The Potential Roles of the Apoptosis-Related Protein PDRG1 in Diapause Embryo Restarting of Artemia sinica

Abstract
:1. Introduction
2. Results
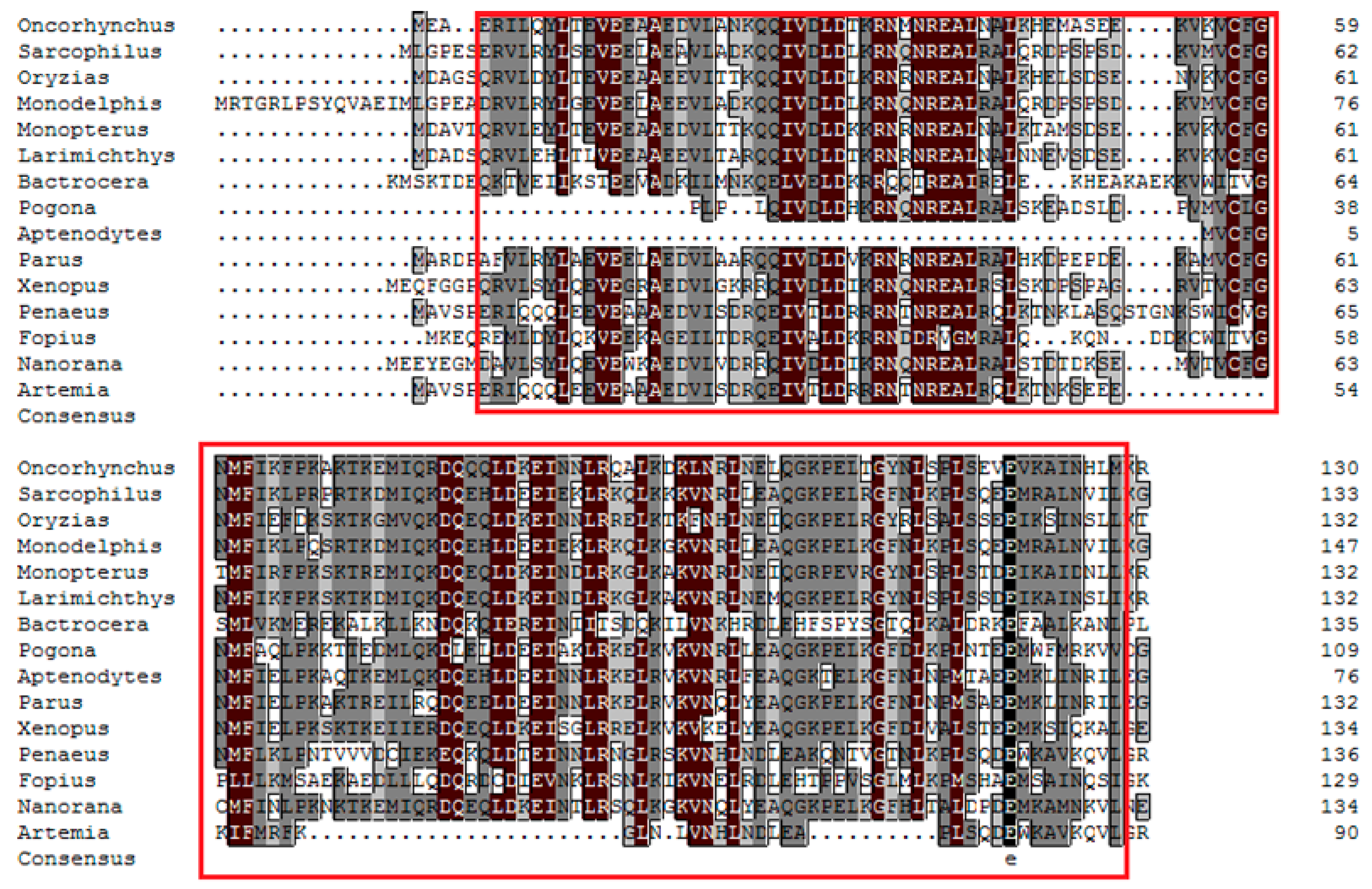
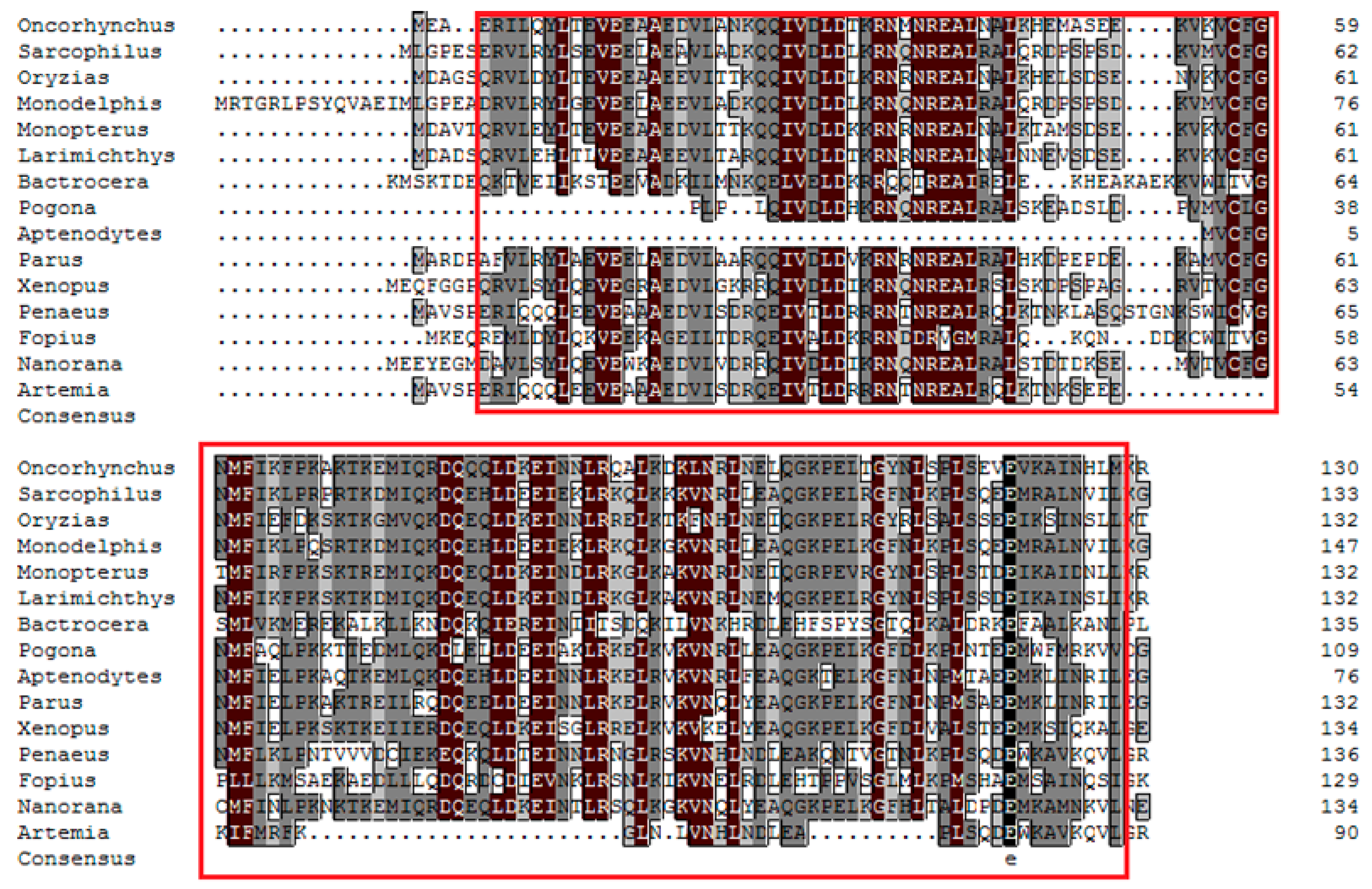
2.1. Cloning and Bioinformatic Analysis of As-pdrg1
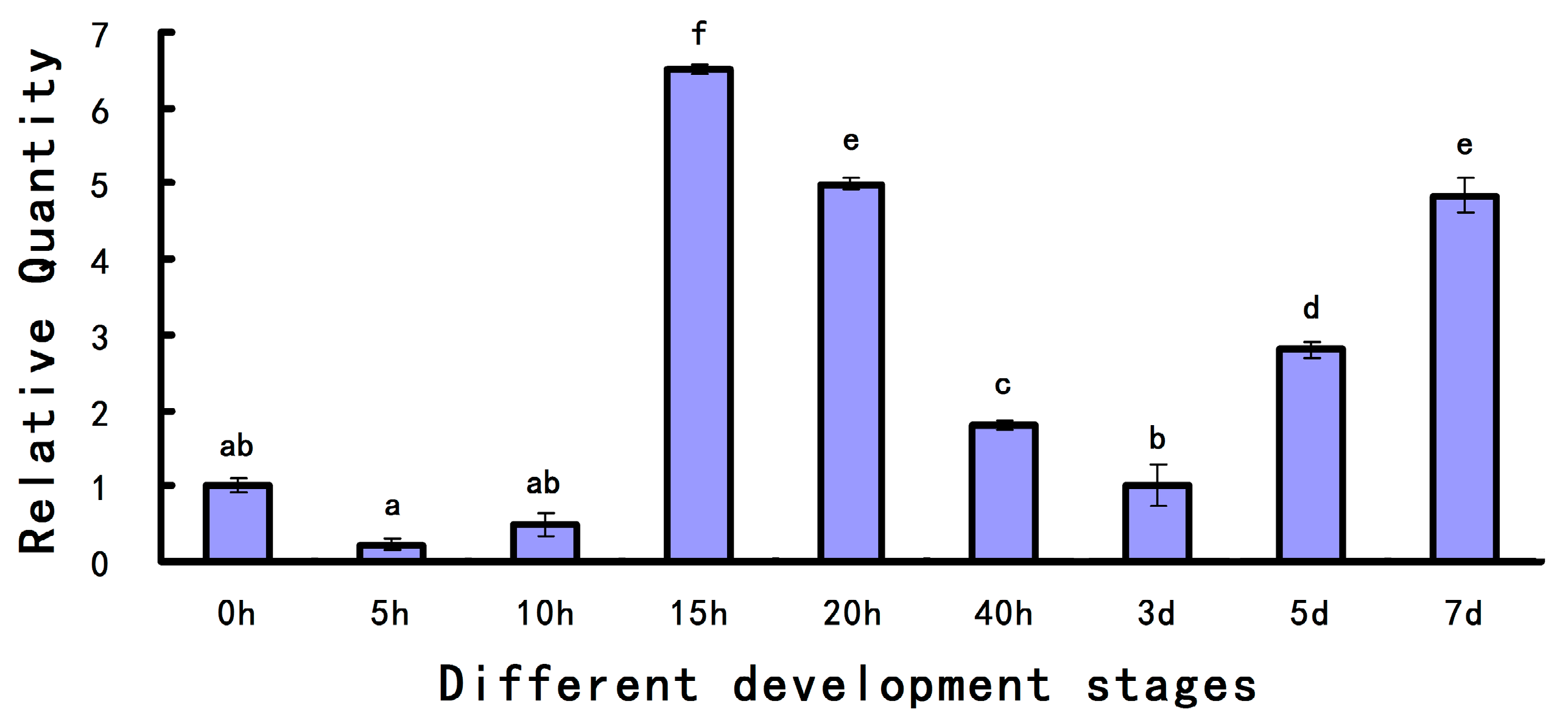
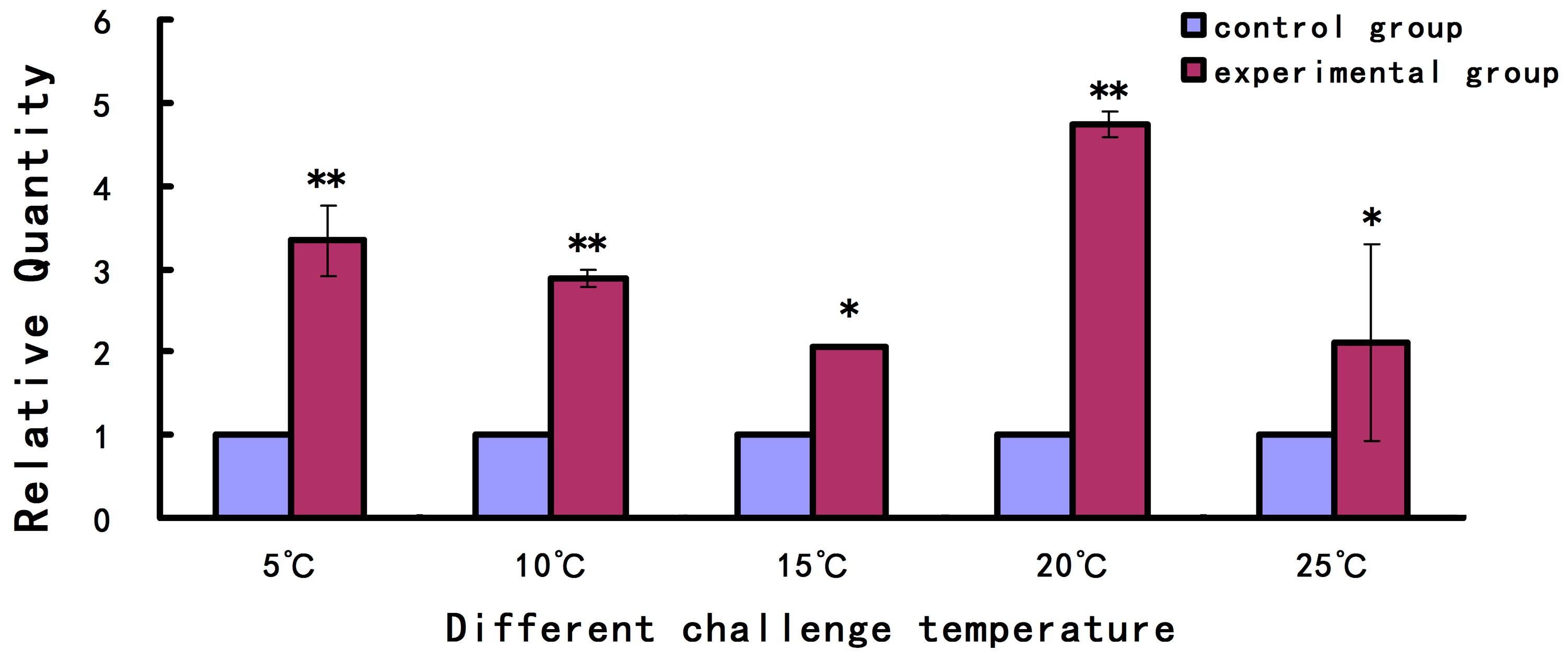
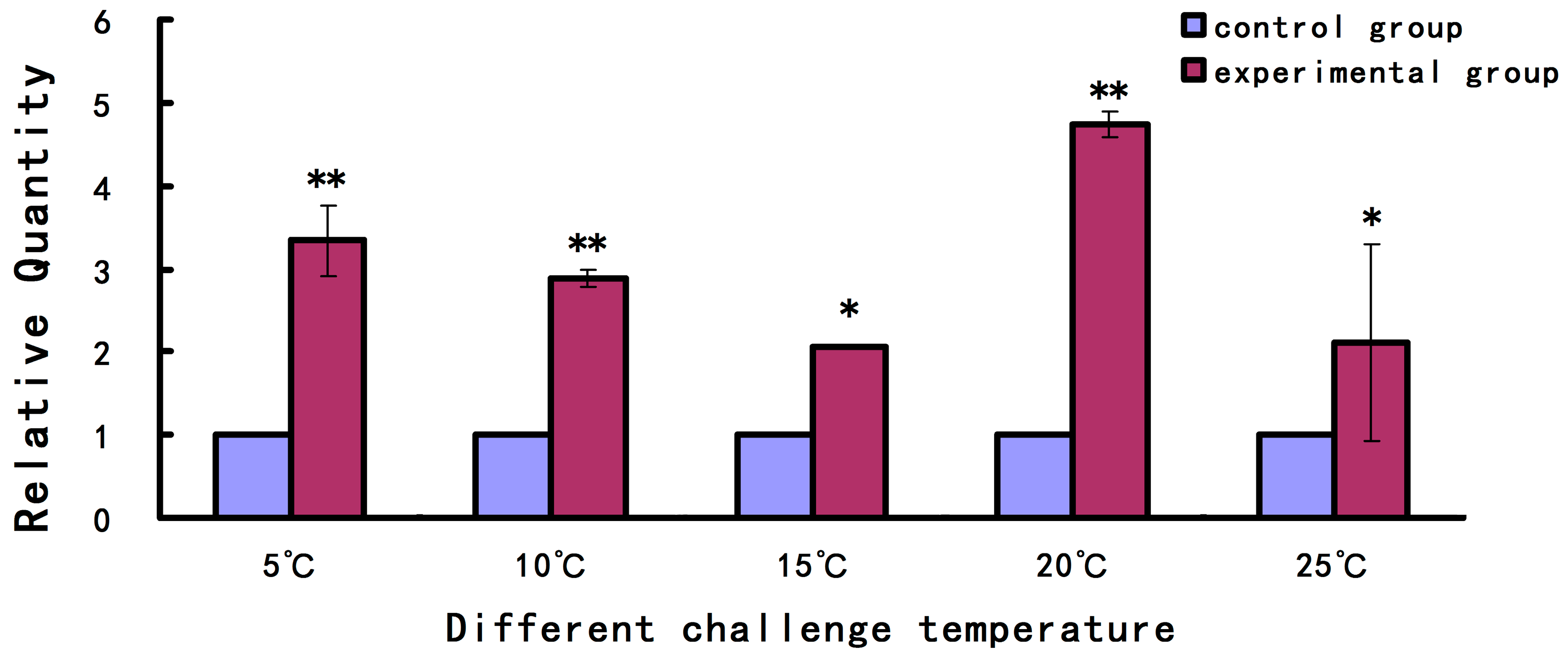
2.2. Expression Level of As-pdrg1 at Different Developmental Stages and under Different Stresses
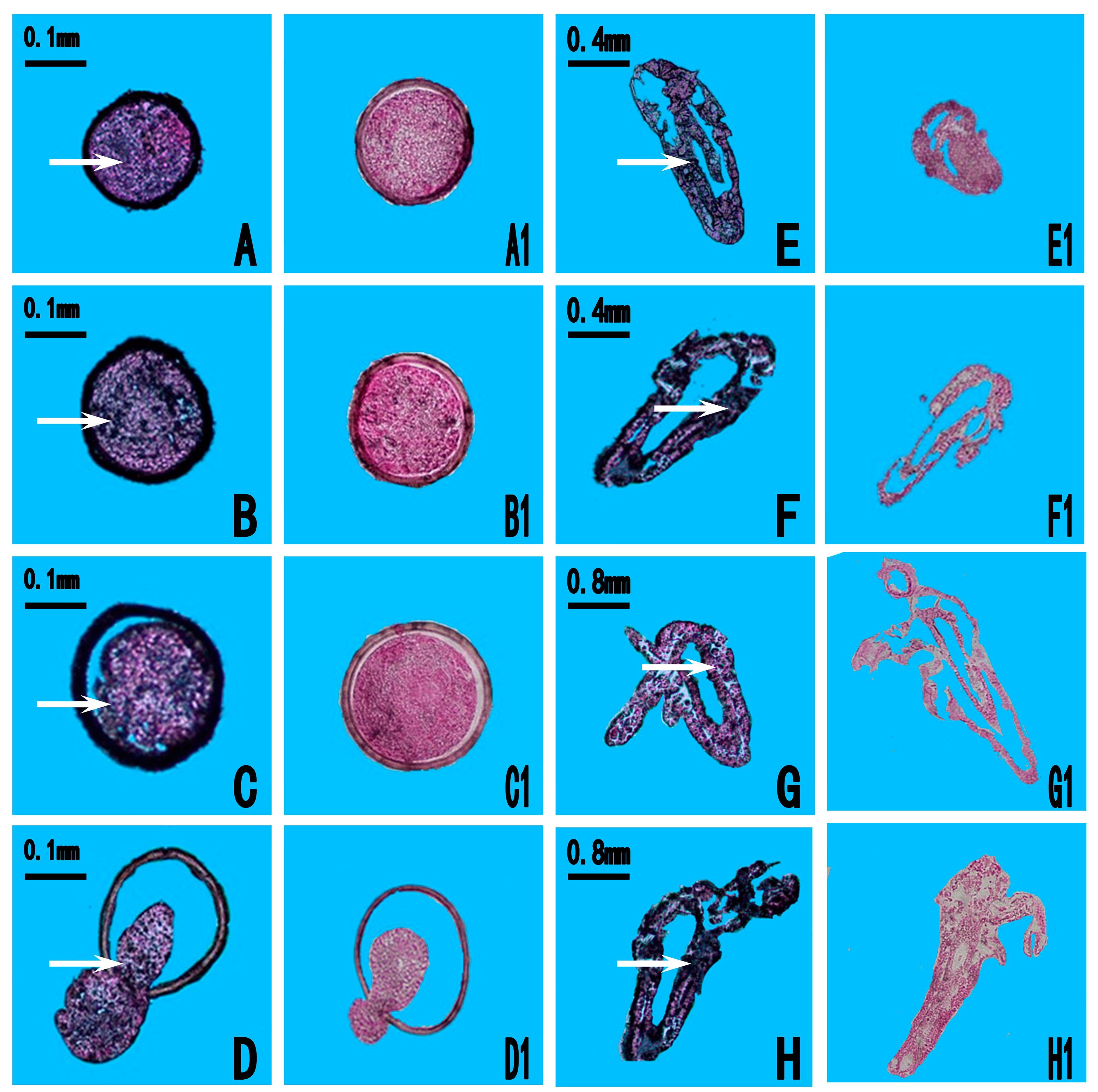
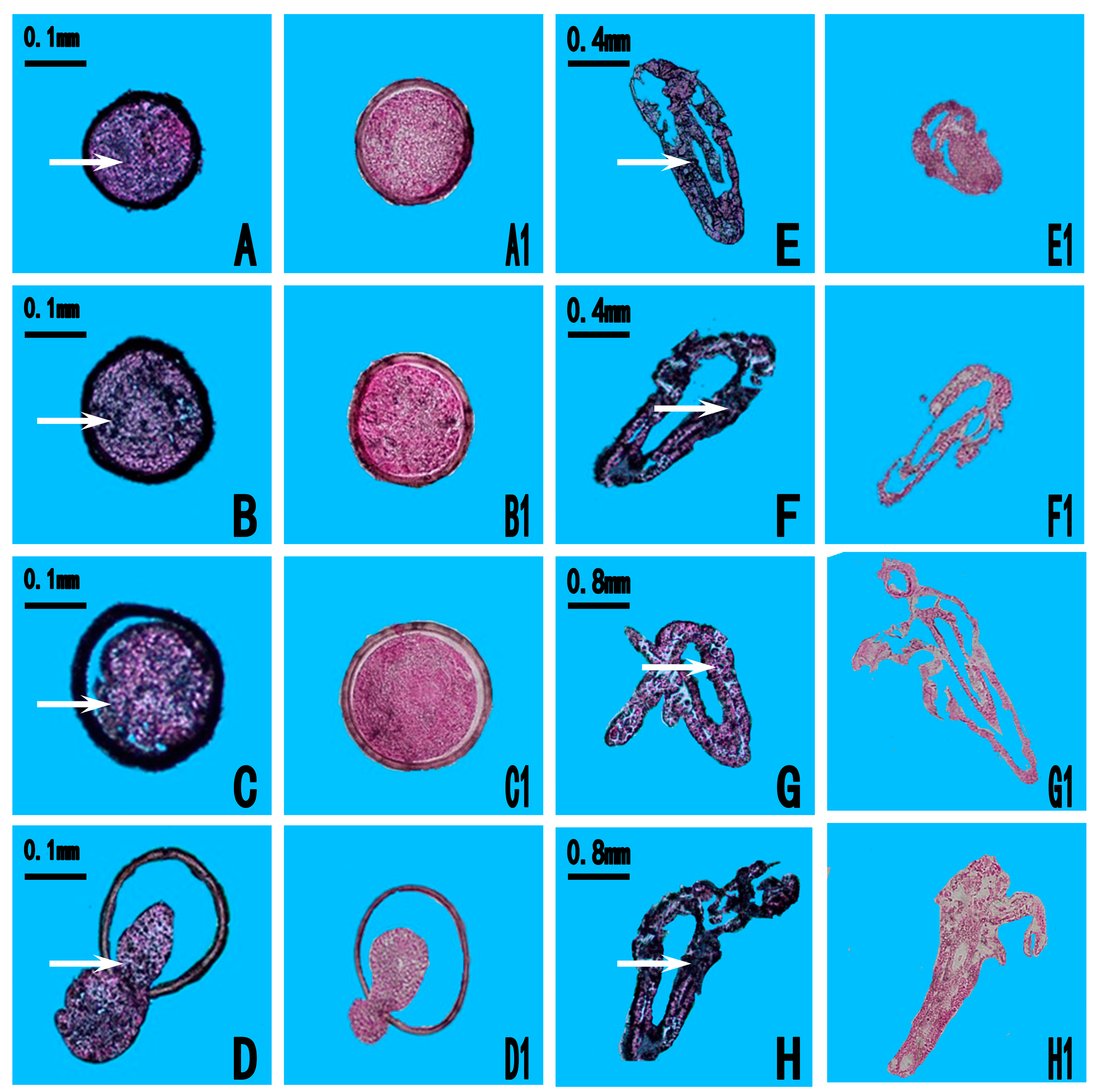
2.3. Expression Location of As-pdrg1
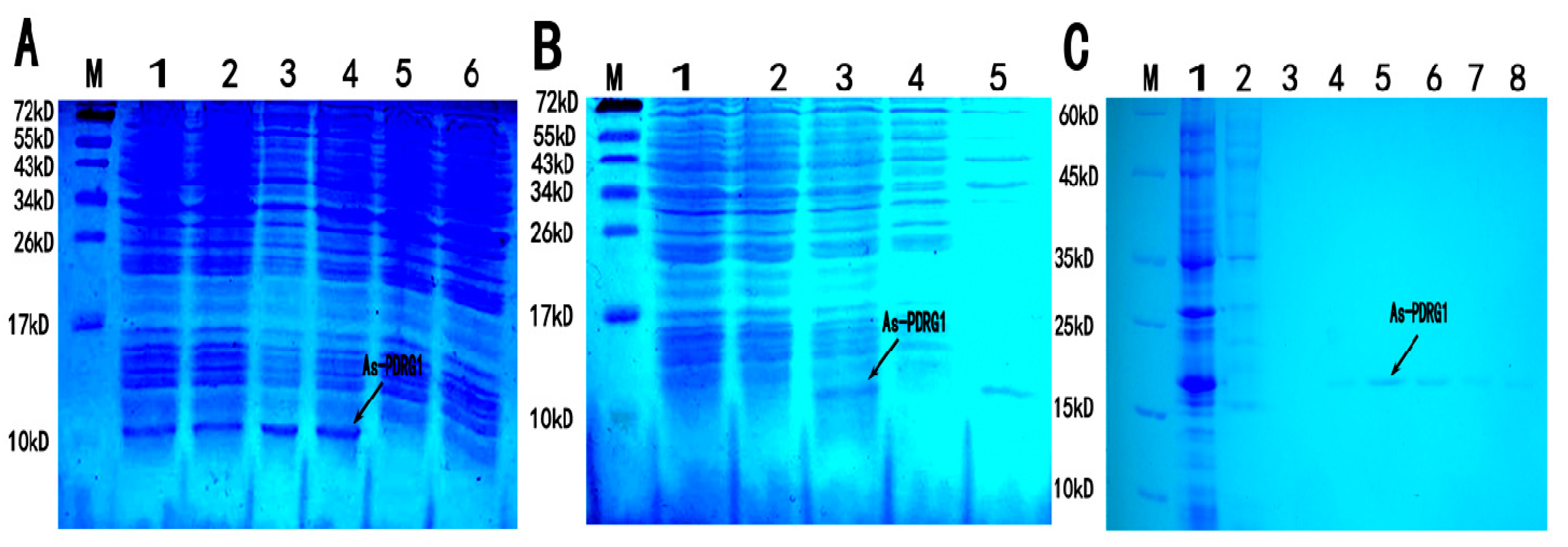
2.4. Purification and Expression of As-PDRG1 Protein
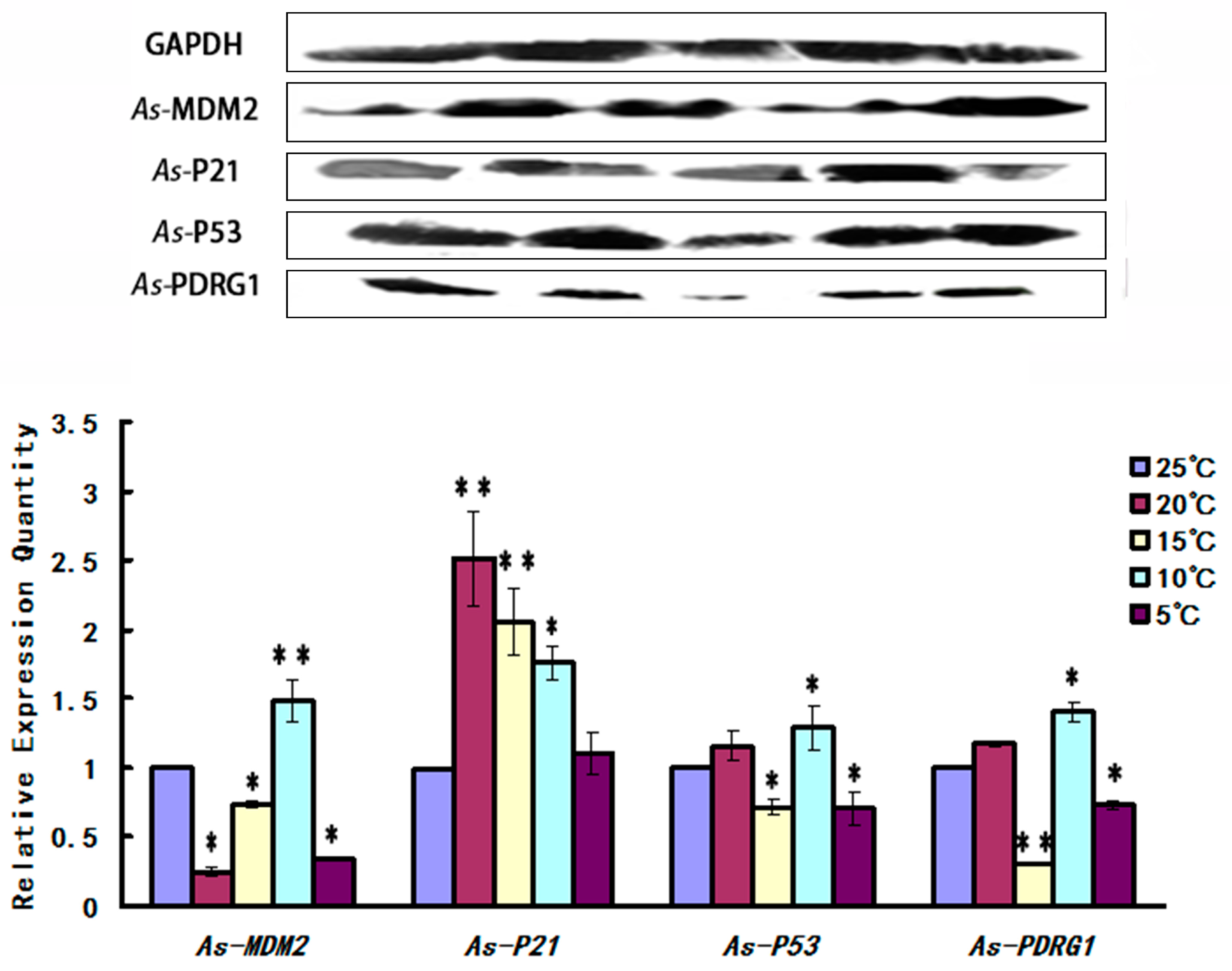
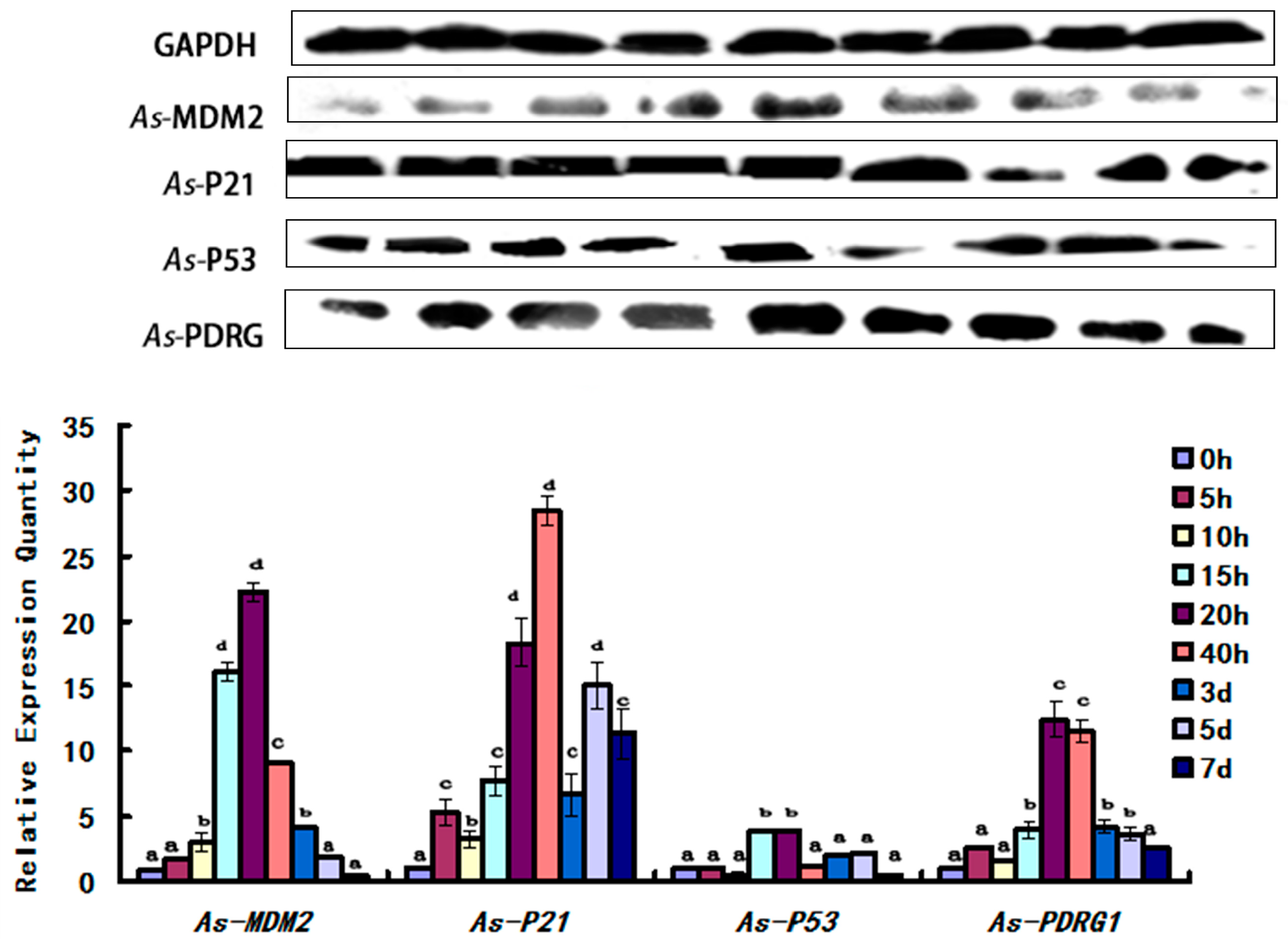
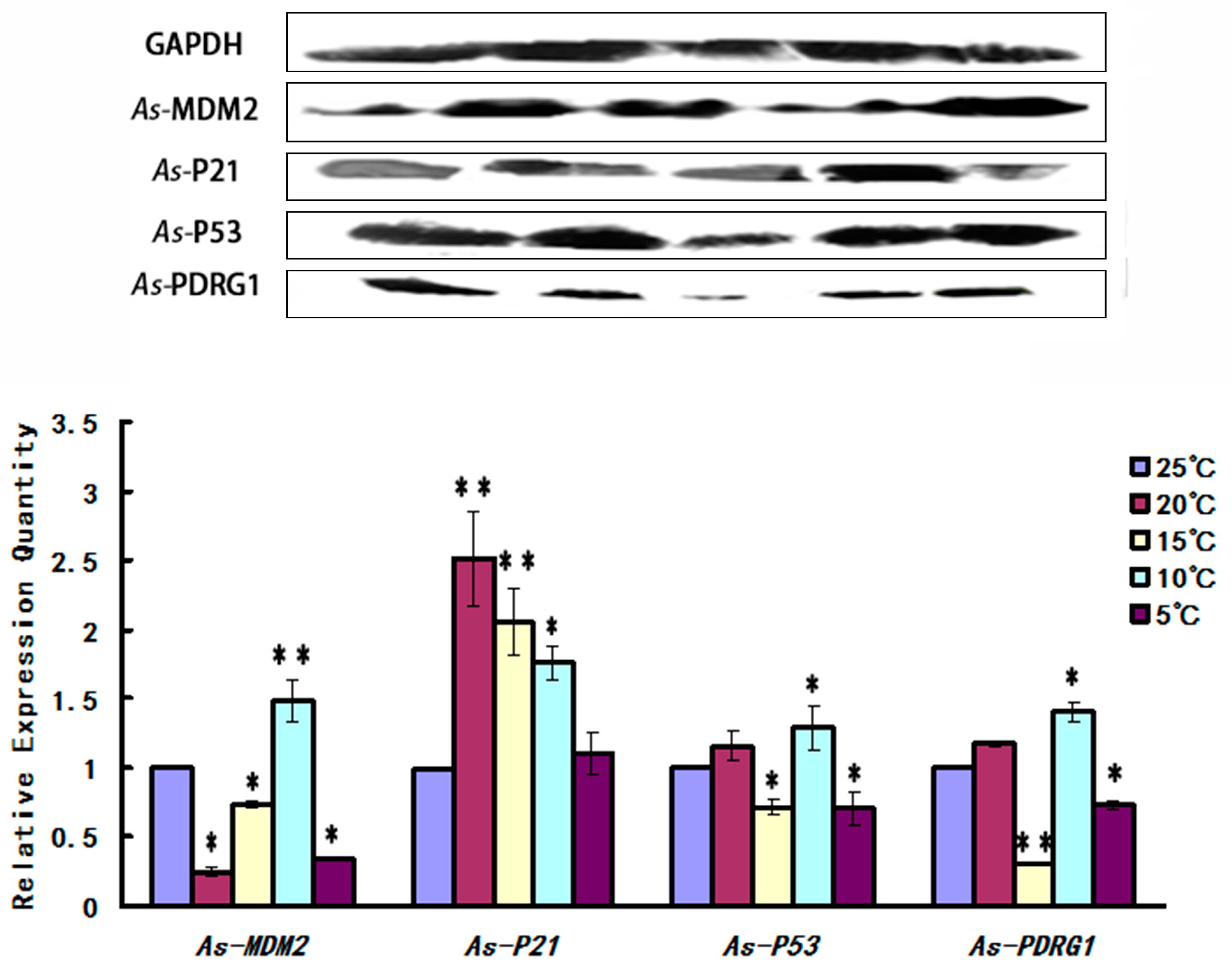
2.5. Expression Pattern of As-PDRG1 Tested by Western Blotting in Early Embryo Development and to Stress
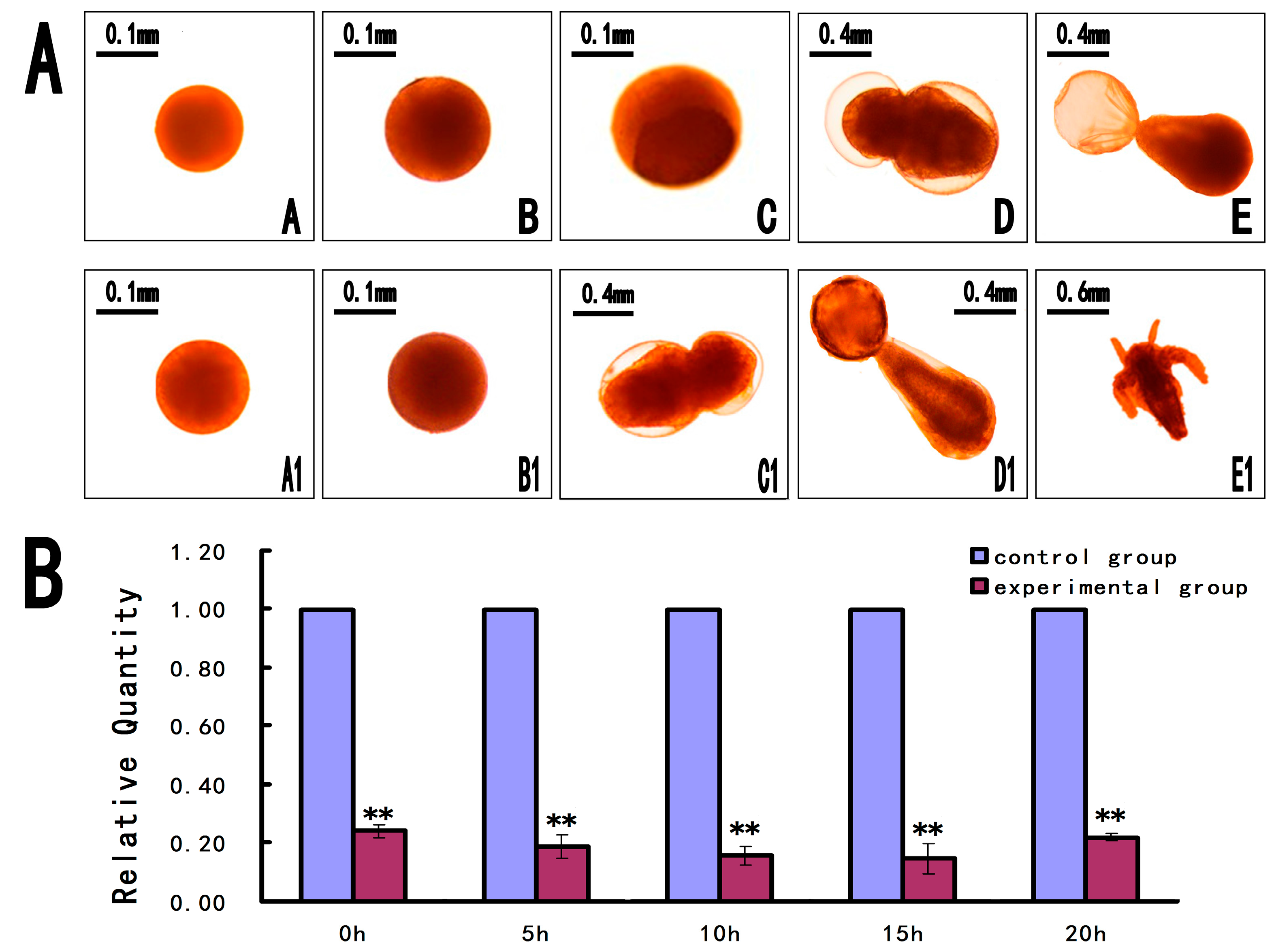
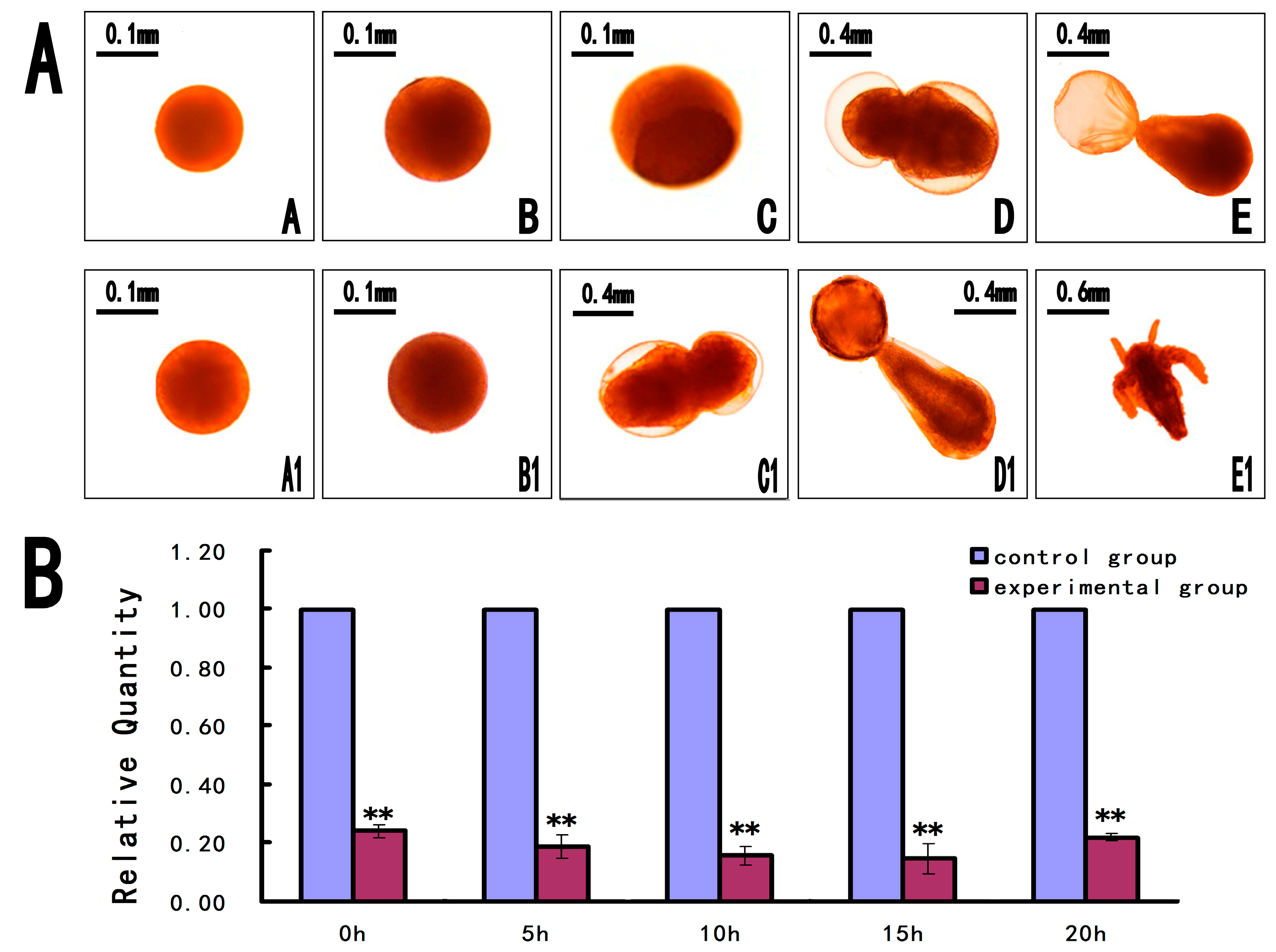
2.6. Short Interfering RNA (siRNA) for Knockdown of As-PDRG1
3. Discussion
4. Materials and Methods
4.1. Sample Reparation
4.2. Cloning the Full-Length pdrg1 cDNA
4.3. Quantitative Real-Time PCR (qPCR)
4.3.1. Expression of As-pdrg1 at Different Developmental Stages
4.3.2. Expression of As-pdrg1 in Response to Stress
4.4. Construction, Expression, and Purification of pET-30a PDRG1 Protein
4.5. Expression and Purification of the Recombinant Protein from Escherichia Coli
4.6. In Situ Hybridization (ISH) Assay
4.7. Western Blotting
4.7.1. Protein Production of As-PDRG1 in Different Stages of Early Embryo Development
4.7.2. Expression of As-PDRG1 under Low Temperature
4.8. Short Interfering RNA (siRNA) Assay
4.9. Bioinformatics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PDRG1 | p53 and DNA damage-regulated gene 1 |
| As-pdrg1 | mRNA from Artemia sinica |
| As-PDRG1 | protein from Artemia sinica |
| ORF | open reading frame |
| PCR | polymerase chain reaction |
| UTR | untranslated region |
| ISH | in situ hybridization |
| siRNA | short interfering RNA |
| DIG | digoxigenin |
References
- Zhang, S.; Yao, F.; Jing, T.; Zhang, M.; Zhao, W.; Zou, X.; Sui, L.; Hou, L. Cloning, expression pattern, and potential role of apoptosis inhibitor 5 in the MARK termination of embryonic diapause and early embryo development of Artemia sinica. Gene 2017, 628, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.C.; Yao, F.; Luan, H.; Zhao, W.; Jing, T.; Zhang, S.; Hou, L.; Zou, X.Y. APC/CCDC20 and APC/C play pivotal roles in the process of embryonic development in Artemina sinica. Sci. Rep. 2016, 6, 39047. [Google Scholar] [CrossRef] [PubMed]
- Abatzopoulos, T.J.; Beardmore, J.A.; Clegg, J.S.; Sorgeloos, P. Artemia: Basic and Applied Biology; Kluwer Academic: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Li, X.; Hou, L.; Ma, J.; Liu, Y.D.; Zheng, L.P.; Zou, X.Y. Cloning and character-ization of β-catenin gene in early embryonic developmental stage of Artemia sinica. Mol. Biol. Rep. 2012, 39, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Jiang, L.J.; Sun, W.J.; Zhang, R.F.; Wang, J.Q.; Zhao, X.T.; An, J.L. Establishment and improvement of real-time fluorescence quantitative PCR for actin gene of Artemia sinica. J. Liaoning Normal Univ. 2006, 29, 15–19. [Google Scholar]
- Wang, X.L.; Yao, F.; Liang, X.Y.; Zhu, X.L.; Zheng, R.; Jia, B.L.; Hou, L.; Zou, X.Y. Cloning and expression of retinoblastoma-binding protein 4 gene in embryo diapause termination and in response to salinity stress from brine shrimp Artemia sinica. Gene 2016, 591, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.J.; Hou, L.; Zou, X.Y.; Zhang, R.F.; Wang, J.Q.; Sun, W.J.; Zhao, X.T.; An, J.L. Cloning and expression analysis of p26 gene in Artemia sinica. Acta Biochim. Biophys. Sin. 2007, 39, 351–358. [Google Scholar] [CrossRef] [PubMed]
- .Zheng, L.P.; Hou, L.; Chang, A.K.; Yu, M.; Ma, J.; Li, X.; Zou, X.Y. Expression pattern of a gram-negative bacteria-binding protein in early embryonic development of Artemia sinica and after bacterial challenge. Dev. Comp. Immunol. 2011, 35, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yao, F.; Chen, Y.; Zhang, R.; Lv, Y.; Zhao, N.; Wang, T.; Xin, W.; Hou, L.; Zou, X. Molecular cloning, characterization and expression analysis of ubiquitin pro-tein ligase gene (As-ubpl) from Artemia sinica. Mol. Cell Biol. 2013, 165, 90–98. [Google Scholar]
- Qing, T.; Zhao, X.; Luan, H.; Ba, H.; Yang, L.; Li, Z.; Hou, L.; Zou, X. Identification, expression pattern and functional characterization of As-MyD88 in bacteria challenge and during different developmental stages of Artemia sinica. Dev. Comp. Immunol. 2015, 50, 9–18. [Google Scholar]
- Zhao, W.; Yao, F.; Zhang, M.C.; Jing, T.; Zhang, S.; Hou, L.; Zou, X.Y. The Potential Roles of the G1LEA and G3LEA Proteins in Early Embryo Development and in Response to Low Temperature and High Salinity in Artemia sinica. PLoS ONE 2016, 11, E0162272. [Google Scholar] [CrossRef] [PubMed]
- Sardiu, M.E.; Cai, Y.; Jin, J.; Swanson, S.K.; Conaway, R.C.; Conaway, J.W.; Florens, L.; Washburn, M.P. Probabilistic assembly of human protein interaction networks from label-free quantitative proteomics. Proc. Natl. Acad. Sci. USA 2008, 105, 1454–1459. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Bertrand, E.; Pradet-Balade, B. HSP90 and the R2TP co-chaperone complex: Building multi-protein machineries essential for cell growth and gene expression. RNA Biol. 2012, 9, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Mita, P.; Savas, J.N.; Ha, S.; Djouder, N.; Yates, J.R., III; Logan, S.K. Analysis of URI nuclear interaction with RPB5 and components of the R2TP/prefoldin-like complex. PLoS ONE 2013, 8, e63879. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yang, H. Cloning and characterization of PDRG gene from sea cucumber Apostichopus japonicus and the expression in intestineduring aestivation. Mar. Sci. 2013, 37, 12. [Google Scholar]
- Luo, X.Q.; Huang, Y.; Sheikh, M.S. Cloning and characterization of a novel gene PDRG that is differential lyregulated by p53 and ultraviolet radiation. Oncogene 2003, 22, 7247–7257. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Luo, X.; Shi, J.; Sun, H.; Sun, Q.; Sheikh, M.S.; Huang, Y. PDRG1, a novel tumor marker for multiple malignancies that is selectively regulated by genotoxic stress. Cancer Biol. Ther. 2011, 11, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.L.; Xu, G.; Han, Q.Y.; Chen, Y. Construction of HeLa-PDRG1-knock Down Stable Cell Line by Lentivirus Infection. Sci. Technol. Eng. 2013, 13, 6385–6388. [Google Scholar]
- Pérez, C.; Pérez-Zúñiga, F.J.; Garrido, F.; Reytor, E.; Portillo, F.; Pajares, M.A. The Oncogene PDRG1 Is an Interaction Target of Methionine Adenosyltransferases. PLoS ONE 2016, 11, e0163761. [Google Scholar]
- Zhao, C.; Dai, W.; Qiu, L. Molecular cloning, characterization and expression analysis of a novel PDRG1 gene from black tiger shrimp (Penaeus monodon). Genet. Mol. Biol. 2017, 40, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Yao, F.; Cheng, C.; Wu, Y.; Mei, Y.L.; Li, X.J.; Liu, Y.; Wang, P.S.; Hou, L.; Zou, X.Y. The potential role of As-sumo-1 in the embryonic diapause process and early embryo development of Artemia sinica. PLoS ONE 2014, 9, e85343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Z.; Hou, M.; Li, Q.Y.; Han, L.L.; Yuan, Z.; Tan, J.; Du, B.; Zou, X.Y.; Hou, L. Expression patterns of As-ClC gene of Artemia sinica in early development and under salinity stress. Mol. Biol. Rep. 2013, 40, 3655–3664. [Google Scholar] [CrossRef] [PubMed]
- Goh, A.M.; Xue, Y.; Leushacke, M.; Li, L.; Wong, J.S.; Chiam, P.C.; Rahmat, S.A.; Mann, M.B.; Mann, K.M.; Barker, N.; et al. Mutant p53 accumulates in cycling and proliferating cells in the normal tissues of p53 R172H mutant mice. Oncotarget 2015, 6, 17968–17980. [Google Scholar] [CrossRef] [PubMed]
- Faried, A.; Faried, L.S.; Kato, H.; Asao, T.; Kuwano, H.; Yazawa, S. Targeting P53 tumor suppressor to induce apoptosis and cell cycle arrest in esophageal cancer cells by novel sugar-cholestanols compounds. Poster Sess. 2008, 7, 83–84. [Google Scholar] [CrossRef]
- Melino, G.; Lu, X.; Gasco, M.; Crook, T.; Knight, R.A. Functional regulation of p73 and p63 development and cancer. Trends Biochem. Sci. 2003, 28, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, J.; Wu, Y.; Shi, G.; Yuan, H.; Lu, Z.; Zhu, Q.; Wu, P.; Lu, C.; Guo, F.; et al. YY1 suppresses proliferation and migration of pancreatic ductal adenocarcinoma by regulating the CDKN3/MdM2/P53/P21 signaling pathway. Int. J. Cancer 2017. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Y.; Wu, M.; Li, X. Oxidative stress-mediated p53/p21WAF1/CIP1 pathway may be involved in microcystin-LR-induced cytotoxicity in HepG2 cells. Chemosphere 2017, 194, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Li, X.R.; Lin, X.Y.; Xiao, Z.X.; Jiang, H.X.; Tang, Q.L.; Liu, X. Structure and Function of P21 and New Research Progress. Med. Recapitul. 2008, 14, 125–134. [Google Scholar]
- Luo, J.L.; Cao, J.P.; Zhu, W.; Feng, S.; Sheng, F.J.; Zhu, C.Y.; Zheng, S.Y. Interaction between ATM and Radiation-activated Phosphorylation of P53 and P21. Chin. J. Cancer 2005, 24, 1059–1063. [Google Scholar]
- Cheng, C.; Yao, F.; Chu, B.; Li, X.J.; Liu, Y.; Wu, Y.; Mei, Y.L.; Wang, P.S.; Hou, L.; Zou, X.Y. Identification of the glycerol kinase gene and its role in diapause embryo restart and early embryo development of Artemia sinica. Gene 2014, 537, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Koci, L.; Chlebova, K.; Hyzdalova, M.; Hofmanova, J.; Jira, M.; Kysela, P.; Kozubik, A.; Kala, Z.; Krejci, P. Apoptosis inhibitor 5 (API-5; AAC-11; FIF) is upregulated in human carcinomas in vivo. Oncol. Lett. 2012, 3, 913–916. [Google Scholar] [PubMed]
- Saigusa, S.; Tanaka, K.; Toiyama, Y.; Matsushita, K.; Kawamura, M.; Okugawa, Y.; Hiro, J.; Inoue, Y.; Uchida, K.; Mohri, Y.; et al. Gene expression profiles of tumor regression grade in locally advanced rectal cancer after neoadjuvant chemoradiotherapy. Oncol. Rep. 2012, 28, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Sorgeloos, P.; Lavens, P.; Leger, P.; Tackaert, W.; Versuchele, D. Manual for the Culture and Use of Brine Shrimp Artemia in Aquaculture; State University of Ghent, Faculty of Agriculture: Ghent, Belgium, 1986; Volume 126, pp. 312–319. [Google Scholar]
- Hallstrom, T.C.; Mori, S.; Nevins, J.R. An E2F1-dependent gene expression pro-gram that determines the balance between proliferation and cell death. Cancer Cell 2008, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative geneexpression data using real-time quantitative PCR andthe 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Verger, A.; Perdomo, J.; Crossley, M. Modification with SUMO A role in transcriptional regulation. EMBO Rep. 2003, 4, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Azmi, T.I.; Oshea, J.D. Mechanism of deletion of Endo-thelial-Cells during regression of the Corpus-Luteum. Lab. Investig. 1984, 51, 206–217. [Google Scholar] [PubMed]
- Erica, S.J.; Gunter, B. Ubc9p Is the Conjugating Enzyme for the Ubiquitin-like Protein Smt3p. J. Biol. Chem. 1997, 272, 26799–26802. [Google Scholar]
- Freichel, M.; Vennekens, R.; Olausson, J.; Stolz, S.; Philipp, S.E.; Weissgerber, P.; Flockerzi, V. Functional role of TRPC proteins in native systems: Implications from knockout and knock-down studies. J. Physiol. 2005, 567, 59–66. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Position | Context a | Score b |
|---|---|---|---|
| Ser | 5 | MRFGSSMFK | 0.500 |
| 6 | RFGSSMFKI | 0.841 | |
| 40 | ILKDSVMVL | 0.675 | |
| 45 | VMVLSQIED | 0.941 | |
| 50 | QIEDSPGMK | 0.950 | |
| 59 | GLNLSPLGS | 0.802 | |
| 63 | SPLGSEERK | 0.812 | |
| 70 | RKAISDLIG | 0.578 |
| Primer | Sequence (5′-3′) | Direction |
|---|---|---|
| As-pdrg1F | CTGAAATAGAAGAGGCAGGT | Forward |
| As-pdrg1R | CTATTTCGGATTTGATTGGT | Reverse |
| 3′As-pdrg1 | TGAGCCCATTGGGTAGTGAAGA | Forward |
| 5′As-pdrg1 | TGAACAGGAGATAAATTTCATAG | Reverse |
| ISH-pdrg1F | TGAACAGGAGATAAATTTCATAGCTG | Forward |
| ISH-pdrg1R | TTGATTGGTGATTAAGGTGTTGAAA | Reverse |
| RT-pdrg1F | TGGTGAAGACGAATAGGTCCA | Forward |
| RT-pdrg1R | AAGTGCAGTGCAAGCTAACC | Reverse |
| β-actin-F | GTGTGACGATGATGTTGCGG | Forward |
| β-actin-R | GCTGTCCTTTTGACCCATTCC | Reverse |
| ORF-pdrg1F | TGAACAGGAGATAAATTTCATAG | Forward |
| ORF-pdrg1R | TTTCAACACCTTAATCACCAATC | Reverse |
| Sense siRNA | GGAUUUGAUUGGUGAUUAAUU | Forward |
| Anti siRNA | UUCCUAAACUAACCACUAAUU | Reverse |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Yao, F.; Zhang, H.; Li, N.; Zou, X.; Sui, L.; Hou, L. The Potential Roles of the Apoptosis-Related Protein PDRG1 in Diapause Embryo Restarting of Artemia sinica. Int. J. Mol. Sci. 2018, 19, 126. https://doi.org/10.3390/ijms19010126
Zhang W, Yao F, Zhang H, Li N, Zou X, Sui L, Hou L. The Potential Roles of the Apoptosis-Related Protein PDRG1 in Diapause Embryo Restarting of Artemia sinica. International Journal of Molecular Sciences. 2018; 19(1):126. https://doi.org/10.3390/ijms19010126
Chicago/Turabian StyleZhang, Wan, Feng Yao, Hong Zhang, Na Li, Xiangyang Zou, Linlin Sui, and Lin Hou. 2018. "The Potential Roles of the Apoptosis-Related Protein PDRG1 in Diapause Embryo Restarting of Artemia sinica" International Journal of Molecular Sciences 19, no. 1: 126. https://doi.org/10.3390/ijms19010126