Genetic Diversity and Genetic Relationships of Purple Willow (Salix purpurea L.) from Natural Locations


1
Department of Plant Breeding and Seed Production, University of Warmia and Mazury in Olsztyn, 10-724 Olsztyn, Poland
2
Institute for Systematic Botany, Friedrich-Schiller-Universität Jena, 07743 Jena, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(1), 105; https://doi.org/10.3390/ijms19010105
Submission received: 9 November 2017
/
Revised: 12 December 2017
/
Accepted: 27 December 2017
/
Published: 30 December 2017
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:In this study, the genetic diversity and structure of 13 natural locations of Salix purpurea were determined with the use of AFLP (amplified length polymorphism), RAPD (randomly amplified polymorphic DNA) and ISSR (inter-simple sequence repeats). The genetic relationships between 91 examined S. purpurea genotypes were evaluated by analyses of molecular variance (AMOVA), principal coordinates analyses (PCoA) and UPGMA (unweighted pair group method with arithmetic mean) dendrograms for both single marker types and a combination of all marker systems. The locations were assigned to distinct regions and the analysis of AMOVA (analysis of molecular variance) revealed a high genetic diversity within locations. The genetic diversity between both regions and locations was relatively low, but typical for many woody plant species. The results noted for the analyzed marker types were generally comparable with few differences in the genetic relationships among S. purpurea locations. A combination of several marker systems could thus be ideally suited to understand genetic diversity patterns of the species. This study makes the first attempt to broaden our knowledge of the genetic parameters of the purple willow (S. purpurea) from natural location for research and several applications, inter alia breeding purposes.
Keywords:
Salix purpurea L.; genetic diversity; genetic relationships; molecular markers; AFLP; RAPD; ISSR; breeding
1. Introduction
Recent years have witnessed the growing popularity of natural remedies, which increased the demand for high-quality herbal material [1,2]. Willow bark is commonly used in the production of natural alternatives to aspirin because of similar medicinal properties to acetylsalicylic acid (synthetic aspirin): it delivers analgesic, antipyretic, anti-inflammatory and anti-rheumatic effects [3,4,5], and it is used as a cold remedy and an alternative treatment for rheumatic diseases [6,7,8,9].
The medicinal properties of willow bark can be attributed mainly to salicylic glycosides (SGs) whose content and composition vary significantly in bark tissue among genotypes [10]. Considerable differences in the content of pharmacologically active ingredients are observed both between and within species of the genus Salix [11,12]. The genus Salix comprises more than 400 identified species in highly diverse natural habitats [13,14], which points to an abundance of natural resources for the production of herbal therapies. The most stable and safe source of high-quality herbal material are identified or bred varieties, which can be grown under field conditions [15]. Highly suitable Salix spp. genotypes with a high content of SGs can be found in natural habitats and included in breeding programs as parental genotypes to obtain as high as possible effect of transgression. Purple willow (Salix purpurea L.) is characterized by one of the highest SG concentrations in the genus Salix. According to the literature, the SG content of purple willow bark ranges from 3% to 11% [10,11,12,16]. Such high variations in the SG content of S. purpurea L. justify breeding efforts to obtain varieties that are most suited for medicinal use.
Breeding of new plant varieties is a long and laborious process that does not always lead to anticipated results. The selection of the most appropriate starting breeding material is thus a key determinant of breeding success. Starting materials for breeding should be selected based on a detailed analysis of genetic diversity of plant material in natural localities to provide potential sources of parental forms. The selection of parental genotypes should guarantee that the offspring will be characterized by considerable variations in value of trait. For this purpose, various S. purpurea genotypes should be collected and subjected to genetic diversity analysis to select the optimal starting parental forms. This goal can be accomplished with the use of DNA markers, which are the main diagnostic tools in modern plant breeding. DNA markers are valuable tools for S. purpurea studies, inter alia to identify species of Salix [17,18,19], genotypes of S. purpurea [20,21] and their genetic diversity [20,22,23]. They facilitate and support structural analyses of genomes, genetic mapping, identification of quantitative trait loci (QTL), marker-assisted selection (MAS) and sequencing of S. purpurea [24,25,26]. Nowadays, numerous marker systems are available and are generating from several hundred (RAPDs, ISSRs) to thousands of markers (DArTs—diversity arrays technology markers, SNPs—single nucleotide polymorphism markers). The optimal marker system, depending on the goal of its application, should be selected based on its efficiency as well as the time and cost of the generation method. The recently observed significant progress in research on the Salix purpurea genome, including the public availability of genomic sequence data [24,26], has made purple willow a model species in research on the genetic improvement of willows. This fact also allows the use of very efficient markers in these studies. The older marker systems begin to give way to methods based on the whole genomes sequencing. However, anonymous and neutral well-established marker systems such as AFLP (amplified length polymorphism), RAPD (randomly amplified polymorphic DNA) and ISSR (inter-simple sequence repeats) are suitable to characterize the basic genetic diversity of plant populations, including starting materials for breeding [27,28,29].
The search for valuable breeding material of S. purpurea can begin in natural localities because the purple willow is a common species in many countries [14,30,31]. Purple willow is native to Europe and Asia, but plants are also often anthropogenically introduced in large areas (inter alia North America and Europe) primarily to reduce erosion along stream banks and lake shores, for cultivation on arable land and for use in basketry [13,14,32]. The species is dioecious and outcrossing. The flowers are pollinated by wind and insects, and blooming starts before leaf development (March–April). Seeds are tiny capsules which are dispersed by wind. Three subspecies have been identified, and hybridization between S. purpurea and several other Salix species is common [13,14,30,31]. In our study area, some species such as S. viminalis, S. triandra, S. caprea and S. fragilis co-occur with S. purpurea, and hybridization among species may exist.
2. Results and Discussion
2.1. Efficiency of the Used Marker Systems
The efficiency of the applied marker systems was evaluated based on data matrices for each marker system and a combination of all systems. This approach was adopted to compare the results in view of their economic and scientific applicability for breeding S. purpurea. The genetic diversity of S. purpurea was analyzed with two combinations of AFLP primers, 62 RAPD primers and 20 ISSR primers, which generated 159, 574 and 221 products, respectively (Table 1). The AFLP procedure is complex, but a relatively high number of polymorphic and reproducible characters can be expected per analysis after restriction, ligation, preamplification and selective amplification. Although lower reproducibility was assumed for RAPD marker system, their significance can be increased by applying the relevant procedures [36]. We therefore repeated the genotyping by RAPDs and ISSRs as well for both biological replicates of every sample, and ambiguous products were excluded from analysis. RAPDs and ISSRs revealed a lower number of scored products per PCR, but these methods are very simple based on only one PCR. Thus, the number of PCRs can be increased with comparatively little effort. In fact, the RAPD primers yielded around nine times fewer products per primer (9.3 products) than one combination of AFLP primers, whereas one ISSR primer produced a higher number of scored products (11.1) than RAPDs, but its yield was seven times lower than that of AFLPs (Table 1). In our study, an average of 711.7 polymorphic and 3.6 private products were identified for the analyzed locations (Table 2). Based on these two parameters, we observed minor differences in the efficiency of the used marker systems. Among all identified AFLP products, an average of 71% polymorphic products were scored for the tested locations, which was somewhat below the values noted for RAPDs and ISSRs (75%). In contrast, the average percentage of private products (8%) was higher for AFLPs than for ISSRs (6%) and RAPDs (4%). The observed differences in technical effort and yield, but also in expected significance of results indicate for the reasonable combination of several marker systems to provide the best approach for elucidating genetic diversity and genetic relationships of S. purpurea locations.
The percentage of polymorphic amplification products (%p), unbiased expected heterozygosities (uHe) and the values of Shannon diversity indices (I) were higher for RAPDs and ISSRs than for AFLPs, and ISSRs which generated higher values of average genetic differentiation between locations (ΦST) than other methods (Table 1). The marker systems associated with highly polymorphic microsatellite regions such as ISSR are expected to reveal higher levels of genetic diversity [37] than RAPD and AFLP polymorphisms which are randomly distributed in the genome [38]. However, AFLPs revealed the highest scored products ratio, which usually lead to the highest efficiency of polymorphisms detection and highest discriminative power. Diversity levels of applied marker systems are dependent from variability of the target regions for primers and/or restriction enzymes, and they differ among species requiring a careful selection of the applied RAPD primer and AFLP primer combinations [37,39]. Despite some differences, our combined application of the three marker systems with different diversity natures could support the identification of complex diversity patterns within and among locations and regions, and it could be suitable and applicable i.e., in molecular breeding issues of S. purpurea. The overall applicability of various types of molecular markers in determinations of species identity, genetic diversity, taxonomic and phylogenetic analyses of the genus Salix has been described by several authors [17,18,19,21,40,41,42,43,44,45,46,47,48]. For example, Alsos et al. [49] demonstrated that AFLP marker system were highly effective in allocating 41 natural locations of S. herbacea to five groups corresponding to their geographic regions. Van Puyvelde and Triest [41] used ISSR marker system to evaluate the spatial isolation of S. alba at the level of individual plants as well as populations. The applicability of natural willow populations in breeding are supported by the studies of Trybush et al. [45] and Berlin et al. [50]. Trybush et al. [45] reported a high level of genetic diversity in 84 S. viminalis genotypes from natural localities, which validated the choice of breeding materials from the group of the examined genotypes. The cited authors used 38 SSR primers generating an average of seven alleles each. Interesting results were also reported by Berlin et al. [50] who relied on 38 SSR loci to analyze the genetic diversity of S. viminalis in natural habitats in Great Britain and Sweden, and concluded for high breeding potential due to the observed high genetic diversity. However, genetic diversity was lower in Sweden and many genotypes were genetically identical; therefore, the observed results were attributed to artificial introduction of the species to Sweden [50]. With regard to S. purpurea, our study confirms the high usefulness of AFLPs, RAPDs and ISSRs in genetic diversity analyses of natural locations [20,22,23,43].
2.2. Genetic Diversity and Analysis of Molecular Variance (AMOVA)
In our study, the genetic diversity of S. purpurea (uHe = 0.179; Table 1) was high despite lower than reported by other authors, but those analyses either relied on different marker systems [43] or did not include natural locations of S. purpurea [20,22,23]. Genetic differentiation (ΦST = 0.212; Table 1) was also slightly lower than expected for plant species with comparable life-history traits (long-lived perennials, outbreeding, wind dispersal: ΦST ≈ 0.25) [51]. Natural occurrence of S. purpurea was usually irregular, and the number of individuals in the sampled locations ranged from several to more than a dozen with a corresponding impact on genetic diversity and structure. Low abundance of S. purpurea is primarily related to the propagation of the species. Although many seeds are produced, establishment of plants is low due to low seed germination ability and viability. In addition, S. purpurea often form interspecific hybrids which additionally decrease the occurrence of homogeneous species in natural populations. In the analyzed locations, the genetic diversity of S. purpurea was similar when calculated with the use of single marker system (uHe = 0.164–0.187) and a combination of all systems (uHe = 0.179; Table 3). Genetic diversity in OL3 (Olsztyn Lakeland) was significantly lower than in the remaining locations (I = 0.134; uHe = 0.110; Table 3), but only three genotypes were analyzed. In most cases, natural locations from the same regions were more similar within than among regions (Table 4). However, higher genetic similarity among than within regions was also observed for locations of Olsztyn Lakeland (OL).
The components of genetic diversity of S. purpurea were determined by AMOVA. In a combined analysis of all marker types, AMOVA revealed 79% genetic variation within locations, 11% between locations and 10% between regions (Table 5). The distribution of genetic variation was similar for single marker systems (77–84% genetic variation within locations; data not shown). The results of this study confirm the general observation that most tree and shrub species are characterized by high genetic variations within natural populations [52]. In the work of Alsos et al. [49], genetic variations within S. herbacea populations ranged from 50% to 80%, depending on the geographic identity of the analyzed natural locations. Compared to our results, similar levels of genetic variation were observed for S. viminalis within natural locations (>90%) [42,48].
2.3. Genetic Relationships
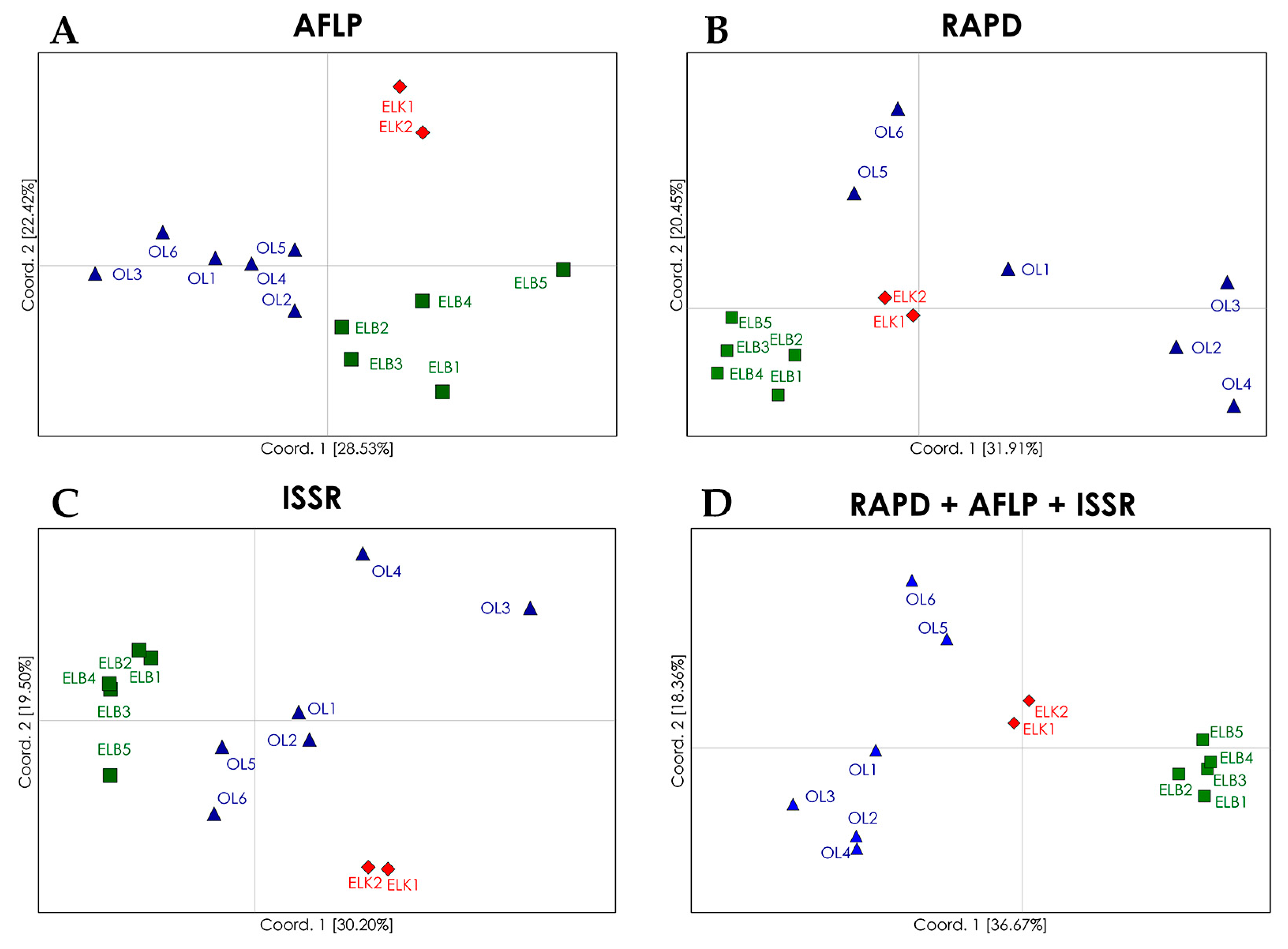
Despite low genetic variations among regions (10%; Table 5), PCoA (principal coordinates analysis) plots and dendrograms based on genetic differentiation (ΦST) values revealed well defined clusters of S. purpurea locations from particular regions (Figure 1). The analyzed locations were grouped in four clusters with the use of RAPD marker system. The locations from the Ełk Lakeland (ELK) and Żuławy Wiślane (ELB) were grouped into two distinct regional clusters. The locations from region OL were separated from the clusters representing regions ELK and ELB, but they formed a non-homogeneous group of two clusters, including Cluster 3 with locations OL2, OL3 and OL4, and Cluster 4 with locations OL5 and OL6. Location OL1 was not assigned to any cluster. The allocation of genotypes from regions ELK and ELB to two separate clusters reflects their geographical distributions as both regions are separated by a considerable distance (more than 200 km): The results for region OL are surprising, because the analyzed locations are situated in relative proximity and could be expected to form a single group. In fact, this assumption is confirmed by a PCoA based on AFLP marker system, which revealed three groups corresponding to the studied regions, whereas regions OL and ELB were less distinct than region ELK. In comparison with RAPD marker system, AFLPs revealed a higher genetic similarity among locations from region OL, and lower genetic similarity among locations from region ELB. ISSR marker system showed the lowest fit between the analyzed locations and regions. Similar to AFLP and RAPD marker systems, ISSRs revealed a separate cluster of locations from region ELK. In the PCoA plot, one location from region ELB (ELB5) was clearly distinct from the remaining locations. The lowest level of genetic similarity was observed among locations from region OL, and the locations OL3 and OL4 were significantly distant from remaining locations. In contrast, in PCoA based on RAPD marker system, locations OL3 and OL4 were assigned to the same cluster. The examined locations were clearly allocated to regional groups in a combined analysis of results for the three marker systems. The locations from regions ELK and ELB formed two autonomous clusters, and locations from region OL formed two clusters, but these differences were justified. Locations OL5 and OL6 formed a separate cluster and were geographically more distant to the remaining OL locations. Despite certain differences in single marker systems, the PCoA for the combination of marker systems can be successfully used to discriminate natural locations of S. purpurea. The differences produced by RAPDs, AFLPs and ISSRs in this study may thus be attributed to their distribution in the genome, and thus, representation of genetic variation. The analysis based on AFLP marker system produced the best fit between the examined locations and their regions of origin, which was also reflected in the combined approach. Alsos et al. [49] also demonstrated that AFLP marker system were highly effective in allocating 41 natural locations of S. herbacea to five groups corresponding to their geographic regions. However, Van Puyvelde and Triest [41] relied on ISSR markers to evaluate the spatial isolation of S. alba at the level of individual plants as well as populations.
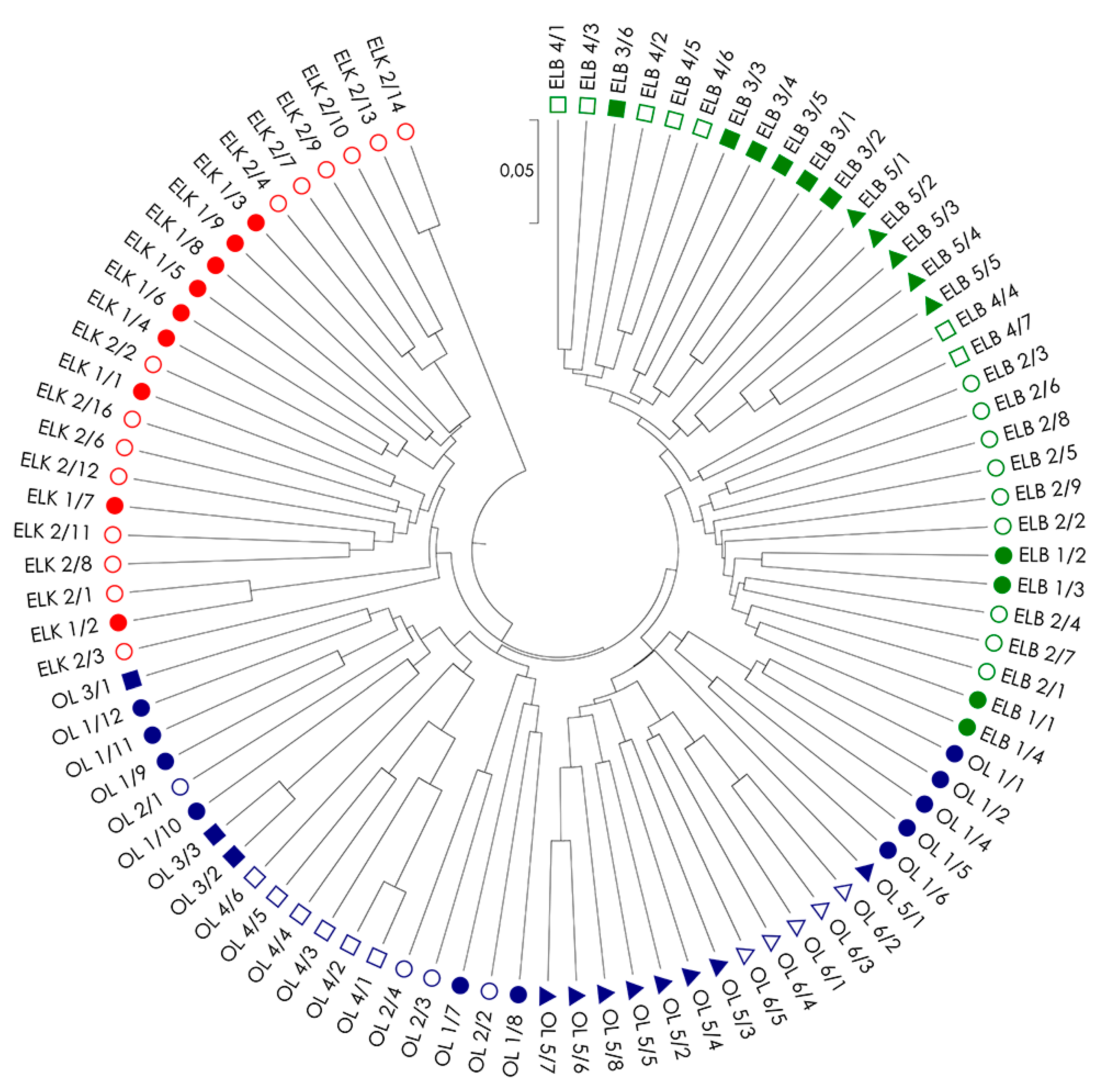
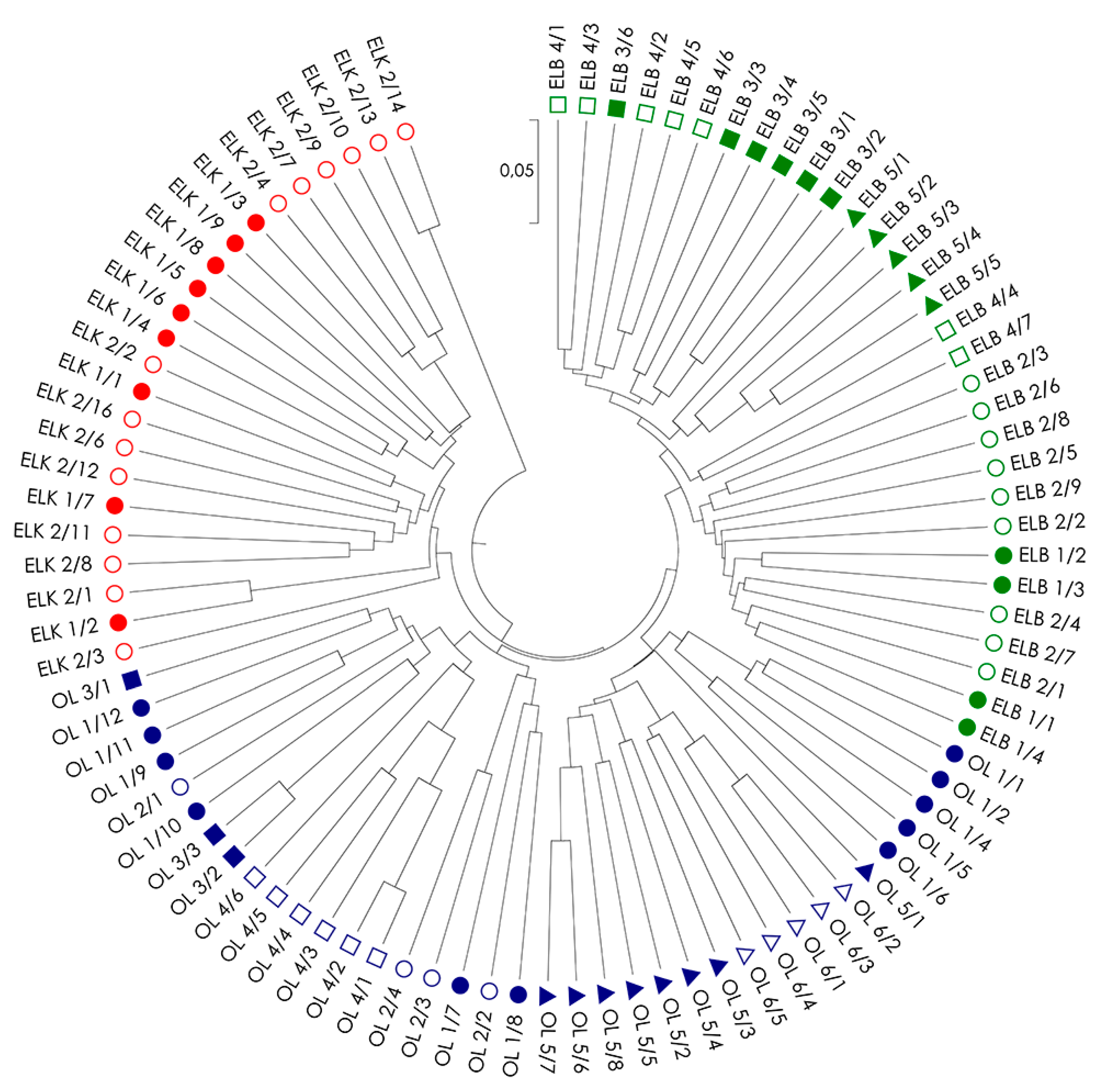
Genetic distances among all examined S. purpurea genotypes (DS; Table S1) were used to create UPGMA dendrograms for single marker systems (Figure S1) and a combination of systems (Figure 2) to illustrate the relationships among the individuals within and among locations and regions. Similar to PCoA, dendrograms also confirms the best fit between genotypes from the same regions in analysis based on a combination of all three marker types (Figure 2). The dendrogram revealed five clusters, including two clusters grouping genotypes from region ELK (with one small cluster of the two genotypes ELK 2/13 and ELK 2/14), one cluster from region ELB, and two clusters formed by genotypes from region OL. The clear separation of genotypes ELK 2/13 and ELK 2/14 from the remaining genotypes in region ELK resulted from a very low genetic distance between these genotypes (DS = 0.082) in contrast to high genetic distances among these genotypes to others in the region ELK (DS = 0.274–0.367), and among the studied genotypes in general (Table S1). The above suggests the presence of a strong kinship relations among the two genotypes. The presence of specific clusters and missing association of genotypes to their location of origin (with the exception of location OL4) could be explained by strong, but random gene flow and fragmented occurrence of the species, which often results from random establishment of the progeny.
3. Materials and Methods
3.1. Plant Material and DNA Isolation
Young leaves growing on shoots were sampled from 91 genotypes of S. purpurea in 13 natural locations in north-eastern Poland (Table 6) [10,53]. For our analyses, plant material was sampled only from exactly defined S. purpurea plants in accordance with the botanical key prepared and based on scientific literature [13,28,29,54]. The number of genotypes in every analyzed locations is related to the actual number of S. purpurea plants found in these locations, excluding identical genotypes (clones) identified by Sulima et al. [10]. The analyzed locations were situated in three geographic regions: Ełk Lakeland (ELK, two localities), Żuławy Wiślane—Delta of the Vistula River (ELB, five localities) and Olsztyn Lakeland (OL, six localities). DNA was isolated twice (in two biological replications) by the method proposed by Milligan with certain modifications [20,55]. The quantity and quality of DNA were evaluated with the NanoDrop 2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA) and confirmed by electrophoresis.
3.2. AFLP Marker System
AFLP amplification products (Figure S2) were generated in accordance with the procedure described by Vos et al. [33] with certain modifications. Genomic DNA (200 ng) was digested with EcoRI and MseI restriction enzymes (Invitrogen, Carlsbad, CA, USA), and double stranded EcoRI+/− and MseI+/− adapters were ligated to restriction fragments. Restriction and ligation were carried out overnight at room temperature, and the obtained mixture was diluted (1:5). Preamplification was carried out using 1× Reaction buffer B (0.8 M Tris HCl pH 9.0, 0.2 M (NH4)2SO4, 0.2% w/v Tween-20; Solis BioDyne, Tartu, Estonia), 2.5 mM MgCl2, 0.2 μM dNTP, 0.2 μM of the preselective primers MseI + C or G and EcoRI + A, 0.3 U FIREPol® DNA Polymerase (Solis BioDyne, Estonia) in 20 cycles (94 °C for 10 s, 56 °C for 30 s, 72 °C for 2 min), with a preliminary step (72 °C for 2 min) and a final step (60 °C for 30 min). In our study, 12 combinations of selective AFLP primers (Table S2) were tested on 16 randomly selected genotypes, and two of them were selected for the study generating a high number of polymorphic, repeatable and unambiguous products. The final amplification mixture was similar to the preamplification mixture, and it contained 3 μL of diluted preamplification product (1:20), 0.2 μM of MseI + GAA/CTT and 0.1 μM of EcoRI + ACA/AAC, respectively. Amplification was carried out according to the touchdown PCR protocol in the following steps: 95 °C for 15 min, 10 cycles with an annealing ramp of 1.0 °C per cycle (94 °C for 10 s, 65–56 °C for 30 s, 72 °C for 2 min), followed by 24 cycles with annealing temperature of 56 °C, and a final polymerization at 60 °C for 30 min.
The obtained AFLP products were separated by capillary electrophoresis with the use of the ABI Prism 3100 genetic analyzer (Applied Biosystems, Foster City, CA, USA) and internal size standard GS 500 ROX™ (Applied Biosystems, Foster City, CA, USA). AFLP fragments were scored using the software Genotyper 3.7 (Applied Biosystems, Foster City, CA, USA).
3.3. RAPD Marker System
The total volume of PCR-RAPD reaction mixtures volume was 25 μL containing 10 ng of DNA, 1× DreamTaq™ Green Buffer (Fermentas Thermo Scientific, Waltham, MA, USA), 0.2 mM of each dNTP (Sigma Aldrich Srl, Milan, Italy), 0.4 μM of the primer and 0.625 U of DreamTaq™ Green DNA Polymerase (Fermentas Thermo Scientific, Waltham, MA, USA). The PCR reaction was performed in 37 cycles (94 °C for 30 s, 40 °C for 2 min, 72 °C for 2 min) with an initial denaturation (94 °C for 10 min) and a final elongation (72 °C for 8 min). A total of 64 RAPD primers with 10 nucleotides each (Operon Biotechnologies GmbH, Cologne, Germany) were tested, and 62 primers were used in genetic diversity analyses of S. purpurea generating clear and repeatable bands (Table S3). All samples were genotyped twice for both biological replications and every RAPD locus was proofed to ensure the repeatability of the results.
3.4. ISSR Marker System
The PCR-ISSR reaction mix (25 μL) contained 10 ng of DNA, 1× DreamTaq™ Green Buffer (Fermentas Thermo Scientific, Waltham, MA, USA), 0.2 mM of each dNTP (Sigma Aldrich Srl, Milan, Italy), 0.4 μM of the primer and 0.625 U of DNA polymerase (Fermentas Thermo Scientific, Waltham, MA, USA). The reaction was carried out in 37 cycles (94 °C for 1 min, 42–63 °C for 2 min, depending on the primer, 72 °C for 2 min) with an initial denaturation (94 °C for 1 min) and a final elongation (72 °C for 8 min). ISSR primers were selected based on literature data [25,56,57]. The annealing temperature for every primer was determined in PCR-ISSR test reactions based on a temperature gradient (Table S4). A total of 20 ISSR primers generating clear and repeatable bands were selected for the analysis. All samples were genotyped twice for both biological replications, and every ISSR locus was proofed to ensure the repeatability of the results.
The amplification products of RAPD and ISSR reactions (Figures S3 and S4) were separated on 1.5% agarose gels with TBE buffer, stained with ethidium bromide (Sigma Aldrich Chemie GmbH, Steinheim, Germany) and visualized under UV light in the DIGIDOC gel imaging system (Biogenet, Warsaw, Poland). The GeneRuler™ 100 bp DNA Ladder (100–1000 bp) (Fermentas Thermo Scientific, Waltham, MA, USA) was used as the standard.
3.5. Data Collection and Analysis
The results for all amplification products were transferred to a binary matrix where “1” denoted the presence and “0” denoted the absence of a product. Genetic diversity analyses were performed in GenAlEx 6.5 (The Australian National University, Canberra, Australia) [58], AFLPsurv 1.0 (Laboratory of Plant Ecology and Biogeochemistry, Université Libre de Bruxelles, Brussels, Belgium [59], Popgene 1.32 (Department of Renewable Resources, University of Alberta, Edmonton, Alberta, Canada) [60] and MEGA 7.0 (The Arizona State University, Tempe, Arizona, USA) [61]. Basic parameters were calculated to describe the effectiveness of the applied marker systems and genetic diversity within the analyzed locations of S. purpurea: the number and percentage of polymorphic amplification products (pP, %p), the number and percentage of private products for each location (pM; %pM), the scored products ratio (SPR) describing the number of scored products per primer/combination of primers, the number of detected products (Na) [62], unbiased expected heterozygosity (uHe) [63] and the Shannon diversity index (I) [64]. These parameters were calculated for each location based on data matrices for single marker systems and based on a combined data matrix from all marker systems. The proportion of genetic diversity components within and among locations was determined by analysis of molecular variance (AMOVA) [65]. Genetic differentiation (ΦST) among the analyzed locations was calculated and presented in principal coordinate analyses (PCoA) [66]. A genetic distance (DS) matrix [67] was used to develop an UPGMA dendrogram [68] to illustrate the genetic relationships among the analyzed S. purpurea genotypes. These analyses were also performed for single marker systems and for a combination of all methods.
4. Conclusions
This is the first study presenting the genetic diversity of S. purpurea from natural localities. The observed genetic diversity is representative for willow species whose genetic structure was largely determined by the geographical regions of the studied localities. Purple willow stands are sometimes small and fragmented, which partially influences their diversity and differentiation. As expected, gene flow and migration could play an important role in wind-pollinated tree species whose small seeds are also widely dispersed by wind. This study relied on three well-established marker systems which are often used in genetic analyses of Salix populations. The results of single marker systems were mostly comparable, but a combined analysis of all systems is a reproducible and highly suitable method for analyzing genetic diversity and variation of S. purpurea. A combination of neutral genetic marker systems with different levels of genetic diversity revealed a significant discriminative power allowing for characterization and identification of genotypes and locations that constitute valuable breeding material for the pharmaceutical industry. However, the applied marker systems could be replaced in the next important breeding steps by new methods and technologies. Effective sequencing techniques reveal thousands of SNPs and contribute to the search for adaptive variation. Nevertheless, our study shows that genetic diversity and structure of natural plant locations can be effectively described with neutral, anonymous, but well-established molecular marker methods.
The presented data relating to the genetic diversity and genetic relationships between S. purpurea locations expand our knowledge about the biology and biogeography of the species. They indicate that analyses combining various types of DNA marker systems produce more reliable results with regard to in understand genetic diversity patterns in the species. The resulting knowledge could be valuable and useful support for breeding programs of S. purpurea, conducting variety surveys and construction of mapping purple willow populations.
Supplementary Materials
Supplementary Materials can be found at https://www.mdpi.com/1422-0067/19/1/105/s1.
Acknowledgments
This study was supported by the Polish Ministry of Science and Higher Education (project No. N N310 088337); by the Department of Plant Breeding and Seed Production of the University of Warmia and Mazury (Olsztyn, Poland); and by the Büsgen-Institute (Göttingen, Germany). We would like to thank Reiner Finkeldey for his support and contribution to the project.
Author Contributions
Paweł Sulima is the corresponding author who designed the experiment, collected plant material, analyzed and interpreted experimental data and drafted the manuscript; Kathleen Prinz designed parts of the experiment, drafted and revised the manuscript, and approved the final version for publication; Jerzy A. Przyborowski designed parts of the experiment, collected plant material, drafted and revised the manuscript, and approved the final version for publication. The authors are accountable for all aspects of the work, and all questions relating to the accuracy and integrity of any part of the work have been appropriately investigated and resolved.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Braun, L.; Cohen, M. Herbs and Natural Supplements, Volume 1: An Evidence-Based Guide, 4th ed.; Elsevier Churchill Livingstone: Sydney, Australia, 2015; pp. 1–1384. [Google Scholar]
- Chrubasik, S.; Eisenberg, E.; Balan, E.; Weinberger, T.; Luzzati, R.; Conradt, C. Treatment of low back pain exacerbations with willow bark extract: A randomized double-blind study. Am. J. Med. 2000, 109, 9–14. [Google Scholar] [CrossRef]
- Chrubasik, S.; Kunzel, O.; Model, A.; Conradt, C.; Black, A. Treatment of low back pain with a herbal or synthetic anti-rheumatic: A randomized controlled study. Willow bark extract for low back pain. Rheumatology 2001, 40, 1388–1393. [Google Scholar] [CrossRef] [PubMed]
- Wichtl, M. Herbal Drugs and Phytopharmaceuticals, 3rd ed.; CRC Press: London, UK, 2004. [Google Scholar]
- Cameron, M.; Gagnier, J.J.; Little, C.V.; Parsons, T.J.; Blümle, A.; Chrubasik, S. Evidence of effectiveness of herbal medicinal products in the treatment of arthritis. Part 1: Osteoarthritis. Phytother. Res. 2009, 23, 1497–1515. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Gagnier, J.J.; Little, C.V.; Parsons, T.J.; Blümle, A.; Chrubasik, S. Evidence of effectiveness of herbal medicinal products in the treatment of arthritis. Part 2: Rheumatoid arthritis. Phytother. Res. 2009, 23, 1647–1662. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, J.G. Medicinal potential of willow: A chemical perspective of aspirin discovery. J. Saudi Chem. Soc. 2010, 14, 317–322. [Google Scholar] [CrossRef]
- Shara, M.; Stohs, S.J. Efficacy and safety of white willow bark (Salix alba) extracts. Phytother. Res. 2015, 29, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Sulima, P.; Krauze-Baranowska, M.; Przyborowski, J.A. Variations in the chemical composition and content of salicylic glycosides in the bark of Salix purpurea from natural locations and their significance for breeding. Fitoterapia 2017, 118, 118–125. [Google Scholar] [CrossRef] [PubMed]
- EMEA. Final assessment report on Salix [various species including S. purpurea L., S. daphnoides Vill., S. fragilis L.], cortex; European Medicines Agency: London, UK, 2017; pp. 4–5. [Google Scholar]
- Förster, N.; Ulrichs, C.; Zander, M.; Kätzel, R. Factors influencing the variability of antioxidative phenolic glycosides in Salix species. J. Agric. Food. Chem. 2010, 58, 8205–8210. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E.; Florineth, F.; Hadacek, F. Weiden in Österreich und Angrenzenden Gebieten; Arbeitsbereich Ingenieurbiologie u. Landschaftsbau, Univ. Bodenkultur Wien: Wien, Austria, 2002; pp. 7–150. [Google Scholar]
- Argus, G.W. Salix (Salicaceae) distribution maps and a synopsis of their classification in North America, north of Mexico. Harv. Pap. Bot. 2007, 12, 335–368. [Google Scholar] [CrossRef]
- Sulima, P.; Przyborowski, J.A.; Wiwart, M. Willow bark—Herbal raw material harvested from plants cultivated on arable lands. Herba Pol. 2006, 52, 4. [Google Scholar]
- Kenstaviciene, P.; Nenortiene, P.; Kiliuviene, G.; Zevzikovas, A.; Lukosius, A.; Kazlauskiene, D. Application of high-performance liquid chromatography for research of salicin in bark of different varieties of Salix. Medicina 2009, 45, 644–651. [Google Scholar] [PubMed]
- Meneghetti, S.; Barcaccia, G.; Paiero, P.; Lucchin, M. Genetic characterization of Salix alba L. and Salix fragilis L. by means of different PCR-derived marker systems. Plant Biosyst. 2007, 141, 283–291. [Google Scholar] [CrossRef]
- Chen, J.H.; Sun, H.; Wen, J.; Yang, Y.P. Molecular phylogeny of Salix L. (Salicaceae) inferred from three chloroplast datasets and its systematic implications. Taxon 2010, 59, 29–37. [Google Scholar]
- Przyborowski, J.A.; Sulima, P.; Kuszewska, A.; Załuski, D.; Kilian, A. Phylogenetic relationships between four Salix L. species based on DArT markers. Int. J. Mol. Sci. 2013, 14, 24113–24125. [Google Scholar] [CrossRef] [PubMed]
- Sulima, P.; Przyborowski, J.A.; Załuski, D. RAPD markers reveal genetic diversity in Salix purpurea L. Crop. Sci. 2009, 49, 857–863. [Google Scholar] [CrossRef]
- Aravanopoulos, F. Clonal identification based on quantitative, co-dominant, and dominant marker data: A comparative analysis of selected willow (Salix L.) clones. Int. J. For. Res. 2010. [Google Scholar] [CrossRef]
- Ngantcha, A.C. DNA Fingerprinting and Genetic Relationships among Willow (Salix spp.). Master’s Thesis, University of Saskatchewan, Saskatoon, Canada, 2010. [Google Scholar]
- Sulima, P.; Przyborowski, J.A. Genetic diversity of Salix purpurea L. genotypes and interspecific hybrids. Acta Biol. Crac. Ser. Bot. 2013, 55, 29–36. [Google Scholar] [CrossRef]
- The Genome Sequence of Salix purpurea Female Clone 94006. Salix purpurea v1.0, DOE-JGI. Available online: http://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Spurpurea (accessed on 9 December 2017).
- Martin, T.; Rönnberg-Wästljung, A.C.; Stenlid, J.; Samils, B. Identification of a differentially expressed TIR-NBS-LRR gene in a major QTL associated to leaf rust resistance in Salix. PLoS ONE 2016, 11, e0168776. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wang, X.; Bi, C.; Xu, Y.; Wu, D.; Ye, N. Assembly and analysis of the complete Salix purpurea L. (Salicaceae) mitochondrial genome sequence. SpringerPlus 2016, 5, 1894. [Google Scholar] [CrossRef] [PubMed]
- Bracci, T.; Busconi, M.; Fogher, C.; Sebastiani, L. Molecular studies in olive (Olea europaea L.): Overview on DNA markers applications and recent advances in genome analysis. Plant Cell Rep. 2011, 30, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.B.; Cetin, O.; Kaya, H.; Sahin, M.; Sefer, F.; Kahraman, A.; Tanyolac, B. SNP discovery by Illumina-based transcriptome sequencing of the olive and the genetic characterization of Turkish olive genotypes revealed by AFLP, SSR and SNP markers. PLoS ONE 2013, 8, e73674. [Google Scholar] [CrossRef] [PubMed]
- Frascaroli, E.; Schrag, T.A.; Melchinger, A.E. Genetic diversity analysis of elite European maize (Zea mays L.) inbred lines using AFLP, SSR, and SNP markers reveals ascertainment bias for a subset of SNPs. Theor. Appl. Genet. 2013, 126, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Skvortsov, A.K. Willows of Russia and Adjacent Countries. Taxonomical and Geographical Revision; University of Joensuu: Joensuu, Finland, 1999; pp. 10–254. [Google Scholar]
- Dolatowski, J.; Seneta, W. Dendrologia, 3rd ed.; PWN: Warszawa, Poland, 2017; ISBN 978-8-30-115369-4. [Google Scholar]
- USDA, NRCS. The PLANTS Database. National Plant Data Team, Greensboro, NC, USA. Available online: https://plants.usda.gov/core/profile?symbol=SAPU2 (accessed on 9 December 2017).
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Penner, G.A.; Bush, A.; Wise, R.; Kim, W.; Domier, L.; Kasha, K.; Laroche, A.; Scoles, G.; Molnar, S.J.; Fedak, G. Reproducibility of random amplified polymorphic DNA (RAPD) analysis among laboratories. Methods Appl. 1993, 2, 341–345. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.M.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Saliba-Colombani, V.; Causse, M.; Gervais, L.; Philouze, J. Efficiency of RFLP, RAPD, and AFLP markers for the construction of an intraspecific map of the tomato genome. Genome 2000, 43, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.; Pereira, G.; Garrido, I.; Tavares-de-Sousa, M.M.; Espinosa, F. Comparison of RAPD, ISSR, and AFLP molecular markers to reveal and classify orchardgrass (Dactylis glomerata L.) germplasm variations. PLoS ONE 2016, 11, e0152972. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.H.A.; Matthes, M.; Arnold, G.M.; Edwards, K.J.; Ahman, I.; Larsson, S.; Karp, A. Characterization of genetic diversity in potential biomass willows (Salix spp.) by RAPD and AFLP analyses. Genome 1999, 42, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Van Puyvelde, K.; Triest, L. ISSRs indicate isolation by distance and spatial structuring in Salix alba populations along Alpine upstream rivers (Alto Adige and Upper Rhine). Belg. J. Bot. 2007, 140, 100–108. [Google Scholar]
- Trybush, S.O.; Jahodová, Š.; Macalpine, W.; Karp, A. A genetic study of a Salix germplasm resource reveals new insights into relationships among sub-genera, sections and species. Bioenergy Res. 2008, 1, 67–79. [Google Scholar] [CrossRef]
- Lin, J.; Gibbs, J.P.; Smart, L.B. Population genetic structure of native versus naturalized sympatric shrub willows (Salix; Salicaceae). Am. J. Bot. 2009, 96, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Przyborowski, J.A.; Sulima, P. The analysis of genetic diversity of Salix viminalis genotypes as a potential source of biomass by RAPD markers. Ind. Crop. Prod. 2010, 31, 395–400. [Google Scholar] [CrossRef]
- Trybush, S.O.; Jahodová, Š.; Čížková, L.; Karp, A.; Hanley, S.J. High levels of genetic diversity in Salix viminalis of the Czech Republic as revealed by microsatellite markers. Bioenergy Res. 2012, 5, 969–977. [Google Scholar] [CrossRef]
- Berlin, S.; Ghelardini, L.; Bonosi, L.; Weih, M.; Rönnberg-Wästljung, A.C. QTL mapping of biomass and nitrogen economy traits in willows (Salix spp.) grown under contrasting water and nutrient conditions. Mol. Breed. 2014, 34, 1987–2003. [Google Scholar] [CrossRef]
- Lauron-Moreau, A.; Pitre, F.E.; Argus, G.W.; Labrecque, M.; Brouillet, L. Phylogenetic relationships of american willows (Salix L., Salicaceae). PLoS ONE 2015, 10, e0121965. [Google Scholar]
- Zhai, F.; Mao, J.; Liu, J.; Peng, X.; Han, L.; Sun, Z. Male and female subpopulations of Salix viminalis present high genetic diversity and high long-term migration rates between them. Front. Plant Sci. 2016, 7, 330. [Google Scholar] [CrossRef] [PubMed]
- Alsos, I.G.; Alm, T.; Normand, S.; Brochmann, C. Past and future range shifts and loss of diversity in dwarf willow (Salix herbacea L.) inferred from genetics, fossils and modelling. Glob. Ecol. Biogeogr. 2009, 18, 223–239. [Google Scholar] [CrossRef]
- Berlin, S.; Trybush, S.O.; Fogelqvist, J.; Gyllenstrand, N.; Hallingbäck, H.R.; Åhman, I.; Nordh, N.E.; Shield, I.; Powers, S.J.; Weih, M.; et al. Genetic diversity, population structure and phenotypic variation in European Salix viminalis L. (Salicaceae). Tree Genet. Genomes 2014, 10, 1595–1610. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Sulima, P.; Przyborowski, J.A. Natural Occurrence of Purple Willow (Salix purpurea). Available online: https://www.google.com/maps/d/u/0/edit?mid=1lFNsWPj63seNFPMTADXIsZP41Yw (accessed on 9 December 2017).
- Neumann, A. Die Mitteleuropäischen Salix-Arten; Mitt Forstlichen Bundes-Versuchsanstalt: Wien, Austria, 1981; pp. 9–144. [Google Scholar]
- Milligan, B.G. Total DNA isolation. In Molecular Genetic Analysis of Populations: A Practical Approach; Hoelzel, A.R., Ed.; Oxford University Press: Oxford, UK, 1998; pp. 29–64. [Google Scholar]
- Hantula, J.; Dusabenygasani, M.; Hamelin, R.C. Random Amplified Microsatellites (RAMS)—A novel method for characterizing genetic variation within fungi. For. Pathol. 1996, 26, 159–166. [Google Scholar] [CrossRef]
- McGregor, C.E.; Lambert, C.A.; Greyling, M.N.; Louw, J.H.; Warnich, L. A comparative assessment of DNA fingerprinting techniques (RAPD, ISSR, AFLP and SSR) in tetraploid potato (Solanum tuberosum L.) germplasm. Euphytica 2000, 113, 135–144. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Vekemans, X. AFLPsurv V.1.0. A Software for Genetic Diversity Analysis with AFLP Population Data; Université Libre de Bruxelles: Brussel, Belgium, 2001; pp. 1–15. [Google Scholar]
- Yeh, F.C.; Yang, R.C.; Boyle, T.B.J.; Ye, Z.H.; Mao, J.X. POPGENE, the User-Friendly Shareware for Population Genetic Analysis; University of Alberta: Edmonton, Canada. Available online: https://sites.ualberta.ca/~fyeh/popgene_download.html (accessed on 9 December 2017).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics, 4th ed.; Sinauer: Sunderland, MA, USA, 2007. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; pp. 3–125. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Gower, J.C. Principal Coordinates Analysis. In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Chichester, UK, 2005. [Google Scholar]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy: The Principles and Practice of Numerical Classification; W.H. Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
Figure 1.
PCoA (principal coordinates analysis) plots of genetic differentiation (ΦST) between the analyzed populations of S. purpurea determined with the use of: AFLP (A); RAPD (B); and ISSR (C) marker systems; and a combination of all marker systems (D). The analyzed locations are marked in different colors.
Figure 1.
PCoA (principal coordinates analysis) plots of genetic differentiation (ΦST) between the analyzed populations of S. purpurea determined with the use of: AFLP (A); RAPD (B); and ISSR (C) marker systems; and a combination of all marker systems (D). The analyzed locations are marked in different colors.
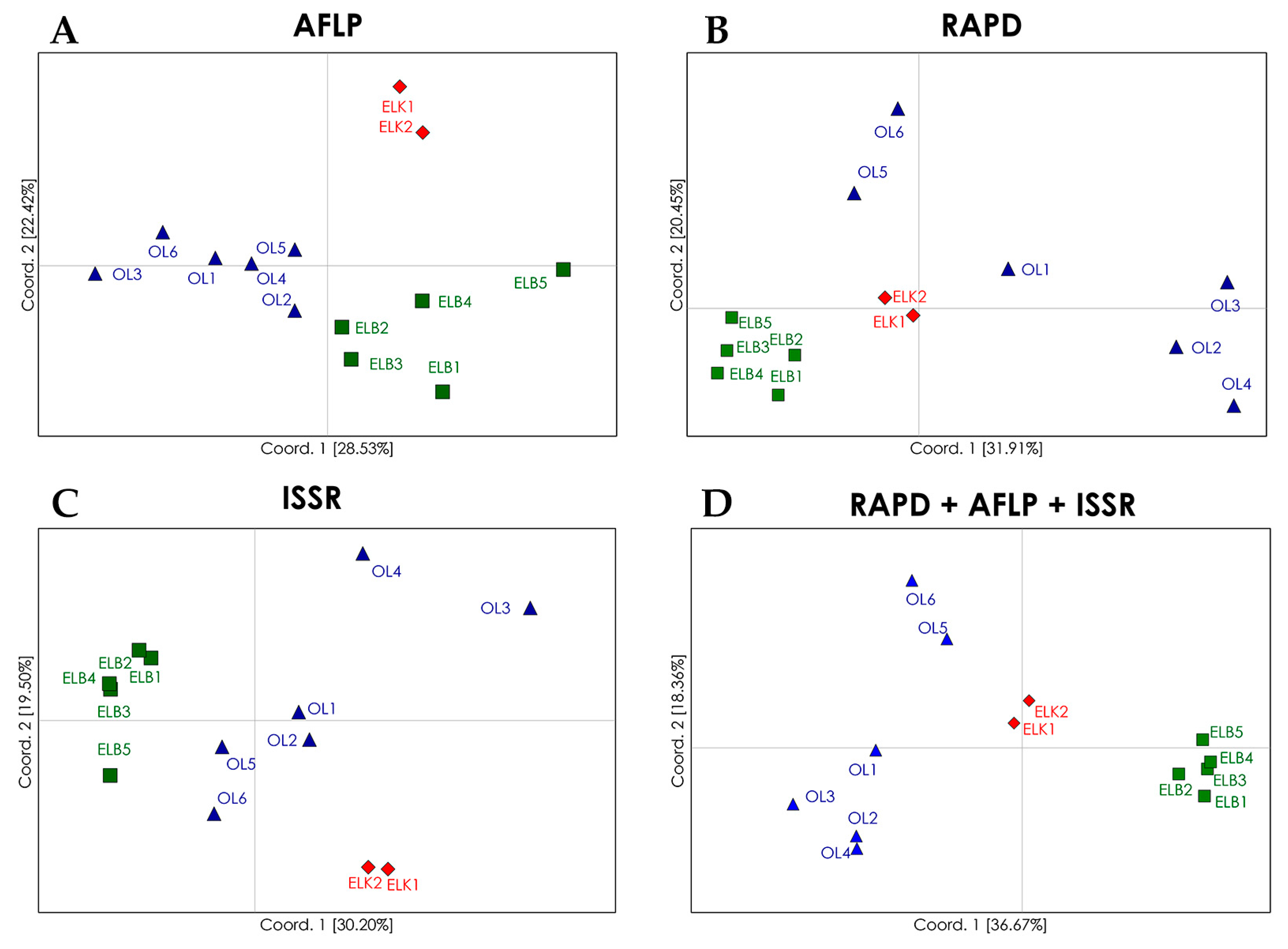
Figure 2.
UPGMA (unweighted pair group method with arithmetic mean) dendrogram of genetic distance between 91 genotypes of S. purpurea based on a combined analysis of all used marker systems (AFLP, RAPD and ISSR). The analyzed locations are marked in different colors.
Figure 2.
UPGMA (unweighted pair group method with arithmetic mean) dendrogram of genetic distance between 91 genotypes of S. purpurea based on a combined analysis of all used marker systems (AFLP, RAPD and ISSR). The analyzed locations are marked in different colors.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Marker systems efficiency for analysis of genetic diversity in S. purpurea using AFLPs (amplified length polymorphisms), RAPDs (randomly amplified polymorphic DNA) and ISSRs (inter-simple sequence repeats) and a combination of all marker systems.
Table 1.
Marker systems efficiency for analysis of genetic diversity in S. purpurea using AFLPs (amplified length polymorphisms), RAPDs (randomly amplified polymorphic DNA) and ISSRs (inter-simple sequence repeats) and a combination of all marker systems.
| Parameter | AFLP | RAPD | ISSR | All Products |
|---|---|---|---|---|
| Total number of products (L) | 159 | 574 | 221 | 954 |
| Percent of polymorphic loci (%p) | 74.8 | 89.9 | 87.3 | 86.8 |
| Scored products ratio (SPR) | 79.5 | 9.3 | 11.1 | 11.4 |
| Shannon diversity index (I) | 0.187 | 0.264 | 0.258 | 0.247 |
| Unbiased expected heterozygosity (uHe) | 0.134 | 0.192 | 0.187 | 0.179 |
| Minimum uHe within a location (MinuHe) | 0.096 | 0.114 | 0.091 | 0.110 |
| Maximum uHe within a location (MaxuHe) | 0.170 | 0.232 | 0.220 | 0.218 |
| Genetic differentiation between locations (ΦST) | 0.150 | 0.159 | 0.225 | 0.212 |
Table 2.
Polymorphic (pP) and private amplification products (pM) for the analyzed locations of S. purpurea.
Table 2.
Polymorphic (pP) and private amplification products (pM) for the analyzed locations of S. purpurea.
| Location | Parameter | AFLP | RAPD | ISSR | All Marker Systems |
|---|---|---|---|---|---|
| ELK1 | pP | 125 | 481 | 186 | 792 |
| pM | 0 | 2 | 1 | 3 | |
| ELK2 | pP | 138 | 517 | 199 | 854 |
| pM | 2 | 7 | 0 | 9 | |
| ELB1 | pP | 108 | 390 | 154 | 652 |
| pM | 1 | 0 | 2 | 3 | |
| ELB2 | pP | 124 | 469 | 186 | 779 |
| pM | 0 | 2 | 1 | 3 | |
| ELB3 | pP | 109 | 444 | 171 | 724 |
| pM | 1 | 0 | 1 | 2 | |
| ELB4 | pP | 116 | 449 | 171 | 736 |
| pM | 0 | 3 | 0 | 3 | |
| ELB5 | pP | 102 | 404 | 158 | 664 |
| pM | 0 | 1 | 1 | 2 | |
| OL1 | pP | 119 | 492 | 185 | 796 |
| pM | 4 | 1 | 1 | 6 | |
| OL2 | pP | 98 | 404 | 158 | 660 |
| pM | 0 | 1 | 2 | 3 | |
| OL3 | pP | 95 | 342 | 125 | 562 |
| pM | 1 | 1 | 1 | 3 | |
| OL4 | pP | 108 | 382 | 142 | 632 |
| pM | 0 | 1 | 1 | 2 | |
| OL5 | pP | 116 | 456 | 168 | 740 |
| pM | 2 | 2 | 0 | 4 | |
| OL6 | pP | 107 | 395 | 159 | 661 |
| pM | 1 | 1 | 2 | 4 | |
| Average | pP | 112.7 | 432.7 | 166.3 | 711.7 |
| pM | 0.9 | 1.7 | 1.0 | 3.6 | |
| %p | 70.9 | 75.4 | 75.3 | 74.6 | |
| %pM | 7.5 | 3.8 | 5.9 | 4.9 |
%p, average percentage of polymorphic amplification products per location; %pM, average percentage of private products per location; ELK, Ełk Lakeland; ELB, Żuławy Wiślane; OL, Olsztyn Lakeland.
Table 3.
Genetic diversity parameters in the studied locations of S. purpurea based on all DNA marker systems (RAPD, AFLP and ISSR).
Table 3.
Genetic diversity parameters in the studied locations of S. purpurea based on all DNA marker systems (RAPD, AFLP and ISSR).
| Location | N | Na | Ne | I | uHe | %p |
|---|---|---|---|---|---|---|
| ELK1 | 9 | 1.421 | 1.325 | 0.295 | 0.205 | 59.78 |
| ELK2 | 14 | 1.582 | 1.348 | 0.322 | 0.218 | 69.16 |
| ELB1 | 4 | 1.057 | 1.250 | 0.212 | 0.164 | 38.04 |
| ELB2 | 9 | 1.382 | 1.323 | 0.288 | 0.202 | 57.61 |
| ELB3 | 6 | 1.220 | 1.286 | 0.248 | 0.181 | 46.88 |
| ELB4 | 7 | 1.285 | 1.300 | 0.266 | 0.190 | 51.77 |
| ELB5 | 5 | 1.082 | 1.261 | 0.220 | 0.165 | 39.40 |
| OL1 | 11 | 1.454 | 1.339 | 0.306 | 0.211 | 62.36 |
| OL2 | 4 | 1.090 | 1.270 | 0.228 | 0.178 | 40.76 |
| OL3 | 3 | 0.829 | 1.157 | 0.134 | 0.110 | 23.51 |
| OL4 | 6 | 1.056 | 1.249 | 0.212 | 0.157 | 38.99 |
| OL5 | 8 | 1.317 | 1.306 | 0.273 | 0.193 | 53.94 |
| OL6 | 5 | 1.067 | 1.245 | 0.209 | 0.156 | 38.45 |
| Average | 7.0 | 1.218 | 1.282 | 0.247 | 0.179 | 47.74 |
N, sample size; Na, number of different alleles; Ne, average number of effective alleles; I, Shannon diversity index; uHe, unbiased expected heterozygosity; %p, percent of polymorphic loci.
Table 4.
Genetic differentiation ΦST (below diagonal) between the analyzed locations of S. purpurea based on AFLP, RAPD and ISSR results.
Table 4.
Genetic differentiation ΦST (below diagonal) between the analyzed locations of S. purpurea based on AFLP, RAPD and ISSR results.
| Location | ELK1 | ELK2 | ELB1 | ELB2 | ELB3 | ELB4 | ELB5 | OL1 | OL2 | OL3 | OL4 | OL5 | OL6 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ELK1 | 0.000 | ||||||||||||
| ELK2 | 0.028 | 0.000 | |||||||||||
| ELB1 | 0.194 | 0.173 | 0.000 | ||||||||||
| ELB2 | 0.166 | 0.146 | 0.004 | 0.000 | |||||||||
| ELB3 | 0.188 | 0.167 | 0.081 | 0.068 | 0.000 | ||||||||
| ELB4 | 0.187 | 0.154 | 0.106 | 0.082 | 0.045 | 0.000 | |||||||
| ELB5 | 0.222 | 0.188 | 0.166 | 0.131 | 0.132 | 0.131 | 0.000 | ||||||
| OL1 | 0.160 | 0.149 | 0.190 | 0.172 | 0.184 | 0.191 | 0.219 | 0.000 | |||||
| OL2 | 0.221 | 0.233 | 0.244 | 0.235 | 0.265 | 0.291 | 0.312 | 0.098 | 0.000 | ||||
| OL3 | 0.244 | 0.224 | 0.322 | 0.257 | 0.319 | 0.318 | 0.369 | 0.101 | 0.222 | 0.000 | |||
| OL4 | 0.241 | 0.238 | 0.287 | 0.260 | 0.269 | 0.280 | 0.317 | 0.158 | 0.190 | 0.222 | 0.000 | ||
| OL5 | 0.181 | 0.157 | 0.216 | 0.172 | 0.209 | 0.202 | 0.229 | 0.098 | 0.234 | 0.223 | 0.242 | 0.000 | |
| OL6 | 0.249 | 0.201 | 0.294 | 0.239 | 0.257 | 0.280 | 0.300 | 0.143 | 0.271 | 0.296 | 0.292 | 0.119 | 0.000 |
Table 5.
Analysis of molecular variance (AMOVA) based on all 954 amplification products (AFLP, RAPD and ISSR) for 91 genotypes of S. purpurea (p < 0.01).
Table 5.
Analysis of molecular variance (AMOVA) based on all 954 amplification products (AFLP, RAPD and ISSR) for 91 genotypes of S. purpurea (p < 0.01).
| Source of Variation | Df | Sum of Squares | Mean Square | Estimated Variation | Total Variance |
|---|---|---|---|---|---|
| Between regions | 2 | 1314.12 | 657.06 | 14.17 | 10% |
| Between locations | 10 | 2005.46 | 200.55 | 14.46 | 11% |
| Within locations | 78 | 8319.15 | 106.66 | 106.66 | 79% |
| Total | 90 | 11,638.74 | – | 135.28 | 100% |
Table 6.
Geographic information relating to the analyzed natural locations of S. purpurea.
| Location | Latitude/Longitude | Geographic Region | Number of Genotypes |
|---|---|---|---|
| ELK1 | 53°48′59.94″/22°23′12.06″ | Ełk Lakeland—ELK | 9 |
| ELK2 | 53°50′23.52″/22°22′41.62″ | Ełk Lakeland—ELK | 14 |
| ELB1 | 54°8′37.35″/19°23′4.98″ | Delta of the Vistula River—ELB | 4 |
| ELB2 | 54°7′34.18″/19°18′28.95″ | Delta of the Vistula River—ELB | 9 |
| ELB3 | 54°11′17.02″/19°12′29.94″ | Delta of the Vistula River—ELB | 6 |
| ELB4 | 54°15′30.93″/19°14′18.96″ | Delta of the Vistula River—ELB | 7 |
| ELB5 | 54°9′14.92″/19°1′10.77″ | Delta of the Vistula River—ELB | 5 |
| OL1 | 53°45′30.02″/20°29′12.62″ | Olsztyn Lakeland—OL | 11 |
| OL2 | 53°46′18.72″/20°26′36.12″ | Olsztyn Lakeland—OL | 4 |
| OL3 | 53°46′43.26″/20°30′42.54″ | Olsztyn Lakeland—OL | 3 |
| OL4 | 53°43′3.06″/20°28′18.72″ | Olsztyn Lakeland—OL | 6 |
| OL5 | 53°52′31.56″/20°21′25.14″ | Olsztyn Lakeland—OL | 8 |
| OL6 | 53°51′54.90″/20°22′45.30″ | Olsztyn Lakeland—OL | 5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sulima, P.; Prinz, K.; Przyborowski, J.A. Genetic Diversity and Genetic Relationships of Purple Willow (Salix purpurea L.) from Natural Locations. Int. J. Mol. Sci. 2018, 19, 105. https://doi.org/10.3390/ijms19010105
AMA Style
Sulima P, Prinz K, Przyborowski JA. Genetic Diversity and Genetic Relationships of Purple Willow (Salix purpurea L.) from Natural Locations. International Journal of Molecular Sciences. 2018; 19(1):105. https://doi.org/10.3390/ijms19010105
Chicago/Turabian StyleSulima, Paweł, Kathleen Prinz, and Jerzy A. Przyborowski. 2018. "Genetic Diversity and Genetic Relationships of Purple Willow (Salix purpurea L.) from Natural Locations" International Journal of Molecular Sciences 19, no. 1: 105. https://doi.org/10.3390/ijms19010105
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.