carcinog. 2009, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B., 3rd. Epidemiology, disease progression, and economic burden of colorectal cancer. J. Manag. Care Pharm. 2007, 13, S5–S18. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D.; Milner, J.A. Gastrointestinal microflora, food components and colon cancer prevention. J. Nutr. Biochem. 2009, 20, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Raju, J. Azoxymethane-induced rat aberrant crypt foci: Relevance in studying chemoprevention of colon cancer. World J. Gastroenterol. 2008, 14, 6632–6635. [Google Scholar] [CrossRef] [PubMed]
- Namasivayam, N. Chemoprevention in experimental animals. Ann. N. Y. Acad. Sci. 2011, 1215, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Schoen, R.E. Aberrant crypt foci: Are they intermediate endpoints of colon carcinogenesis in humans? Curr. Opin. Gastroenterol. 2009, 25, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P.; Good, C.K. The significance of aberrant crypt foci in understanding the pathogenesis of colon cancer. Toxicol. Lett. 2000, 112–113, 395–402. [Google Scholar] [CrossRef]
- Khare, S.; Chaudhary, K.; Bissonnette, M.; Carroll, R. Aberrant crypt foci in colon cancer epidemiology. Methods Mol. Biol. 2009, 472, 373–386. [Google Scholar] [PubMed]
- Fotiadis, C.I.; Stoidis, C.N.; Spyropoulos, B.G.; Zografos, E.D. Role of probiotics, prebiotics and synbiotics in chemoprevention for colorectal cancer. World J. Gastroenterol. 2008, 14, 6453–6457. [Google Scholar] [CrossRef] [PubMed]
- Femia, A.P.; Luceri, C.; Dolara, P.; Giannini, A.; Biggeri, A.; Salvadori, M.; Clune, Y.; Collins, K.J.; Paglierani, M.; Caderni, G. Antitumorigenic activity of the prebiotic inulin enriched with oligofructose in combination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis on azoxymethane-induced colon carcinogenesis in rats. Carcinogenesis 2002, 23, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Shukla, G. Administration of prebiotic inulin suppresses 1,2-dimethylhydrazine dihydrochloride induced procarcinogenic biomarkers fecal enzymes and preneoplastic lesions in early colon carcinogenesis in Sprague Dawley rats. J. Funct. Foods 2013, 5, 991–996. [Google Scholar] [CrossRef]
- Dias, M.C.; Vieiralves, N.F.; Gomes, M.I.; Salvadori, D.M.; Rodrigues, M.A.; Barbisan, L.F. Effects of lycopene, synbiotic and their association on early biomarkers of rat colon carcinogenesis. Food Chem. Toxicol. 2010, 48, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, M.V.; Appel, M.J.; Hollanders, V.M.; Woutersen, R.A. A comparison of the effects of dietary cellulose and fermentable galacto-oligosaccharide, in a rat model of colorectal carcinogenesis: Fermentable fibre confers greater protection than non-fermentable fibre in both high and low fat backgrounds. Carcinogenesis 1999, 20, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, M.V.; Schoterman, H.C.; Bruijntjes, J.B.; Hollanders, V.M.; Woutersen, R.A. Effect of dietary galacto-oligosaccharides on azoxymethane-induced aberrant crypt foci and colorectal cancer in Fischer 344 rats. Carcinogenesis 2001, 22, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Donini, L.M.; Serra-Majem, L.; Bullo, M.; Gil, A.; Salas-Salvado, J. The Mediterranean diet: Culture, health and science. Br. J. Nutr. 2015, 113, S1–S3. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the human large intestine: Its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [PubMed]
- Sivieri, K.; Spinardi-Barbisan, A.L.T.; Barbisan, L.F.; Bedani, R.; Pauly, N.D.; Carlos, I.Z.; Benzatti, F.; Vendramini, R.C.; Rossi, E.A. Probiotic Enterococcus faecium CRL 183 inhibit chemically induced colon cancer in male Wistar rats. Eur. Food Res. Technol. 2008, 228, 231–237. [Google Scholar] [CrossRef]
- Gallaher, D.D.; Khil, J. The effect of synbiotics on colon carcinogenesis in rats. J. Nutr. 1999, 129, 1483S–1487S. [Google Scholar] [PubMed]
- Reddy, B.S. Prevention of colon cancer by pre-and probiotics: Evidence from laboratory studies. Br. J. Nutr. 1998, 80, S219–S223. [Google Scholar] [PubMed]
- Aachary, A.A.; Gobinath, D.; Srinivasan, K.; Prapulla, S.G. Protective effect of xylooligosaccharides from corncob on 1,2-dimethylhydrazine induced colon cancer in rats. Biol. Carbohydr. Diet. Fiber 2015, 5, 146–152. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, H.; Lu, C.; Kang, Z.; Luo, Y.; Feng, L.; Lu, X. Fermented milk supplemented with probiotics and prebiotics can effectively alter the intestinal microbiota and immunity of host animals. J. Dairy Sci. 2012, 95, 4813–4822. [Google Scholar] [CrossRef] [PubMed]
- Weir, T.L.; Manter, D.K.; Sheflin, A.M.; Barnett, B.A.; Heuberger, A.L.; Ryan, E.P. Stool microbiome and metabolome differences between colorectal cancer patients and healthy adults. PLoS ONE 2013, 8, e70803. [Google Scholar] [CrossRef] [PubMed]
- Pool-Zobel, B.L.; Sauer, J. Overview of experimental data on reduction of colorectal cancer risk by inulin-type fructans. J. Nutr. 2007, 137, 2580S–2584S. [Google Scholar] [PubMed]
- Hijova, E.; Szabadosova, V.; Stofilova, J.; Hrckova, G. Chemopreventive and metabolic effects of inulin on colon cancer development. J. Vet. Sci. 2013, 14, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hijova, E.; Szabadosova, V.; Strojny, L.; Bomba, A. Changes chemopreventive markers in colorectal cancer development after inulin supplementation. Bratisl. Lek. Listy 2014, 115, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Scharlau, D.; Borowicki, A.; Habermann, N.; Hofmann, T.; Klenow, S.; Miene, C.; Munjal, U.; Stein, K.; Glei, M. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res. 2009, 682, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.S.; Sengupta, S.; Tjandra, J.J.; Gibson, P.R. The influence of specific luminal factors on the colonic epithelium: High-dose butyrate and physical changes suppress early carcinogenic events in rats. Dis. Colon Rectum. 2005, 48, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Goldin, B.R.; Gorbach, S.L. The relationship between diet and rat fecal bacterial enzymes implicated in colon cancer. J. Natl. Cancer Inst. 1976, 57, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Juśkiewicz, J.; Wróblewska, M.; Jarosławska, J.; Baliński, P.; Matusevičius, P.; Zduńczyk, P.; Zduńczyk, Z. Effects of inulin supplemented to cellulose-free or cellulose-rich diets on caecal environment and biochemical blood parameters in rats. J. Anim. Feed Sci. 2009, 18, 709–722. [Google Scholar] [CrossRef]
- Lahouar, L.; Pochart, P.; Salem, H.B.; El Felah, M.; Mokni, M.; Magne, F.; Mangin, I.; Suau, A.; Pereira, E.; Hammami, M.; Achour, L. Effect of dietary fibre of barley variety ‘Rihane’ on azoxymethane-induced aberrant crypt foci development and on colonic microbiota diversity in rats. Br. J. Nutr. 2012, 108, 2034–2042. [Google Scholar] [CrossRef] [PubMed]
- Suau, A.; Bonnet, R.; Sutren, M.; Godon, J.J.; Gibson, G.R.; Collins, M.D.; Dore, J. Direct analysis of genes encoding 16S rRNA from complex communities reveals many novel molecular species within the human gut. Appl. Environ. Microbiol. 1999, 65, 4799–4807. [Google Scholar] [PubMed]
- Manning, T.S.; Gibson, G.R. Microbial-gut interactions in health and disease. Prebiotics. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, R.W.; George, S.E.; Claxton, L.D. Role of the gastrointestinal mucosa and microflora in the bioactivation of dietary and environmental mutagens or carcinogens. Drug. Metab. Rev. 1992, 24, 425–492. [Google Scholar] [CrossRef] [PubMed]
- Tzortzis, G.; Goulas, A.K.; Gibson, G.R. Synthesis of prebiotic galactooligosaccharides using whole cells of a novel strain, Bifidobacterium bifidum NCIMB 41171. Appl. Microbiol. Biotechnol. 2005, 68, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Tzortzis, G.; Goulas, A.K.; Gee, J.M.; Gibson, G.R. A novel galactooligosaccharide mixture increases the bifidobacterial population numbers in a continuous in vitro fermentation system and in the proximal colonic contents of pigs in vivo. J. Nutr. 2005, 135, 1726–1731. [Google Scholar] [PubMed]
- Reeves, P.G.; Rossow, K.L.; Lindlauf, J. Development and testing of the AIN-93 purified diets for rodents: Results on growth, kidney calcification and bone mineralization in rats and mice. J. Nutr. 1993, 123, 1923–1931. [Google Scholar] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Spinardi-Barbisan, A.L.; Rodrigues, M.A.; de Camargo, J.L.; Teran, E.; Barbisan, L.F. Lack of chemopreventive effects of ginger on colon carcinogenesis induced by 1,2-dimethylhydrazine in rats. Food Chem. Toxicol. 2006, 44, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Nguyen, T.H.; Nguyen, T.T.; Maischberger, T.; Haltrich, D. β-Galactosidase from Lactobacillus plantarum WCFS1: Biochemical characterization and formation of prebiotic galacto-oligosaccharides. Carbohydr. Res. 2010, 345, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Splechtna, B.; Nguyen, T.-H.; Steinböck, M.; Kulbe, K.D.; Lorenz, W.; Haltrich, D. Production of prebiotic galacto-oligosaccharides from lactose using β-galactosidases from Lactobacillus reuteri. J. Agric. Food Chem. 2006, 54, 4999–5006. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett. 1995, 93, 55–71. [Google Scholar] [CrossRef]
- Qamar, T.R.; Syed, F.; Nasir, M.; Rehman, H.; Zahid, M.N.; Liu, R.H.; Iqbal, S. Novel combination of prebiotics galacto-oligosaccharides and inulin-inhibited aberrant crypt foci formation and biomarkers of colon cancer in wistar rats. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Nyman, M.; Åke Jönsson, J. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef] [PubMed]
- De Wiele, T.V.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Prebiotic effects of chicory inulin in the simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2004, 51, 143–153. [Google Scholar] [CrossRef] [PubMed]
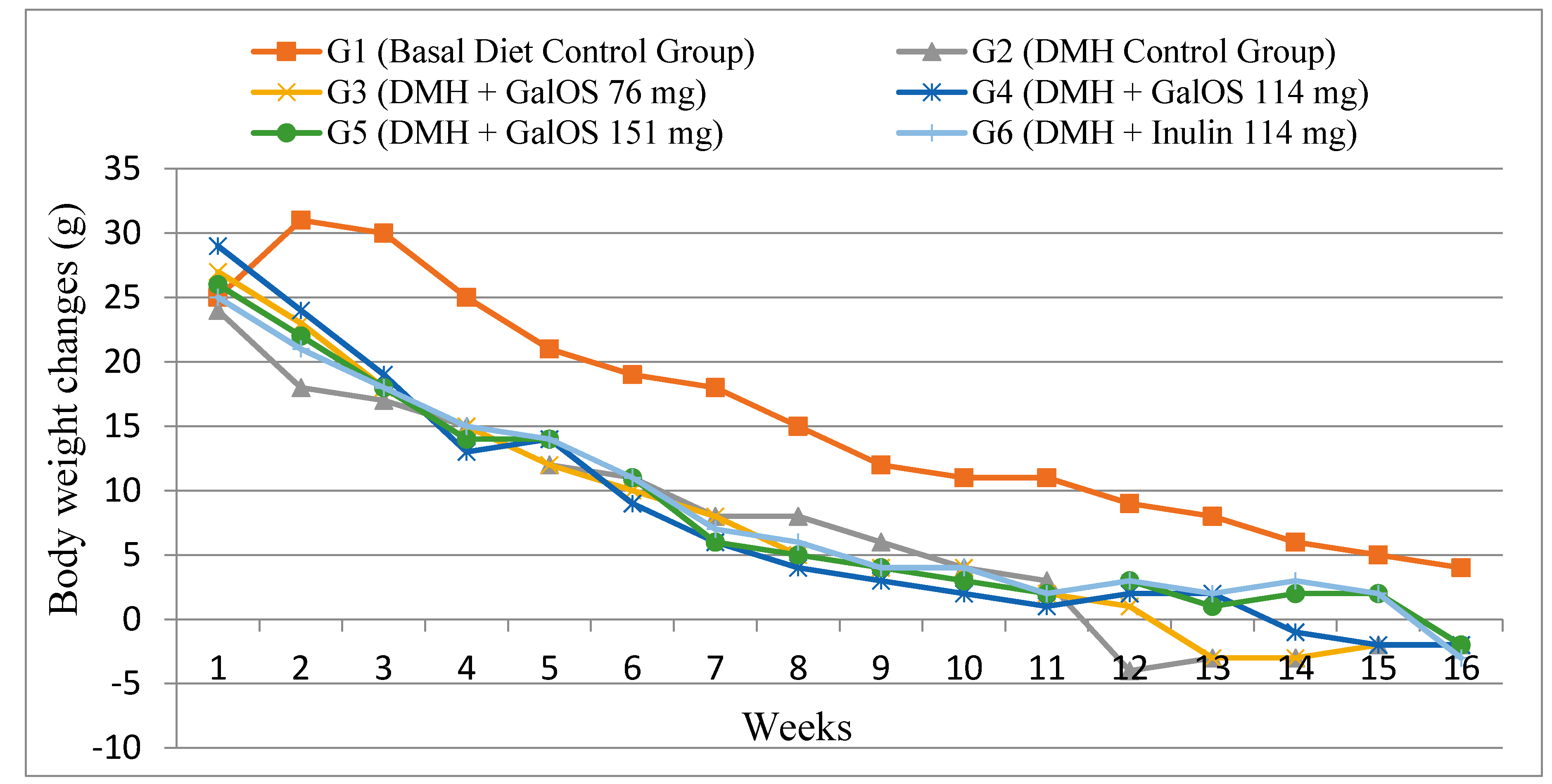


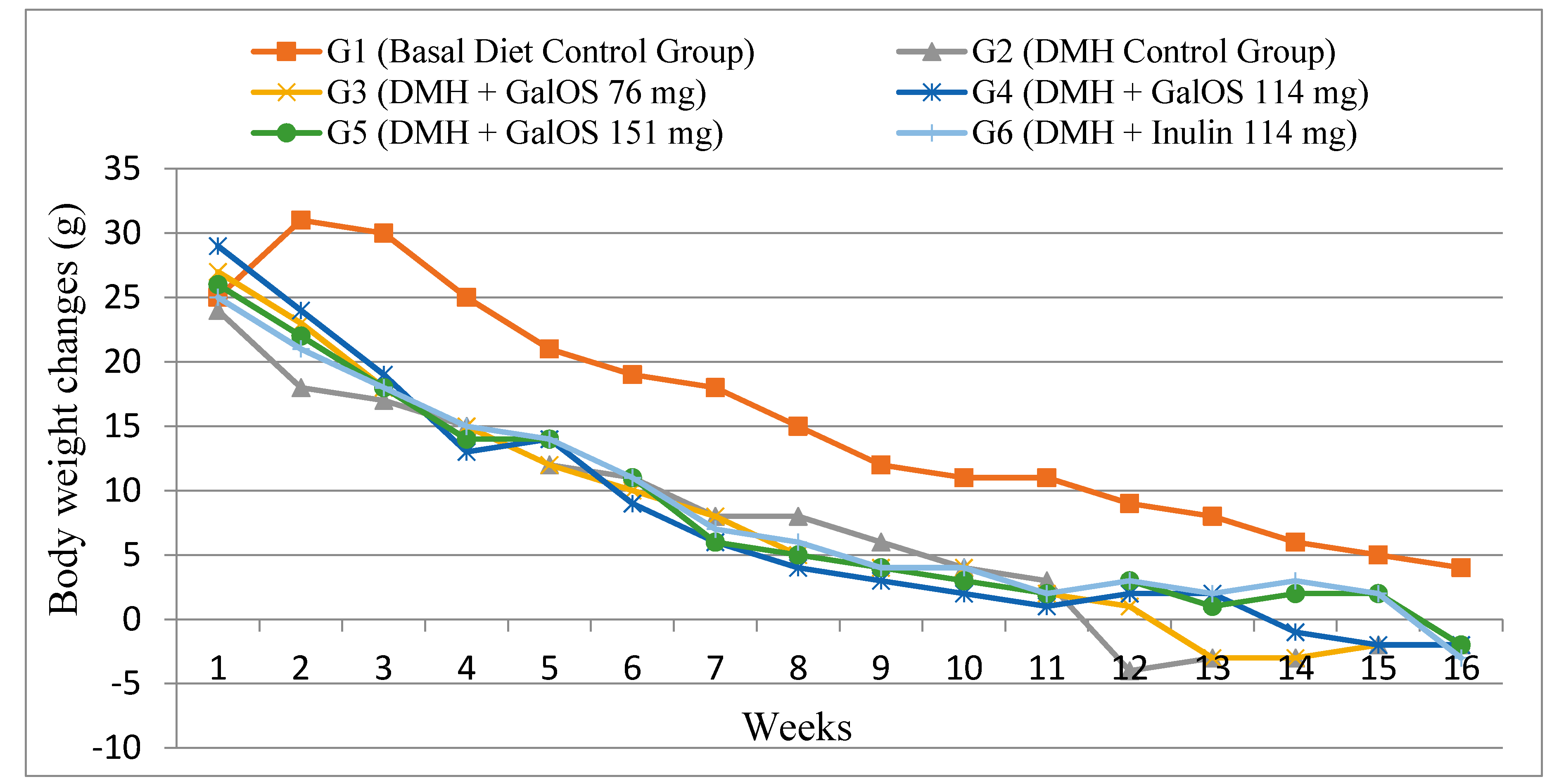{kind=link}
{kind=link}
| ACF | G1 (Basal Diet Control Group) | G2 (DMH Control Group) | G3 (DMH + GalOS 76 mg) | G4 (DMH + GalOS 114 mg) | G5 (DMH + GalOS 151 mg) | G6 (DMH + Inulin 114 mg) |
|---|---|---|---|---|---|---|
| ACF/proximal colon | ND | 27.4 ± 2.10 a | 24.7 ± 3.11 a,b | 18.7 ± 2.56 b,c | 14.3 ± 1.78 c | 16.4 ± 2.83 c |
| ACF/middle colon | ND | 44.7 ± 2.98 a | 42.3 ± 2.86 a,b | 34.3 ± 3.8 b,c | 22.9 ± 3.45 d | 28.3 ± 3.23 c,d |
| ACF/distal colon | ND | 119.1 ± 5.56 a | 115.3 ± 7.22 a,b | 110.4 ± 4.63 a,b | 101.7 ± 3.32 b,c | 90.3 ± 3.31 c |
| Total ACF/colon | ND | 191.2 ± 6.46 a | 182.4 ± 7.92 a | 163.4 ± 4.72 b | 139.0 ± 3.24 c | 135.1 ± 3.73 c |
| Percent inhibition of total ACF | - | - | 4.6 | 14.53 | 27.3 | 29.3 |
| Parameters | G1 (Basal Diet Control Group) | G2 (DMH Control Group) | G3 (DMH + GalOS 76 mg) | G4 (DMH + GalOS 114 mg) | G5 (DMH + GalOS 151 mg) | G6 (DMH + Inulin 114 mg) |
|---|---|---|---|---|---|---|
| Cecum | ||||||
| Acetate | 80.8 ± 3.41 c | 79.2 ± 3.28 c | 82.8 ± 3.59 b,c | 86.2 ± 3.85 a,b,c | 92.4 ± 3.49 a,b | 94.9 ± 3.83 a |
| Propionate | 21.9 ± 2.03 a,b | 20.6 ± 1.85 b | 22.1 ± 2.11 a,b | 23.4 ± 1.78 a,b | 26.1 ± 1.54 a,b | 26.6 ± 1.44 a |
| Butyrate | 14.1 ± 1.11 c | 13.9 ± 1.03 c | 14.8 ± 0.93 b,c | 16.4 ± 1.21 a,b,c | 18.5 ± 1.36 a | 17.8 ± 1.09 a,b |
| Fecal | ||||||
| Acetate | 51.7 ± 2.58 b | 50.6 ± 2.71 b | 53.2 ± 3.11 b | 56.8 ± 2.35 a,b | 62.1 ± 3.13 a | 64.8 ± 2.67 a |
| Propionate | 14.3 ± 0.99 c,d | 14.1 ± 0.79 d | 15.5 ± 0.68 c,d | 17.4 ± 1.05 b,c | 20.3 ± 1.26 a,b | 21.2 ± 1.34 a |
| Butyrate | 5.2 ± 0.51 b | 4.92 ± 0.48 b | 5.5 ± 0.53 b | 6.1 ± 0.53 a,b | 7.3 ± 0.57 a | 7.5 ± 0.67 a |
| Enzymes | G1 (Basal Diet Control Group) | G2 (DMH Control Group) | G3 (DMH + GalOS 76 mg) | G4 (DMH + GalOS 114 mg) | G5 (DMH + GalOS 151 mg) | G6 (DMH + Inulin 114 mg) |
|---|---|---|---|---|---|---|
| Cecum | ||||||
| β-Glucosidase | 0.82 ± 0.11 a,b | 0.96 ± 0.11 a | 0.94 ± 0.09 a | 0.80 ± 0.10 a,b | 0.77 ± 0.11 a,b | 0.59 ± 0.09 b |
| β-Glucoronidase | 3.43 ± 0.18 a,b | 3.52 ± 0.19 a | 3.25 ± 0.12 a,b | 3.12 ± 0.18 a,b | 2.96 ± 0.15 b | 2.99 ± 0.19 a,b |
| Nitroreductase | 4.38 ± 0.29 a | 4.47 ± 0.28 a | 4.27 ± 0.25 a | 3.83 ± 0.25 a,b | 3.44 ± 0.19 b | 3.26 ± 0.17 b |
| Azoreductase | 11.67 ± 1.01 a | 11.42 ± 0.56 a,b | 10.08 ± 0.67 a,b | 10.17 ± 0.81 a,b | 9.16 ± 0.79 b | 9.25 ± 0.72 a,b |
| Fecal | ||||||
| β-Glucosidase | 0.72 ± 0.11 a,b | 0.91 ± 0.10 a | 0.76 ± 0.08 a,b | 0.69 ± 0.09 a,b | 0.64 ± 0.11 a,b | 0.53 ± 0.09 b |
| β-Glucoronidase | 2.97 ± 0.25 a | 3.07 ± 0.16 a | 2.93 ± 0.10 a,b | 2.72 ± 0.14 a,b | 2.43 ± 0.15 b | 2.55 ± 0.16 a,b |
| Nitroreductase | 3.13 ± 0.18 a | 3.18 ± 0.19 a | 2.86 ± 0.15 a,b | 2.76 ± 0.15 a,b | 2.49 ± 0.17 b | 2.55 ± 0.18 b |
| Azoreductase | 8.17 ± 0.51 a,b | 8.42 ± 0.42 a | 7.92 ± 0.47 a,b | 7.41 ± 0.59 a,b | 6.91 ± 0.45 a,b | 6.67 ± 0.57 b |
| Microbes | G1 (Basal Diet Control Group) | G2 (DMH Control Group) | G3 (DMH + GalOS 76 mg) | G4 (DMH + GalOS 114 mg) | G5 (DMH + GalOS 151 mg) | G6 (DMH + Inulin 114 mg) |
|---|---|---|---|---|---|---|
| Lactobacillus | 7.18 ± 0.24 b | 7.14 ± 0.17 b | 7.39 ± 0.18 b | 7.64 ± 0.09 a,b | 7.93 ± 0.16 a | 7.52 ± 0.18 a,b |
| Bifidobacteria | 8.11 ± 0.14 c,d | 8.07 ± 0.15 d | 8.18 ± 0.14 c,d | 8.24 ± 0.13 b,c,d | 8.51 ± 0.15 b,c | 9.12 ± 0.11 a |
| Clostridia | 8.82 ± 0.12 a,b | 8.92 ± 0.11 a | 8.77 ± 0.15 a,b | 8.59 ± 0.20 a,b,c | 8.51 ± 0.14 a,b,c | 8.45 ± 0.11 b,c |
| Enterococci | 7.42 ± 0.11 | 7.48 ± 0.10 | 7.53 ± 0.18 | 7.37 ± 0.17 | 7.22 ± 0.15 | 7.27 ± 0.09 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qamar, T.R.; Iqbal, S.; Syed, F.; Nasir, M.; Rehman, H.; Iqbal, M.A.; Liu, R.H. Impact of Novel Prebiotic Galacto-Oligosaccharides on Various Biomarkers of Colorectal Cancer in Wister Rats. Int. J. Mol. Sci. 2017, 18, 1785. https://doi.org/10.3390/ijms18091785
Qamar TR, Iqbal S, Syed F, Nasir M, Rehman H, Iqbal MA, Liu RH. Impact of Novel Prebiotic Galacto-Oligosaccharides on Various Biomarkers of Colorectal Cancer in Wister Rats. International Journal of Molecular Sciences. 2017; 18(9):1785. https://doi.org/10.3390/ijms18091785
Chicago/Turabian StyleQamar, Tahir Rasool, Sanaullah Iqbal, Fatima Syed, Muhammad Nasir, Habib Rehman, Muhammad Aamir Iqbal, and Rui Hai Liu. 2017. "Impact of Novel Prebiotic Galacto-Oligosaccharides on Various Biomarkers of Colorectal Cancer in Wister Rats" International Journal of Molecular Sciences 18, no. 9: 1785. https://doi.org/10.3390/ijms18091785