Involvement of Bradykinin B2 Receptor in Pathological Vascularization in Oxygen-Induced Retinopathy in Mice and Rabbit Cornea

 , , ,
, , ,
Abstract
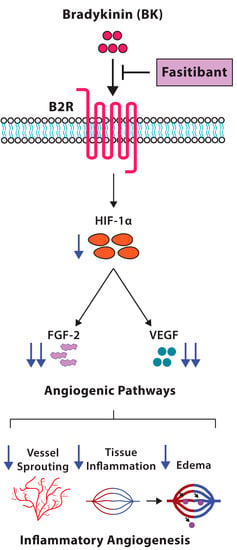
:
1. Introduction
2. Results
2.1. B2R Blockade Reduces the Extent of Retinal Vascularization and Maturation in Mouse Pups
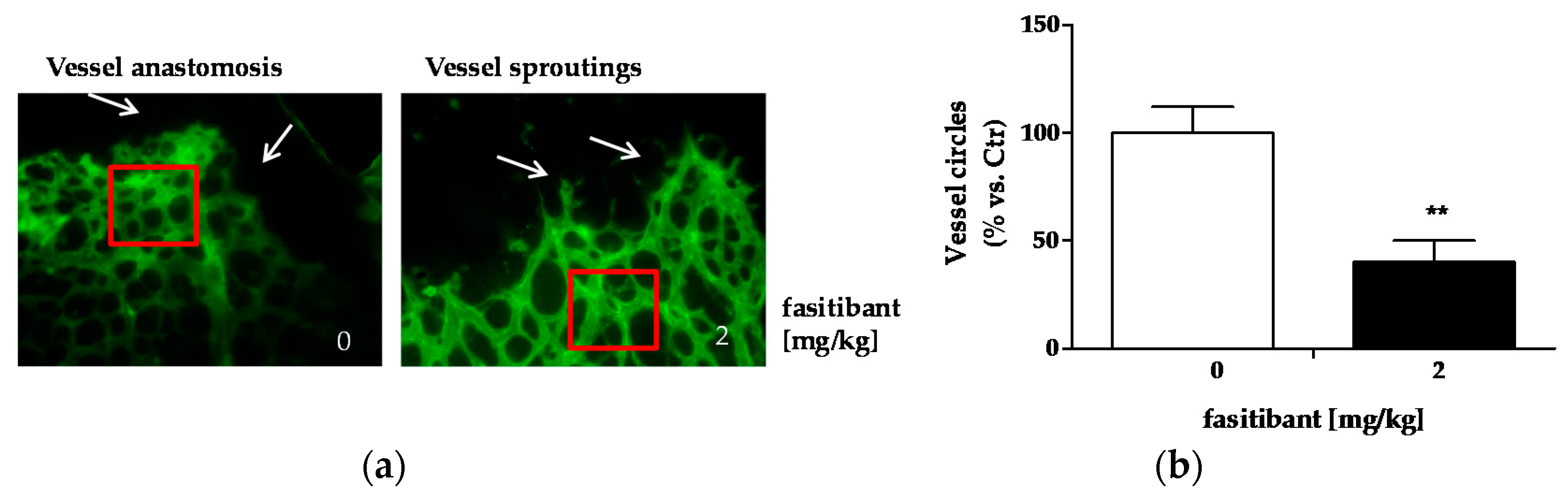
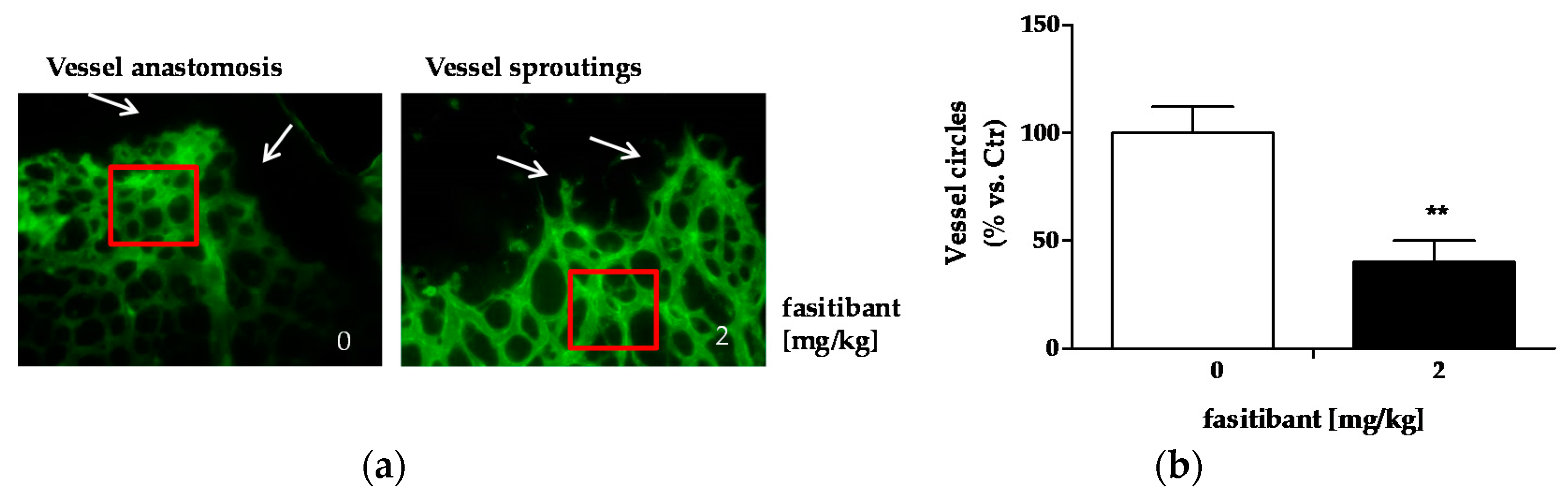
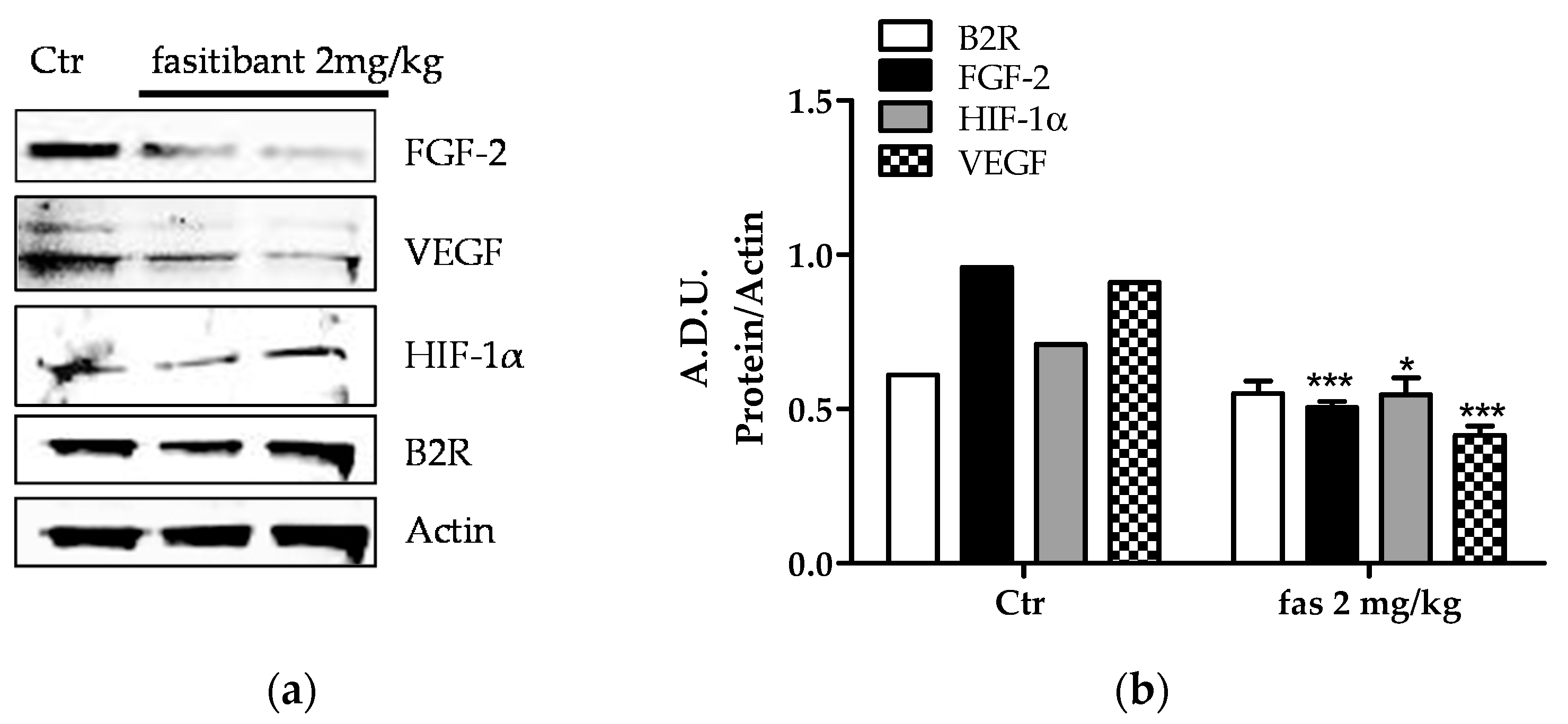
2.2. B2R Signaling Promotes Pathological Neoangiogenesis
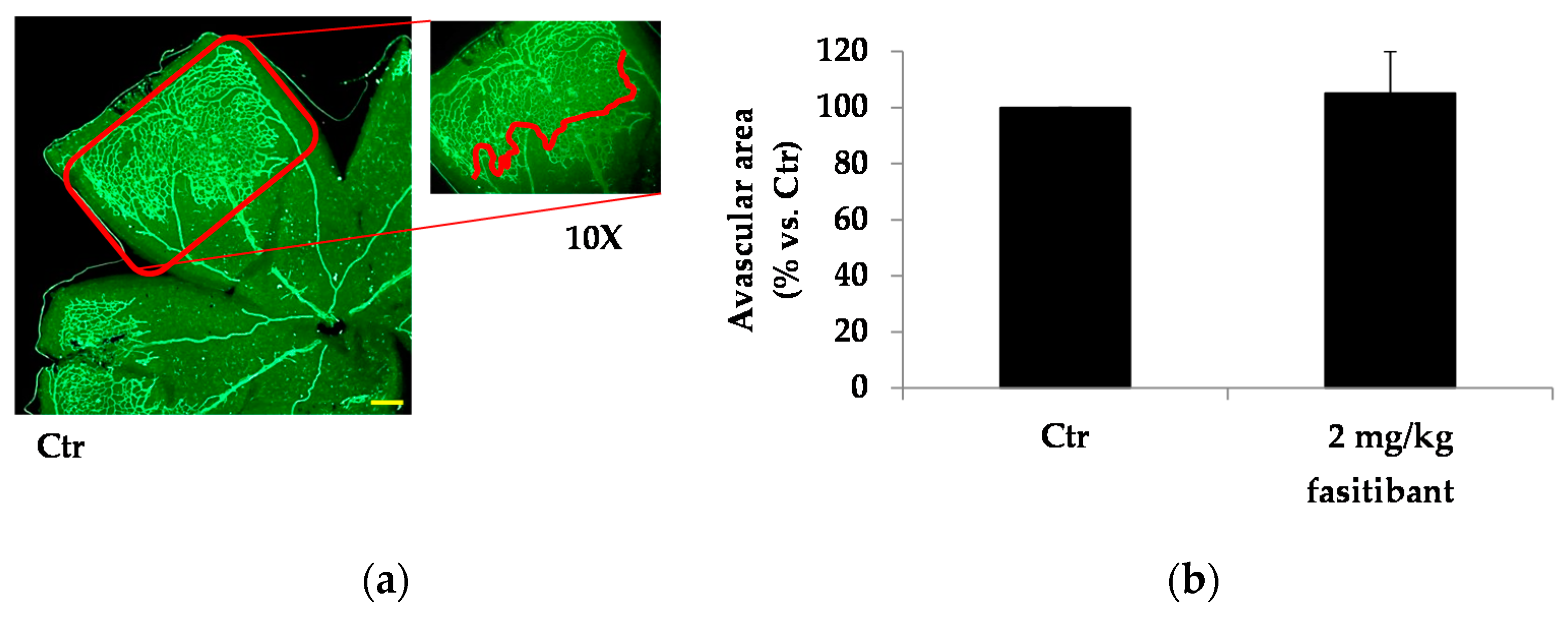
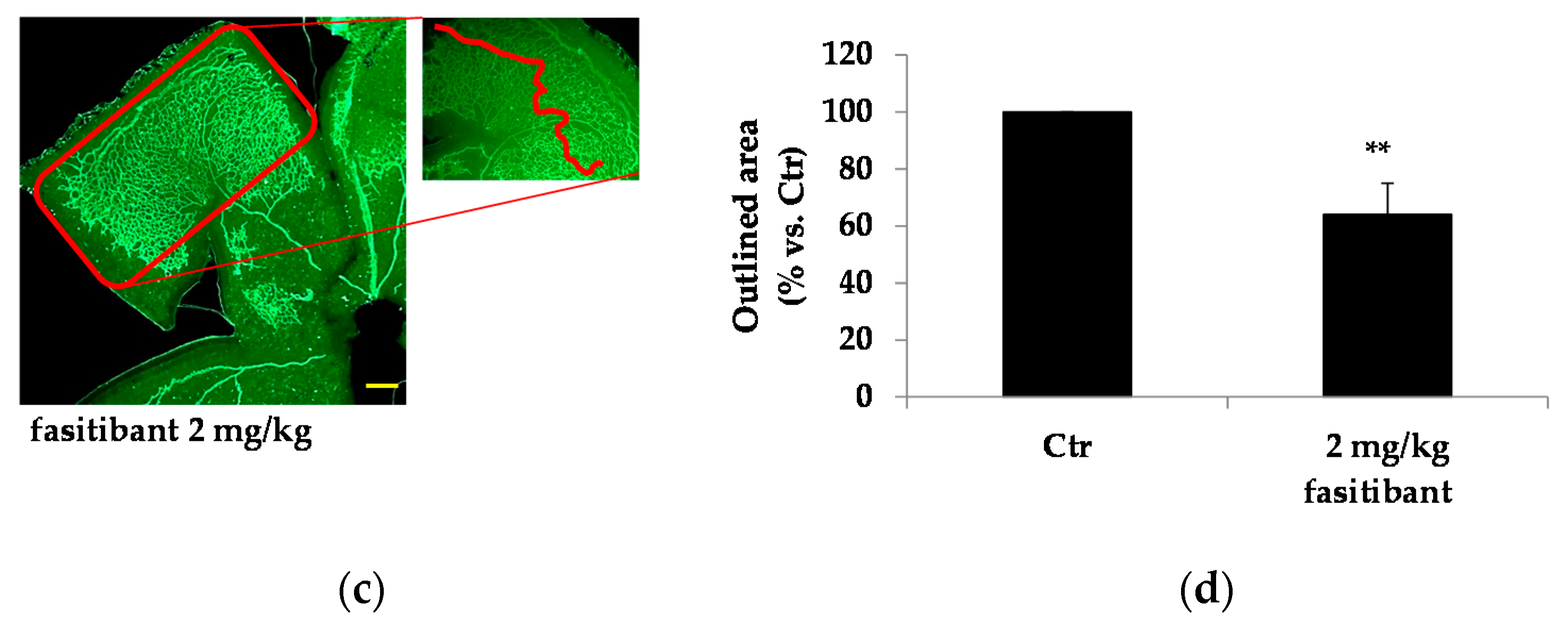
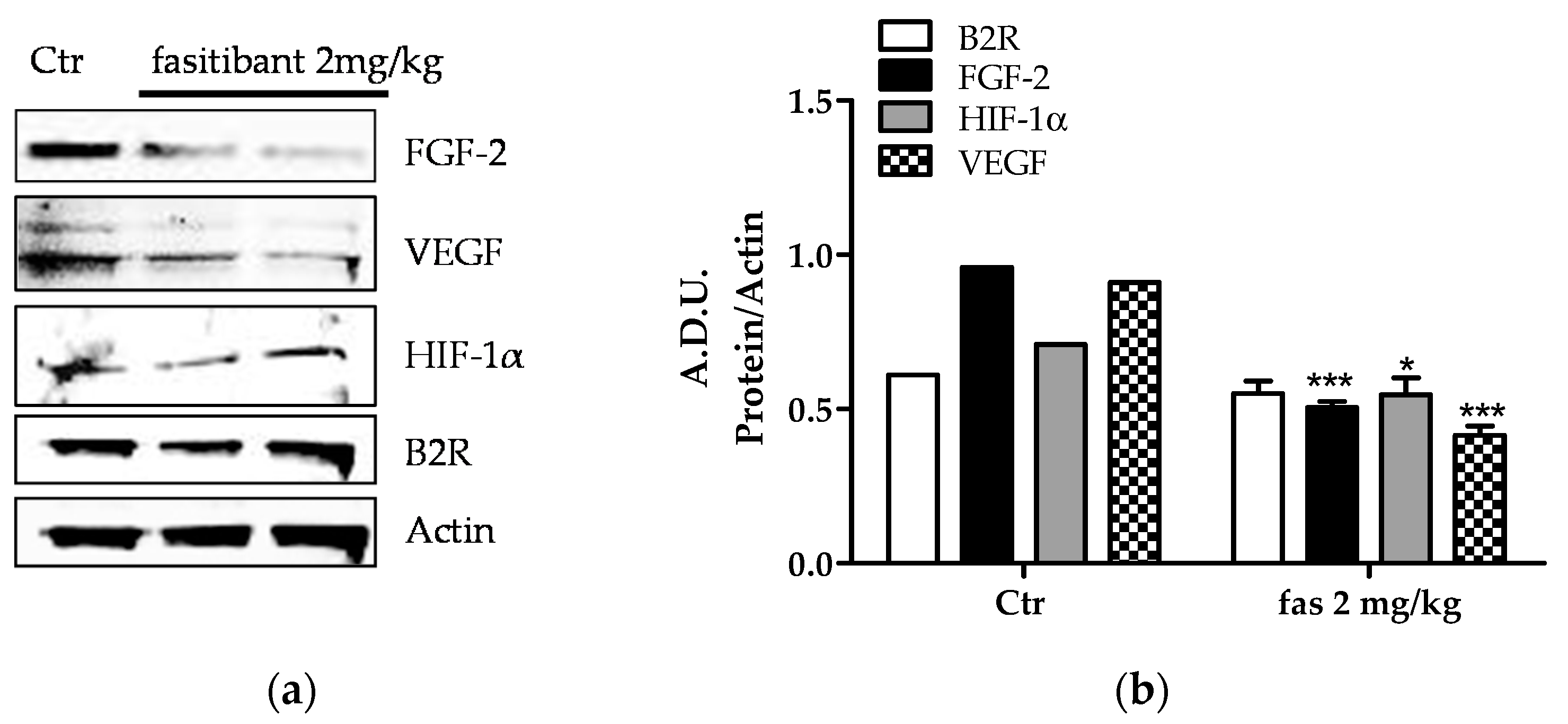
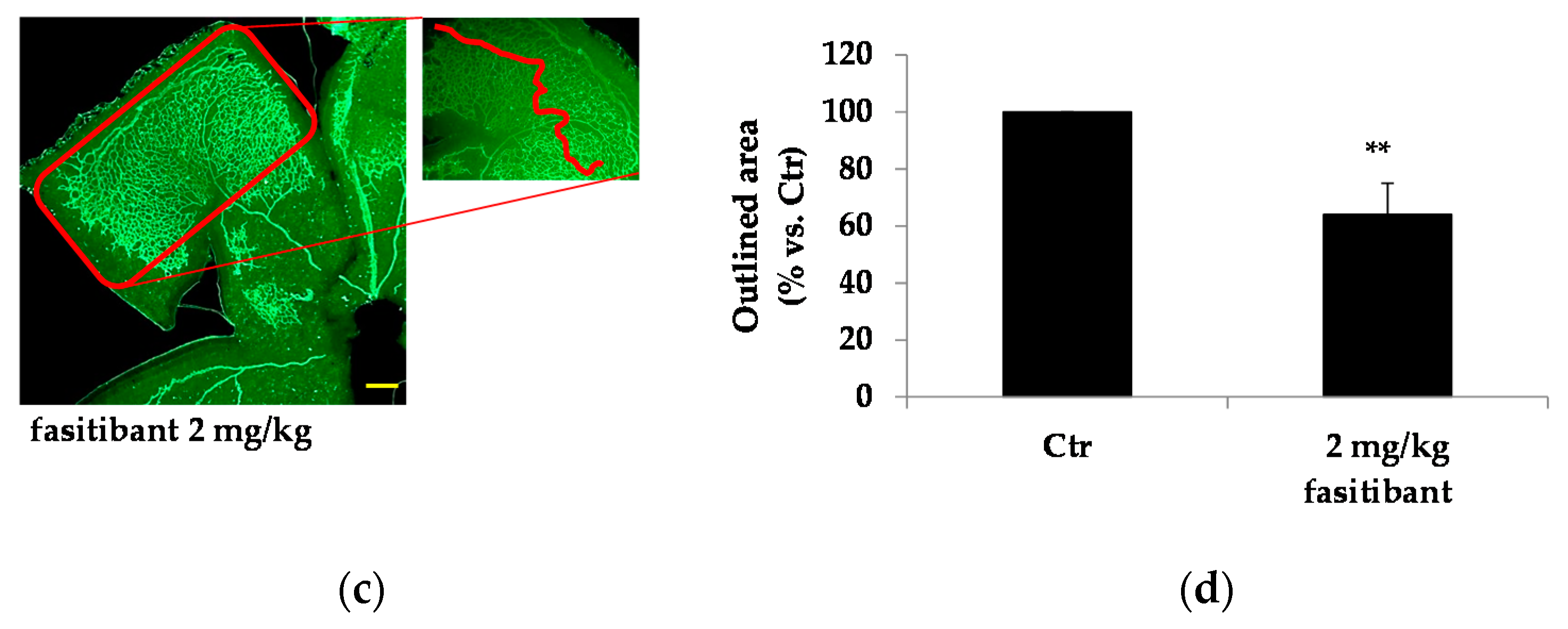
2.3. Fasitibant Reduces Retinal Neovascularization in the OIR Model
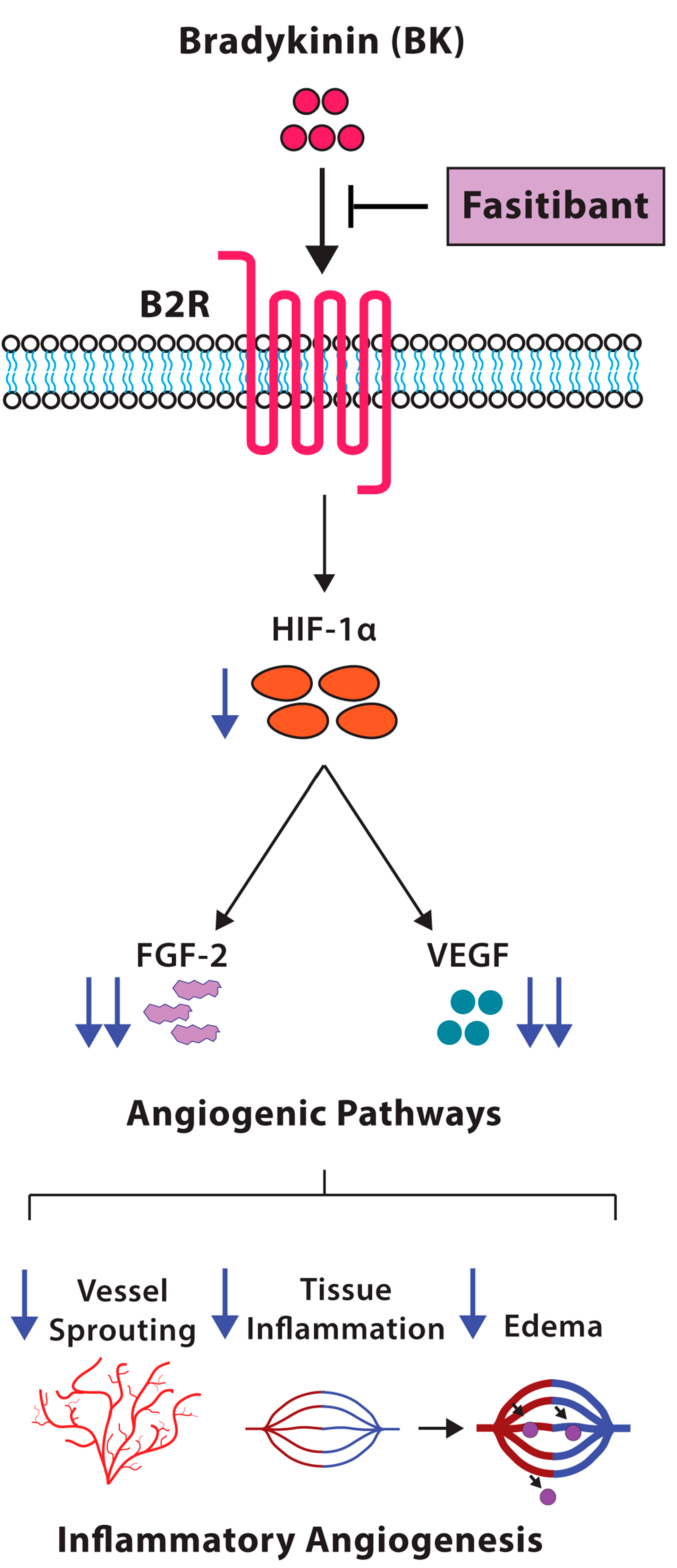
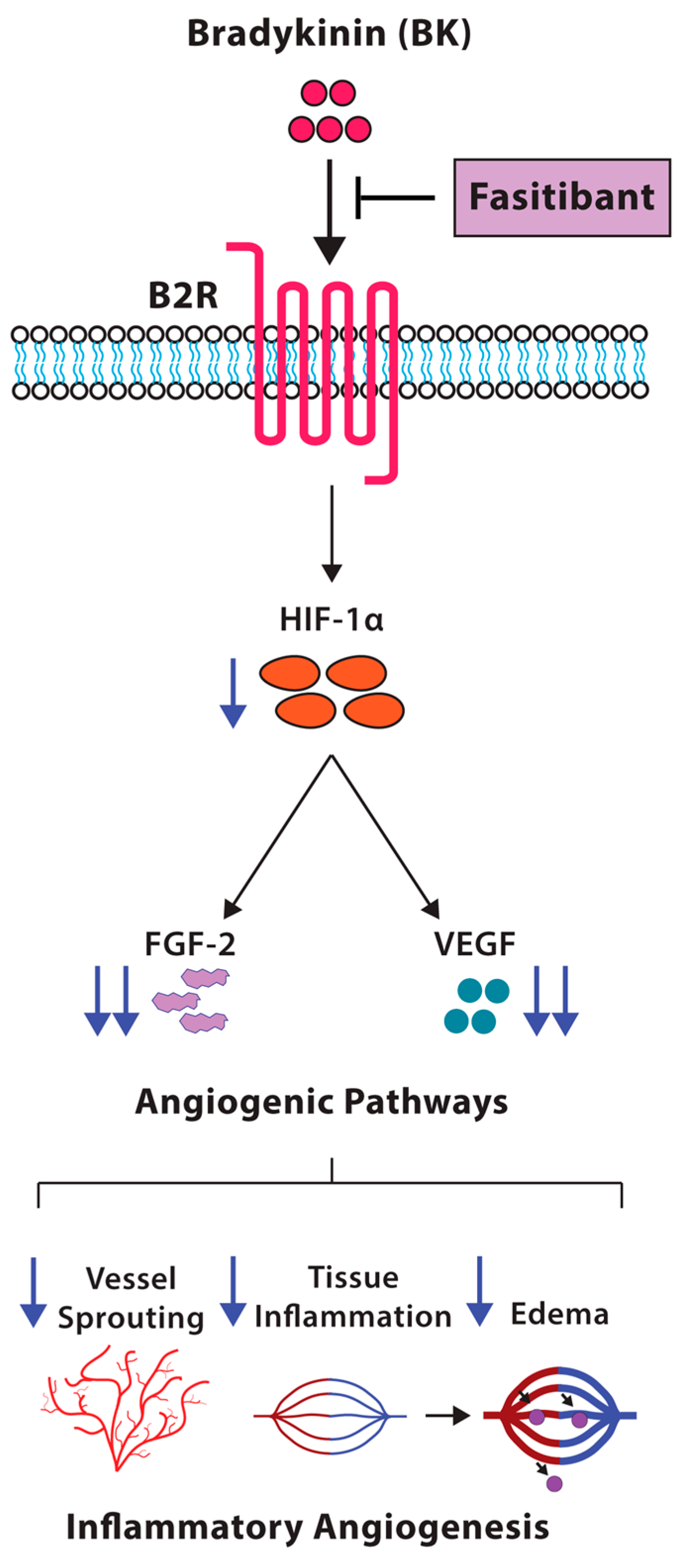
3. Discussion
4. Materials and Methods
4.1. Fasitibant Treatment of Mouse Pups
4.2. Immunofluorescence Analysis
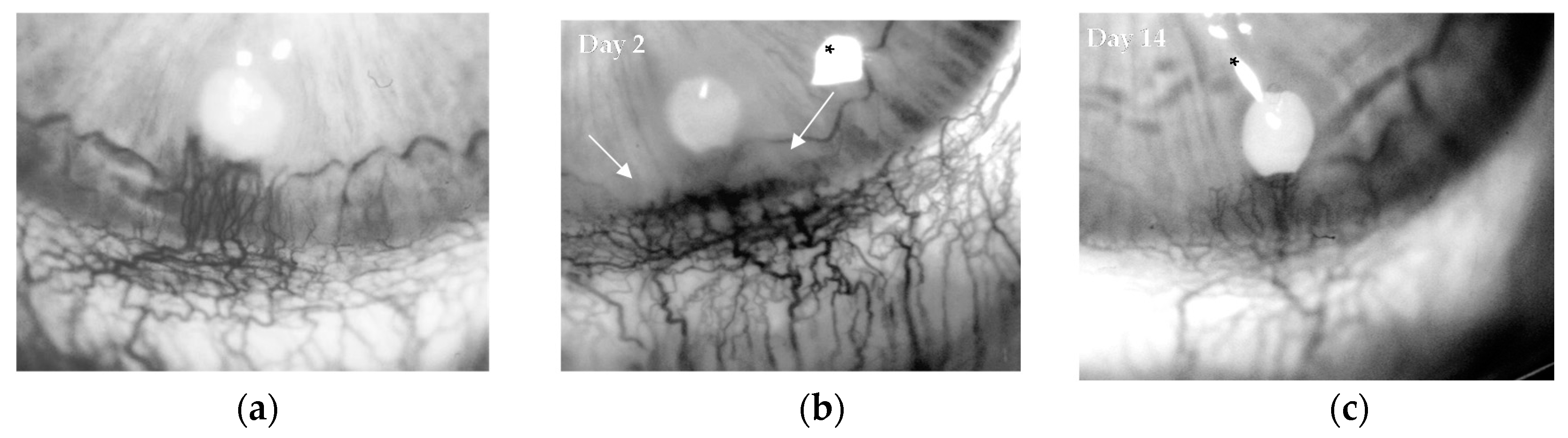
4.3. Angiogenesis In Vivo: Rabbit Cornea Assay
4.4. The Mouse Model of OIR
4.5. Western Blot Assay
4.6. Materials and Reagents
4.7. Data Analysis and Statistical Procedures
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| A.D.U. | Arbitrary density unit |
| BK | Bradykinin |
| B1R | BK receptor 1 |
| B2R | BK receptor 2 |
| Fas | Fasitibant |
| FGF-2 | Fibroblast growth factor-2 |
| HIF-1α | Hypoxia-inducible factor 1-alpha |
| HK | High–molecular-weight kininogen |
| i.p. | Intraperitoneal |
| KKS | Kallikrein–kinin system |
| NV | Neovessel |
| OIR | Oxygen-induced retinopathy |
| PB | Phosphate buffer |
| PBS | Phosphate buffer saline |
| VEGF | Vascular endothelial growth factor |
References
- Pruneau, D.; Bélichard, P.; Sahel, J.A.; Combal, J.P. Targeting the kallikrein-kinin system as a new therapeutic approach to diabetic retinopathy. Curr. Opin. Investig. Drugs 2010, 11, 507–514. [Google Scholar] [PubMed]
- Gao, B.B.; Clermont, A.; Rook, S.; Fonda, S.J.; Srinivasan, V.J.; Wojtkowski, M.; Fujimoto, J.G.; Avery, R.L.; Arrigg, P.G.; Bursell, S.E.; et al. Extracellular carbonic anhydrase mediates hemorrhagic retinal and cerebral vascular permeability through prekallikrein activation. Nat. Med. 2007, 13, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Feener, E.P. Plasma kallikrein kinin system and diabetic retinopathy. Biol. Chem. 2013, 394, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Parenti, A.; Morbidelli, L.; Ledda, F.; Granger, H.J.; Ziche, M. The bradykinin/B1 receptor promotes angiogenesis by up-regulation of endogenous FGF-2 in endothelium via the nitric oxide synthase pathway. FASEB J. 2001, 15, 1487–1489. [Google Scholar] [CrossRef] [PubMed]
- Terzuoli, E.; Meini, S.; Cucchi, P.; Catalani, C.; Cialdai, C.; Maggi, C.A.; Giachetti, A.; Ziche, M.; Donnini, S. Antagonism of bradykinin B2 receptor prevents inflammatory responses in human endothelial cells by quenching the NF-kB pathway activation. PLoS ONE 2014, 9, e84358. [Google Scholar] [CrossRef] [PubMed]
- Bossi, F.; Peerschke, E.I.; Ghebrehiwet, B.; Tedesco, F. Cross-talk between the complement and the kinin system in vascular permeability. Immunol. Lett. 2011, 140, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hofman, Z.L.; Relan, A.; Zeerleder, S.; Drouet, C.; Zuraw, B.; Hack, C.E. Angioedema attacks in patients with hereditary angioedema: Local manifestations of a systemic activation process. J. Allergy Clin. Immunol. 2016, 138, 359–366. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, P.A. Mechanism of retinal and choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3974–3979. [Google Scholar]
- Thuringer, D.; Maulon, L.; Frelin, C. Rapid transactivation of the vascular endothelial growth factor receptor KDR/Flk-1 by the bradykinin B2 receptor contributes to endothelial nitric-oxide synthase activation in cardiac capillary endothelial cells. J. Biol. Chem. 2002, 277, 2028–2032. [Google Scholar] [CrossRef] [PubMed]
- Vinores, S.A.; Youssri, A.I.; Luna, J.D.; Chen, Y.S.; Bhargave, S.; Vinores, M.A.; Schoenfeld, C.L.; Peng, B.; Chan, C.C.; LaRochelle, W.; et al. Upregulation of vascular endothelial growth factor in ischemic and non-ischemic human and experimental retinal disease. Histol. Histopathol. 1997, 12, 99–109. [Google Scholar] [PubMed]
- Monti, M.; Donnini, S.; Morbidelli, L.; Giachetti, A.; Mochly-Rosen, D.; Mignatti, P.; Ziche, M. PKCε activation promotes FGF-2 exocytosis and induces endothelial cell proliferation and sprouting. J. Mol. Cell. Cardiol. 2013, 63, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Stavri, G.T.; Zachary, I.C.; Baskerville, P.A.; Martin, J.F.; Erusalimsky, J.D. Basic fibroblast growth factor upregulates the expression of vascular endothelial growth factor in vascular smooth muscle cells. Synergistic interaction with hypoxia. Circulation 1995, 92, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Li, Y.; Hendrickson, A.; Sage, E.H. Regulation of retinal capillary cells by basic fibroblast growth factor, vascular endothelial growth factor, and hypoxia. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 45–49. [Google Scholar] [CrossRef]
- Calvani, M.; Rapisarda, A.; Uranchimeg, B.; Shoemaker, R.H.; Melillo, G. Hypoxic induction of an HIF-1alpha-dependent bFGF autocrine loop drives angiogenesis in human endothelial cells. Blood 2006, 107, 2705–2712. [Google Scholar] [CrossRef] [PubMed]
- Castellon, R.; Hamdi, H.K.; Sacerio, I.; Aoki, A.M.; Kenney, M.C.; Ljubimov, A.V. Effects of angiogenic growth factor combinations on retinal endothelial cells. Exp. Eye Res. 2002, 74, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Holborn, M.; Jahn, K.; Limb, G.A.; Kohen, L.; Wiedemann, P.; Bringmann, A. Characterization of the basic fibroblast growth factor-evoked proliferation of the human Müller cell line, MIO-M1. Graefes Arch. Clin. Exp. Ophthalmol. 2004, 242, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.; Terasaki, H. Biological involvement in microRNA in proliferative vitreoretinopathy. Transl. Vis. Sci. Technol. 2017, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Zittermann, S.I.; Issekutz, A.C. Basic Fibroblast Growth Factor (bFGF, FGF-2) potentiates leukocyte recruitment to inflammation by enhancing endothelia adhesion molecule expression. Am. J. Path 2006, 168, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Meini, S.; Maggi, C.A. Knee osteoarthritis: A role for bradykinin? Inflamm. Res. 2008, 57, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Ellman, B.M.; Kroin, J.S.; Chen, D.; Yan, D.; Mikecz, K.; Ranjan, K.C.; Xiao, G.; Stein, G.S.; Kim, S.; et al. Species-specific biological effects of FGF-2 in articular cartilage: Implication for distinct roles within the FGF receptor family. J. Cell. Biochem. 2012, 113, 2532–2542. [Google Scholar] [CrossRef]
- Zubakova, R.; Gille, A.; Faussner, A.; Hilgenfeldt, U. Ca2+ signalling of kinins in cells expressing rat, mouse and human B1/B2-receptor. Int. Immunopharmacol. 2008, 2, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Ambati, B.K.; Nozaki, M.; Singh, N.; Takeda, A.; Jani, P.D.; Suthar, T.; Albuquerque, R.J.; Richter, E.; Sakurai, E.; Newcomb, M.T.; et al. Corneal avascularity is due to soluble VEGF receptor-1. Nature 2006, 443, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Bader, M. Cardiovascular Hormone Systems: From Molecular Mechanisms to Novel Therapeutics; Wiley-Blackwell: Hoboken, NJ, USA, 2008; p. 105. ISBN 978-3-527-31920-6. [Google Scholar]
- Renné, T.; Schuh, K.; Müller-Esterl, W. Local bradykinin formation is controlled by glycosaminoglycans. J. Immunol. 2005, 175, 3377–3385. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, P.L.N.; Sirois, P.; Tannock, I.F.; Chammas, R. The role of kinin receptors in cancer and therapeutic opportunities. Cancer Lett. 2014, 345, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Morbidelli, L.; Parenti, A.; Giovannelli, L.; Granger, H.J.; Ledda, F.; Ziche, M. B1 receptor involvement in the effect of bradykinin on venular endothelial cell proliferation and potentiation of FGF-2 effects. Br. J. Pharmacol. 1998, 124, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Donnini, S.; Giachetti, A.; Morbidelli, L.; Ziche, M. Prostaglandin E(2) primes the angiogenic switch via a synergic interaction with the fibroblast growth factor-2 pathway. Circ. Res. 2009, 105, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Pitulescu, M.E.; Schmidt, I.; Benedito, R.; Adams, R.H. Inducible gene targeting in the neonatal vasculature and analysis of retinal angiogenesis in mice. Nat. Protoc. 2010, 5, 1518–1534. [Google Scholar] [CrossRef] [PubMed]
- Connor, K.M.; Krah, N.M.; Dennison, R.J.; Aderman, C.M.; Chen, J.; Guerin, K.I.; Sapieha, P.; Stahl, A.; Willett, K.L.; Smith, L.E. Quantification of oxygen-induced retinopathy in the mouse: A model of vessel loss, vessel regrowth and pathological angiogenesis. Nat. Protoc. 2009, 4, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Ziche, M.; Morbidelli, L.; Masini, E.; Amerini, S.; Granger, H.J.; Maggi, C.A.; Geppetti, P.; Ledda, L. Nitric oxide mediates angiogenesis in vivo and endothelial cell growth and migration in vitro promoted by substance P. J. Clin. Investig. 1994, 94, 2036–2044. [Google Scholar] [CrossRef] [PubMed]
- Ziche, M.; Morbidelli, L.; Choudhuri, R.; Zhang, H.-T.; Donnini, S.; Granger, H.J.; Bicknell, R. Nitric oxide-synthase lies downstream of vascular endothelial growth factor but not basic fibroblast growth factor induced angiogenesis. J. Clin. Investig. 1997, 99, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A.; Connor, K.M.; Sapieha, P.; Chen, J.; Dennison, R.J.; Krah, N.M.; Seaward, M.R.; Willett, K.L.; Aderman, C.M.; Guerin, K.I.; et al. The mouse retina as an angiogenesis model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2813–2826. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stimulus (Dose/Pellet) | Corneal Opacity (Day 2) | Sprouting (Day 8) | Sprouting (Day 14) |
|---|---|---|---|
| BK (1 μg) | ++ | 3.6 ± 0.4 | 5.0 ± 0.1 |
| Kallidin (1 μg) | ++ | 1.8 ± 0.2 | 1.3 ± 0.9 |
| Kallidin (5 μg) | +++ | 3.7 ± 0.9 | 4.6 ± 0.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terzuoli, E.; Morbidelli, L.; Nannelli, G.; Giachetti, A.; Donnini, S.; Ziche, M. Involvement of Bradykinin B2 Receptor in Pathological Vascularization in Oxygen-Induced Retinopathy in Mice and Rabbit Cornea. Int. J. Mol. Sci. 2018, 19, 330. https://doi.org/10.3390/ijms19020330
Terzuoli E, Morbidelli L, Nannelli G, Giachetti A, Donnini S, Ziche M. Involvement of Bradykinin B2 Receptor in Pathological Vascularization in Oxygen-Induced Retinopathy in Mice and Rabbit Cornea. International Journal of Molecular Sciences. 2018; 19(2):330. https://doi.org/10.3390/ijms19020330
Chicago/Turabian StyleTerzuoli, Erika, Lucia Morbidelli, Ginevra Nannelli, Antonio Giachetti, Sandra Donnini, and Marina Ziche. 2018. "Involvement of Bradykinin B2 Receptor in Pathological Vascularization in Oxygen-Induced Retinopathy in Mice and Rabbit Cornea" International Journal of Molecular Sciences 19, no. 2: 330. https://doi.org/10.3390/ijms19020330