Overall Survival of Ovarian Cancer Patients Is Determined by Expression of Galectins-8 and -9

 and
and
Abstract
:
1. Introduction
2. Results
2.1. In Silico Analysis of Gal-8 and -9 Expression in Normal Ovarian Tissues and Ovarian Cancer
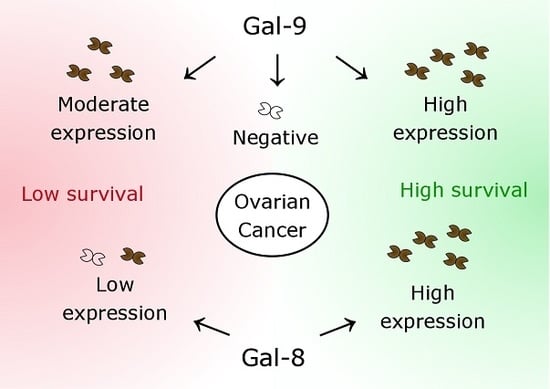
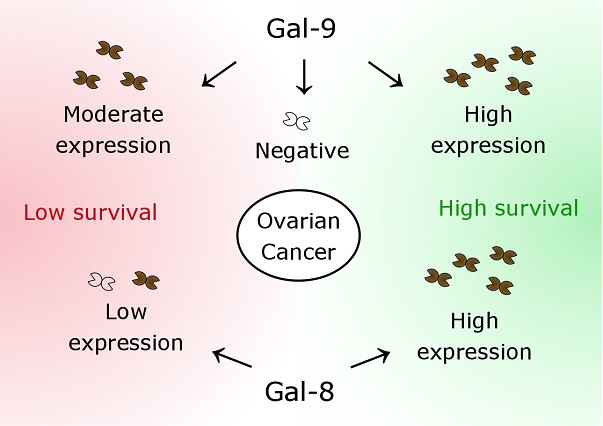
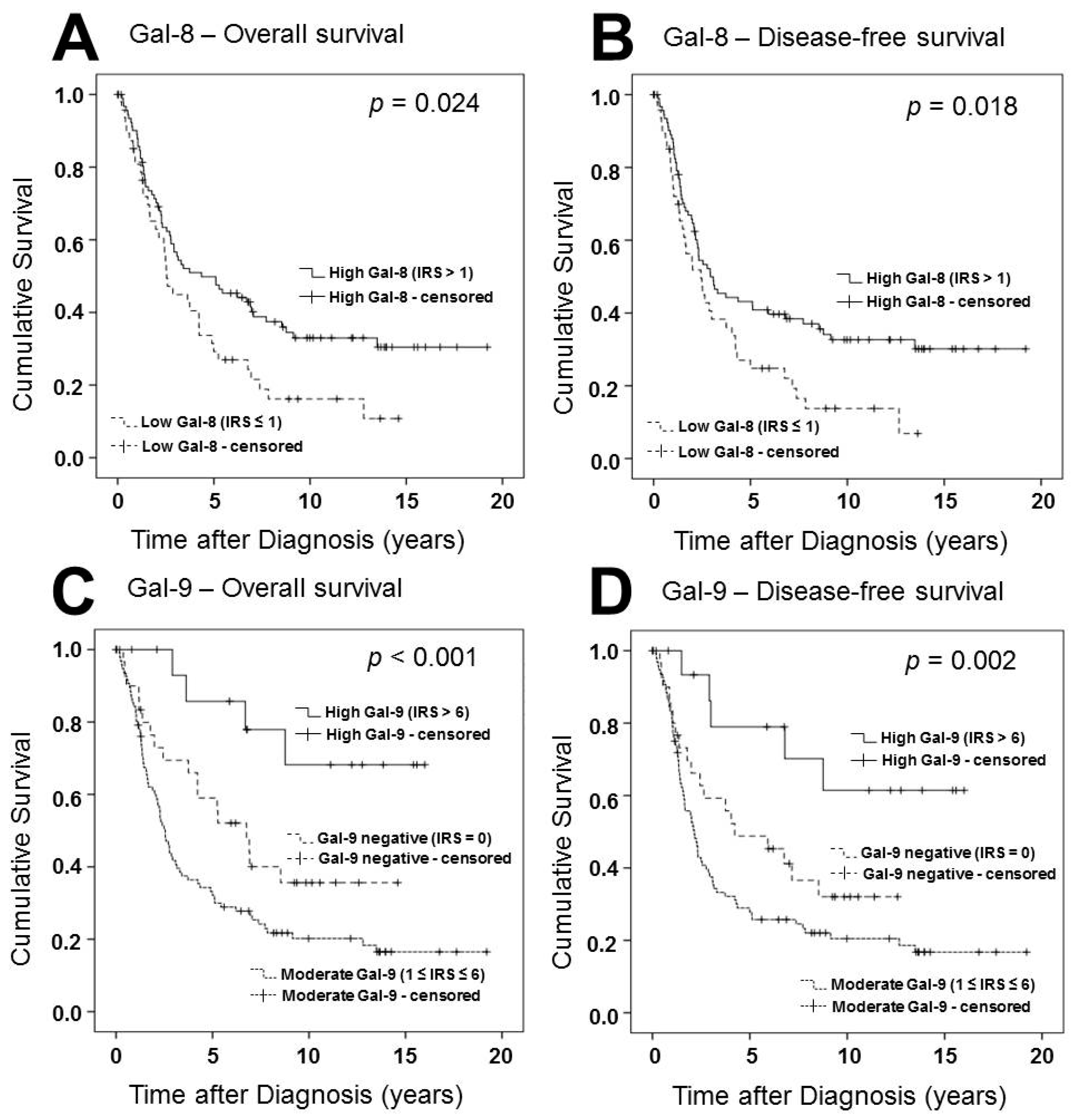
2.2. Gal-8 is a Positive Prognostic Factor for OS and DFS in Ovarian Cancer Patients
2.3. Gal-9 Expression Determines DFS and OS of Ovarian Cancer Patients in Two Different Ways
2.4. Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Patients
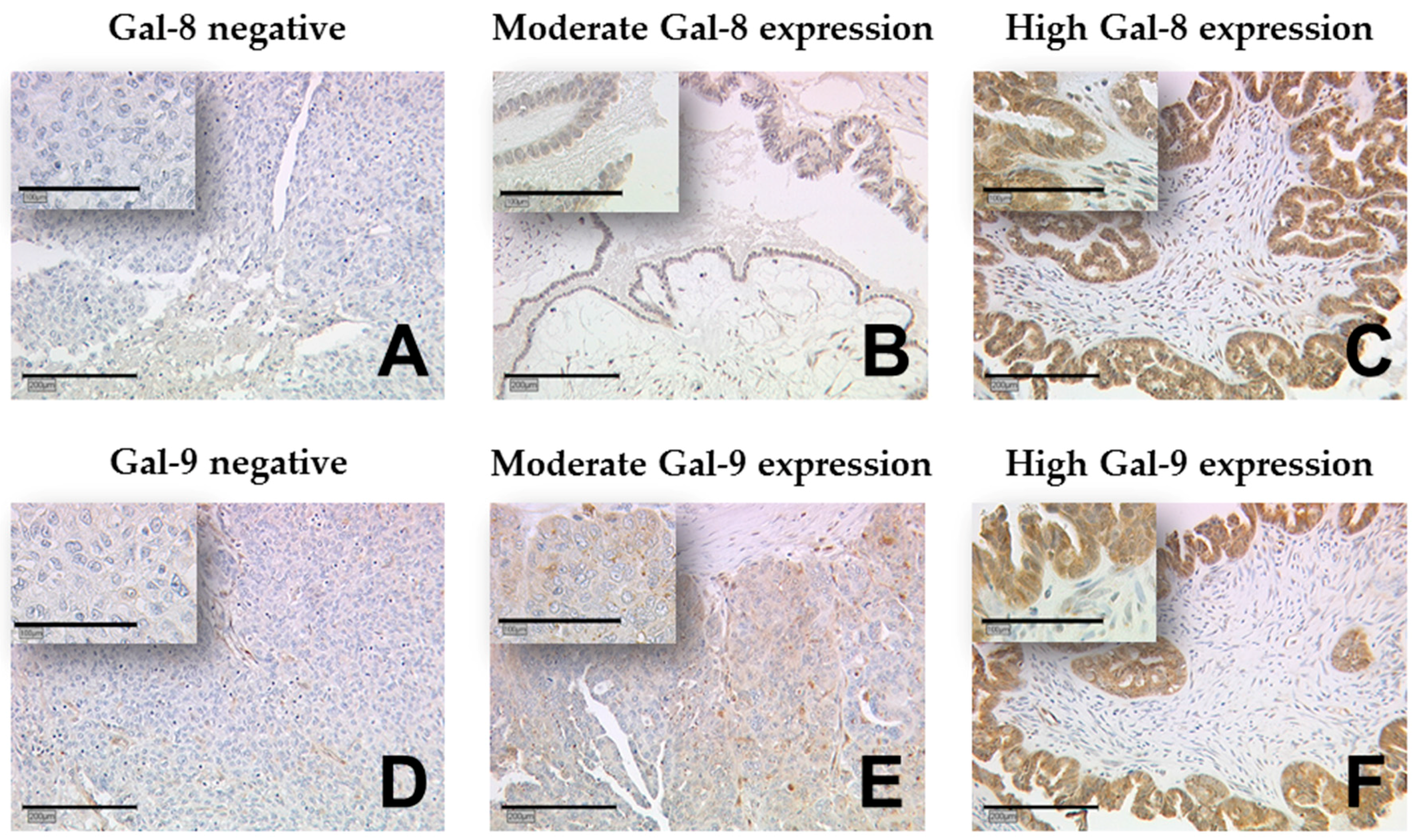
4.2. Immunohistochemistry
4.3. Statistical Analysis
4.4. Ethics Statement
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Gal | Galectin |
| IHC | Immunohistochemistry |
| IRS | Immunoreactivity score |
| DFS | Disease-free survival |
| OS | Overall survival |
| BRCA | Breast cancer gene |
| CRD | Carbohydrate-recognition domain |
| LGALS8 | Human Gal-8 gene |
| LGALS9 | Human Gal-9 gene |
| FIGO | Fédération Internationale de Gynécologie et d’Obstétrique |
| UICC | Union for International Cancer Control |
| PCTA-1 | Prostate carcinoma tumor antigen-1 |
| CD8 | cluster of differentiation 8 |
| CD4 | cluster of differentiation 4 |
| TIM3 | T-cell immunoglobulin and mucin-domain containing-3 |
| T reg | T-regulatory |
| TMA | Tissue micro array |
| FFPE | Formalin-fixed paraffin-embedded |
| WHO | World Health Organization |
| TNM | T = tumor, N = lymph nodes, M = metastasis |
| PBS | phosphate buffered saline |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Thibault, B.; Castells, M.; Delord, J.P.; Couderc, B. Ovarian cancer microenvironment: Implications for cancer dissemination and chemoresistance acquisition. Cancer Metastasis Rev. 2014, 33, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, L.A.; Huang, B.; Miller, R.W.; Tucker, T.; Goodrich, S.T.; Podzielinski, I.; DeSimone, C.P.; Ueland, F.R.; van Nagell, J.R.; Seamon, L.G. Ten-year relative survival for epithelial ovarian cancer. Obstet. Gynecol. 2012, 120, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Trope, C.G. Ovarian cancer: Diagnostic, biological and prognostic aspects. Womens Health (Lond.) 2014, 10, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H.; Castronovo, V.; Cooper, D.N.; Cummings, R.D.; Drickamer, K.; Feizi, T.; Gitt, M.A.; Hirabayashi, J.; Hughes, C.; Kasai, K.; et al. Galectins: A family of animal beta-galactoside-binding lectins. Cell 1994, 76, 597–598. [Google Scholar] [CrossRef]
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [PubMed]
- Ebrahim, A.H.; Alalawi, Z.; Mirandola, L.; Rakhshanda, R.; Dahlbeck, S.; Nguyen, D.; Jenkins, M.; Grizzi, F.; Cobos, E.; Figueroa, J.A.; et al. Galectins in cancer: Carcinogenesis, diagnosis and therapy. Ann. Transl. Med. 2014, 2, 88. [Google Scholar] [PubMed]
- Leffler, H.; Carlsson, S.; Hedlund, M.; Qian, Y.; Poirier, F. Introduction to galectins. Glycoconj. J. 2002, 19, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Rabinovich, G.A.; Liu, F.T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Rabinovich, G.A. Galectins as modulators of tumour progression. Nat. Rev. Cancer 2005, 5, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Grosset, A.A.; Labrie, M.; Vladoiu, M.C.; Yousef, E.M.; Gaboury, L.; St-Pierre, Y. Galectin signatures contribute to the heterogeneity of breast cancer and provide new prognostic information and therapeutic targets. Oncotarget 2016, 7, 18183–18203. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Jin, M.S.; Kong, F.; Cao, D.; Ma, H.X.; Jia, Z.; Wang, Y.P.; Suo, J.; Cao, X. Decreased galectin-9 and increased tim-3 expression are related to poor prognosis in gastric cancer. PLoS ONE 2013, 8, e81799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Shi, B.; Zhou, M.; Jiang, H.; Zhang, H.; Pan, X.; Gao, H.; Sun, H.; Li, Z. Galectin-1 overexpression promotes progression and chemoresistance to cisplatin in epithelial ovarian cancer. Cell Death Dis. 2014, 5, e991. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeon, H.K.; Cho, Y.J.; Park, Y.A.; Choi, J.J.; Do, I.G.; Song, S.Y.; Lee, Y.Y.; Choi, C.H.; Kim, T.J.; et al. High galectin-1 expression correlates with poor prognosis and is involved in epithelial ovarian cancer proliferation and invasion. Eur. J. Cancer 2012, 48, 1914–1921. [Google Scholar] [CrossRef] [PubMed]
- Mirandola, L.; Yu, Y.; Cannon, M.J.; Jenkins, M.R.; Rahman, R.L.; Nguyen, D.D.; Grizzi, F.; Cobos, E.; Figueroa, J.A.; Chiriva-Internati, M. Galectin-3 inhibition suppresses drug resistance, motility, invasion and angiogenic potential in ovarian cancer. Gynecol. Oncol. 2014, 135, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Brustmann, H. Epidermal growth factor receptor expression in serous ovarian carcinoma: An immunohistochemical study with galectin-3 and cyclin d1 and outcome. Int. J. Gynecol. Pathol. 2008, 27, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeon, H.K.; Lee, J.K.; Sung, C.O.; Do, I.G.; Choi, C.H.; Kim, T.J.; Kim, B.G.; Bae, D.S.; Lee, J.W. Clinical significance of galectin-7 in epithelial ovarian cancer. Anticancer Res. 2013, 33, 1555–1561. [Google Scholar] [PubMed]
- Labrie, M.; Vladoiu, M.C.; Grosset, A.A.; Gaboury, L.; St-Pierre, Y. Expression and functions of galectin-7 in ovarian cancer. Oncotarget 2014, 5, 7705–7721. [Google Scholar] [CrossRef] [PubMed]
- Schulz, H.; Schmoeckel, E.; Kuhn, C.; Hofmann, S.; Mayr, D.; Mahner, S.; Jeschke, U. Galectins-1, -3, and -7 are prognostic markers for survival of ovarian cancer patients. Int. J. Mol. Sci. 2017, 18, 1230. [Google Scholar] [CrossRef] [PubMed]
- Labrie, M.; De Araujo, L.O.F.; Communal, L.; Mes-Masson, A.M.; St-Pierre, Y. Tissue and plasma levels of galectins in patients with high grade serous ovarian carcinoma as new predictive biomarkers. Sci. Rep. 2017, 7, 13244. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Hadari, Y.R.; Paz, K.; Dekel, R.; Mestrovic, T.; Accili, D.; Zick, Y. Galectin-8. A new rat lectin, related to galectin-4. J. Biol. Chem. 1995, 270, 3447–3453. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Z.; Lin, J.; Shen, R.; Fisher, P.E.; Goldstein, N.I.; Fisher, P.B. Surface-epitope masking and expression cloning identifies the human prostate carcinoma tumor antigen gene pcta-1 a member of the galectin gene family. Proc. Natl. Acad. Sci. USA 1996, 93, 7252–7257. [Google Scholar] [CrossRef] [PubMed]
- Nishi, N.; Itoh, A.; Shoji, H.; Miyanaka, H.; Nakamura, T. Galectin-8 and galectin-9 are novel substrates for thrombin. Glycobiology 2006, 16, 15C–20C. [Google Scholar] [CrossRef] [PubMed]
- Troncoso, M.F.; Ferragut, F.; Bacigalupo, M.L.; Cardenas Delgado, V.M.; Nugnes, L.G.; Gentilini, L.; Laderach, D.; Wolfenstein-Todel, C.; Compagno, D.; Rabinovich, G.A.; et al. Galectin-8: A matricellular lectin with key roles in angiogenesis. Glycobiology 2014, 24, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Delgado, V.M.; Nugnes, L.G.; Colombo, L.L.; Troncoso, M.F.; Fernandez, M.M.; Malchiodi, E.L.; Frahm, I.; Croci, D.O.; Compagno, D.; Rabinovich, G.A.; et al. Modulation of endothelial cell migration and angiogenesis: A novel function for the “tandem-repeat” lectin galectin-8. FASEB J. 2011, 25, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Levy, Y.; Arbel-Goren, R.; Hadari, Y.R.; Eshhar, S.; Ronen, D.; Elhanany, E.; Geiger, B.; Zick, Y. Galectin-8 functions as a matricellular modulator of cell adhesion. J. Biol. Chem. 2001, 276, 31285–31295. [Google Scholar] [CrossRef] [PubMed]
- Metz, C.; Doger, R.; Riquelme, E.; Cortes, P.; Holmes, C.; Shaughnessy, R.; Oyanadel, C.; Grabowski, C.; Gonzalez, A.; Soza, A. Galectin-8 promotes migration and proliferation and prevents apoptosis in u87 glioblastoma cells. Biol. Res. 2016, 49, 33. [Google Scholar] [CrossRef] [PubMed]
- Tureci, O.; Schmitt, H.; Fadle, N.; Pfreundschuh, M.; Sahin, U. Molecular definition of a novel human galectin which is immunogenic in patients with hodgkin’s disease. J. Biol. Chem. 1997, 272, 6416–6422. [Google Scholar] [CrossRef] [PubMed]
- Heusschen, R.; Griffioen, A.W.; Thijssen, V.L. Galectin-9 in tumor biology: A jack of multiple trades. Biochim. Biophys. Acta 2013, 1836, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Kageshita, T.; Kashio, Y.; Yamauchi, A.; Seki, M.; Abedin, M.J.; Nishi, N.; Shoji, H.; Nakamura, T.; Ono, T.; Hirashima, M. Possible role of galectin-9 in cell aggregation and apoptosis of human melanoma cell lines and its clinical significance. Int. J. Cancer 2002, 99, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, A.; Kontani, K.; Kihara, M.; Nishi, N.; Yokomise, H.; Hirashima, M. Galectin-9, a novel prognostic factor with antimetastatic potential in breast cancer. Breast J. 2006, 12, S196–S200. [Google Scholar] [CrossRef] [PubMed]
- Irie, A.; Yamauchi, A.; Kontani, K.; Kihara, M.; Liu, D.; Shirato, Y.; Seki, M.; Nishi, N.; Nakamura, T.; Yokomise, H.; et al. Galectin-9 as a prognostic factor with antimetastatic potential in breast cancer. Clin. Cancer Res. 2005, 11, 2962–2968. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, V.L.; Hulsmans, S.; Griffioen, A.W. The galectin profile of the endothelium: Altered expression and localization in activated and tumor endothelial cells. Am. J. Pathol. 2008, 172, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The tim-3 ligand galectin-9 negatively regulates t helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; He, W.; Zhou, H.; Yuan, J.; Wu, K.; Xu, L.; Chen, Z.K. The tim-3 ligand galectin-9 negatively regulates cd8+ alloreactive t cell and prolongs survival of skin graft. Cell Immunol. 2007, 250, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Oomizu, S.; Sakata, K.M.; Sakata, A.; Arikawa, T.; Watanabe, K.; Ito, K.; Takeshita, K.; Niki, T.; Saita, N.; et al. Galectin-9 suppresses the generation of th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis. Clin. Immunol. 2008, 127, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Facciabene, A.; Motz, G.T.; Coukos, G. T-regulatory cells: Key players in tumor immune escape and angiogenesis. Cancer Res. 2012, 72, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Anz, D.; Engel, J.; Kirchner, T.; Endres, S.; Mayr, D. Analysis of foxp3+ t-regulatory cells and cd8+ T-cells in ovarian carcinoma: Location and tumor infiltration patterns are key prognostic markers. PLoS ONE 2014, 9, e111757. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Gal-8 Expression (Cytoplasm) | p-Value | Gal-8 Expression (Nucleus) | p-Value | Gal-9 Expression (Cytoplasm) | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low | High | Negative | Positive | Negative | Moderate | High | ||||
| Histology | ||||||||||
| Serous | 40 | 62 | NS | 54 | 48 | NS | 24 | 71 | 9 | 0.024 |
| Clear cell | 2 | 9 | 3 | 8 | 1 | 10 | 0 | |||
| Endometrioid | 2 | 17 | 8 | 11 | 6 | 10 | 5 | |||
| Mucinous | 3 | 8 | 5 | 7 | 1 | 6 | 4 | |||
| Tumor Stage | ||||||||||
| pT1 | 10 | 26 | NS | 14 | 23 | NS | 8 | 20 | 9 | 0.018 |
| pT2+ | 37 | 69 | 55 | 51 | 24 | 77 | 8 | |||
| Lymph node | ||||||||||
| pN0/ pNX | 25 | 70 | 0.019 | 43 | 53 | NS | 26 | 61 | 12 | NS |
| pN1 | 22 | 26 | 27 | 21 | 6 | 36 | 6 | |||
| Distant Metastasis | ||||||||||
| pM0/pMX | 45 | 94 | NS | 68 | 72 | NS | 32 | 92 | 17 | NS |
| pM1 | 2 | 2 | 2 | 2 | 0 | 5 | 1 | |||
| Grading | ||||||||||
| G1 | 7 | 26 | NS | 13 | 21 | NS | 7 | 19 | 8 | 0.006 |
| G2+ | 37 | 62 | 53 | 46 | 24 | 72 | 5 | |||
| FIGO | ||||||||||
| I/ II | 8 | 33 | 0.033 | 14 | 28 | 0.011 | 6 | 25 | 11 | 0.002 |
| III/ IV | 37 | 60 | 55 | 42 | 25 | 69 | 6 | |||
| Age | ||||||||||
| ≤60 years | 24 | 51 | NS | 32 | 43 | NS | 10 | 52 | 17 | <0.001 |
| >60 years | 23 | 45 | 38 | 31 | 22 | 45 | 1 | |||
| Covariate | Coefficient (bi) | HR Exp (bi) | 95% CI | p-Value | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| Histology | −0.005 | 0.995 | 0.989 | 1.002 | 0.135 |
| Grading | 0.614 | 1.848 | 1.342 | 2.544 | <0.001 |
| FIGO | 0.763 | 2.144 | 1.503 | 3.058 | <0.001 |
| Patients’ age (≤60 vs. >60 years) | 0.737 | 2.089 | 1.265 | 3.447 | 0.004 |
| Gal-8 staining (low vs. high) | −0.487 | 0.615 | 0.388 | 0.973 | 0.038 |
| Gal-9 staining (neg. vs. low vs. high) | 0.687 | 1.988 | 1.257 | 3.145 | 0.003 |
| Gal-9 Cytoplasm | |
|---|---|
| Gal-8 cytoplasm | |
| cc | 0.464 |
| p | <0.001 |
| n | 142 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulz, H.; Kuhn, C.; Hofmann, S.; Mayr, D.; Mahner, S.; Jeschke, U.; Schmoeckel, E. Overall Survival of Ovarian Cancer Patients Is Determined by Expression of Galectins-8 and -9. Int. J. Mol. Sci. 2018, 19, 323. https://doi.org/10.3390/ijms19010323
Schulz H, Kuhn C, Hofmann S, Mayr D, Mahner S, Jeschke U, Schmoeckel E. Overall Survival of Ovarian Cancer Patients Is Determined by Expression of Galectins-8 and -9. International Journal of Molecular Sciences. 2018; 19(1):323. https://doi.org/10.3390/ijms19010323
Chicago/Turabian StyleSchulz, Heiko, Christina Kuhn, Simone Hofmann, Doris Mayr, Sven Mahner, Udo Jeschke, and Elisa Schmoeckel. 2018. "Overall Survival of Ovarian Cancer Patients Is Determined by Expression of Galectins-8 and -9" International Journal of Molecular Sciences 19, no. 1: 323. https://doi.org/10.3390/ijms19010323