Natural Antisense Transcripts: Molecular Mechanisms and Implications in Breast Cancers
by
 and
and
Guillaume Latgé
1, 
Christophe Poulet
1,
Vincent Bours
1,2,
Claire Josse
1,3,4 and
Guy Jerusalem
3,4,* 1
Laboratory of Human Genetics, GIGA-Institute, University of Liège, 4500 Liège, Belgium
2
Center of Genetics, University Hospital (CHU), 4500 Liège, Belgium
3
Department of Medical Oncology, University Hospital (CHU), 4500 Liège, Belgium
4
Laboratory of Medical Oncology, GIGA-Institute, University of Liège, 4500 Liège, Belgium
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(1), 123; https://doi.org/10.3390/ijms19010123
Submission received: 13 October 2017
/
Revised: 7 December 2017
/
Accepted: 29 December 2017
/
Published: 2 January 2018
(This article belongs to the Collection Regulation by Non-coding RNAs)
Abstract
:Natural antisense transcripts are RNA sequences that can be transcribed from both DNA strands at the same locus but in the opposite direction from the gene transcript. Because strand-specific high-throughput sequencing of the antisense transcriptome has only been available for less than a decade, many natural antisense transcripts were first described as long non-coding RNAs. Although the precise biological roles of natural antisense transcripts are not known yet, an increasing number of studies report their implication in gene expression regulation. Their expression levels are altered in many physiological and pathological conditions, including breast cancers. Among the potential clinical utilities of the natural antisense transcripts, the non-coding|coding transcript pairs are of high interest for treatment. Indeed, these pairs can be targeted by antisense oligonucleotides to specifically tune the expression of the coding-gene. Here, we describe the current knowledge about natural antisense transcripts, their varying molecular mechanisms as gene expression regulators, and their potential as prognostic or predictive biomarkers in breast cancers.
1. Introduction
After an international effort, the scientific community has revealed that up to 90% of the human genome is transcribed. Thanks to the FANTOM project (functional annotation of the mammalian genome, available online: http://fantom.gsc.riken.jp/), started in 2000 with the mouse genome [1,2], which was quickly followed by the human genome in 2003 by the ENCODE project (encyclopedia of DNA elements, available online: https://www.encodeproject.org/) [3,4], we know that 98% of the human genome is composed of non-coding (nc) sequences, previously considered “Junk DNA” due to their heterogeneity, low expression levels, and unknown functions [5,6,7,8,9,10,11]. This huge part of the transcriptome could therefore play a role in protein-coding (pc) RNA expression regulation. Databases specialized in genome annotation, such as the GENCODE project (encyclopædia of genes and gene variants, available online: http://www.gencodegenes.org/) [12,13], specialized in ncRNA, such as the NONCODE (integrated knowledge database dedicated to ncRNAs, especially lncRNAs, available online: http://www.noncode.org/) and RNAcentral projects (the non-coding RNA sequence database, available online: http://rnacentral.org/) [14,15], or specialized in human long non-coding RNAs (lncRNAs), such as the LNCipedia project (a comprehensive compendium of human long non-coding RNAs, available online: https://lncipedia.org/) [16], are now illustrating this new vision of the transcriptome.
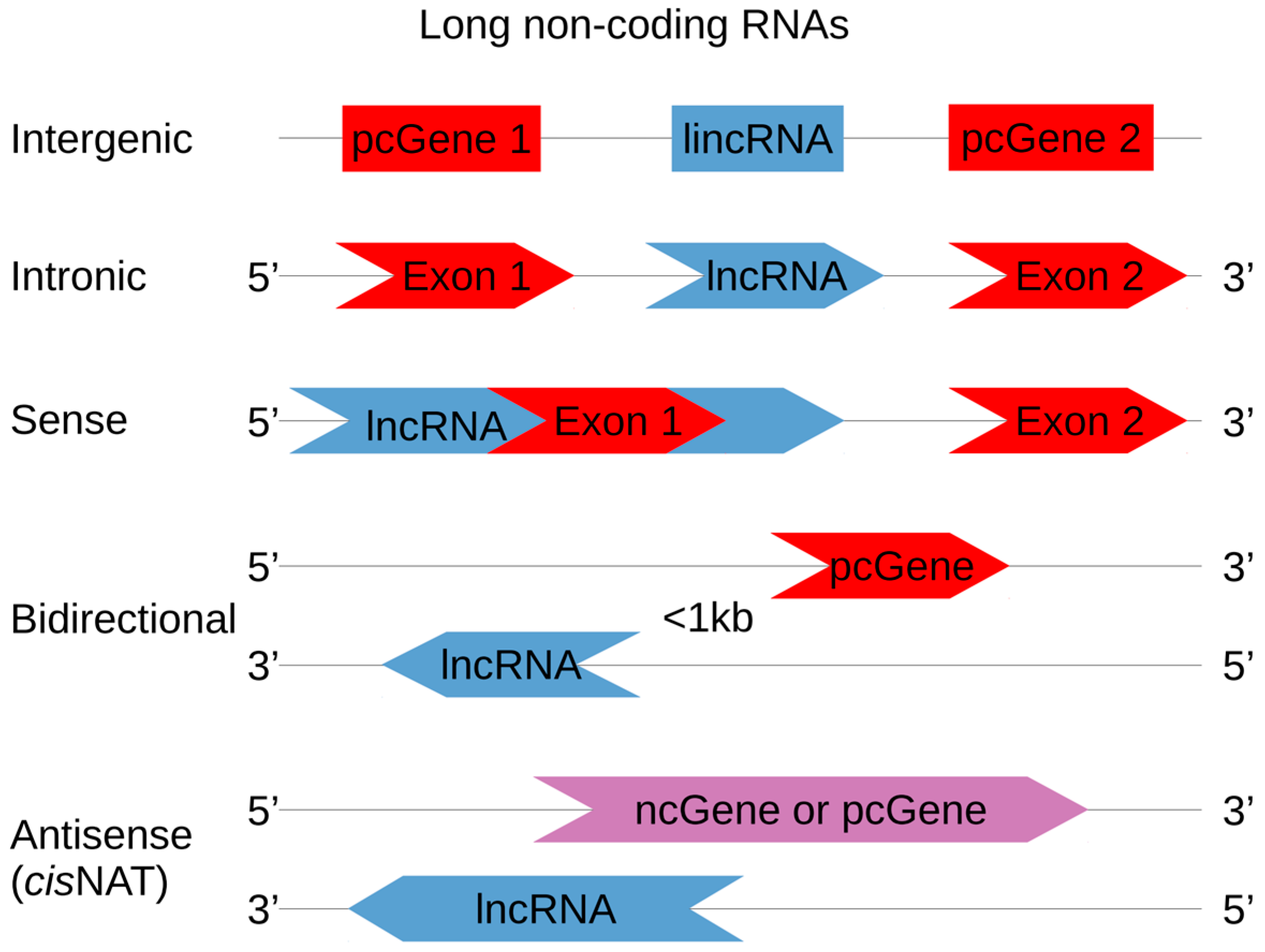
Non-coding RNAs are classified according to their transcript length. With few exceptions, a 200-nt arbitrary threshold is used to separate short and long ncRNAs [17]. The long non-coding RNAs (lncRNAs) make up the largest portion of ncRNAs. With approximatively 98% of the genome containing non-coding regions and only 1.1% containing exons, it is obvious that many lncRNAs do not overlap exons. lncRNAs are classified according to their original genomic location and their context when compared to a protein-coding gene (pcGene) [18,19,20]. Figure 1 depicts the five current categories of lncRNAs, named intergenic, intronic, bidirectional (or divergent), sense and antisense. Sense and antisense lncRNAs are defined according to the nearest pcGene position. Both can overlap, partially or entirely, one or more exons of a pcGene [2]. Bidirectional lncRNA transcription starts close to a pcGene (less than 1 kb from the pcGene transcription start site) and proceeds in the opposite direction as pcGene transcription. Intronic lncRNAs are transcribed entirely from introns, and do not overlap with any exon [20,21,22]. Those in intergenic regions are named long intergenic non-coding RNAs (lincRNAs), and their transcription can occur in both directions [23,24]. Some ncRNA classification exceptions are also worth noting. A few lncRNAs, such as BC1 or snaR, contain less than or close to 200 nt, but they are classified as lncRNAs in the lncRNAdb database (the reference database for functional long noncoding RNAs, available online: http://lncrnadb.org/) [25]. Other lncRNAs can reach lengths of 1 Mbase and are thus called very long intergenic RNA (vlincRNA) [26].
lncRNAs are defined as endogenous cellular RNAs without a significant ORF (open reading frame) [27,28,29]. However, some ncRNAs containing an ORF smaller than 100 amino-acids may be classified as lncRNAs [27]. The known biological roles of lncRNAs are very heterogeneous and cover various molecular and cellular functions such as pcGene regulation [30], stem cell pluripotency and differentiation [31], allelic expression [32], cell cycle control [33], apoptosis and senescence [34], heat shock response [35], and control of chromatin modifications [36]. It is worth noting that lncRNAs are found in all tissues and show pronounced tissue-specific expression. Their cellular location may vary, probably reflecting their function [20,37,38]. There is a structural similarity between lncRNAs and mRNAs, in the sense that they may be multi-exonic, 5′ capped, 3′ polyadenylated, and spliced [23]. RNA polymerase II (RNA Pol II) is responsible for the transcription of most of the lncRNAs, and their expression is under the control of promoters and enhancers, that can be induced by external stimuli [23].
2. Generic Definition of NATs
Natural antisense transcripts (NATs) are coding or non-coding RNA sequences that are complementary to and overlap with either protein-coding or non-coding transcripts [39]. As 98% of the transcriptome is non-coding, the vast majority of paired transcripts are composed of nc|nc or nc|pc pairs. Therefore, NATs are defined in regard to the relative genomic position from their paired transcript origins, in cis or in trans. Cis-NAT pairs are transcribed from the opposite strand of the same genomic locus and display perfect RNA|RNA sequence complementarity with the opposite strand transcript (if no RNA modifications, such as RNA editing, occur). Trans-NAT pairs are transcribed from different genomic loci, and the two RNA molecules may hybridize to each other with imperfect RNA|RNA sequence complementarity [40,41].
Because whole genome sequencing of the antisense transcription has only been available for less than a decade, many NATs were described as lncRNAs without information about the co-existence of other transcripts from the same genomic origin. This convergence between NAT and lncRNA classifications may thus lead to some confusion in the literature and will probably disappear with the increasing knowledge in the antisense transcription field.
3. NAT: Structure, Localization, and Expression Regulation
Like lncRNAs and mRNAs, NATs may be capped and poly-adenylated transcripts that are maturated to excise introns. NAT expression is also controlled by promoters and enhancers. In addition, many examples of bidirectional promoters that control transcript expression originating from both strands are described in the literature [42,43]. In this case, several transcription factors, such as GABPA or E2F1, are preferentially implicated [44,45,46]. NATs may originate from cryptic promoters that are then inserted within the intronic regions of a gene or close to the transcription start site of neighboring genes [43,47,48].
NATs accumulate preferentially in the nucleus, associating with chromatin, unlike coding mRNAs which accumulate in the cytoplasm. NATs are also found in other cellular compartments, such as mitochondria, and have been reported to accumulate at polysomes [3,18,49]. Moreover, NAT expression is closely linked with the activity of their sense or neighboring genes [43].
4. NAT: Role, Function and Mechanism of Action
The biological significance of NATs remains under scientific investigation with major key questions yet to be answered. Specific pcGene regulation by their corresponding overlapping ncNATs has been reported. Our team and others have shown that up to 50% of the pcGenes also express ncNATs [2,39,50] and that transcript levels of nc|pc pairs are often tightly correlated [39,46,50]. Altogether, this suggests that NATs could be implicated in a new level of gene expression regulation [5,51].
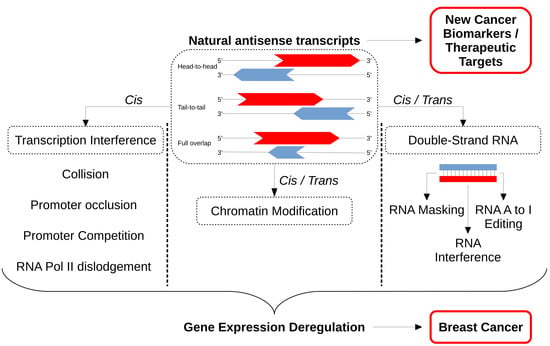
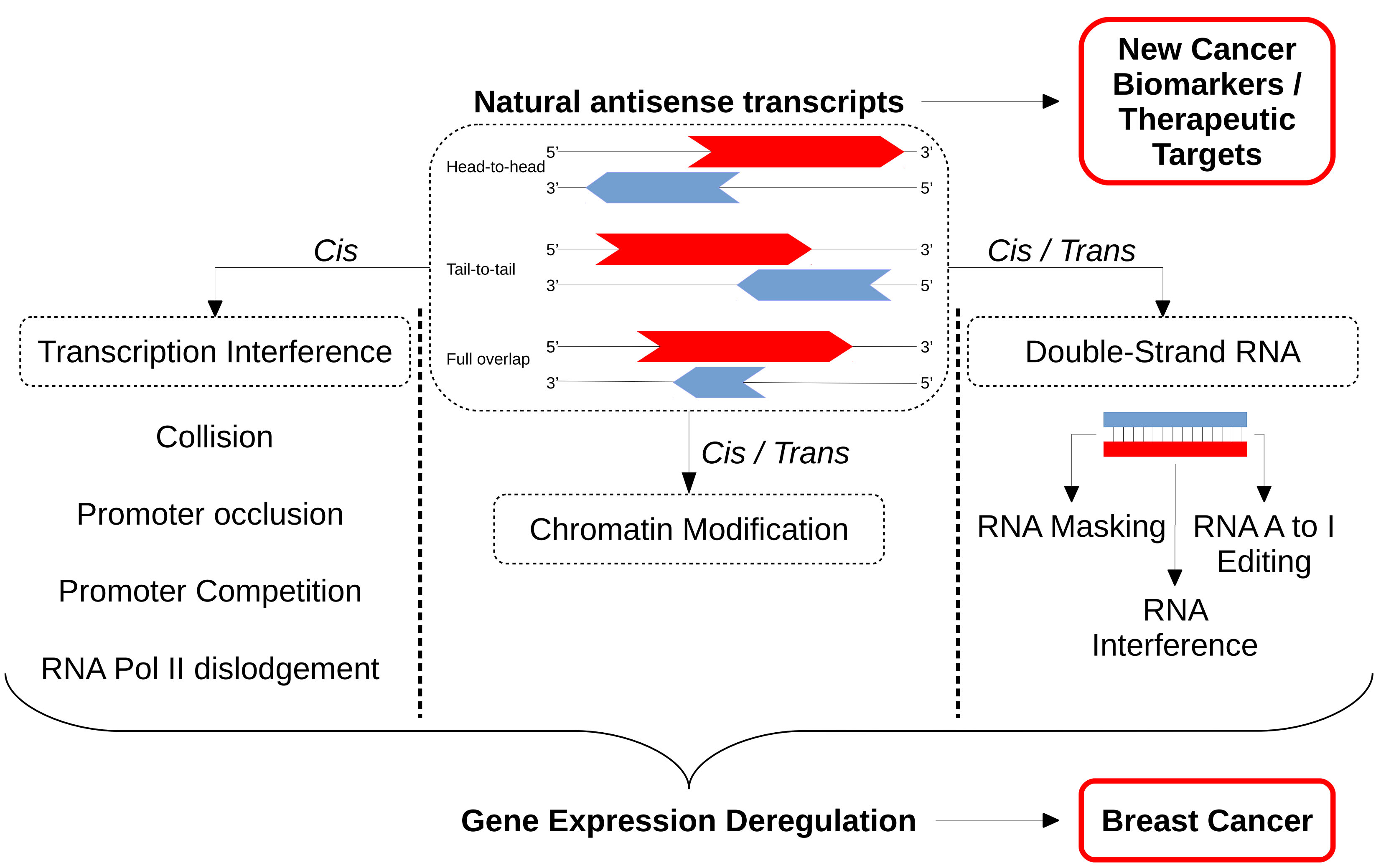
Both transcriptional and post-transcriptional regulations of expression have also been explained as the result of the creation of natural sense and antisense transcript pairs. The regulatory processes implicated can be more or less complex, ranging from simple transcriptional interference to modulation of chromatin changes or the formation of double-stranded RNA (dsRNA). The latter leads to RNA masking, RNA interference or RNA editing [52].
Several examples of pcGene expression regulation by their NATs are described hereafter to illustrate the different molecular mechanisms of action.
4.1. Action in Cis or Trans
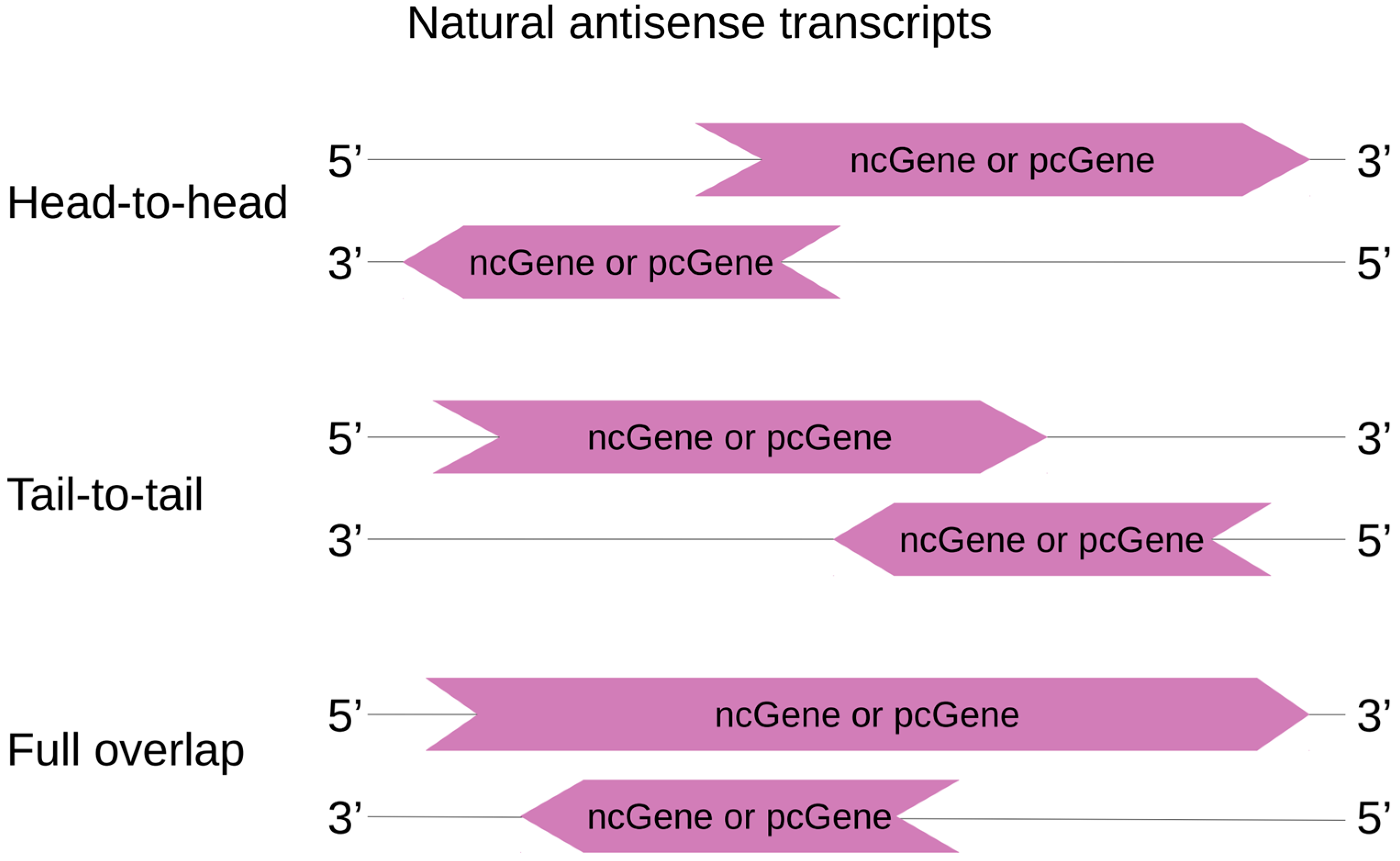
While NATs are more likely to handle regulation of other genes in cis, they may also tune gene expression elsewhere in the genome by trans regulation. Based on the definition of overlapping genes from Makalowska et al., cis-NATs are here classified according to the relative position of the DNA coding sequence of the RNA transcripts [53]. Three categories can thus be described and are depicted in Figure 2: (1) “head-to-head”, where sense and antisense transcripts overlap on their 5′ ends; (2) “tail-to-tail”, where sense and antisense transcripts overlap on their 3′ ends; and (3) “embedded overlap” (also called “full overlap”), where one of the entire transcript overlaps the other.
4.2. Transcriptional Interference
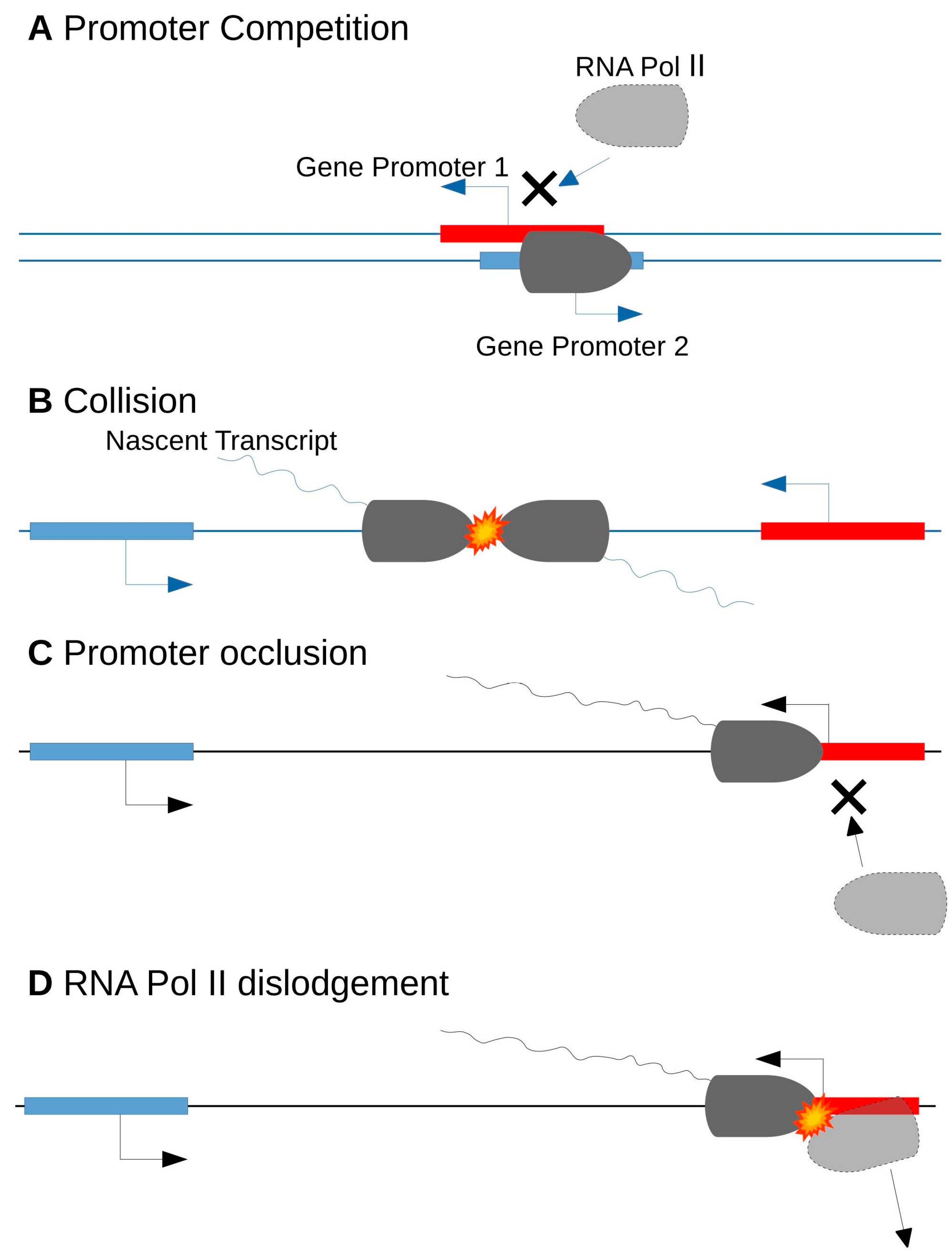
Antisense transcription can modulate in cis the sense transcription of the opposite strand, although this effect may not be caused by the pairing of the RNA molecules themselves. The proximity of the two transcriptional events, sense and antisense, leads to a downregulation of both transcripts [54]. Transcriptional interference can occur during the initiation or elongation phases of transcription. In the initiation phase, promoters of head-to-head NATs are competing for the use of RNA Pol II and common regulatory elements (Figure 3A). In the elongation phase, interference can occur after the following events: a collision between RNA Pol II complexes, leading to a machinery blockage (Figure 3B); a promoter occlusion by RNA Pol II during the antisense transcript elongation (Figure 3C); or an RNA Pol II dislodgement by the RNA Pol II standing on the opposite strand, when the first one was too slow to start (Figure 3D) [54]. It is worth noting that the transcriptional interference investigation field is still young and that formal proof of gene expression regulation by this mechanism was only recently reported [55]. Nevertheless, a negative correlation between sense and antisense transcript levels are less frequently observed than a positive correlation or no correlation. This suggests that only a minority of NATs could be involved in transcriptional interference processes [50,56,57,58,59].
Despite difficulties in discriminating transcription interference from gene expression regulation by RNA transcripts, Stojic et al. [55] have demonstrated such a mechanism by screening an siRNA library. Whereas nearly all siRNAs dampen GNG12-AS1 (a non-coding natural antisense transcript of the tumor suppressor coding gene DIRAS3) post-transcriptionally, siRNA targeting exon 1 of GNG12-AS1 downregulates its transcription by recruiting Argonaute 2 and inhibiting RNA polymerase II binding. In this case, the active transcription of GNG12-AS1 causes the transcriptional silencing of DIRAS3, leading to increased cell proliferation.
4.3. Chromatin Modification
ncNATs may regulate the expression levels of the sense pcGenes by regulating chromatin modifications. Such epigenetic modifications encompass DNA methylations of cytosine in CpG islands and histone modifications by methylation or acetylation of lysine residues. NATs and, more widely, ncRNAs, are thought to affect DNA methylation by interacting with various types of proteins involved in histone modification or chromatin remodeling such as, in particular, the polycomb repressive complex 2 (PRC2) [60]. A current hypothesis considers that nascent NATs guide PRC2 to specific-target sites on the chromatin. The tethering would occur by pairing the nascent NAT with DNA or mRNA sequences, during or after NAT transcription (Figure 4A) [61].
Additionally, a “decoy” mechanism can be described, where the NAT binds a protein complex, such as PRC2, and prevents this complex from binding the sense transcript by competition. This complex can also prevent the interaction of the sense gene with RNA Pol II or the chromatin [61,62].
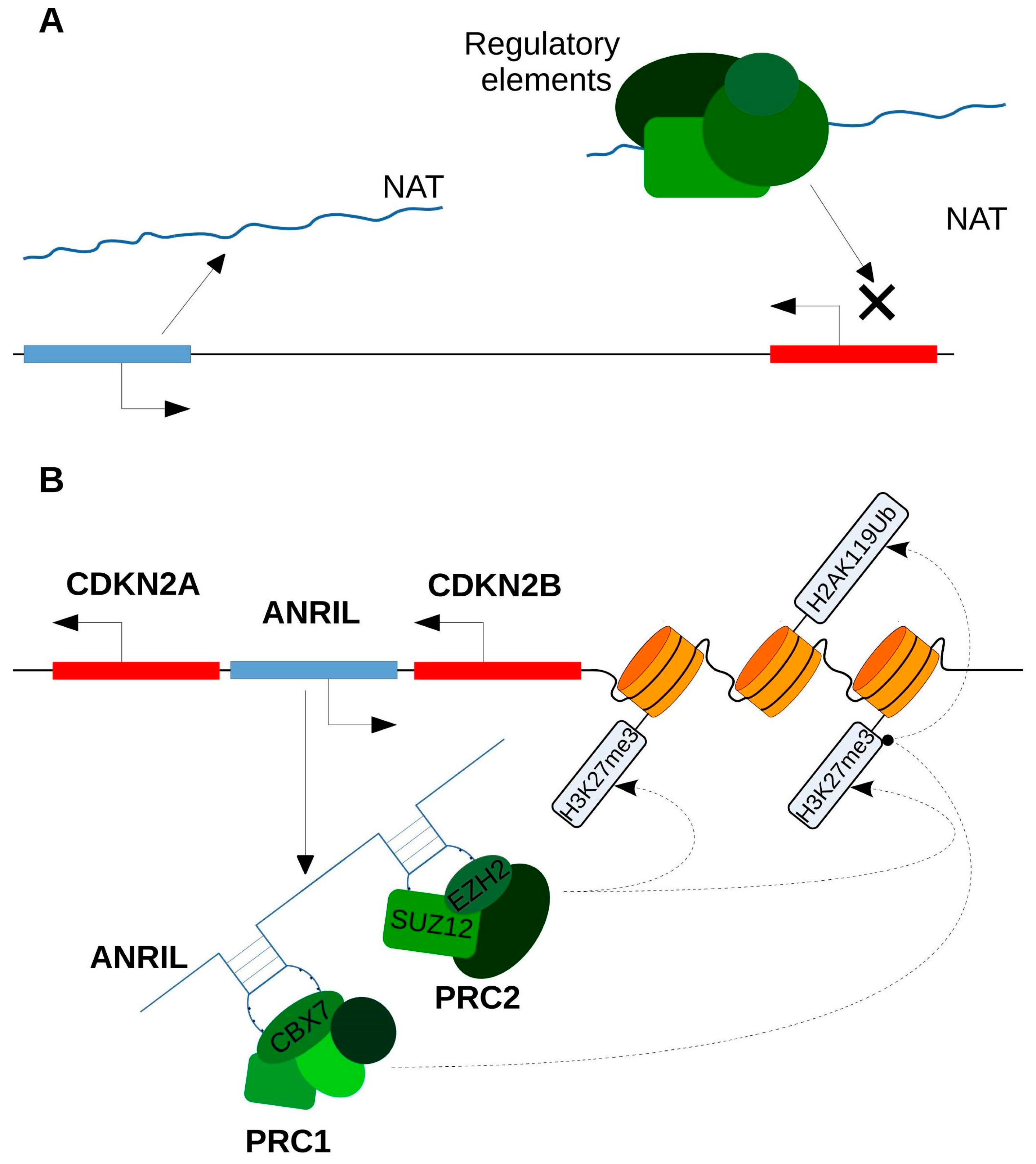
Here are two examples of lncRNA/NAT that play a role in the tethering of PRC2 with chromatin. The first example is the combined action of ANRIL and PRC1–PCR2 on INK4b-ARF-INK4a gene expression and on the chromatin structure of this locus. ANRIL is a cisNAT that is dysregulated in breast cancer. It is located in the INK4b-ARF-INK4a gene cluster, which contains three genes encoding the three tumor-suppressor proteins p15, p14 and p16 [63]. Polycomb repressive complexes 1 and 2 (PRC1 and PRC2) are implicated in epigenetic silencing mechanisms. ANRIL can recruit those complexes to the chromatin of the INK4b-ARF-INK4a locus, recruiting PRC2 through interaction with SUZ12 and EZH2 components, and recruiting PRC1 by binding CBX7 [63,64,65]. Next, PRC2 silences INK4b-ARF-INK4a gene expression by inducing H3K27 tri-methylation, and PRC1 maintains a repressive chromatin structure by mono-ubiquitination of H2AK119 (Figure 4B) [66].
A second example is HOTAIR, which is implicated and dysregulated in many types of cancer and displays an active and critical role in chromatin dynamics [67,68]. Like ANRIL, HOTAIR interacts with PRC2 through its 5′ end to induce H3K27 tri-methylation. In addition, HOTAIR binds to LSD1 (lysine-specific demethylase 1) by its 3′ end, leading to H3K4 demethylation. These combined modifications lead, in trans, to a repressive chromatin structure and thus to the silencing of multiple genes [68,69].
4.3.1. Double-Stranded RNA/RNA Masking
NATs can regulate gene expression through the formation of a complex of two overlapping NAT sequences. This double-stranded RNA (dsRNA) molecule thus creates a physical protection against post-transcriptional regulation factors that target the pcGene. RNA masking will then interfere with splicing or translation machineries. This mechanism will also prevent miRNA binding or RNAse activities, which often target single-stranded RNA and influence their complex stability [52]. Under this condition and in opposition with other mechanisms described above, NAT positively regulates pcGene expression.
In osteocarcinoma, upregulated FGFR3-AS1 forms a tail-to-tail dsRNA with FGFR3, its sense transcript. FGFR3 mRNA is thus protected against RNase activity, leading to an increase in both its mRNA stability and its protein production [70]. Conversely, binding of the MALAT1 3′ UTR by its ncNAT TALAM1 allows for RNase P cleavage, leading to 3′ end processing and maturation that is essential for MALAT1 stability and function [71].
While forming dsRNAs, NATs can also interfere with splicing and translation mechanisms. For example, the protein coded from the gene ZEB2 is a transcriptional factor that downregulates E-cadherin and its antisense transcript, ZEB2-AS1. ZEB2 also contains an IRES (internal ribosome entry site) required for its translation. By binding this sequence, ZEB2-AS1 promotes ZEB2 splicing and downregulates its protein expression [72].
4.3.2. Double-Stranded RNA/RNA A to I Editing
ADARs (adenosine deaminases that act on RNA) are enzymes responsible for RNA editing by site-specific adenosine deamination. They target dsRNA molecules such as those formed by NAT pairs. After adenosine to inosine (A-to-I) editing, inosines (I) are interpreted as a guanosines (G) during splicing or translation. Such modification may modulate the localization or the stability of the edited transcripts [73,74]. The occurrence frequency of RNA editing by NATs is not yet characterized [52,75,76]. Indeed, few NATs display edited sequences, but they may be quickly degraded or retained in the nucleus, thus disappearing from the bulk of the expressed sequences [77].
An example of this A to I editing mechanism has been found in human prostate cancers with the sense/antisense couple of PRUNE2 and PCA3 transcripts. PRUNE2 is a pcGene that has a tumor suppressor role. PCA3 is an NAT that originates from introns, and is fully overlapped by PRUNE2’s 6th intron. The dsRNA created by PCA3 and PRUNE2’s pre-mRNA forms a complex with ADAR proteins. An A-to-I editing of this dsRNA leads to a downregulation of protein expression and an increase in tumor cell growth [78]. It is important to note that PCA3 was also approved as a specific biomarker for diagnostic tests.
4.3.3. Double-Stranded RNA/RNA Interference
RNA interference is an additional mechanism whereby NATs are implicated in pcGene post-transcriptional regulation [79]. RNA interference is the endogenous siRNA formation from NAT-derived dsRNA. RNA interference is DICER-dependent and is followed by the action of the RNA-induced silencing complex (RISC) [80,81,82]. NATs may thus serve as precursors in endo-siRNA and miRNA production [83]. NATs form internal hairpins or duplexes with sense RNA, leading to a dsRNA that can be handled and digested by DICER. Short RNA duplexes will then be bound by the RISC complex, where one strand of the RNA duplex is used as a guide for mRNA recognition. This mRNA is then cleaved by the RISC complex, which will decrease the protein expression. Even with scarce evidence of NAT involvement in the RNA interference process, recent transcriptome sequencing studies have shown the widespread occurrence of endo-siRNAs and their regulatory potential during stages of development and differentiation [82,83].
5. NATs in Breast Cancer
Numerous studies have highlighted a link between lncRNA/NAT and cancers, especially breast cancers. Most of these transcripts were either highlighted by high-throughput transcriptomic studies that lacked the strand origin, or explored one by one due to their implication in oncogenic pathways. Therefore, many lncRNA listed in Table 1 are generally not described as NAT in the literature. In addition, the expression correlation between the NAT pair transcripts, as well as the ncNAT regulatory role with regard to the paired pcGene, are often unknown. It is also worth noting that most genomic loci coding for NAT transcript pairs also display numerous alternative transcripts. Therefore, each lncRNA transcript may belong to different classes among NAT pc|nc, lincRNA, lncRNA, or NAT nc|nc.
To the best of our knowledge, only three strand-specific whole genome transcriptomic studies were performed on breast cancer samples [39,46,50]. The main concordant conclusions were that: (i) pcGene transcription coincides with an antisense ncNAT transcription in 50% of the cases; (ii) NAT transcripts are 1000 times less abundant than pcGene transcripts; and (iii) positive expression correlations between ncNATs and their paired pcGenes are approximately six times more frequent than negative correlations. This latest suggests that if ncNATs can affect the expression of their corresponding pcGene, positive regulation of expression should be more frequently observed than repression. However, a comparison of transcript levels between tumors and paired non-malignant adjacent healthy tissues showed that the ncNAT/pcGene transcript balance is disrupted in tumors. Therefore, new positive correlations of NAT/pcGene pairs are created in tumor tissues, while others that were present in the normal tissue decline [50].
The mechanism by which lncRNA/NAT regulates pcGene expression is known in several instances, and two mechanisms are often described in breast cancer. The first is driven by the polycomb repressing complexes (PRC), and the second by microRNAs. Here are three examples of PRC2 involvement in cancer pathways. The NAT ANRASSF1 leads PRC2 binding on the RASSF1 promoter to regulate RASSF1 expression [84]. The INK4b-ARF-INK4A locus coding for the cell cycle associated proteins p14, p15 and p16 is regulated by the NAT ANRIL via PRC2, and in addition, the lncRNA PANDAR recruits PRC1 to also regulate p16 expression [63,64,65,85]. Similarly, the p53 pathway is regulated at several levels via PCR2 by HOTAIR and MEG3 lncRNAs [86,87,88,89]. The importance of gene regulation by PRC2 is well known in breast cancers, as the expression of its targeted genes can be used to predict patient outcomes [90].
As displayed in Table 1, microRNAs are also frequently involved in gene regulation by lncRNA. One particular example is the epithelial to mesenchymal transition (EMT) that is regulated by three lncRNAs, namely, H19, linc-RoR and TP73-AS, which capture multiple microRNAs and prevent their binding to other mRNA targets [91,92,93,94].
5.1. NATs as Cancer Biomarkers
Like mRNAs, the expression levels of NATs and lncRNAs are affected under cancerous conditions. Differences in mRNA expression patterns between different subgroups of breast cancer patients have been used to develop genomic tests able to predict patient’s prognosis, or to predict treatment response by breast cancers. Among them, we can underline the MammaPrint and PAM50 microarray-based gene signatures, or the Oncotype DX RT-PCR–based assay that can help clinicians make treatment decisions based on the calculation of the recurrence risk, and/or the benefits of chemotherapy in the case of Oncotype DX test [179,180,181].
Similarly, multiple NATs/lncRNAs display expression levels that are associated with the disease prognosis, the treatment response or the clinical classification of breast cancers (Table 1). Although no clinically validated test has emerged yet, several studies report prognostic ncRNA gene signatures [119,182,183,184,185].
5.2. NATs as Therapeutic Targets
The understanding of antisense transcription is important for therapies. Indeed, NATs represent a potential highly specific entry point for therapeutic intervention on targeted genes by the use of ASO (antisense oligonucleotides) that are drugs already FDA-approved for several diseases [186].
Functionally characterized NATs can be targeted by ASOs, called in this case antagoNATs, to block the interaction of the sense and antisense transcripts. The hybridization of ASOs with the antisense transcript would lead to its degradation, or to transcriptional de-repression at the chromatin level [187]. The first in vivo demonstration of antagoNAT efficacy was shown by Modarresi et al. [188] and has been validated in other clinical contexts, detailed in the review by MacLeod et al. [187].
6. Conclusions
Up to 90% of the human genome length is transcribed: ~2% of the genomic DNA is coding for proteins; ~88% is transcribed but do not encode proteins; and ~10% is not transcribed. In contrast, ~80% of the RNA transcripts are coding for proteins and the remaining ~20% do not. These sequences are thus less expressed than the coding ones. They are also less conserved between species than coding genes, but more conserved than the non-coding and the non-transcribed genes. Such transcripts must therefore play a biological role, which has yet to be described.
Among lncRNAs, NATs are coding or non-coding RNA sequences, which are complementary to and overlapping with either protein-coding or non-coding transcripts. Their main biological role is thought to be the regulation of pcGene expression through a variety of molecular mechanisms. High-throughput transcriptomic studies have demonstrated that the expression of NATs and lncRNAs is modified under cancerous conditions, making them good cancer biomarkers. Finally, non-coding/pcGene transcript pairs are interesting, especially for specific target-gene treatments using ASO.
Acknowledgments
This work was supported by the French Community of Belgium, Belgian Funds for Scientific Research (F.R.S.-FNRS), the F.R.S.-FNRS-Televie, CHU Liege (F.I.R.S) and the Region Wallonne (XSPRELTRIN).
Author Contributions
Guillaume Latgé and Claire Josse compiled the current literature and wrote the first draft of the article. Christophe Poulet contributed to scientific and language editing. Vincent Bours, Claire Josse and Guy Jerusalem supervised the project. All authors provided critical feedback and helped shape the manuscript writing.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| NAT | Natural antisense transcripts |
| nc | Non-coding |
| pc | Protein-coding |
| ds | Double-stranded |
| pcGene | Protein-coding gene |
| lncRNA | Long non-coding RNA |
| lincRNA | Long intergenic non-coding RNA |
| ncRNA | Non-coding RNA |
| mRNA | Messenger RNA |
| RNA Pol II | RNA polymerase II |
| PRC | Polycomb Repressive Complex |
| ASO | Antisense oligonucleotide |
References
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawai, J.; et al. Antisense transcription in the mammalian transcriptome. Science 2005, 309, 1564–1566. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium; Birney, E.; Stamatoyannopoulos, J.A.; Dutta, A.; Gingeras, T.R.; Margulies, E.H.; Weng, Z.; Snyder, M.; Dermitzakis, E.T.; Thurman, R.E.; et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 2007, 447, 799–816. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Ikeya, Y.; Okumura, T.; Kimura, T. Post-transcriptional inducible gene regulation by natural antisense RNA. Front. Biosci. 2015, 20, 1–36. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed]
- Taft, R.J.; Pang, K.C.; Mercer, T.R.; Dinger, M.; Mattick, J.S. Non-coding RNAs: Regulators of disease. J. Pathol. 2010, 220, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Kellis, M.; Wold, B.; Snyder, M.P.; Bernstein, B.E.; Kundaje, A.; Marinov, G.K.; Ward, L.D.; Birney, E.; Crawford, G.E.; Dekker, J.; et al. Defining functional DNA elements in the human genome. Proc. Natl. Acad. Sci. USA 2014, 111, 6131–6138. [Google Scholar] [CrossRef] [PubMed]
- Frankish, A.; Uszczynska, B.; Ritchie, G.R.S.; Gonzalez, J.M.; Pervouchine, D.; Petryszak, R.; Mudge, J.M.; Fonseca, N.; Brazma, A.; Guigo, R.; et al. Comparison of GENCODE and RefSeq gene annotation and the impact of reference geneset on variant effect prediction. BMC Genom. 2015, 16 (Suppl. 8), S2. [Google Scholar] [CrossRef] [PubMed]
- Jalali, S.; Gandhi, S.; Scaria, V. Navigating the dynamic landscape of long noncoding RNA and protein-coding gene annotations in GENCODE. Hum. Genom. 2016, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q.; et al. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Res. 2016, 44, D203–D208. [Google Scholar] [CrossRef] [PubMed]
- The RNAcentral Consortium. RNAcentral: A comprehensive database of non-coding RNA sequences. Nucleic Acids Res. 2017, 45, D128–D134. [Google Scholar] [CrossRef]
- Volders, P.-J.; Helsens, K.; Wang, X.; Menten, B.; Martens, L.; Gevaert, K.; Vandesompele, J.; Mestdagh, P. LNCipedia: A database for annotated human lncRNA transcript sequences and structures. Nucleic Acids Res. 2013, 41, D246–D251. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, H.I.; Amaral, P.P.; Louro, R.; Lopes, A.; Fachel, A.A.; Moreira, Y.B.; El-Jundi, T.A.; da Silva, A.M.; Reis, E.M.; Verjovski-Almeida, S. Genome mapping and expression analyses of human intronic noncoding RNAs reveal tissue-specific patterns and enrichment in genes related to regulation of transcription. Genome Biol. 2007, 8, R43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Laurent, G.; Savva, Y.A.; Kapranov, P. Dark matter RNA: An intelligent scaffold for the dynamic regulation of the nuclear information landscape. Front. Genet. 2012, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Gardner, E.J.; Nizami, Z.F.; Talbot, C.C., Jr.; Gall, J.G. Stable intronic sequence RNA (sisRNA), a new class of noncoding RNA from the oocyte nucleus of Xenopus tropicalis. Genes Dev. 2012, 26, 2550–2559. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. lncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res. 2011, 39, D146–D151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Laurent, G.; Shtokalo, D.; Dong, B.; Tackett, M.R.; Fan, X.; Lazorthes, S.; Nicolas, E.; Sang, N.; Triche, T.J.; McCaffrey, T.A.; et al. VlincRNAs controlled by retroviral elements are a hallmark of pluripotency and cancer. Genome Biol. 2013, 14, R73. [Google Scholar] [CrossRef] [PubMed]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Mattick, J.S. Differentiating protein-coding and noncoding RNA: Challenges and ambiguities. PLoS Comput. Biol. 2008, 4, e1000176. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Carmichael, G.G. Long noncoding RNAs in mammalian cells: What, where, and why? Wiley Interdiscip. Rev. RNA 2010, 1, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef] [PubMed]
- Vance, K.W.; Ponting, C.P. Transcriptional regulatory functions of nuclear long noncoding RNAs. Trends Genet. 2014, 30, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinger, M.E.; Amaral, P.P.; Mercer, T.R.; Pang, K.C.; Bruce, S.J.; Gardiner, B.B.; Askarian-Amiri, M.E.; Ru, K.; Soldà, G.; Simons, C.; et al. Long noncoding RNAs in mouse embryonic stem cell pluripotency and differentiation. Genome Res. 2008, 18, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Maenner, S.; Müller, M.; Becker, P.B. Roles of long, non-coding RNA in chromosome-wide transcription regulation: Lessons from two dosage compensation systems. Biochimie 2012, 94, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, M.; Kitagawa, K.; Kotake, Y.; Niida, H.; Ohhata, T. Cell cycle regulation by long non-coding RNAs. Cell. Mol. Life Sci. 2013, 70, 4785–4794. [Google Scholar] [CrossRef] [PubMed]
- Grammatikakis, I.; Panda, A.C.; Abdelmohsen, K.; Gorospe, M. Long noncoding RNAs(lncRNAs) and the molecular hallmarks of aging. Aging 2014, 6, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Dammert, M.A.; Hoppe, S.; Bierhoff, H.; Grummt, I. Heat shock represses rRNA synthesis by inactivation of TIF-IA and lncRNA-dependent changes in nucleosome positioning. Nucleic Acids Res. 2016, 44, 8144–8152. [Google Scholar] [CrossRef] [PubMed]
- Angrand, P.-O.; Vennin, C.; Le Bourhis, X.; Adriaenssens, E. The role of long non-coding RNAs in genome formatting and expression. Front. Genet. 2015, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Tsuiji, H.; Yoshimoto, R.; Hasegawa, Y.; Furuno, M.; Yoshida, M.; Nakagawa, S. Competition between a noncoding exon and introns: Gomafu contains tandem UACUAAC repeats and associates with splicing factor-1. Genes Cells 2011, 16, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Balbin, O.A.; Malik, R.; Dhanasekaran, S.M.; Prensner, J.R.; Cao, X.; Wu, Y.-M.; Robinson, D.; Wang, R.; Chen, G.; Beer, D.G.; et al. The landscape of antisense gene expression in human cancers. Genome Res. 2015, 25, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-T.; Zhang, Y.; Kong, L.; Liu, Q.-R.; Wei, L. Trans-natural antisense transcripts including noncoding RNAs in 10 species: Implications for expression regulation. Nucleic Acids Res. 2008, 36, 4833–4844. [Google Scholar] [CrossRef] [PubMed]
- Vanhée-Brossollet, C.; Vaquero, C. Do natural antisense transcripts make sense in eukaryotes? Gene 1998, 211, 1–9. [Google Scholar] [CrossRef]
- Seila, A.C.; Calabrese, J.M.; Levine, S.S.; Yeo, G.W.; Rahl, P.B.; Flynn, R.A.; Young, R.A.; Sharp, P.A. Divergent transcription from active promoters. Science 2008, 322, 1849–1851. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, L.; Luo, W.; Zhang, X. Characteristics of antisense transcript promoters and the regulation of their activity. Int. J. Mol. Sci. 2016, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.; Block, S.; Coombs, C.; Martin, M.E. Identification of functional elements in the murine Gabp alpha/ATP synthase coupling factor 6 bi-directional promoter. Gene 2006, 369, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Schuettengruber, B.; Doetzlhofer, A.; Kroboth, K.; Wintersberger, E.; Seiser, C. Alternate activation of two divergently transcribed mouse genes from a bidirectional promoter is linked to changes in histone modification. J. Biol. Chem. 2003, 278, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Grinchuk, O.V.; Motakis, E.; Yenamandra, S.P.; Ow, G.S.; Jenjaroenpun, P.; Tang, Z.; Yarmishyn, A.A.; Ivshina, A.V.; Kuznetsov, V.A. Sense-antisense gene-pairs in breast cancer and associated pathological pathways. Oncotarget 2015, 6, 42197–42221. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, I.; Rando, O.J.; Delrow, J.; Tsukiyama, T. Chromatin remodelling at promoters suppresses antisense transcription. Nature 2007, 450, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Xu, Z.; Clauder-Münster, S.; Steinmetz, L.M.; Buratowski, S. Set3 HDAC mediates effects of overlapping noncoding transcription on gene induction kinetics. Cell 2012, 150, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Giannakakis, A.; Zhang, J.; Jenjaroenpun, P.; Nama, S.; Zainolabidin, N.; Aau, M.Y.; Yarmishyn, A.A.; Vaz, C.; Ivshina, A.V.; Grinchuk, O.V.; et al. Contrasting expression patterns of coding and noncoding parts of the human genome upon oxidative stress. Sci. Rep. 2015, 5, 9737. [Google Scholar] [CrossRef] [PubMed]
- Wenric, S.; ElGuendi, S.; Caberg, J.-H.; Bezzaou, W.; Fasquelle, C.; Charloteaux, B.; Karim, L.; Hennuy, B.; Frères, P.; Collignon, J.; et al. Transcriptome-wide analysis of natural antisense transcripts shows their potential role in breast cancer. Sci. Rep. 2017, 7, 17452. [Google Scholar] [CrossRef] [PubMed]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Rosikiewicz, W.; Makałowska, I. Biological functions of natural antisense transcripts. Acta Biochim. Pol. 2016, 63, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Makalowska, I.; Lin, C.-F.; Makalowski, W. Overlapping genes in vertebrate genomes. Comput. Biol. Chem. 2005, 29, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shearwin, K.E.; Callen, B.P.; Egan, J.B. Transcriptional interference--a crash course. Trends Genet. 2005, 21, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Stojic, L.; Niemczyk, M.; Orjalo, A.; Ito, Y.; Ruijter, A.E.M.; Uribe-Lewis, S.; Joseph, N.; Weston, S.; Menon, S.; Odom, D.T.; et al. Transcriptional silencing of long noncoding RNA GNG12-AS1 uncouples its transcriptional and product-related functions. Nat. Commun. 2016, 7, 10406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeder, S.; Mages, J.; Flicek, P.; Lang, R. Uncovering information on expression of natural antisense transcripts in Affymetrix MOE430 datasets. BMC Genom. 2007, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Grigoriadis, A.; Oliver, G.R.; Tanney, A.; Kendrick, H.; Smalley, M.J.; Jat, P.; Neville, A.M. Identification of differentially expressed sense and antisense transcript pairs in breast epithelial tissues. BMC Genom. 2009, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Conley, A.B.; Jordan, I.K. Epigenetic regulation of human cis-natural antisense transcripts. Nucleic Acids Res. 2012, 40, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.H.T.; Ban, Y.; Wen, H.; Wang, S.M.; Ge, S.X. Conserved expression of natural antisense transcripts in mammals. BMC Genom. 2013, 14, 243. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, H.; Wang, H. Long noncoding RNAs in DNA methylation: New players stepping into the old game. Cell Biosci. 2016, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Khorkova, O.; Myers, A.J.; Hsiao, J.; Wahlestedt, C. Natural antisense transcripts. Hum. Mol. Genet. 2014, 23, R54–R63. [Google Scholar] [CrossRef] [PubMed]
- Davidovich, C.; Cech, T.R. The recruitment of chromatin modifiers by long noncoding RNAs: Lessons from PRC2. RNA 2015, 21, 2007–2022. [Google Scholar] [CrossRef] [PubMed]
- Meseure, D.D.; Vacher, S.; Alsibai, K.D.; Nicolas, A.; Chemlali, W.; Caly, M.; Lidereau, R.; Pasmant, E.; Callens, C.; Bieche, I. Expression of ANRIL-Polycomb Complexes-CDKN2A/B/ARF Genes in Breast Tumors: Identification of a Two-Gene (EZH2/CBX7) Signature with Independent Prognostic Value. Mol. Cancer Res. 2016, 14, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Aguilo, F.; Zhou, M.-M.; Walsh, M.J. Long noncoding RNA, polycomb, and the ghosts haunting INK4b-ARF-INK4a expression. Cancer Res. 2011, 71, 5365–5369. [Google Scholar] [CrossRef] [PubMed]
- Congrains, A.; Kamide, K.; Ohishi, M.; Rakugi, H. ANRIL: Molecular mechanisms and implications in human health. Int. J. Mol. Sci. 2013, 14, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Sun, H.; He, B.; Pan, Y.; Gao, T.; Chen, J.; Ying, H.; Liu, X.; Wang, F.; Xu, Y.; et al. Prognostic value of long non-coding RNA HOTAIR in various cancers. PLoS ONE 2014, 9, e110059. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Mandal, S.S. LncRNA HOTAIR: A master regulator of chromatin dynamics and cancer. Biochim. Biophys. Acta 2015, 1856, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, X.; Fu, C.; Wang, X.; Zou, J.; Hua, H.; Bi, Z. Long noncoding RNA FGFR3-AS1 promotes osteosarcoma growth through regulating its natural antisense transcript FGFR3. Mol. Biol. Rep. 2016, 43, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Nakagawa, S.; Freier, S.M.; Fei, J.; Ha, T.; Prasanth, S.G.; Prasanth, K.V. Natural antisense RNA promotes 3′ end processing and maturation of MALAT1 lncRNA. Nucleic Acids Res. 2016, 44, 2898–2908. [Google Scholar] [CrossRef] [PubMed]
- Beltran, M.; Puig, I.; Peña, C.; García, J.M.; Alvarez, A.B.; Peña, R.; Bonilla, F.; de Herreros, A.G. A natural antisense transcript regulates Zeb2/Sip1 gene expression during Snail1-induced epithelial-mesenchymal transition. Genes Dev. 2008, 22, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and regulation of RNA editing by ADAR deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Carmichael, G.G. The fate of dsRNA in the nucleus: A p54(nrb)-containing complex mediates the nuclear retention of promiscuously A-to-I edited RNAs. Cell 2001, 106, 465–475. [Google Scholar] [CrossRef]
- Levanon, E.Y.; Eisenberg, E.; Yelin, R.; Nemzer, S.; Hallegger, M.; Shemesh, R.; Fligelman, Z.Y.; Shoshan, A.; Pollock, S.R.; Sztybel, D.; et al. Systematic identification of abundant A-to-I editing sites in the human transcriptome. Nat. Biotechnol. 2004, 22, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, G.; Li, J.B. RADAR: A rigorously annotated database of A-to-I RNA editing. Nucleic Acids Res. 2014, 42, D109–D113. [Google Scholar] [CrossRef] [PubMed]
- Neeman, Y.; Dahary, D.; Levanon, E.Y.; Sorek, R.; Eisenberg, E. Is there any sense in antisense editing? Trends Genet. 2005, 21, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Salameh, A.; Lee, A.K.; Cardó-Vila, M.; Nunes, D.N.; Efstathiou, E.; Staquicini, F.I.; Dobroff, A.S.; Marchiò, S.; Navone, N.M.; Hosoya, H.; et al. PRUNE2 is a human prostate cancer suppressor regulated by the intronic long noncoding RNA PCA3. Proc. Natl. Acad. Sci. USA 2015, 112, 8403–8408. [Google Scholar] [CrossRef] [PubMed]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Czech, B.; Hannon, G.J. Small RNA sorting: Matchmaking for Argonautes. Nat. Rev. Genet. 2011, 12, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Polikepahad, S.; Corry, D.B. Profiling of T helper cell-derived small RNAs reveals unique antisense transcripts and differential association of miRNAs with argonaute proteins 1 and 2. Nucleic Acids Res. 2013, 41, 1164–1177. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Cockell, S.; Falconer, J.; Carlile, M.; Alnumeir, S.; Robinson, J. Contribution of natural antisense transcription to an endogenous siRNA signature in human cells. BMC Genom. 2014, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Meng, Y.; Zuo, Z.; Xue, J.; Wang, H. NATpipe: An integrative pipeline for systematical discovery of natural antisense transcripts (NATs) and phase-distributed nat-siRNAs from de novo assembled transcriptomes. Sci. Rep. 2016, 6, 21666. [Google Scholar] [CrossRef] [PubMed]
- Beckedorff, F.C.; Ayupe, A.C.; Crocci-Souza, R.; Amaral, M.S.; Nakaya, H.I.; Soltys, D.T.; Menck, C.F.M.; Reis, E.M.; Verjovski-Almeida, S. The intronic long noncoding RNA ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the expression of RASSF1A and increasing cell proliferation. PLoS Genet. 2013, 9, e1003705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, Y.; Tang, J.; Li, S.; Li, L.; Tang, X.; Cheng, C.; Luo, Y.; Qian, X.; Deng, L.-M.; Liu, L.; et al. LncRNA PANDAR regulates the G1/S transition of breast cancer cells by suppressing p16(INK4A) expression. Sci. Rep. 2016, 6, 22366. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lv, F.; Liang, D.; Yang, Q.; Zhang, B.; Lin, H.; Wang, X.; Qian, G.; Xu, J.; You, W. HOTAIR may regulate proliferation, apoptosis, migration and invasion of MCF-7 cells through regulating the P53/Akt/JNK signaling pathway. Biomed. Pharmacother. 2017, 90, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Y.; Yang, B. Downregulated long non-coding RNA MEG3 in breast cancer regulates proliferation, migration and invasion by depending on p53’s transcriptional activity. Biochem. Biophys. Res. Commun. 2016, 478, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Y.; Yu, M.-S.; Li, X.; Zhang, Z.; Han, C.-R.; Yan, B. Overexpression of long non-coding RNA MEG3 suppresses breast cancer cell proliferation, invasion, and angiogenesis through AKT pathway. Tumour. Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long non-coding RNA HOTAIR regulates the proliferation, self-renewal capacity, tumor formation and migration of the cancer stem-like cell (CSC) subpopulation enriched from breast cancer cells. PLoS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef] [PubMed]
- Jene-Sanz, A.; Váraljai, R.; Vilkova, A.V.; Khramtsova, G.F.; Khramtsov, A.I.; Olopade, O.I.; Lopez-Bigas, N.; Benevolenskaya, E.V. Expression of polycomb targets predicts breast cancer prognosis. Mol. Cell. Biol. 2013, 33, 3951–3961. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Liu, Y.; Wei, H.-Y.; Lv, K.-Z.; Fu, P. Linc-ROR induces epithelial-mesenchymal transition and contributes to drug resistance and invasion of breast cancer cells. Tumour. Biol. 2016, 37, 10861–10870. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ye, X.-L.; Xu, J.; Cao, M.-G.; Fang, Z.-Y.; Li, L.-Y.; Guan, G.-H.; Liu, Q.; Qian, Y.-H.; Xie, D. The lncRNA H19 mediates breast cancer cell plasticity during EMT and MET plasticity by differentially sponging miR-200b/c and let-7b. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Zhao, Y.; Li, Z.; Yao, R.; Ma, M.; Gao, Y.; Zhao, L.; Zhang, Y.; Huang, B.; Lu, J. LincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014, 5, e1287. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Zhou, E.; Xu, F.; Zhang, D.; Yi, W.; Yao, J. A TP73-AS1/miR-200a/ZEB1 regulating loop promotes breast cancer cell invasion and migration. J. Cell. Biochem. 2017. [CrossRef] [PubMed]
- Chi, Y.; Huang, S.; Yuan, L.; Liu, M.; Huang, N.; Zhou, S.; Zhou, B.; Wu, J. Role of BC040587 as a predictor of poor outcome in breast cancer. Cancer Cell Int. 2014, 14, 123. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Lin, C.; Yang, L. Unraveling the therapeutic potential of the LncRNA-dependent noncanonical Hedgehog pathway in cancer. Mol. Cell. Oncol. 2015, 2, e998900. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Park, P.K.; Lin, C.; Yang, L. LncRNA BCAR4 wires up signaling transduction in breast cancer. RNA Biol. 2015, 12, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Booy, E.P.; McRae, E.K.; Koul, A.; Lin, F.; McKenna, S.A. The long non-coding RNA BC200 (BCYRN1) is critical for cancer cell survival and proliferation. Mol. Cancer 2017, 16, 109. [Google Scholar] [CrossRef] [PubMed]
- Redis, R.S.; Sieuwerts, A.M.; Look, M.P.; Tudoran, O.; Ivan, C.; Spizzo, R.; Zhang, X.; de Weerd, V.; Shimizu, M.; Ling, H.; et al. CCAT2, a novel long non-coding RNA in breast cancer: Expression study and clinical correlations. Oncotarget 2013, 4, 1748–1762. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhao, Y.; Wu, X.; Song, G. Upregulation of CCAT2 promotes cell proliferation by repressing the P15 in breast cancer. Biomed. Pharmacother. 2017, 91, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-J.; Li, Y.; Wu, Y.-Z.; Wang, Y.; Nian, W.-Q.; Wang, L.-L.; Li, L.-C.; Luo, H.-L.; Wang, D.-L. Long non-coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF-beta signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 706–714. [Google Scholar] [PubMed]
- Cai, Y.; He, J.; Zhang, D. Long noncoding RNA CCAT2 promotes breast tumor growth by regulating the Wnt signaling pathway. OncoTargets Ther. 2015, 8, 2657–2664. [Google Scholar] [CrossRef]
- Sarrafzadeh, S.; Geranpayeh, L.; Tasharrofi, B.; Soudyab, M.; Nikpayam, E.; Iranpour, M.; Mirfakhraie, R.; Gharesouran, J.; Ghafouri-Fard, S.; Ghafouri-Fard, S. Expression study and clinical correlations of MYC and CCAT2 in breast cancer patients. Iran. Biomed. J. 2017, 21, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Huan, J.; Xing, L.; Lin, Q.; Xui, H.; Qin, X. Long noncoding RNA CRNDE activates Wnt/beta-catenin signaling pathway through acting as a molecular sponge of microRNA-136 in human breast cancer. Am. J. Transl. Res. 2017, 9, 1977–1989. [Google Scholar] [PubMed]
- Sha, S.; Yuan, D.; Liu, Y.; Han, B.; Zhong, N. Targeting long non-coding RNA DANCR inhibits triple negative breast cancer progression. Biol. Open 2017, 6, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Miano, V.; Ferrero, G.; Reineri, S.; Caizzi, L.; Annaratone, L.; Ricci, L.; Cutrupi, S.; Castellano, I.; Cordero, F.; de Bortoli, M. Luminal long non-coding RNAs regulated by estrogen receptor alpha in a ligand-independent manner show functional roles in breast cancer. Oncotarget 2015, 7, 3201–3216. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, Y.S.; Han, S.; Ma, T.; Speers, C.; Zhang, C.; Wilder-Romans, K.; Iyer, M.K.; Pitchiaya, S.; Malik, R.; Hosono, Y.; et al. The lncRNA landscape of breast cancer reveals a role for DSCAM-AS1 in breast cancer progression. Nat. Commun. 2016, 7, 12791. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Lv, S.-X.; Lv, L.; Liu, Y.-H.; Dong, S.-Y.; Yao, Z.-H.; Dai, X.-X.; Zhang, X.-H.; Wang, O.-C. Identification of lncRNA FAM83H-AS1 as a novel prognostic marker in luminal subtype breast cancer. OncoTargets Ther. 2016, 9, 7039–7045. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Song, W.-Q.; Sun, P.; Jin, L.; Dai, H.-Y. LncRNA-GAS5 induces PTEN expression through inhibiting miR-103 in endometrial cancer cells. J. Biomed. Sci. 2015, 22, 100. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, Z. Long non-coding RNA growth arrest-specific transcript 5 in tumor biology. Oncol. Lett. 2015, 10, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Pickard, M.R.; Williams, G.T. The hormone response element mimic sequence of GAS5 lncRNA is sufficient to induce apoptosis in breast cancer cells. Oncotarget 2016, 7, 10104–10116. [Google Scholar] [CrossRef] [PubMed]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhai, L.; Wang, H.; Liu, C.; Zhang, J.; Chen, W.; Wei, Q. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget 2016, 7, 27778–27786. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Ma, P.; Liu, S.-M.; Zhou, X. Circulating long noncoding RNA GAS5 as a potential biomarker in breast cancer for assessing the surgical effects. Tumour. Biol. 2016, 37, 6847–6854. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Y.; Li, Y.; Ren, K.; Li, X.; Han, X.; Wang, J. Long non-coding RNA H19 promotes the proliferation and invasion of breast cancer through upregulating DNMT1 expression by sponging miR-152. J. Biochem. Mol. Toxicol. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Zang, R.; Zhang, E.; Liu, Y.; Shi, X.; Zhang, E.; Shao, L.; Li, A.; Yang, N.; Han, X.; et al. LncRNA H19 confers chemoresistance in ERalpha-positive breast cancer through epigenetic silencing of the pro-apoptotic gene BIK. Oncotarget 2016, 7, 81452–81462. [Google Scholar] [CrossRef] [PubMed]
- Vennin, C.; Spruyt, N.; Dahmani, F.; Julien, S.; Bertucci, F.; Finetti, P.; Chassat, T.; Bourette, R.P.; Le Bourhis, X.; Adriaenssens, E. H19 non coding RNA-derived miR-675 enhances tumorigenesis and metastasis of breast cancer cells by downregulating c-Cbl and Cbl-b. Oncotarget 2015, 6, 29209–29223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Luo, Z.; Zhang, Y.; Zhang, L.; Wu, L.; Liu, L.; Yang, J.; Song, X.; Liu, J. Circulating lncRNA H19 in plasma as a novel biomarker for breast cancer. Cancer Biomark. 2016, 17, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-Z.; Liu, Y.-R.; Xu, X.-E.; Jin, X.; Hu, X.; Yu, K.-D.; Shao, Z.-M. Transcriptome analysis of triple-negative breast cancer reveals an integrated mRNA-lncRNA signature with predictive and prognostic value. Cancer Res. 2016, 76, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xing, L.-Q.; Liu, Y.-J. A three-long noncoding RNA signature as a diagnostic biomarker for differentiating between triple-negative and non-triple-negative breast cancers. Medicine 2017, 96, e6222. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ma, J.; Cheng, Y.; Zhang, H.; Luo, W.; Zhang, H. HMMR antisense RNA 1, a novel long noncoding RNA, regulates the progression of basal-like breast cancer cells. Breast Cancer 2016, 8, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Hussain, I.; Ansari, K.I.; Kasiri, S.; Bashyal, A.; Mandal, S.S. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J. Mol. Biol. 2013, 425, 3707–3722. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Kogo, R.; Shimamura, T.; Mimori, K.; Kawahara, K.; Imoto, S.; Sudo, T.; Tanaka, F.; Shibata, K.; Suzuki, A.; Komune, S.; et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011, 71, 6320–6326. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Abdelmohsen, K.; Kim, J.; Yang, X.; Martindale, J.L.; Tominaga-Yamanaka, K.; White, E.J.; Orjalo, A.V.; Rinn, J.L.; Kreft, S.G.; et al. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat. Commun. 2013, 4, 2939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.-J.; Zhang, Y.; Luo, Z.-L.; Liu, L.; Yang, J.; Wu, L.-C.; Yu, S.-S.; Liu, J.-B. Long non-coding RNA HOTAIR in plasma as a potential biomarker for breast cancer diagnosis. Nan Fang Yi Ke Da Xue Xue Bao 2016, 36, 488–492. [Google Scholar] [PubMed]
- Ozes, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.D.; Dostie, J. Reciprocal regulation of chromatin state and architecture by HOTAIRM1 contributes to temporal collinear HOXA gene activation. Nucleic Acids Res. 2017, 45, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Malouf, G.G.; Chen, Y.; Zhang, J.; Yao, H.; Valero, V.; Weinstein, J.N.; Spano, J.-P.; Meric-Bernstam, F.; Khayat, D.; et al. Comprehensive analysis of long non-coding RNAs in human breast cancer clinical subtypes. Oncotarget 2014, 5, 9864–9876. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, J.; Wu, F.; Song, Y.; Zhao, S.; Zhang, Q. Long non-coding RNA HOXA-AS2 promotes proliferation and invasion of breast cancer by acting as a miR-520c-3p sponge. Oncotarget 2017, 8, 46090–46103. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jia, G.; Qu, Y.; Du, Q.; Liu, B.; Liu, B. Long Non-Coding RNA (LncRNA) HOXA11-AS Promotes Breast Cancer Invasion and Metastasis by Regulating Epithelial-Mesenchymal Transition. Med. Sci. Monit. 2017, 23, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-X.; Sha, R.-L.; Bao, J.-Q.; Luan, W.; Su, R.-L.; Sun, S.-R. Expression of long non-coding RNA linc-ITGB1 in breast cancer and its influence on prognosis and survival. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 3397–3401. [Google Scholar] [PubMed]
- Yan, M.; Zhang, L.; Li, G.; Xiao, S.; Dai, J.; Cen, X. Long noncoding RNA linc-ITGB1 promotes cell migration and invasion in human breast cancer. Biotechnol. Appl. Biochem. 2017, 64, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, C.; Chen, J.; Zhang, K.; Chu, X.; Wang, R.; Chen, L. The Emerging Roles of Long Noncoding RNA ROR (lincRNA-ROR) and its Possible Mechanisms in Human Cancers. Cell. Physiol. Biochem. 2016, 40, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Katsaros, D.; Loo, L.W.M.; Hernandez, B.Y.; Chong, C.; Canuto, E.M.; Biglia, N.; Lu, L.; Risch, H.; Chu, W.-M.; et al. Prognostic and predictive values of long non-coding RNA LINC00472 in breast cancer. Oncotarget 2015, 6, 8579–8592. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wang, Z.; Loo, L.W.M.; Ni, Y.; Jia, W.; Fei, P.; Risch, H.A.; Katsaros, D.; Yu, H. LINC00472 expression is regulated by promoter methylation and associated with disease-free survival in patients with grade 2 breast cancer. Breast Cancer Res. Treat. 2015, 154, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.; Boczek, N.J.; Berres, M.W.; Ma, X.; Smith, D.I. LSINCT5 is over expressed in breast and ovarian cancer and affects cellular proliferation. RNA Biol. 2011, 8, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Jadaliha, M.; Zong, X.; Malakar, P.; Ray, T.; Singh, D.K.; Freier, S.M.; Jensen, T.; Prasanth, S.G.; Karni, R.; Ray, P.S.; et al. Functional and prognostic significance of long non-coding RNA MALAT1 as a metastasis driver in ER negative lymph node negative breast cancer. Oncotarget 2016, 7, 40418–40436. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Hammerle, M.; Diederichs, S. MALAT1—A paradigm for long noncoding RNA function in cancer. J. Mol. Med. 2013, 91, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.-S.; Chi, Y.-Y.; Xue, J.-Y.; Liu, M.-Y.; Huang, S.; Mo, M.; Zhou, S.-L.; Wu, J. Long non-coding RNA metastasis associated in lung adenocarcinoma transcript 1 (MALAT1) interacts with estrogen receptor and predicted poor survival in breast cancer. Oncotarget 2016, 7, 37957–37965. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Fan, R.; Chen, L.; Qian, H. Clinical Significance of Long Non-coding RNA MALAT1 Expression in Tissue and Serum of Breast Cancer. Ann. Clin. Lab. Sci. 2016, 46, 418–424. [Google Scholar] [PubMed]
- Zhang, W.; Shi, S.; Jiang, J.; Li, X.; Lu, H.; Ren, F. LncRNA MEG3 inhibits cell epithelial-mesenchymal transition by sponging miR-421 targeting E-cadherin in breast cancer. Biomed. Pharmacother. 2017, 91, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA-DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef] [PubMed]
- Nie, F.-Q.; Ma, S.; Xie, M.; Liu, Y.-W.; De, W.; Liu, X.-H. Decreased long noncoding RNA MIR31HG is correlated with poor prognosis and contributes to cell proliferation in gastric cancer. Tumour. Biol. 2016, 37, 7693–7701. [Google Scholar] [CrossRef] [PubMed]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. miR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lu, B.; Wang, J.; Wang, J.; Shi, Y.; Lian, Y.; Zhu, Y.; Wang, J.; Fan, Y.; Wang, Z.; et al. Long non-coding RNA Loc554202 induces apoptosis in colorectal cancer cells via the caspase cleavage cascades. J. Exp. Clin. Cancer Res. 2015, 34, 100. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, Z.; Liu, X.; Cheng, X.; Zhang, Y.; Han, X.; Zhang, Y.; Liu, S.; Yang, J.; Xu, B.; et al. The FOXN3-NEAT1-SIN3A repressor complex promotes progression of hormonally responsive breast cancer. J. Clin. Investig. 2017, 127, 3421–3440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, W.-B.; Wang, Z.-W.; Wang, X.-H. lncRNA NEAT1 is closely related with progression of breast cancer via promoting proliferation and EMT. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1020–1026. [Google Scholar] [PubMed]
- Hirose, T.; Virnicchi, G.; Tanigawa, A.; Naganuma, T.; Li, R.; Kimura, H.; Yokoi, T.; Nakagawa, S.; Benard, M.; Fox, A.H.; et al. NEAT1 long noncoding RNA regulates transcription via protein sequestration within subnuclear bodies. Mol. Biol. Cell 2014, 25, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Qian, K.; Liu, G.; Tang, Z.; Hu, Y.; Fang, Y.; Chen, Z.; Xu, X. The long non-coding RNA NEAT1 interacted with miR-101 modulates breast cancer growth by targeting EZH2. Arch. Biochem. Biophys. 2017, 615, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shao, C.; Wu, Q.-J.; Chen, G.; Zhou, J.; Yang, B.; Li, H.; Gou, L.-T.; Zhang, Y.; Wang, Y.; et al. NEAT1 scaffolds RNA-binding proteins and the Microprocessor to globally enhance pri-miRNA processing. Nat. Struct. Mol. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Zhao, L.; Feng, X.; Xu, H.; Zou, L.; Yang, Q.; Su, X.; Peng, L.; Jiao, B. NEAT1 is Required for Survival of Breast Cancer Cells Through FUS and miR-548. Gene Regul. Syst. Biol. 2016, 10, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, M.; Soudyab, M.; Geranpayeh, L.; Mirfakhraie, R.; Azargashb, E.; Movafagh, A.; Ghafouri-Fard, S. Expression analysis of four long noncoding RNAs in breast cancer. Tumour. Biol. 2016, 37, 2933–2940. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Luo, J.; Jiao, S. Comprehensive characterization of cancer subtype associated long non-coding RNAs and their clinical implications. Sci. Rep. 2014, 4, 6591. [Google Scholar] [CrossRef] [PubMed]
- Colombo, T.; Farina, L.; Macino, G.; Paci, P. PVT1: A rising star among oncogenic long noncoding RNAs. BioMed Res. Int. 2015, 2015, 304208. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-Y.; Bagchi, A. The PVT1-MYC duet in cancer. Mol. Cell. Oncol. 2015, 2, e974467. [Google Scholar] [CrossRef] [PubMed]
- Sarver, A.L.; Murray, C.D.; Temiz, N.A.; Tseng, Y.-Y.; Bagchi, A. MYC and PVT1 synergize to regulate RSPO1 levels in breast cancer. Cell Cycle 2016, 15, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Shahryari, A.; Jazi, M.S.; Samaei, N.M.; Mowla, S.J. Long non-coding RNA SOX2OT: Expression signature, splicing patterns, and emerging roles in pluripotency and tumorigenesis. Front. Genet. 2015, 6, 196. [Google Scholar] [CrossRef] [PubMed]
- Askarian-Amiri, M.E.; Seyfoddin, V.; Smart, C.E.; Wang, J.; Kim, J.E.; Hansji, H.; Baguley, B.C.; Finlay, G.J.; Leung, E.Y. Emerging role of long non-coding RNA SOX2OT in SOX2 regulation in breast cancer. PLoS ONE 2014, 9, e102140. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Li, J.; Liu, Y.; Ding, J.; Fan, Y.; Tian, Y.; Wang, L.; Lian, Y.; Wang, K.; Shu, Y. The long noncoding RNA SPRY4-IT1 increases the proliferation of human breast cancer cells by upregulating ZNF703 expression. Mol. Cancer 2015, 14, 51. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Y.; Chen, Z.; He, A.; Xie, H.; Zhang, Q.; Cai, Z.; Liu, Y.; Huang, W. SPRY4-IT1: A novel oncogenic long non-coding RNA in human cancers. Tumour. Biol. 2017, 39, 1010428317711406. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Chartrand, P. Telomeric repeat-containing RNA TERRA: A noncoding RNA connecting telomere biology to genome integrity. Front. Genet. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Lingner, J. TERRA: Telomeric repeat-containing RNA. EMBO J. 2009, 28, 2503–2510. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Xu, F.; Zhang, D.; Yi, W.; Chen, X.; Chen, G.; Zhou, E. TP73-AS1 promotes breast cancer cell proliferation through miR-200a-mediated TFAM inhibition. J. Cell. Biochem. 2017. [CrossRef] [PubMed]
- Gumireddy, K.; Li, A.; Yan, J.; Setoyama, T.; Johannes, G.J.; Orom, U.A.; Tchou, J.; Liu, Q.; Zhang, L.; Speicher, D.W.; et al. Identification of a long non-coding RNA-associated RNP complex regulating metastasis at the translational step. EMBO J. 2013, 32, 2672–2684. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhou, N.; Watabe, K.; Lu, Z.; Wu, F.; Xu, M.; Mo, Y.-Y. Long non-coding RNA UCA1 promotes breast tumor growth by suppression of p27 (Kip1). Cell Death Dis. 2014, 5, e1008. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.-L.; Li, X.-M.; Luo, J. Long noncoding RNA UCA1 modulates breast cancer cell growth and apoptosis through decreasing tumor suppressive miR-143. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3403–3411. [Google Scholar] [PubMed]
- Xiao, C.; Wu, C.-H.; Hu, H.-Z. LncRNA UCA1 promotes epithelial-mesenchymal transition (EMT) of breast cancer cells via enhancing Wnt/beta-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2819–2824. [Google Scholar] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. Long non-coding RNA UCA1 enhances tamoxifen resistance in breast cancer cells through a miR-18a-HIF1alpha feedback regulatory loop. Tumour. Biol. 2016, 37, 14733–14743. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Luo, J. Long Non-Coding RNA (lncRNA) Urothelial Carcinoma-Associated 1 (UCA1) Enhances Tamoxifen Resistance in Breast Cancer Cells via Inhibiting mTOR Signaling Pathway. Med. Sci. Monit. 2016, 22, 3860–3867. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.-M.; Meng, F.-Q.; Wang, X.-B. Overexpression of long-noncoding RNA ZFAS1 decreases survival in human NSCLC patients. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5126–5131. [Google Scholar] [PubMed]
- Askarian-Amiri, M.E.; Crawford, J.; French, J.D.; Smart, C.E.; Smith, M.A.; Clark, M.B.; Ru, K.; Mercer, T.R.; Thompson, E.R.; Lakhani, S.R.; et al. SNORD-host RNA Zfas1 is a regulator of mammary development and a potential marker for breast cancer. RNA 2011, 17, 878–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Xing, C. Upregulation of long noncoding RNA ZFAS1 predicts poor prognosis and prompts invasion and metastasis in colorectal cancer. Pathol. Res. Pract. 2016, 212, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, F.; Chen, H.; Tan, Q.; Qiu, S.; Chen, S.; Jing, W.; Yu, M.; Liang, C.; Ye, S.; et al. Increased expression of long-noncoding RNA ZFAS1 is associated with epithelial-mesenchymal transition of gastric cancer. Aging 2016, 8, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Hansji, H.; Leung, E.Y.; Baguley, B.C.; Finlay, G.J.; Cameron-Smith, D.; Figueiredo, V.C.; Askarian-Amiri, M.E. ZFAS1: A long noncoding RNA associated with ribosomes in breast cancer cells. Biol. Direct 2016, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.; Shak, S.; Tang, G.; Kim, C.; Baker, J.; Cronin, M.; Baehner, F.L.; Walker, M.G.; Watson, D.; Park, T.; et al. A multigene assay to predict recurrence of tamoxifen-treated, node-negative breast cancer. N. Engl. J. Med. 2004, 351, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Gnant, M.; Sestak, I.; Filipits, M.; Dowsett, M.; Balic, M.; Lopez-Knowles, E.; Greil, R.; Dubsky, P.; Stoeger, H.; Rudas, M.; et al. Identifying clinically relevant prognostic subgroups of postmenopausal women with node-positive hormone receptor-positive early-stage breast cancer treated with endocrine therapy: A combined analysis of ABCSG-8 and ATAC using the PAM50 risk of recurrence. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2015, 26, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Mook, S.; Schmidt, M.K.; Viale, G.; Pruneri, G.; Eekhout, I.; Floore, A.; Glas, A.M.; Bogaerts, J.; Cardoso, F.; Piccart-Gebhart, M.J.; et al. The 70-gene prognosis-signature predicts disease outcome in breast cancer patients with 1-3 positive lymph nodes in an independent validation study. Breast Cancer Res. Treat. 2009, 116, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, Q.; Zhan, Y.; Chen, X.; Yu, Q.; Zhang, J.; Wang, Y.; Xu, X.-J.; Zhu, L. Transcriptome sequencing uncovers a three-long noncoding RNA signature in predicting breast cancer survival. Sci. Rep. 2016, 6, 27931. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhong, L.; Xu, W.; Sun, Y.; Zhang, Z.; Zhao, H.; Yang, L.; Sun, J. Discovery of potential prognostic long non-coding RNA biomarkers for predicting the risk of tumor recurrence of breast cancer patients. Sci. Rep. 2016, 6, 31038. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, J.; Koirala, P.; Ding, X.; Chen, B.; Wang, Y.; Wang, Z.; Wang, C.; Zhang, X.; Mo, Y.-Y. Long non-coding RNAs as prognostic markers in human breast cancer. Oncotarget 2016, 7, 20584–20596. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-R.; Jiang, Y.-Z.; Xu, X.-E.; Hu, X.; Yu, K.-D.; Shao, Z.-M. Comprehensive Transcriptome Profiling Reveals Multigene Signatures in Triple-Negative Breast Cancer. Clin. Cancer Res. 2016, 22, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Wahlestedt, C. Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 2013, 12, 433–446. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.R.; Crooke, S.T. RNA Therapeutics in Oncology: Advances, Challenges, and Future Directions. J. Clin. Pharmacol. 2017, 57, S43–S59. [Google Scholar] [CrossRef] [PubMed]
- Modarresi, F.; Faghihi, M.A.; Lopez-Toledano, M.A.; Fatemi, R.P.; Magistri, M.; Brothers, S.P.; van der Brug, M.P.; Wahlestedt, C. Inhibition of natural antisense transcripts in vivo results in gene-specific transcriptional upregulation. Nat. Biotechnol. 2012, 30, 453–459. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
lncRNA classification according to their orientation and position in the genome. lincRNAs are located between two pcGenes, regardless of their orientation. Intronic lncRNAs are entirely encoded in pcGene introns, while sense lncRNAs overlap pcGene exons. Bidirectional lncRNA transcription starts less than 1 kb from a pcGene transcription start site and goes in its opposite direction. Cis-NATs (natural antisense transcript) are RNA sequences that are transcribed from the two strands of the same genomic locus, in the antisense direction. NAT pairs can be protein-coding sequences (pc, red colored) or non-coding sequences (nc, blue colored), forming nc|pc, nc|nc or pc|pc pairs. NAT pairs that are nc|pc or nc|nc sequences only belong to the lncRNA classification (purple colored sequences are pc or nc).
Figure 1.
lncRNA classification according to their orientation and position in the genome. lincRNAs are located between two pcGenes, regardless of their orientation. Intronic lncRNAs are entirely encoded in pcGene introns, while sense lncRNAs overlap pcGene exons. Bidirectional lncRNA transcription starts less than 1 kb from a pcGene transcription start site and goes in its opposite direction. Cis-NATs (natural antisense transcript) are RNA sequences that are transcribed from the two strands of the same genomic locus, in the antisense direction. NAT pairs can be protein-coding sequences (pc, red colored) or non-coding sequences (nc, blue colored), forming nc|pc, nc|nc or pc|pc pairs. NAT pairs that are nc|pc or nc|nc sequences only belong to the lncRNA classification (purple colored sequences are pc or nc).
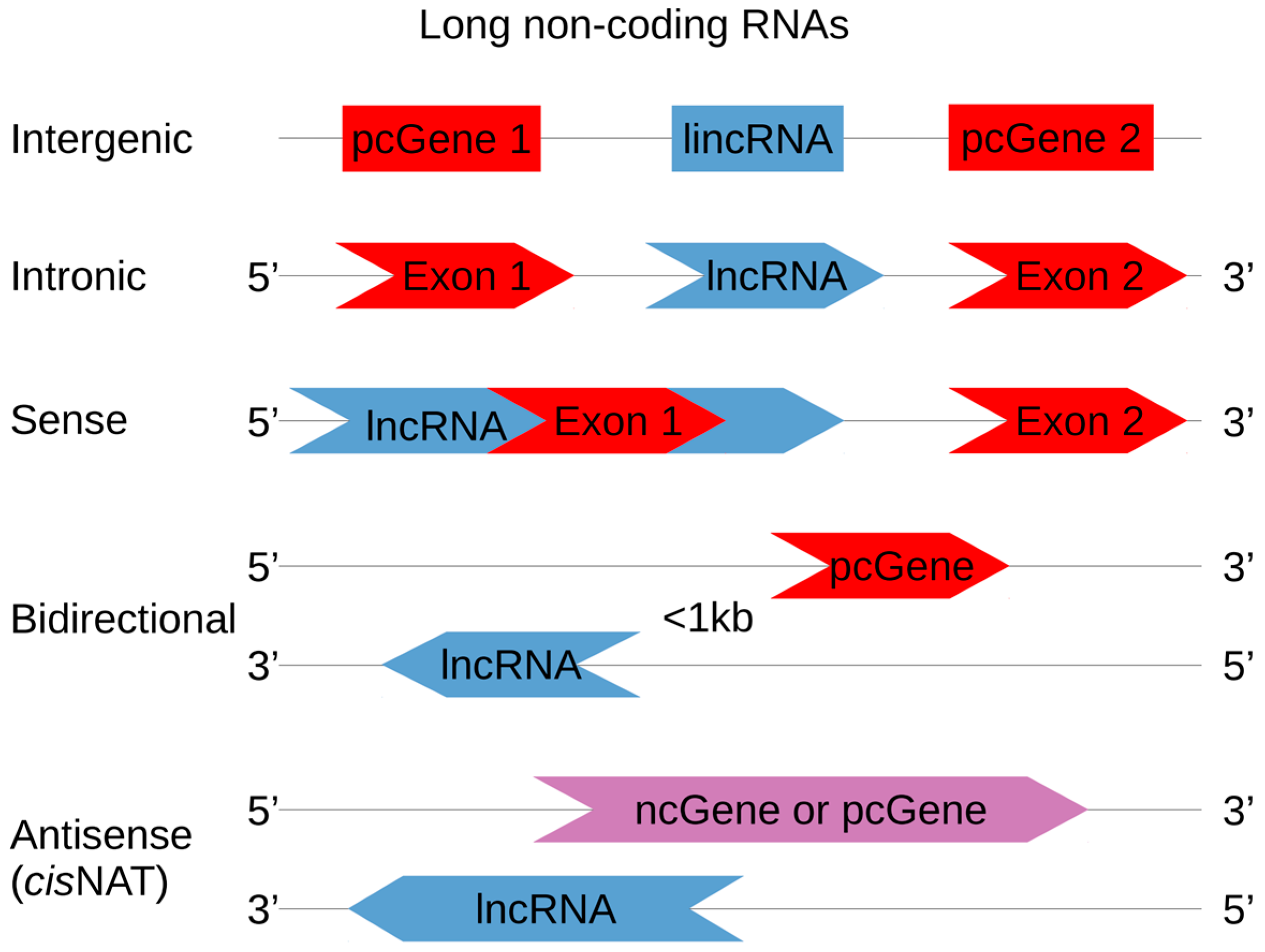
Figure 2.
cis-NAT classification. cisNAT pairs can be protein coding sequences (pc) or non-coding sequences (nc), forming nc|pc, nc|nc or pc|pc pairs. In head-to-head orientation, sense and antisense transcripts overlap on their 5′ ends. Inversely, tail-to-tail describes an overlap of the 3′ ends. In a full overlap (or embedded overlap), one transcript is totally included in the other one.
Figure 2.
cis-NAT classification. cisNAT pairs can be protein coding sequences (pc) or non-coding sequences (nc), forming nc|pc, nc|nc or pc|pc pairs. In head-to-head orientation, sense and antisense transcripts overlap on their 5′ ends. Inversely, tail-to-tail describes an overlap of the 3′ ends. In a full overlap (or embedded overlap), one transcript is totally included in the other one.
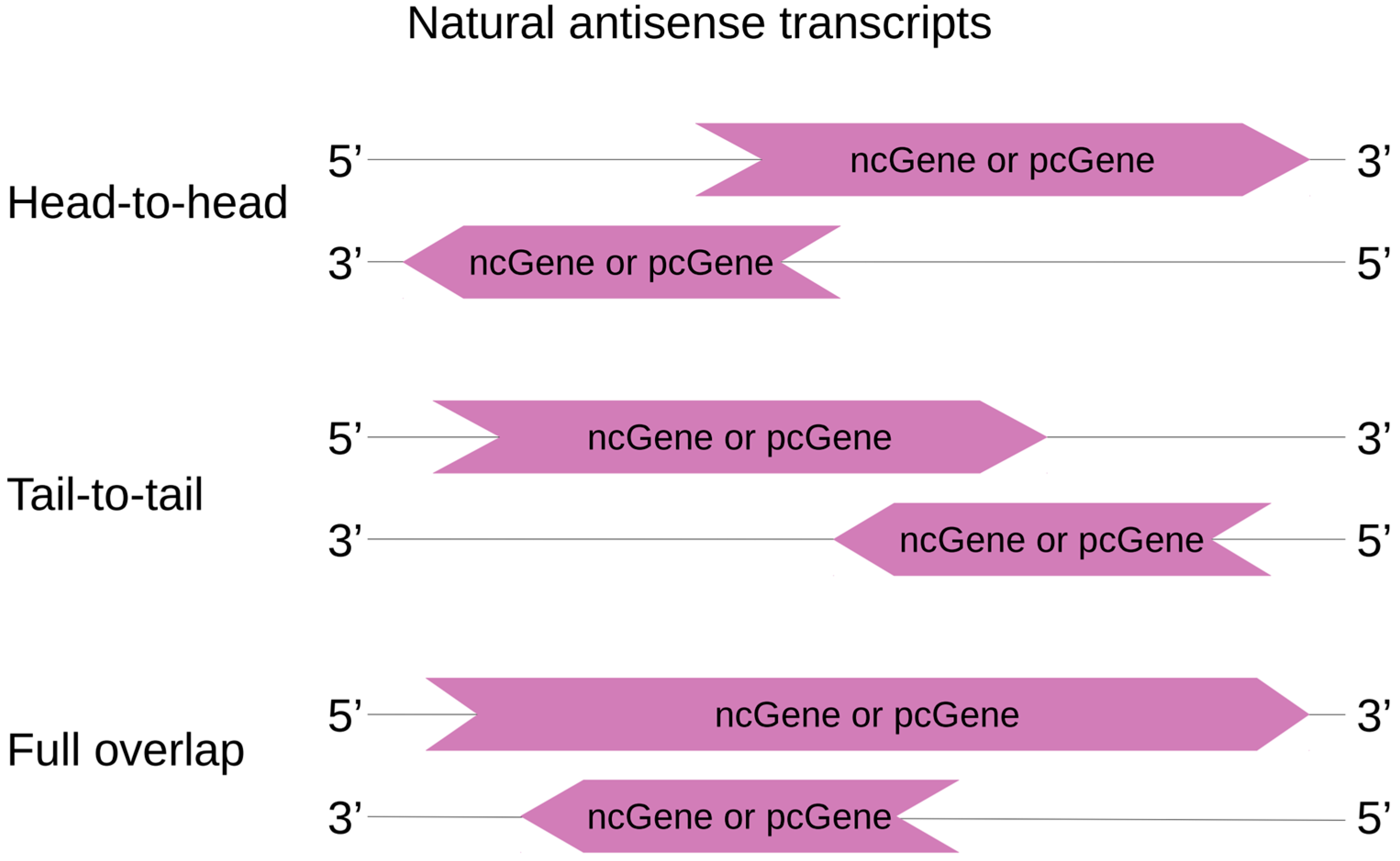
Figure 3.
Transcriptional Interference: (A) in the initiation phase, promoters of head-to-head NATs are competing for the use of RNA Pol II and common regulatory elements; (B) in the elongation phase, interference can occur after the following events: a collision between RNA Pol II complexes, leading to a machinery blockage; (C) a promoter occlusion by RNA Pol II during the antisense transcript; and (D) a RNA Pol II dislodgement by the RNA Pol II standing on the opposite strand, when the first one was too slow to start. Promoters of protein coding sequences are represented in red, and promoters of non-coding sequences in blue. RNA pol II enzyme is represented in dark grey when able to transcribe the sequence, and light grey when its binding and thus activity, is prevented.
Figure 3.
Transcriptional Interference: (A) in the initiation phase, promoters of head-to-head NATs are competing for the use of RNA Pol II and common regulatory elements; (B) in the elongation phase, interference can occur after the following events: a collision between RNA Pol II complexes, leading to a machinery blockage; (C) a promoter occlusion by RNA Pol II during the antisense transcript; and (D) a RNA Pol II dislodgement by the RNA Pol II standing on the opposite strand, when the first one was too slow to start. Promoters of protein coding sequences are represented in red, and promoters of non-coding sequences in blue. RNA pol II enzyme is represented in dark grey when able to transcribe the sequence, and light grey when its binding and thus activity, is prevented.
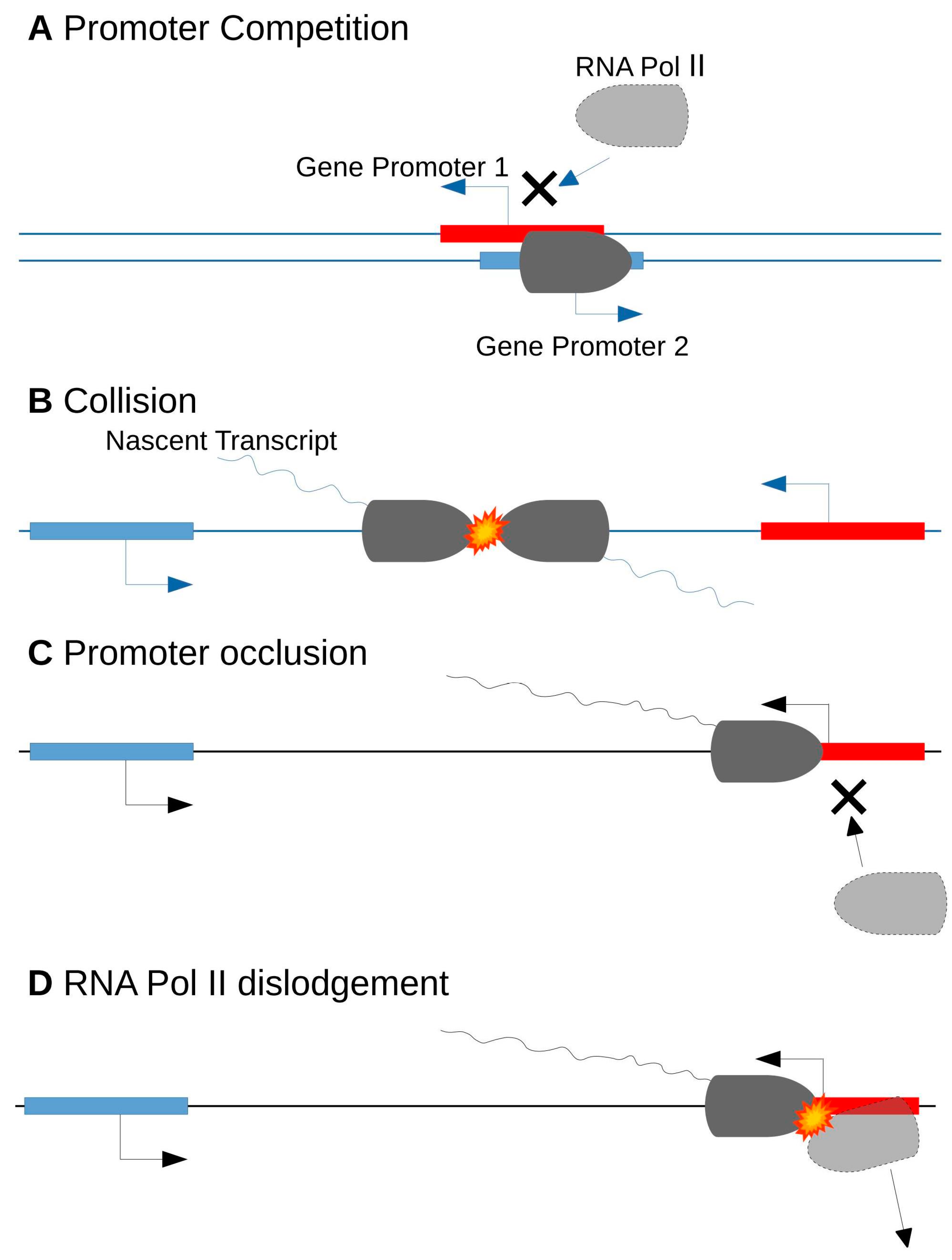
Figure 4.
ncNATs (non-coding natural antisense transcripts) may regulate the expression levels of the sense pcGenes (protein coding genes) by regulating chromatin modifications by the following. (A) A decoy mechanism: The NAT binds a protein complex that can trigger chromatin modifications and prevents, by competition, this complex from binding the sense transcript. This complex can also prevent the interaction of the sense gene with RNA Pol II (RNA polymerase II); (B) a tethering mechanism, such as ANRIL (antisense non-coding RNA in the INK4 locus): ANRIL recruits PRC2 (polycomb repressive complex) through interaction with SUZ12 (suppressor of zeste 12 homolog) and EZH2 (enhancer of zeste 2 polycomb repressive complex 2 subunit) components and PRC1 by binding CBX7 (chromobox homolog 7). Next, PRC2 silences the INK4 locus expression by inducing H3K27 tri-methylation, and PRC1 maintains a repressive chromatin structure by mono-ubiquitination of H2AK119. Protein coding sequences or promoters are represented in red, and non-coding in blue.
Figure 4.
ncNATs (non-coding natural antisense transcripts) may regulate the expression levels of the sense pcGenes (protein coding genes) by regulating chromatin modifications by the following. (A) A decoy mechanism: The NAT binds a protein complex that can trigger chromatin modifications and prevents, by competition, this complex from binding the sense transcript. This complex can also prevent the interaction of the sense gene with RNA Pol II (RNA polymerase II); (B) a tethering mechanism, such as ANRIL (antisense non-coding RNA in the INK4 locus): ANRIL recruits PRC2 (polycomb repressive complex) through interaction with SUZ12 (suppressor of zeste 12 homolog) and EZH2 (enhancer of zeste 2 polycomb repressive complex 2 subunit) components and PRC1 by binding CBX7 (chromobox homolog 7). Next, PRC2 silences the INK4 locus expression by inducing H3K27 tri-methylation, and PRC1 maintains a repressive chromatin structure by mono-ubiquitination of H2AK119. Protein coding sequences or promoters are represented in red, and non-coding in blue.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Role and therapeutic utility of lncRNAs in breast cancer. lncRNAs implicated in breast cancer pathology are listed and classified in different categories: lincRNA (long intergenic non-coding RNA), bidirectional lncRNA, sense-overlapping lncRNA, sense-intronic lncRNA, and NAT composed of nc|nc or nc|pc transcripts pairs. In the case of nc|pc pairs, the pcGene name is provided. As ncRNAs often display multiple transcript variants, some lncRNAs may belong to multiple categories.
Table 1.
Role and therapeutic utility of lncRNAs in breast cancer. lncRNAs implicated in breast cancer pathology are listed and classified in different categories: lincRNA (long intergenic non-coding RNA), bidirectional lncRNA, sense-overlapping lncRNA, sense-intronic lncRNA, and NAT composed of nc|nc or nc|pc transcripts pairs. In the case of nc|pc pairs, the pcGene name is provided. As ncRNAs often display multiple transcript variants, some lncRNAs may belong to multiple categories.
| lncRNA in Breast Cancer | Access Number | Type | pcGene | Alteration/Role in Breast Cancer | Mechanism of Action | Therapeutic Utility | Ref. |
|---|---|---|---|---|---|---|---|
| ANRASSF1 | ENSG00000281358 | NAT (nc|pc) | RASSF1 | Upregulation/Oncogenic | Binds to PRC2 and silences the tumor suppressor gene RASSF1A. | [84] | |
| ANRIL | ENSG00000240498 | NAT (nc|pc) | CDKN2A, CDKN2B | Upregulation/Oncogenic | ANRIL is the NAT of CDKN2B gene (p15); binds to components of PRC1 (CBX7) and PRC2 (SUZ12) to silence the INK4 locus by epigenetic mechanisms. | Overexpressed in a variety of cancers and diseases. | [63,64,65] |
| LINC00901 | ENSG00000242385 | NAT (nc|pc) | LSAMP | Downregulation/Tumor suppressor | Low expression is associated with low overall survival. | Potential prognostic marker. | [95] |
| BCAR4 | ENSG00000262117 | lincRNA | Upregulation/Oncogenic | Interaction with SNIP1 and PNUTS in Hedgehog canonical pathway leads to a resistance to cancer treatments with SMO inhibitors. | Responsible for the acquisition of resistance to treatments and upregulation of non-canonical hedgehog pathway. | [96,97] | |
| BCYRN1 | ENSG00000236824 | NAT (nc|nc) | Upregulation/Oncogenic | BCYRN1 expression is associated with cell proliferation. | Knockdown BCYRN1 impacts viability of actively proliferating cells through growth arrest and apoptosis. Potential therapeutic target for various cancers. | [98] | |
| CCAT2 | ENSG00000280997 | NAT (nc|nc) | Upregulation/Oncogenic | Downregulates p15 through interaction with EZH2. Regulates TGF-β and Wnt signaling pathways. Promotes cell proliferation, invasion, tumor growth and metastasis. | Potential prognosis biomarker and therapeutic target. | [99,100,101,102,103] | |
| CRNDE | ENSG00000245694 | lincRNA | Upregulation/Oncogenic | Molecular sponge of miRNA-136 in breast cancer, activating Wnt/β-catenin. | Associated with unfavorable prognosis. | [104] | |
| DANCR | ENSG00000226950 | lincRNA | Upregulation/Oncogenic | Participates in cell proliferation and invasion. | Associated with a worse prognosis in TNBC. | [105] | |
| DSCAM-AS1 | ENSG00000235123 | NAT (nc|pc) | DSCAM | Upregulation/Oncogenic | Expression induced by estrogen stimulation. Positive correlation with genes associated with cancer aggression, tamoxifen resistance, and metastasis. | Biomarker for luminal subtype. | [106,107] |
| FAM83H-AS1 | ENSG00000282685 | lincRNA | Upregulation/Oncogenic | Most upregulated in luminal subtype of breast cancer. | Prognostic marker of luminal subtype. | [108] | |
| GAS5 | ENSG00000234741 | NAT (nc|pc)/bidirectional lncRNA/lincRNA (multiple transcripts) | ZBTB37 | Downregulation/Tumor suppressor | Required for decoy of glucocorticoid receptor (GR), inhibits transcriptional induction by GR, stops growth and triggers apoptosis, induces PTEN through miR-103 inhibition. | Responsible for the acquisition of trastuzumab resistance. Potential circulating biomarker. | [109,110,111,112,113,114] |
| H19 | ENSG00000130600 | lincRNA | Upregulation/Oncogenic | Mediates breast cancer cell plasticity, invasion, and proliferation by sponging several miR (miR-200b/c, let-7b, miR-152), silences pro-apoptotic gene BIK through epigenetic modifications, precursor of miR-675 (pro-tumoral and pro-metastatic). | Upregulated in cancer. Potential circulating biomarker for early screening and prognosis monitoring in breast cancer. | [92,115,116,117,118] | |
| HIF1A-AS2 | ENSG00000258667 | NAT (nc|pc) | HIF1A | Upregulation/Oncogenic | Involved in cell proliferation and invasion, contributes to chemotherapy resistance. | In TNBC, biomarker for detection, prognosis and prediction for recurrence and response to taxane chemotherapy. | [119,120] |
| HMMR-AS1 | ENSG00000251018 | NAT (nc|pc) | HMMR | Upregulation/Oncogenic | Involved in cell proliferation and invasion. | Positive correlation with HMMR, BRCA1, BRCA2 (oncogenic), biomarker and potential target in basal-like breast cancer. | [121] |
| HOTAIR | ENSG00000228630 | NAT (nc|pc)/lincRNA (multiple transcripts) | HOXC11 | Upregulation/Oncogenic | Guides epigenetic mechanisms to silence tumor suppressor genes through interaction with PRC2 and LSD1, involved in protein degradation by interaction with E3-ubiquitin ligases, tumor invasion, apoptosis and EMT. | Over-expressed in cancer, biomarker and potential therapeutic target. | [86,89,122,123,124,125,126,127] |
| HOTAIRM1 | ENSG00000233429 | NAT (nc|pc)/bidirectional lncRNA (multiple transcripts) | HOXA1, HOXA2 | Upregulation/Oncogenic | Modulates gene expression in HoxA gene cluster by interacting with PRC1 and PRC2 complexes. High positive correlation with HOXA1 expression. | Increased in basal-like subtype breast cancer. | [128,129] |
| HOXA-AS2 | ENSG00000253552 | NAT (nc|pc) | HOXA3, HOXA4 | Upregulation/Oncogenic | Acts as an endogenous sponge of miR-520c-3p and indirectly controls the expression of miR520c-3p target genes (TGFBR2 and RELA). | [130] | |
| HOXA11-AS | ENSG00000240990 | NAT (nc|pc) | HOXA11 | Upregulation/Oncogenic | Promotes cell proliferation, invasion and metastasis by regulating EMT. | Biomarker for metastasis and prognosis in breast cancer. Blocked relation between HOXA11-AS and EMT may have therapeutic utility. | [131] |
| Lnc-ITGB1-6:7 | Lnc-ITGB1-6 (LNCipedia) | lincRNA | Upregulation/Oncogenic | Promotes cell proliferation, invasion and metastasis by regulating EMT. High linc-ITGB1 expression is associated with poor prognosis. | Biomarker in prognosis of breast cancer. | [132,133] | |
| Linc-RoR | ENSG00000258609 | lincRNA | Upregulation/Oncogenic | Induces EMT. Contributes to tumor growth, invasion, metastasis and drug resistance through endogenous competition with various miR (145, 205, 133, 34) and inhibition of p53 expression. | Upregulation is a marker in multi-drug resistance, chemotherapy tolerance. Potential therapeutic target for aggressive and metastatic breast cancer. | [91,134] | |
| LINC00472 | ENSG00000233237 | lincRNA | Downregulation/Tumor suppressor | Associated with tumor grade, estrogen receptor status and molecular subtype in breast cancer. Repressed by methylation of its promoter. | Potential prognosis and predictive biomarker. | [135,136] | |
| LSINCT5 | ENSG00000281560 | lincRNA | Upregulation/Oncogenic | Promotes cell proliferation. | [137] | ||
| MALAT1 | ENSG00000251562 | NAT (nc|nc) | Upregulation/Oncogenic | Plays a critical role in pre-mRNA alternative splicing. Regulates EMT gene expression. | Knockdown reduces cell growth, invasion, migration and differentiation into cystic tumors. Potential prognosis marker in ER− and prediction marker for endocrine treatment sensitivity in ER+. | [138,139,140,141] | |
| MEG3 | ENSG00000214548 | NAT (nc|nc) | Downregulation/Tumor suppressor | Represses MDM2, leading to p53 accumulation. Silences genomic loci of TGFβ-associated genes by interaction with PRC2. Represses AKT signaling pathway. Inhibits EMT by sponging miR-421. | Expression promotes apoptosis, inhibits proliferation and angiogenesis. | [87,88,142,143] | |
| MIR31HG (LOC554202) | ENSG00000171889 | Sense-overlapping lncRNA | Downregulation/Tumor suppressor | Host gene of miR-31. Silenced in TNBC by promoter hypermethylation. Inhibits invasion-metastasis cascade by targeting pro-metastasis genes (i.e., RhoA and WAVE3). | [144,145,146] | ||
| NEAT1 | ENSG00000245532 | lincRNA | Upregulation/Oncogenic | Modulates miRNA biogenesis by organizing key components of paraspeckles and regulates transcription through protein sequestration into paraspeckles. Promotes proliferation and EMT. In ER+, NEAT1 is indispensable for interaction between FOXN3 and SINA3 complex. Regulates EZH2 through miR-101. | Overexpression of miR-548ar-3p downregulates NEAT1 and results in inhibition of cell growth. | [147,148,149,150,151,152] | |
| PANDAR | ENSG00000281450 | lincRNA | Upregulation/Oncogenic | Represses p16INK4A expression through modulating the recruitment of Bmi1 to the p16INK4A promoter. Removes cycle arrest possibility during G1/S transition. | Potential therapeutic target. | [85] | |
| PTPRG-AS1 | ENSG00000241472 | NAT (nc|pc) | PTPRG, C3ORF14 | Upregulation/Oncogenic | Differentially expressed between ER+ and ER− subtypes. | [153,154] | |
| PVT1 | ENSG00000249859 | NAT (nc|pc) | TMEM75 | Upregulation/Oncogenic | Co-operation between c-Myc and PVT1. Enhances c-Myc stability through inhibiting its phosphorylation. | Due to synergy between c-Myc and PVT1, silencing PVT1 expression decreases cell proliferation and increases apoptosis. Potential therapeutic target. | [155,156,157,158] |
| SNHG17 | ENSG00000196756 | lincRNA | Differentially expressed between ER+ and ER− subtypes. Low expression associated with overall survival. Expression correlates with tumor grade. | [154] | |||
| SOX2-OT | ENSG00000242808 | NAT (nc|pc), sense-overlapping lncRNA, lincRNA | DNAJC19 | Upregulation/Oncogenic | Through positive effect on SOX2 expression, SOX2OT plays a key role in pluripotency and tumorigenesis. | Potential prognosis marker and therapeutic target. | [159,160] |
| SPRY4-IT1 | ENSG00000281881 | Sense-intronic lncRNA | Upregulation/Oncogenic | Upregulates ZNF703 involved in the activation of the mTor signaling pathway. Promotes cell proliferation and inhibits apoptosis. | SPRY4-IT1 positively correlates with tumor size and pathological stage. Prognostic marker and potential therapeutic target. | [161,162] | |
| TERRA (Telomeric repeat-containing RNA) | lncRNA | Misregulation | Transcribed from telomeric C-rich strand. Interacts with TRF1 and TRF2 to facilitate heterochromatin formation. Provides RNA template to aid telomerase function. | Potential therapeutic target to impair telomerase activity. | [163,164,165,166] | ||
| TP73-AS1 | ENSG00000227372 | NAT (nc|pc) | TP73 | Upregulation/Oncogenic | TP73-AS1/miR-200a/ZEB1 forms a regulating loop. TP73-AS1 competes with ZEB1 for binding to miR-200a. ZEB1 binds to TP73-AS1 promoter and activates its expression. Upregulation of TP73-AS1/ZEB1 promotes cell invasion and migration. | Potential therapeutic target. | [94,167] |
| treRNA | ENSG00000231265 | lincRNA | Upregulation/Oncogenic | Regulates translation through interaction with ribonucleoprotein complex, which will bind to the translation initiation factor (EIF4G1). Overexpressed in lymph-node metastasis. Promotes tumor invasion and metastasis. Regulates expression of metastasis promoting-gene Snail. Suppresses epithelial markers and translation of E-cadherin mRNA. | [168] | ||
| UCA1 | ENSG00000214049 | lincRNA | Upregulation/Oncogenic | Enhances chemotherapy resistance (tamoxifen) through mTor pathway inhibition and miR-18a downregulation. Promotes EMT through activating Wnt/β-catenin signaling. UCA1/hnRNP1 suppresses p27 protein level by competition. Downregulates tumor suppressor miR-143. | Potential urine biomarker. Knockdown reduces chemoresistance, cell migration and tumor size. | [169,170,171,172,173] | |
| ZFAS1 | ENSG00000177410 | NAT (nc|pc) | ZNFX1 | Downregulation in breast cancer/Upregulated in other cancers | Associated with ribosomes in breast cancer. Role in development and cell differentiation in mammary gland. | Potential biomarker. | [174,175,176,177,178] |
Abbreviations: nc: non-coding; pc: protein coding; lncRNA: long intergenic non-coding RNA; TNBC: triple negative breast cancer; NAT: natural antisense transcript; EMT: epithelial to mesenchymal transition; ER+ and ER−: estrogen receptor positive and negative breast cancers; G1/S transition: The transition between the two first phases of the cell cycle; ANRASSF1: RASSF1 antisense RNA 1; RASSF1: Ras association domain family member 1; PRC: polycomb repressive complex; ANRIL: antisense non-coding RNA in the INK4 locus; CDKN2A: cyclin dependent kinase inhibitor 2A; CDKN2B: cyclin dependent kinase inhibitor 2B; CBX7: chromobox 7; SUZ12: SUZ12 polycomb repressive complex 2 subunit; INK: cyclin dependent kinase inhibitor; LSAMP: limbic system-associated membrane protein; BCAR4: breast cancer anti-estrogen resistance 4; SNIP: SRC kinase signaling inhibitor 1; PNUTS: protein phosphatase 1 regulatory subunit; SMO: smoothened, frizzled class receptor; BCYRN1: brain cytoplasmic RNA 1; CCAT2: colon cancer associated transcript 2; TGFbeta: transforming growth factor beta 1; Wnt: wingless-type MMTV integration site family; CRNDE: colorectal neoplasia differentially expressed; DANCR: differentiation antagonizing non-protein coding RNA; DSCAM-AS1: DSCAM antisense RNA 1; DSCAM: DS cell adhesion molecule; FAM83H-AS1: FAM83H antisense RNA 1; GAS5: growth arrest specific 5; ZBTB37: zinc finger and BTB domain containing 37; GR: glucocorticoïd receptor; H19: H19, imprinted maternally expressed transcript; BIK: BCL2 interacting killer; HIF1A-AS2: HIF1A antisense RNA 2; HIF1A: hypoxia inducible factor 1 alpha subunit; HMMR-AS1: HMMR antisense RNA 1; HMMR: hyaluronan mediated motility receptor; BRCA1: BRCA1, DNA repair associated; BRCA2: BRCA2, DNA repair associated; HOTAIR: HOX transcript antisense RNA; HOXC11: homeobox C11; LSD1: lysine demethylase 1A; HOTAIRM1: HOXA transcript antisense RNA, myeloid-specific 1; HOX: homeobox; HOXA-AS2: HOXA cluster antisense RNA 2; TGFBR2: transforming growth factor beta receptor 2; RELA: RELA proto-oncogene, NF-kB subunit; HOXA11-AS: HOXA11 antisense RNA; LSINCT5: long stress-induced non-coding transcript 5; MALAT1: metastasis associated lung adenocarcinoma transcript 1; MEG3: maternally expressed 3; MDM2: transformed mouse 3T3 cell double minute 2 proto-oncogene; AKT: thymoma viral proto-oncogene serine/threonine kinase 1; RhoA: ras homolog family member A; RhoA: ras homolog family member A; WAVE3: WAS protein family member 3; NEAT1: nuclear paraspeckle assembly transcript 1; FOXN3: forkhead box N3; SINA3: E3 ubiquitin-protein ligase SINAT5-like; PANDAR: promoter of CDKN1A antisense DNA damage activated RNA; Bmi1: BMI1 proto-oncogene, polycomb ring finger; PTPRG-AS1: PTPRG antisense RNA 1; PTPRG: protein tyrosine phosphatase, receptor type G; C3ORF14: chromosome 3 open reading frame 14; PVT1: Pvt1 oncogene; TMEM75: transmembrane protein 75; c-Myc: MYC proto-oncogene, bHLH transcription factor; SNHG17: small nucleolar RNA host gene 17; SOX2-OT: SOX2 overlapping transcript; DNAJC19: DnaJ heat shock protein family (Hsp40) member C19; SOX2: SRY-box 2; SPRY4-IT1: SPRY4 intronic transcript 1; ZNF703: zinc finger protein 703; mTor: mechanistic target of rapamycin kinase; TRF: telomeric repeat binding factor; TP73-AS1: TP73 antisense RNA 1; TP73: tumor protein p73; ZEB1: zinc finger E-box binding homeobox 1; treRNA: translation regulatory long non-coding RNA 1; EIF4G1: eukaryotic translation initiation factor 4 gamma 1; Snail: snail family transcriptional repressor 1; UCA1: urothelial cancer associated 1; ZFAS1: ZNFX1 antisense RNA 1; ZNFX1: zinc finger NFX1-type containing 1.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Latgé, G.; Poulet, C.; Bours, V.; Josse, C.; Jerusalem, G. Natural Antisense Transcripts: Molecular Mechanisms and Implications in Breast Cancers. Int. J. Mol. Sci. 2018, 19, 123. https://doi.org/10.3390/ijms19010123
AMA Style
Latgé G, Poulet C, Bours V, Josse C, Jerusalem G. Natural Antisense Transcripts: Molecular Mechanisms and Implications in Breast Cancers. International Journal of Molecular Sciences. 2018; 19(1):123. https://doi.org/10.3390/ijms19010123
Chicago/Turabian StyleLatgé, Guillaume, Christophe Poulet, Vincent Bours, Claire Josse, and Guy Jerusalem. 2018. "Natural Antisense Transcripts: Molecular Mechanisms and Implications in Breast Cancers" International Journal of Molecular Sciences 19, no. 1: 123. https://doi.org/10.3390/ijms19010123
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.