Small Myristoylated Protein-3, Identified as a Potential Virulence Factor in Leishmania amazonensis, Proves to be a Protective Antigen against Visceral Leishmaniasis

 , ,
, ,
Abstract
:
1. Introduction
2. Results
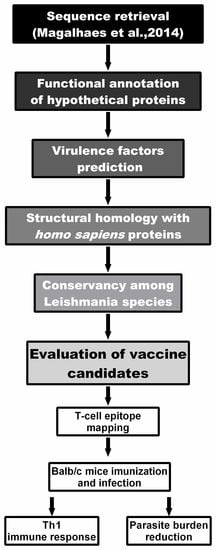
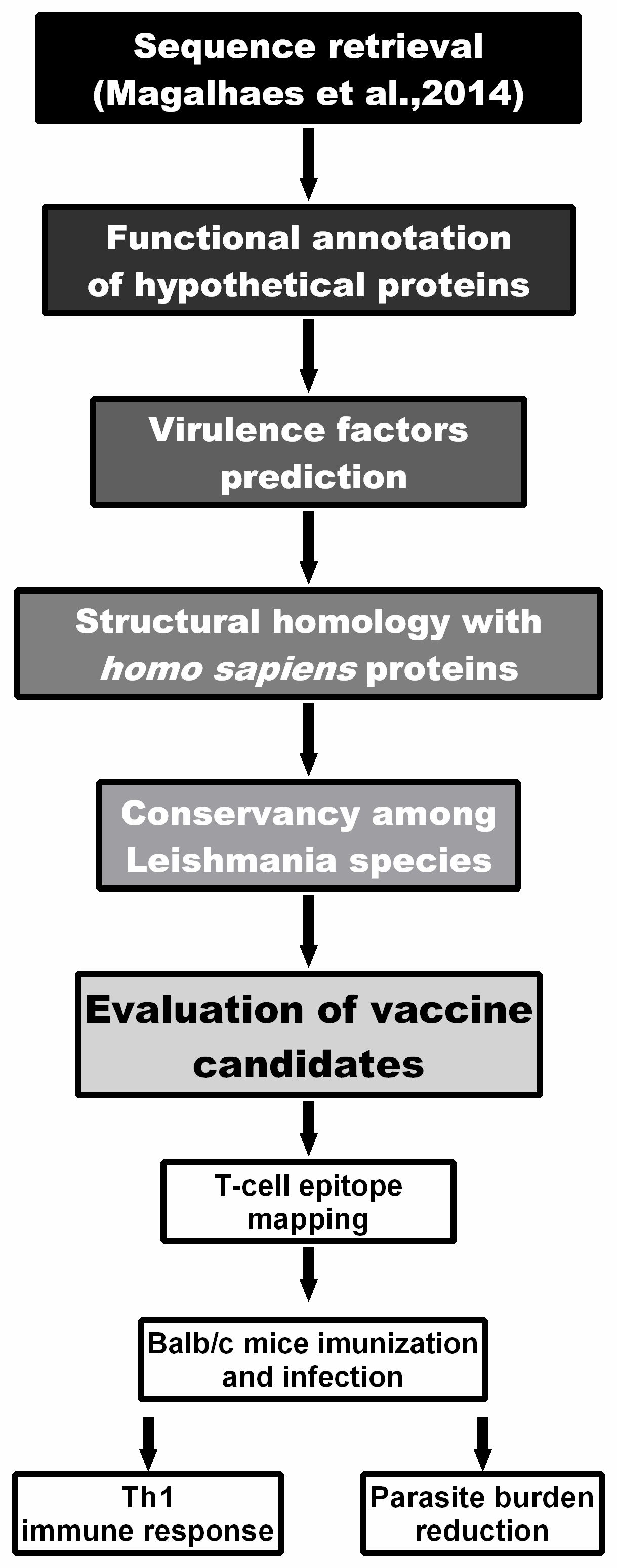
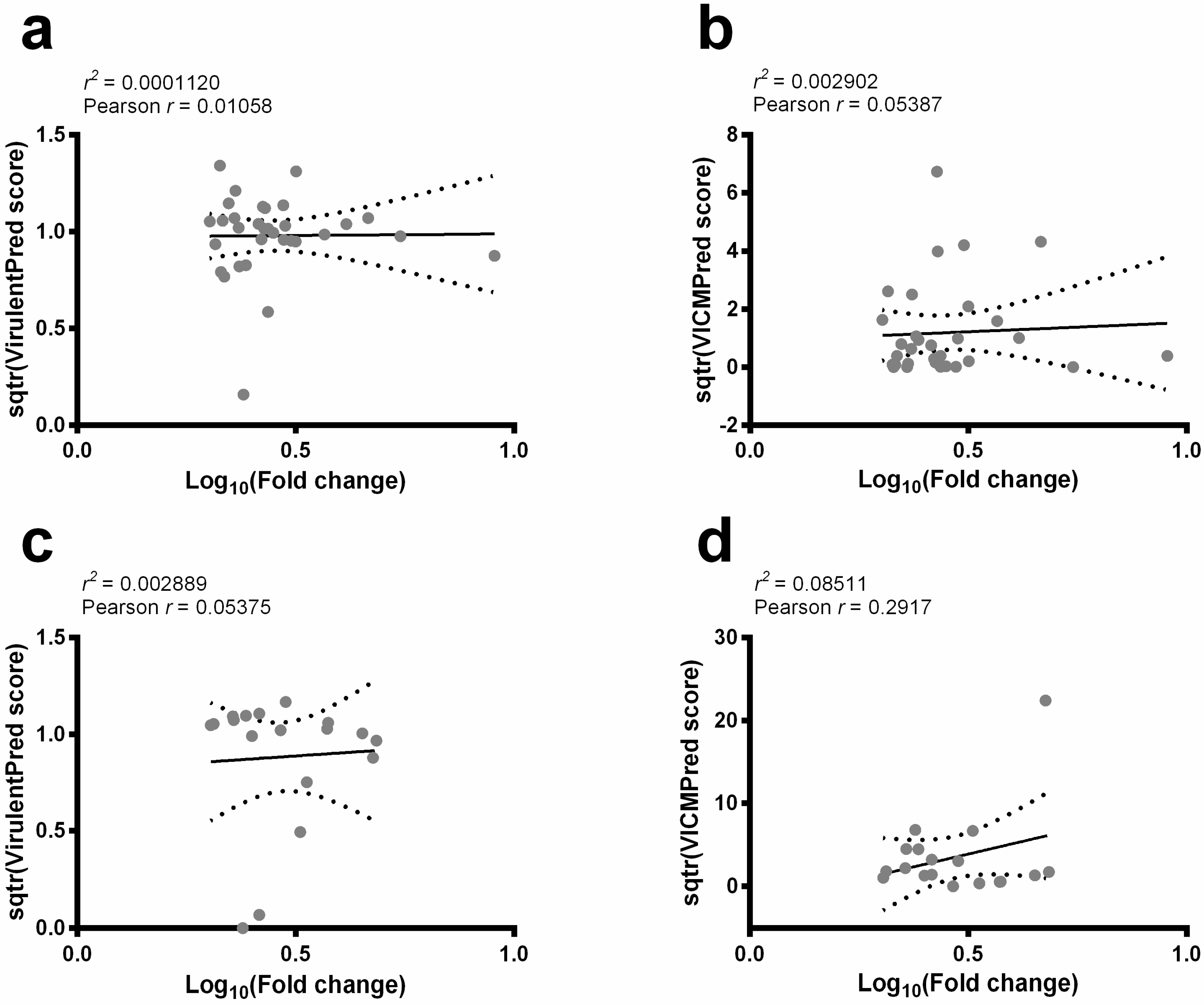
2.1. Computational Strategy to Predict Virulence Factors in L. amazonensis Proteins
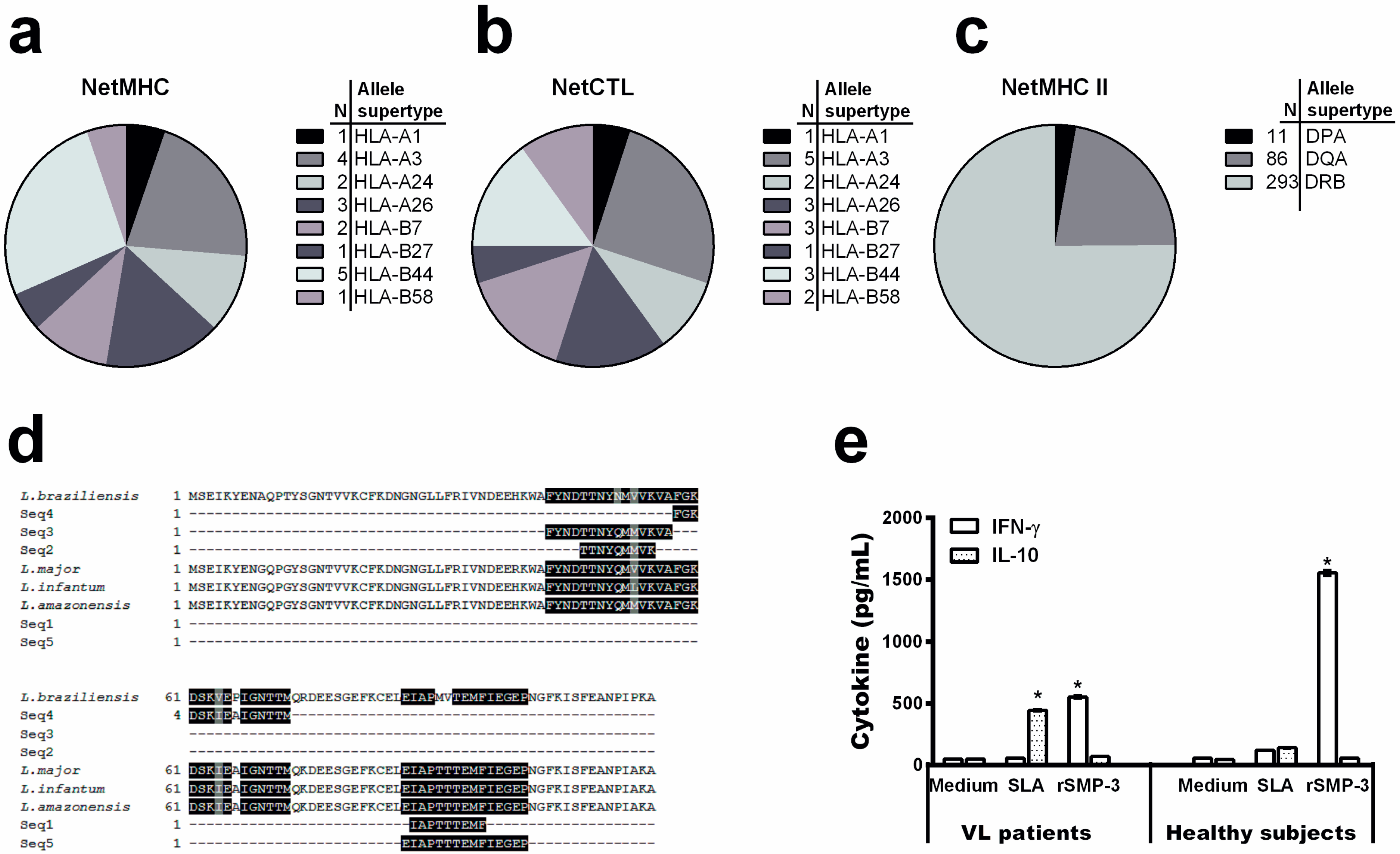
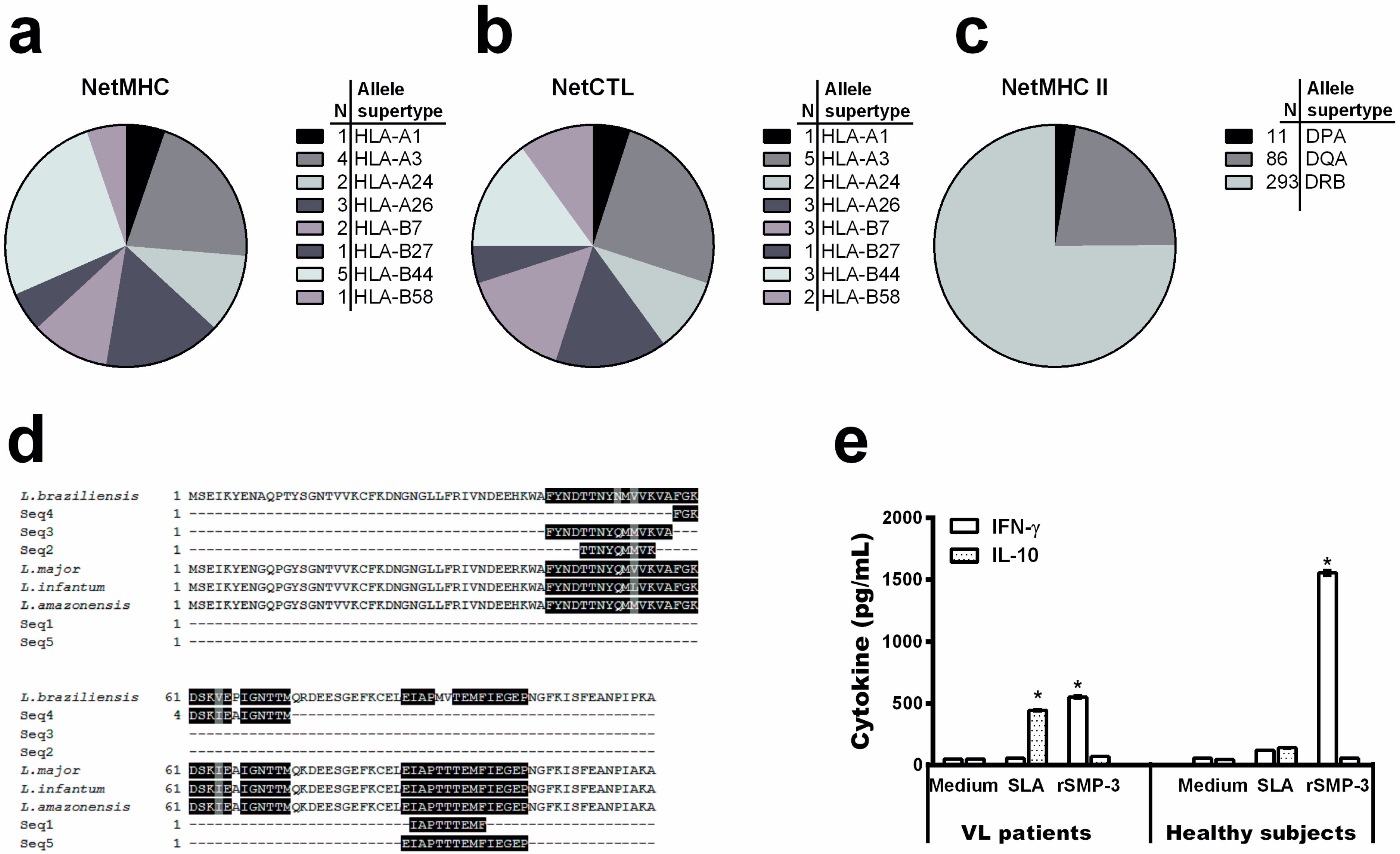
2.2. Bioinformatics Assays and Cytokine Response Using Human PBMCs
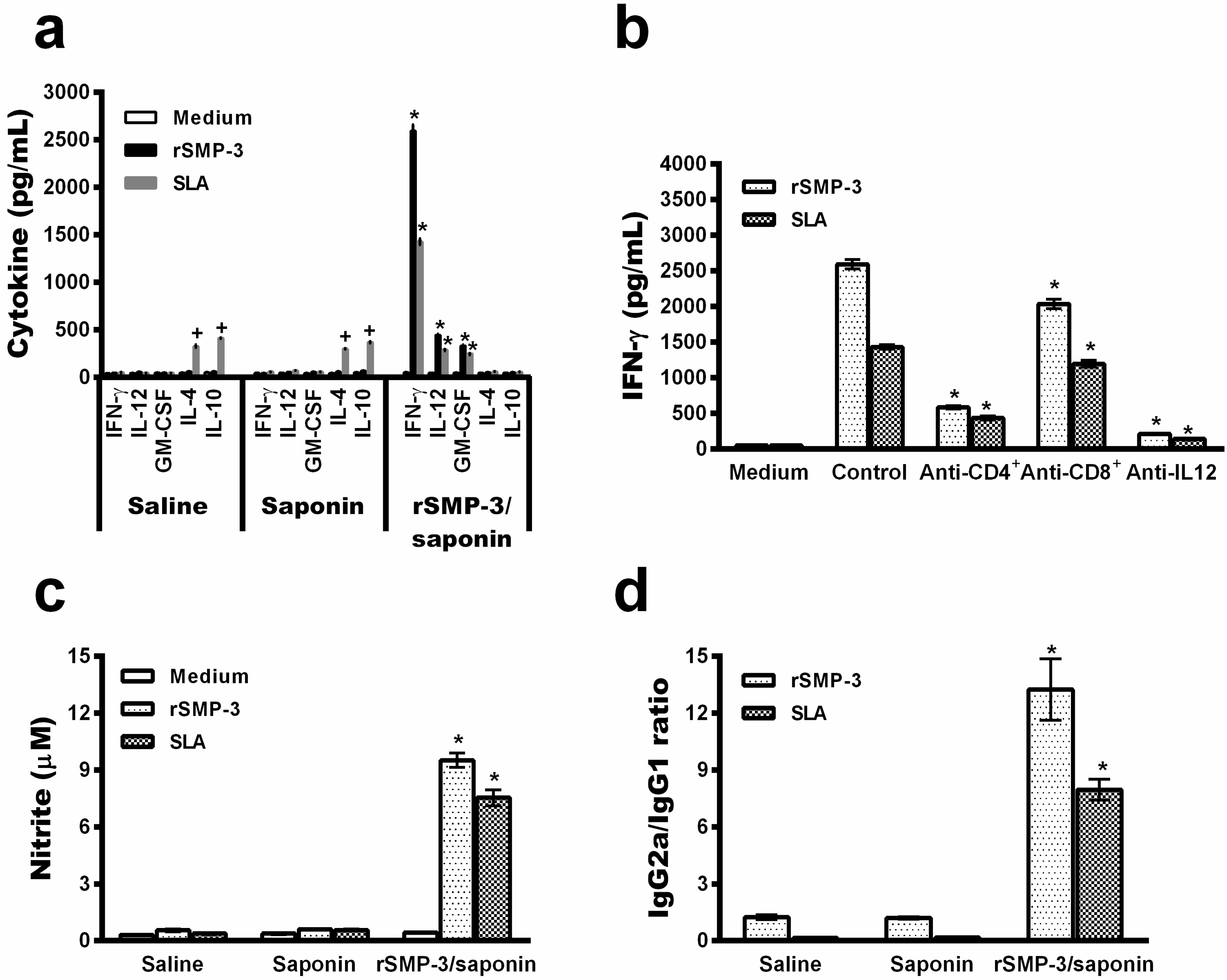
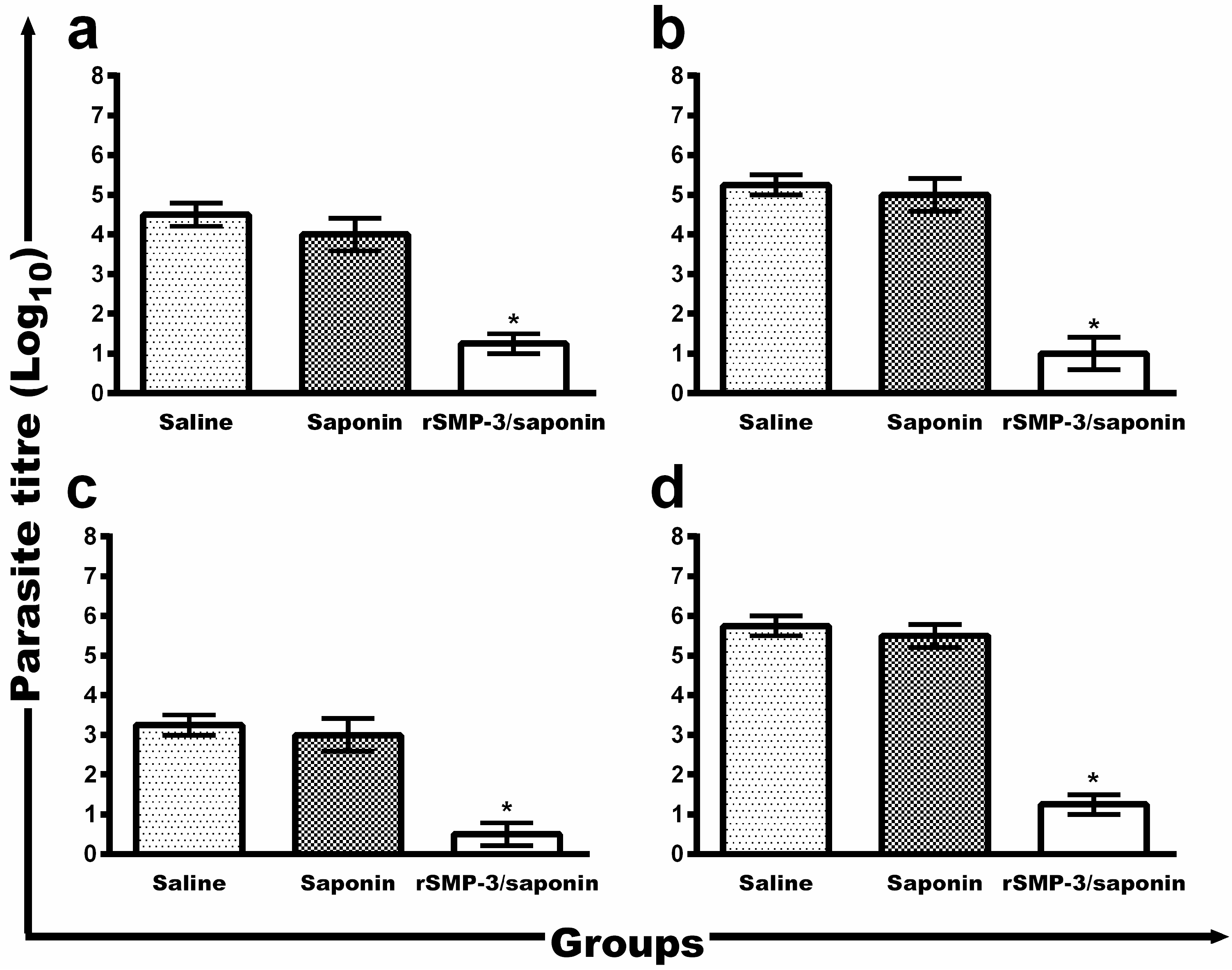
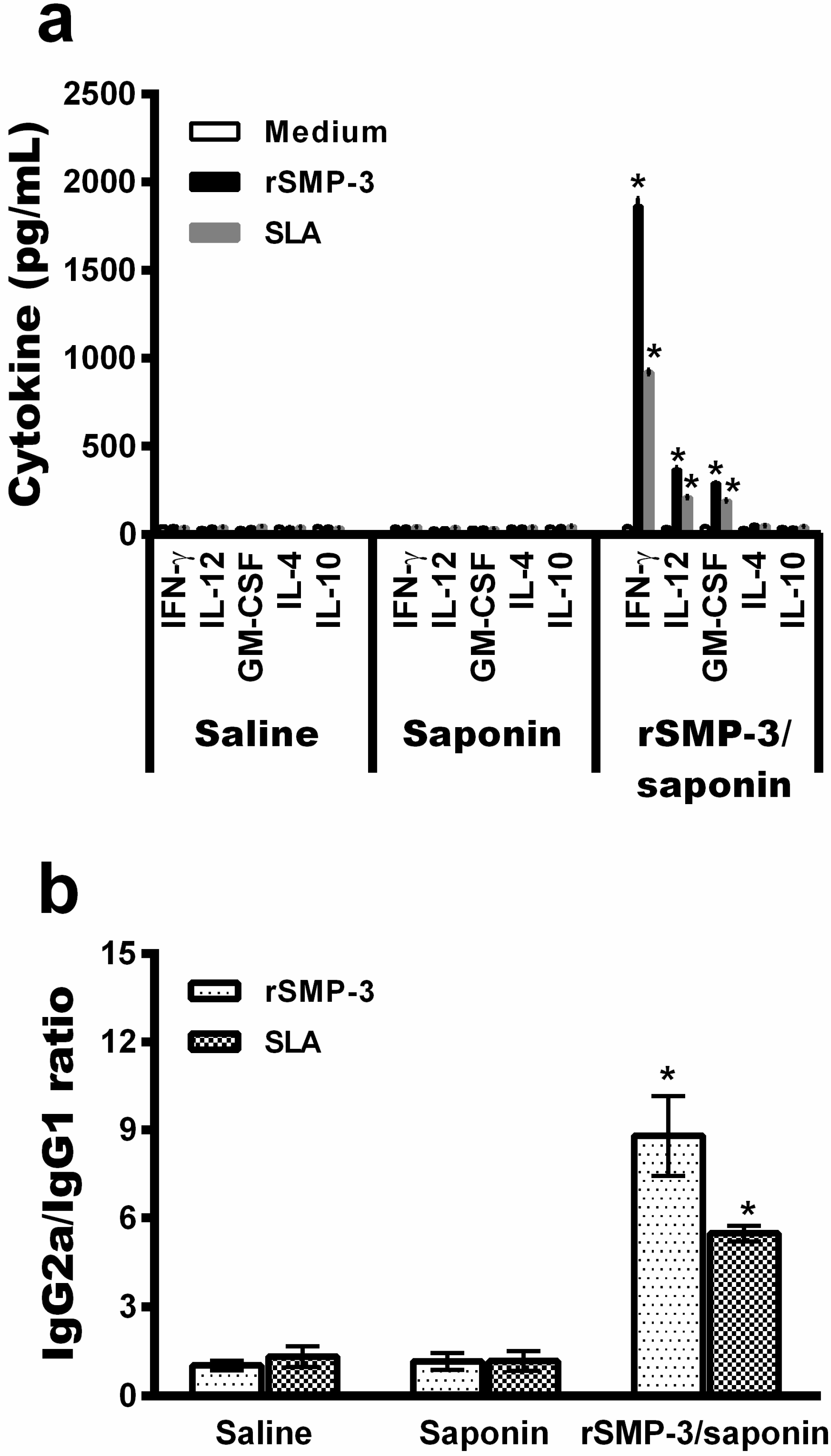
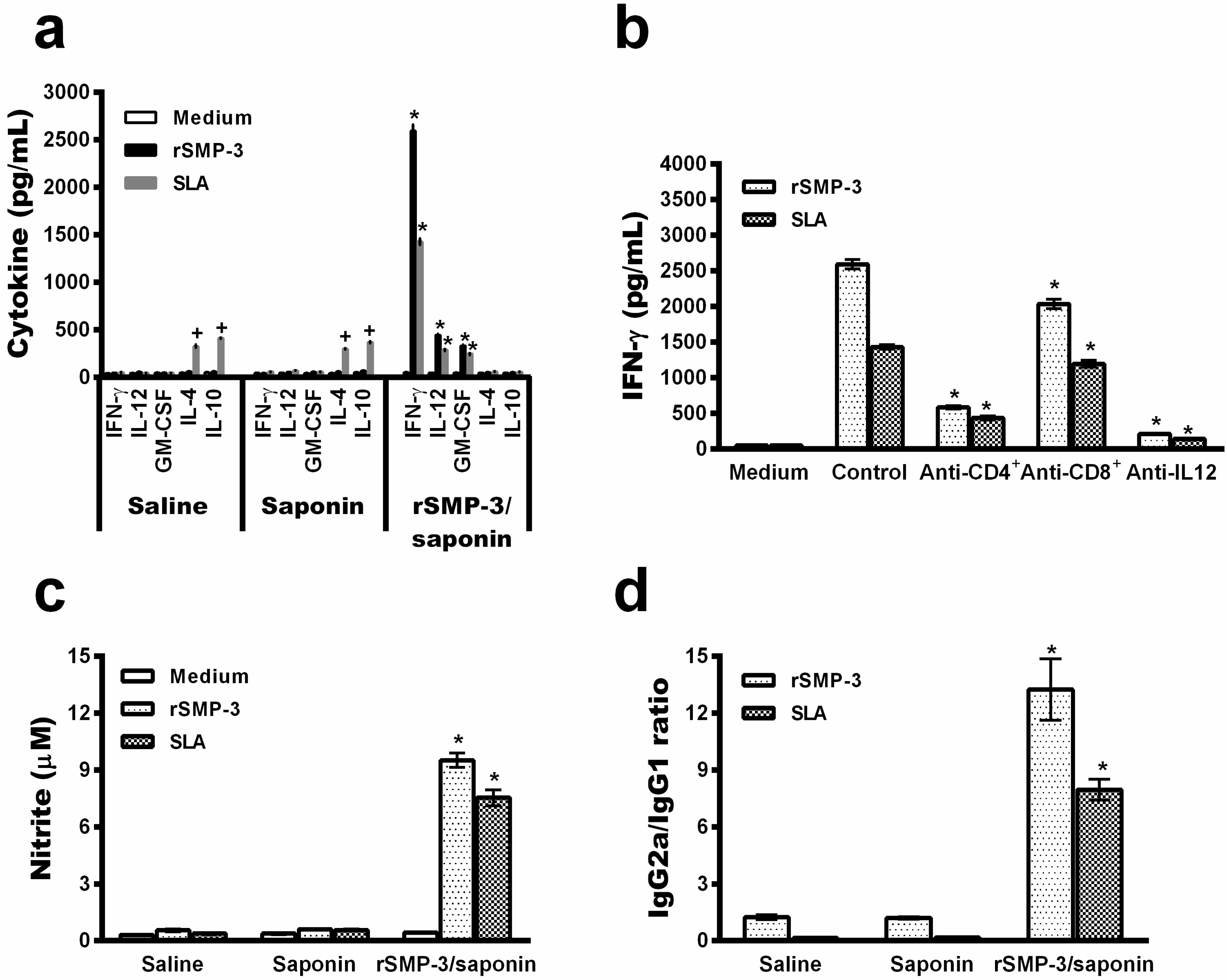
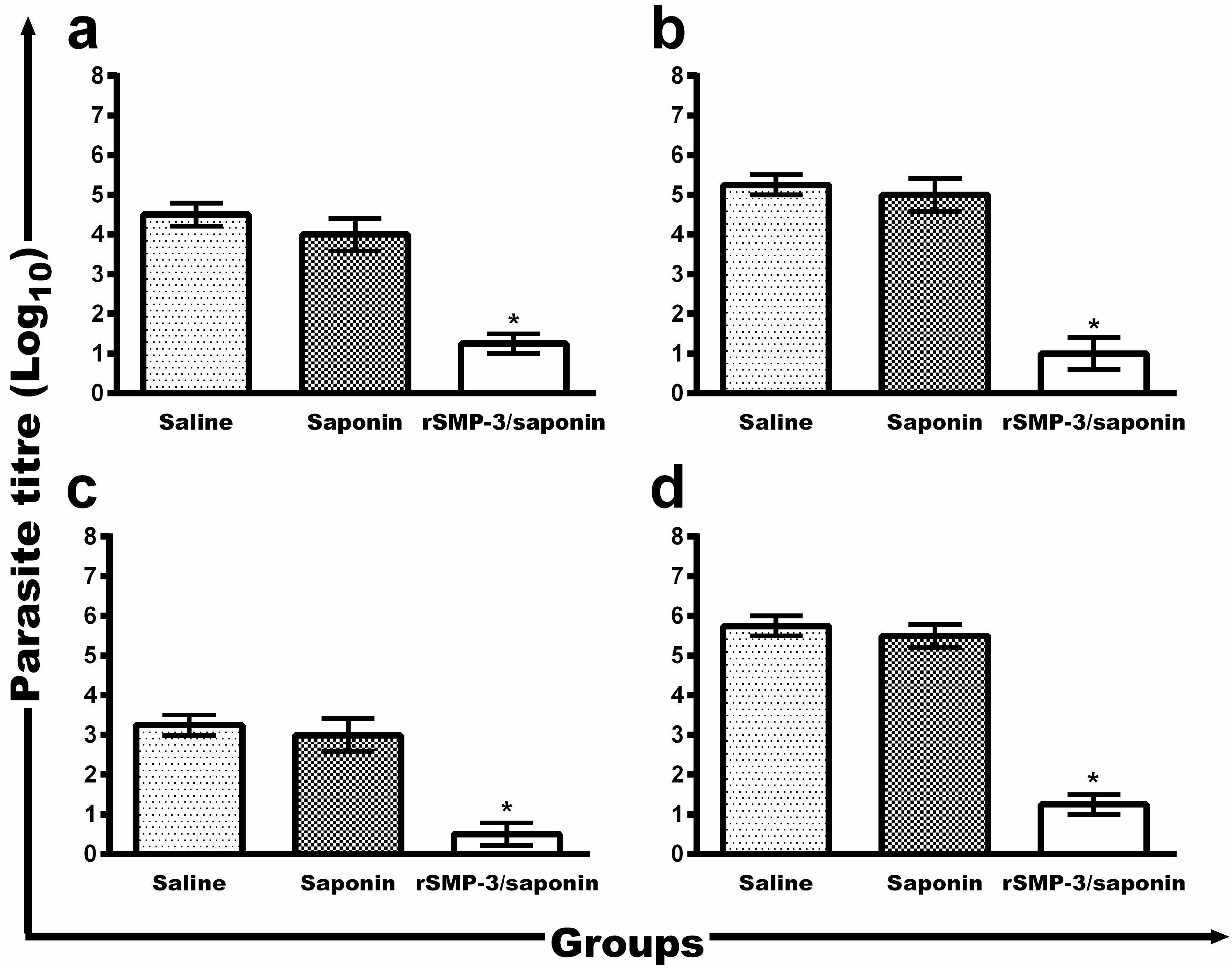
2.3. In Vivo Validation against L. infantum Infection
3. Discussion
4. Materials and Methods
4.1. Sequence Retrieval, Subcellular Localization, and Functional Annotation
4.2. Virulence Factors Prediction
4.3. Protein Sequence Comparison
4.4. Mapping Linear MHC Classes I and II T-Cell Epitopes
4.5. Blood Samples, PBMCs Culture and Cytokine Assay
4.6. Parasites, Mice, and Preparation of the Recombinant Antigen
4.7. Vaccination, Infection, and Cell Response
4.8. Humoral Response
4.9. Evaluation of the Parasite Burden
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; de Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Lockwood, D.N. Clinical aspects of visceral leishmaniasis in HIV infection. Curr. Opin. Infect. Dis. 2013, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Asp. Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Garcez, L.M.; Goto, H.; Ramos, P.K.; Brigido, M.D.C.; Gomes, P.A.F.; Souza, R.A.; De Luca, P.M.; Mendonça, S.C.; Muniz, J.A.P.C.; Shaw, J.J. Leishmania (Leishmania) amazonensis-induced cutaneous leishmaniasis in the primate Cebus apella: A model for vaccine trials. Int. J. Parasitol. 2002, 32, 1755–1764. [Google Scholar] [CrossRef]
- Barral, A.; Pedral-Sampaio, D.; Grimaldi, G.; Momen, H.; McMahon-Pratt, D.; Ribeiro-de-Jesus, A.; Almeida, R.; Badaro, R.; Barral-Netto, M.; Carvalho, E.M.; et al. Leishmaniasis in Bahia, Brazil: Evidence that Leishmania amazonensis produces a wide spectrum of clinical disease. Am. J. Trop. Med. Hyg. 1991, 44, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Shaw, J.J. New World leishmaniasis. In Topley and Wilson’s Microbiology and Microbial Infections; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Elmahallawy, E.K.; Sampedro Martínez, A.; Rodriguez-Granger, J.; Hoyos-Mallecot, Y.; Agil, A.; Navarro Mari, J.M.; Gutierrez Fernández, J. Diagnosis of leishmaniasis. J. Infect. Dev. Ctries. 2014, 8, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Lindoso, J.A.L.; Costa, J.M.L.; Queiroz, I.T.; Goto, H. Review of the current treatments for leishmaniases. Res. Rep. Trop. Med. 2012, 3, 69–77. [Google Scholar]
- Costa, C.H.N.; Peters, N.C.; Maruyama, S.R.; Brito, E.C.; de Miranda Santos, I.K.; Ali, N.; Brodskyn, C.; Campos-Neto, A.; Carvalho, E.M.; Chang, K.P.; et al. Vaccines for the leishmaniases: Proposals for a research agenda. PLoS Negl. Trop. Dis. 2011, 5, e943. [Google Scholar]
- Kedzierski, L. Leishmaniasis. Hum. Vaccin. 2011, 7, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Engwerda, C. Vaccines to prevent leishmaniasis. Clin. Transl. Immunol. 2014, 3, e13. [Google Scholar] [CrossRef] [PubMed]
- Gannavaram, S.; Bhattacharya, P.; Ismail, N.; Kaul, A.; Singh, R.; Nakhasi, H.L. Modulation of innate immune mechanisms to enhance Leishmania vaccine-induced immunity: Role of coinhibitory molecules. Front. Immunol. 2016, 7, 187. [Google Scholar] [CrossRef] [PubMed]
- White, A.C.; Castes, M.; Garcia, L.; Trujillo, D.; Zambrano, L. Leishmania chagasi antigens recognized in cured visceral leishmaniasis and asymptomatic infection. Am. J. Trop. Med. Hyg. 1992, 46, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Nylén, S. Immunobiology of visceral leishmaniasis. Front. Immunol. 2012, 3, 251. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.A.; Taylor, P.; Flower, D.R. Proteomics in vaccinology and immunobiology: An informatics perspective of the immunone. J. Biomed. Biotechnol. 2003, 2003, 267–290. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, R.D.M.; Duarte, M.C.; Mattos, E.C.; Martins, V.T.; Lage, P.S.; Chávez-Fumagalli, M.A.; Lage, D.P.; Menezes-Souza, D.; Régis, W.C.B.; Manso-Alves, M.J.; et al. Identification of differentially expressed proteins from Leishmania amazonensis associated with the loss of virulence of the parasites. PLoS Negl. Trop. Dis. 2014, 8, e2764. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R. Twenty-first century vaccines. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2756–2758. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Miele, A.E.; Ardini, M.; Boumis, G.; Saccoccia, F.; Bellelli, A. Typical 2-Cys peroxiredoxins in human parasites: Several physiological roles for a potential chemotherapy target. Mol. Biochem. Parasitol. 2016, 206, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Pires, S.; Fialho, L.C.; Silva, S.O.; Melo, M.N.; Souza, C.C.; Tafuri, W.L.; Bruna-Romero, O.; Andrade, H.M. Identification of virulence factors in Leishmania infantum strains by a proteomic approach. J. Proteome Res. 2014, 13, 1860–1872. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Samant, M.; Misra, P.; Khare, P.; Sisodia, B.; Shasany, A.K.; Dube, A. Th1-stimulatory polyproteins of soluble Leishmania donovani promastigotes ranging from 89.9 to 97.1 kDa offers long-lasting protection against experimental visceral leishmaniasis. Vaccine 2008, 26, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, P.C.; Velasquez, L.G.; Lepique, A.P.; Rezende, E.; Bonatto, J.M.C.; Barcinski, M.A.; Cunha-Neto, E.; Stolf, B.S. Regulation of Leishmania (L.) amazonensis protein expression by host T cell dependent responses: Differential expression of oligopeptidase B, tryparedoxin peroxidase and HSP70 isoforms in amastigotes isolated from BALB/c and BALB/c nude mice. PLoS Negl. Trop. Dis. 2015, 9, e0003411. [Google Scholar] [CrossRef] [PubMed]
- Grimm, F.; Brun, R.; Jenni, L. Promastigote infectivity in Leishmania infantum. Parasitol. Res. 1991, 77, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Segovia, M.; Artero, J.M.; Mellado, E.; Chance, M.L. Effects of long-term in vitro cultivation on the virulence of cloned lines of Leishmania major promastigotes. Ann. Trop. Med. Parasitol. 1992, 86, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Santarém, N.; Loureiro, I.; Tavares, J.; Silva, A.M.; Amorim, A.M.; Ouaissi, A.; Cordeiro-da-Silva, A.; Silvestre, R. Impact of continuous axenic cultivation in Leishmania infantum virulence. PLoS Negl. Trop. Dis. 2012, 6, e1469. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Reed, S.G.; McGwire, B.S.; Soong, L. Leishmania model for microbial virulence: The relevance of parasite multiplication and pathoantigenicity. Acta Trop. 2003, 85, 375–390. [Google Scholar] [CrossRef]
- Almeida-Silva, M.; Pereira, B.A.; Ribeiro-Guimarães, M.; Alves, C. Proteinases as virulence factors in Leishmania spp. infection in mammals. Parasit. Vectors 2012, 5, 160. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kapil, R.; Dhakan, D.B.; Sharma, V.K. MP3: A software tool for the prediction of pathogenic proteins in genomic and metagenomic data. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Underwood, A.P.; Mulder, A.; Gharbia, S.; Green, J. Virulence Searcher: A tool for searching raw genome sequences from bacterial genomes for putative virulence factors. Clin. Microbiol. Infect. 2005, 11, 770–772. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.T.; Huang, W.L.; Ho, S.J.; Shu, L.S.; Ho, S.Y. Virulent-GO: Prediction of virulent proteins in bacterial pathogens utilizing gene ontology terms. Int. J. Biol. Life Sci. 2009, 5, 159–166. [Google Scholar]
- Saha, S.; Raghava, G.P.S. VICMpred: An SVM-based method for the prediction of functional proteins of gram-negative bacteria using amino acid patterns and composition. Genom. Proteom. Bioinform. 2006, 4, 42–47. [Google Scholar] [CrossRef]
- Garg, A.; Gupta, D. VirulentPred: A SVM based prediction method for virulent proteins in bacterial pathogens. BMC Bioinform. 2008, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.D.O.; Beauchamp, E.; Berthiaume, L.G. Post-translational myristoylation: Fat matters in cellular life and death. Biochimie 2011, 93, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; Price, H.P.; Johner, A.; Emerson, J.E.; Smith, D.F. Kinetoplastid PPEF phosphatases: Dual acylated proteins expressed in the endomembrane system of Leishmania. Mol. Biochem. Parasitol. 2007, 152, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Paape, D.; Storck, E.M.; Serwa, R.A.; Smith, D.F.; Tate, E.W. Global analysis of protein N-myristoylation and exploration of N-myristoyltransferase as a drug target in the neglected human pathogen Leishmania donovani. Chem. Biol. 2015, 22, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Tull, D.; Naderer, T.; Spurck, T.; Mertens, H.D.T.; Heng, J.; McFadden, G.I.; Gooley, P.R.; McConville, M.J. Membrane protein SMP-1 is required for normal flagellum function in Leishmania. J. Cell Sci. 2010, 123, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Udenwobele, D.I.; Su, R.C.; Good, S.V.; Ball, T.B.; Shrivastav, V.S.; Shrivastav, A. Myristoylation: An important protein modification in the immune response. Front. Immunol. 2017, 8, 751. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Heal, W.P.; Mann, D.J.; Tate, E.W. Protein myristoylation in health and disease. J. Chem. Biol. 2010, 3, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Mutiso, J.M.; Macharia, J.C.; Kiio, M.N.; Ichagichu, J.M.; Rikoi, H.; Gicheru, M.M. Development of Leishmania vaccines: Predicting the future from past and present experience. J. Biomed. Res. 2013, 27, 85–102. [Google Scholar] [PubMed]
- Faleiro, R.J.; Kumar, R.; Hafner, L.M.; Engwerda, C.R. Immune regulation during chronic visceral leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Coelho, E.A.F.; Tavares, C.A.P.; Carvalho, F.A.A.; Chaves, K.F.; Teixeira, K.N.; Rodrigues, R.C.; Charest, H.; Matlashewski, G.; Gazzinelli, R.T.; Fernandes, A.P. Immune responses induced by the Leishmania (Leishmania) donovani A2 antigen, but not by the LACK antigen, are protective against experimental Leishmania (Leishmania) amazonensis infection. Infect. Immun. 2003, 71, 3988–3994. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.P.; Costa, M.M.S.; Coelho, E.A.F.; Michalick, M.S.M.; Freitas, E.; Melo, M.N.; Tafuri, W.L.; Resende, D.D.M.; Hermont, V.; Abrantes, C.D.F.; et al. Protective immunity against challenge with Leishmania (Leishmania) chagasi in beagle dogs vaccinated with recombinant A2 protein. Vaccine 2008, 26, 5888–5895. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, G.; Teva, A.; Porrozzi, R.; Pinto, M.A.; Marchevsky, R.S.; Rocha, M.G.L.; Dutra, M.S.; Bruña-Romero, O.; Fernandes, A.P.; Gazzinelli, R.T. Clinical and parasitological protection in a Leishmania infantum-macaque model vaccinated with adenovirus and the recombinant A2 antigen. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Najera, C.; Piña-Aguilar, R.; Xacur-Garcia, F.; Ramirez-Sierra, M.J.; Dumonteil, E. Mining the Leishmania genome for novel antigens and vaccine candidates. Proteomics 2009, 9, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Resende, D.M.; Rezende, A.M.; Oliveira, N.J.; Batista, I.C.; Corrêa-Oliveira, R.; Reis, A.B.; Ruiz, J.C. An assessment on epitope prediction methods for protozoa genomes. BMC Bioinform. 2012, 13, 309. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, P.M.; Beaumier, C.M.; Strych, U.; Hayward, T.; Hotez, P.J.; Bottazzi, M.E. Status of vaccine research and development of vaccines for leishmaniasis. Vaccine 2016, 34, 2992–2995. [Google Scholar] [CrossRef] [PubMed]
- Rezende, A.M.; Folador, E.L.; Resende, D.D.M.; Ruiz, J.C. Computational prediction of protein-protein interactions in Leishmania predicted proteomes. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Brito, R.; Guimarães, F.; Velloso, J.; Corrêa-Oliveira, R.; Ruiz, J.; Reis, A.; Resende, D. Immunoinformatics features linked to Leishmania vaccine development: Data integration of experimental and in silico studies. Int. J. Mol. Sci. 2017, 18, 371. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Silva, R.; Ferreira, L.F.G.R.; Hernandes, M.Z.; Brito, M.E.F.; Oliveira, B.C.; Silva, A.A.; Melo-Neto, O.P.; Rezende, A.M.; Pereira, V.R.A. Combination of in silico methods in the search for potential CD4+ and CD8+ T cell epitopes in the proteome of Leishmania braziliensis. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Mansueto, P.; Vitale, G.; Di Lorenzo, G.; Rini, G.B.; Mansueto, S.; Cillari, E. Immunopathology of leishmaniasis: An update. Int. J. Immunopathol. Pharmacol. 2007, 20, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.; Teixeira, C.; Teixeira, M.J.; Oliveira, F.; Menezes, M.J.; Silva, C.; Oliveira, C.I.; Miranda, J.C.; Elnaiem, D.E.; Kamhawi, S.; et al. Immunity to a salivary protein of a sand fly vector protects against the fatal outcome of visceral leishmaniasis in a hamster model. Proc. Natl. Acad. Sci. USA 2008, 105, 7845–7850. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Fumagalli, M.A.; Costa, M.A.; Oliveira, D.M.; Ramírez, L.; Costa, L.E.; Duarte, M.C.; Martins, V.T.; Oliveira, J.S.; Olortegi, C.C.; Bonay, P.; et al. Vaccination with the Leishmania infantum ribosomal proteins induces protection in BALB/c mice against Leishmania chagasi and Leishmania amazonensis challenge. Microbes Infect. 2010, 12, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Carrión, J.; Nieto, A.; Iborra, S.; Iniesta, V.; Soto, M.; Folgueira, C.; Abanades, D.R.; Requena, J.M.; Alonso, C. Immunohistological features of visceral leishmaniasis in BALB/c mice. Parasite Immunol. 2006, 28, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Kaur, H.; Kaur, S. Evaluation of the immunoprophylactic potential of a killed vaccine candidate in combination with different adjuvants against murine visceral leishmaniasis. Parasitol. Int. 2015, 64, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.M.; Valadares, D.G.; Duarte, M.C.; Costa, L.E.; Martins, V.T.; Gomes, R.F.; Melo, M.N.; Soto, M.; Tavares, C.A.P.; Coelho, E.A.F. Evaluation of parasitological and immunological parameters of Leishmania chagasi infection in BALB/c mice using different doses and routes of inoculation of parasites. Parasitol. Res. 2012, 110, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Colmenares, M.; Soong, L.; Goldsmith-Pestana, K.; Munstermann, L.; Molina, R.; McMahon-Pratt, D. Intradermal infection model for pathogenesis and vaccine studies of murine visceral leishmaniasis. Infect. Immun. 2003, 71, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Mizbani, A.; Taheri, T.; Zahedifard, F.; Taslimi, Y.; Azizi, H.; Azadmanesh, K.; Papadopoulou, B.; Rafati, S. Recombinant Leishmania tarentolae expressing the A2 virulence gene as a novel candidate vaccine against visceral leishmaniasis. Vaccine 2009, 28, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.; Karmakar, S.; De, T. TLR-mediated distinct IFN-γ/IL-10 pattern induces protective immunity against murine visceral leishmaniasis. Eur. J. Immunol. 2012, 42, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.L.; Wille, U.; Villegas, E.N.; Hunter, C.A.; Farrell, J.P. IL-10 mediates susceptibility to Leishmania donovani infection. Eur. J. Immunol. 2001, 31, 2848–2856. [Google Scholar] [CrossRef]
- Murray, H.W.; Lu, C.M.; Mauze, S.; Freeman, S.; Moreira, A.L.; Kaplan, G.; Coffman, R.L. Interleukin-10 (IL-10) in experimental visceral leishmaniasis and IL-10 receptor blockade as immunotherapy. Infect. Immun. 2002, 70, 6284–6293. [Google Scholar] [CrossRef] [PubMed]
- Padigel, U.M.; Alexander, J.; Farrell, J.P.; Padigel, U.M.; Alexander, J.; Farrell, J.P. The role of interleukin-10 in susceptibility of BALB/c mice to infection with Leishmania mexicana and Leishmania amazonensis. J. Immunol. 2003, 171, 3705–3710. [Google Scholar] [CrossRef] [PubMed]
- Singh, O.P.; Hasker, E.; Sacks, D.; Boelaert, M.; Sundar, S. Asymptomatic Leishmania infection: A new challenge for Leishmania control. Clin. Infect. Dis. 2014, 58, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Topno, R.K.; Das, V.N.R.; Ranjan, A.; Pandey, K.; Singh, D.; Kumar, N.; Siddiqui, N.A.; Singh, V.P.; Kesari, S.; Kumar, N.; et al. Asymptomatic infection with visceral leishmaniasis in a disease-endemic area in Bihar, India. Am. J. Trop. Med. Hyg. 2010, 83, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Fumagalli, M.A.; Schneider, M.S.; Lage, D.P.; Machado-de-Ávila, R.A.; Coelho, E.A.F. An in silico functional annotation and screening of potential drug targets derived from Leishmania spp. hypothetical proteins identified by immunoproteomics. Exp. Parasitol. 2017, 176, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.uniprot.org/.
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Cenk-Sahinalp, S.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, M.; Garg, A.; Raghava, G.P.S. PSLpred: Prediction of subcellular localization of bacterial proteins. Bioinformatics 2005, 21, 2522–2524. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-S.; Lin, C.-J.; Hwang, J.-K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; Von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. 2004, 17, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Pethica, R.; Zhou, Y.; Talbot, C.; Vogel, C.; Madera, M.; Chothia, C.; Gough, J. SUPERFAMILY—Sophisticated comparative genomics, data mining, visualization and phylogeny. Nucleic Acids Res. 2009, 37, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Sillitoe, I.; Lewis, T.E.; Cuff, A.; Das, S.; Ashford, P.; Dawson, N.L.; Furnham, N.; Laskowski, R.A.; Lee, D.; Lees, J.G.; et al. CATH: Comprehensive structural and functional annotations for genome sequences. Nucleic Acids Res. 2015, 43, D376–D381. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Lee, C.S.; Van Gelder, R.N.; Comin, M.; Leoni, A.; Schimd, M.; Lindner, M.; Renard, B.; Benson, D.; Karsch-Mizrachi, I.; et al. Scalable metagenomics alignment research tool (SMART): A scalable, rapid, and complete search heuristic for the classification of metagenomic sequences from complex sequence populations. BMC Bioinform. 2016, 17, 292. [Google Scholar] [CrossRef] [PubMed]
- Rappoport, N.; Karsenty, S.; Stern, A.; Linial, N.; Linial, M. ProtoNet 6.0: Organizing 10 million protein sequences in a compact hierarchical family tree. Nucleic Acids Res. 2012, 40, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.-L.; Li, Y.-X.; Ding, J.; Guo, X.-K.; Feng, K.-Y.; Wang, Y.-J.; Hu, L.-L.; Cai, Y.-D.; Hao, P.; Chou, K.-C. A comparison of computational methods for identifying virulence factors. PLoS ONE 2012, 7, e42517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://tcoffee.crg.cat/.
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; Mc William, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, M.; Nielsen, M. Gapped sequence alignment using artificial neural networks: Application to the MHC class I system. Bioinformatics 2015, 32, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Lundegaard, C.; Lamberth, K.; Buus, S.; Lund, O.; Nielsen, M. Large-scale validation of methods for cytotoxic T-lymphocyte epitope prediction. BMC Bioinform. 2007, 8, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.; Lundegaard, C.; Lund, O. Prediction of MHC class II binding affinity using SMM-align, a novel stabilization matrix alignment method. BMC Bioinform. 2007, 8, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://www.ncbi.nlm.nih.gov/gv/mhc.
- Fleri, W.; Paul, S.; Dhanda, S.K.; Mahajan, S.; Xu, X.; Peters, B.; Sette, A. The immune epitope database and analysis resource in epitope discovery and synthetic vaccine design. Front. Immunol. 2017, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.T.; Chávez-Fumagalli, M.A.; Lage, D.P.; Duarte, M.C.; Garde, E.; Costa, L.E.; Silva, V.G.; Oliveira, J.S.; Magalhães-Soares, D.F.; Teixeira, S.M.R.; et al. Antigenicity, immunogenicity and protective efficacy of three proteins expressed in the promastigote and amastigote stages of Leishmania infantum against visceral leishmaniasis. PLoS ONE 2015, 10, e0137683. [Google Scholar] [CrossRef]
- Santos, T.T.O.; Martins, V.T.; Lage, D.P.; Costa, L.E.; Salles, B.C.S.; Carvalho, A.M.R.S.; Dias, D.S.; Ribeiro, P.A.F.; Chavez-Fumagalli, M.A.; Machado-de-Avila, R.A.; et al. Probing the efficacy of a heterologous Leishmania/L. Viannia braziliensis recombinant enolase as a candidate vaccine to restrict the development of L. infantum in BALB/c mice. Acta Trop. 2017, 171, 8–16. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uniprot ID | Gene ID | Predicted Function | Predicted Localization |
|---|---|---|---|
| E9ASM0 | LMXM36.1000 | WD40 repeat-like | Cytoplasmic |
| E9AXT3 | LMXM25.2010 | Phosphoenol pyruvate-like | Cytoplasmic |
| E9B549 | LMXM33.2580 | AlbA-like | Cytoplasmic |
| E9ATK7 | LMXM36.4230 | Leucine rich-like | Cytoplasmic |
| Uniprot ID | Protein Name | Gene ID | Major HHpred Hit to Homo sapien Sequences | ||
|---|---|---|---|---|---|
| PDB | Probability (%) | E-Value | |||
| E9ATK7 | WD40 repeat-like protein a | LMXM36.4230 | 1Z7X | 100 | 4.3 × 10−29 |
| E9AQ29 | Ribonucleoprotein p18 | LMXM15.0275 | 4XGL | 99.8 | 1 × 10−17 |
| E9APT0 | Putative small myristoylated protein-3 | LMXM14.0850 | 2JX8 | 45.1 | 27 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, M.P.; Martins, V.T.; Santos, T.T.O.; Lage, D.P.; Ramos, F.F.; Salles, B.C.S.; Costa, L.E.; Dias, D.S.; Ribeiro, P.A.F.; Schneider, M.S.; et al. Small Myristoylated Protein-3, Identified as a Potential Virulence Factor in Leishmania amazonensis, Proves to be a Protective Antigen against Visceral Leishmaniasis. Int. J. Mol. Sci. 2018, 19, 129. https://doi.org/10.3390/ijms19010129
Oliveira MP, Martins VT, Santos TTO, Lage DP, Ramos FF, Salles BCS, Costa LE, Dias DS, Ribeiro PAF, Schneider MS, et al. Small Myristoylated Protein-3, Identified as a Potential Virulence Factor in Leishmania amazonensis, Proves to be a Protective Antigen against Visceral Leishmaniasis. International Journal of Molecular Sciences. 2018; 19(1):129. https://doi.org/10.3390/ijms19010129
Chicago/Turabian StyleOliveira, Marcelo P., Vívian T. Martins, Thaís T. O. Santos, Daniela P. Lage, Fernanda F. Ramos, Beatriz C. S. Salles, Lourena E. Costa, Daniel S. Dias, Patrícia A. F. Ribeiro, Mônica S. Schneider, and et al. 2018. "Small Myristoylated Protein-3, Identified as a Potential Virulence Factor in Leishmania amazonensis, Proves to be a Protective Antigen against Visceral Leishmaniasis" International Journal of Molecular Sciences 19, no. 1: 129. https://doi.org/10.3390/ijms19010129