Elucidating the Role of CD84 and AHR in Modulation of LPS-Induced Cytokines Production by Cruciferous Vegetable-Derived Compounds Indole-3-Carbinol and 3,3′-Diindolylmethane

Abstract
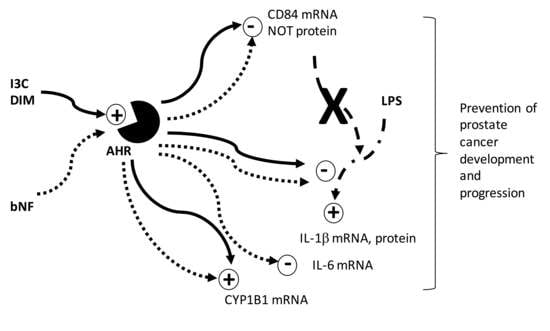
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
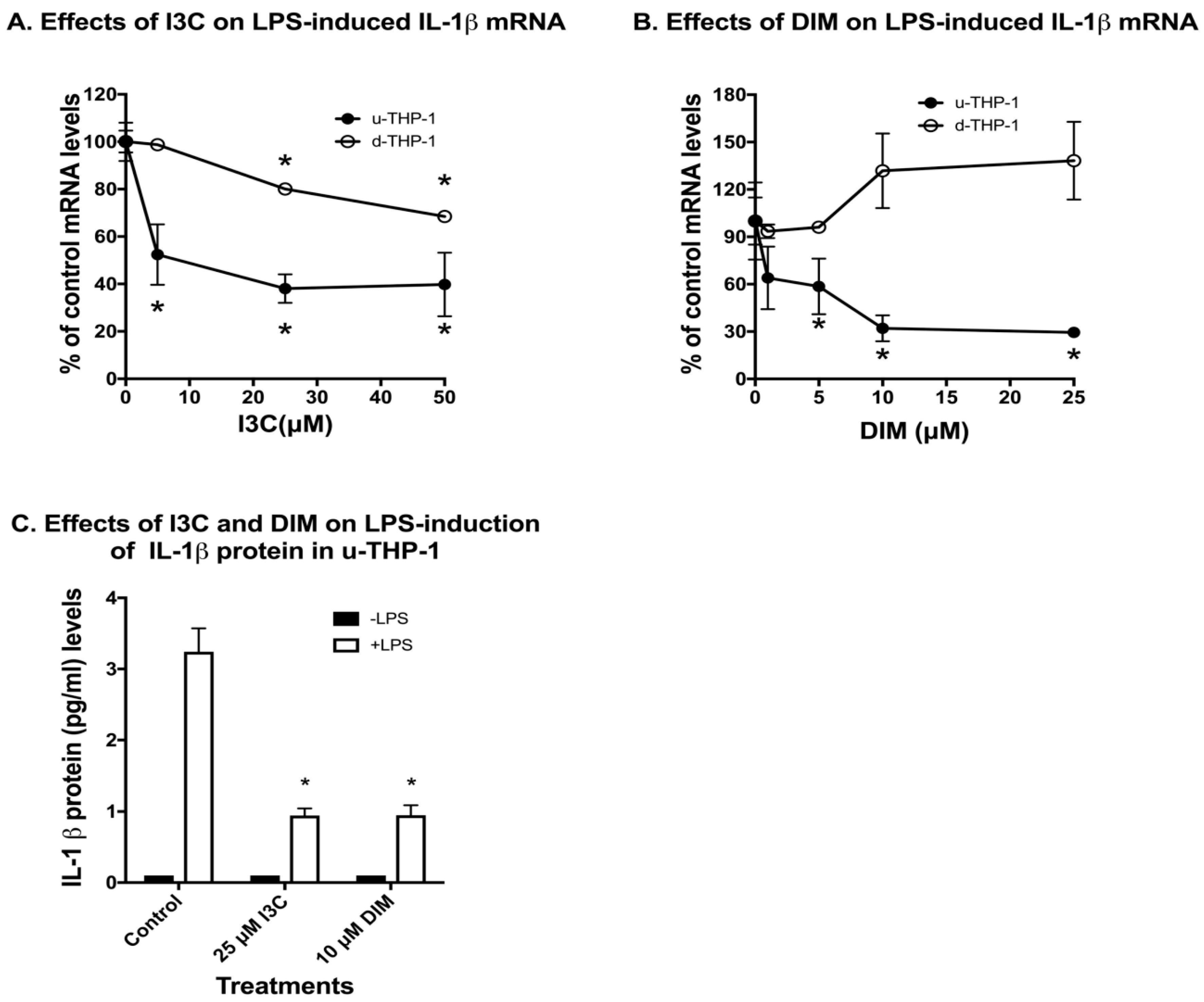
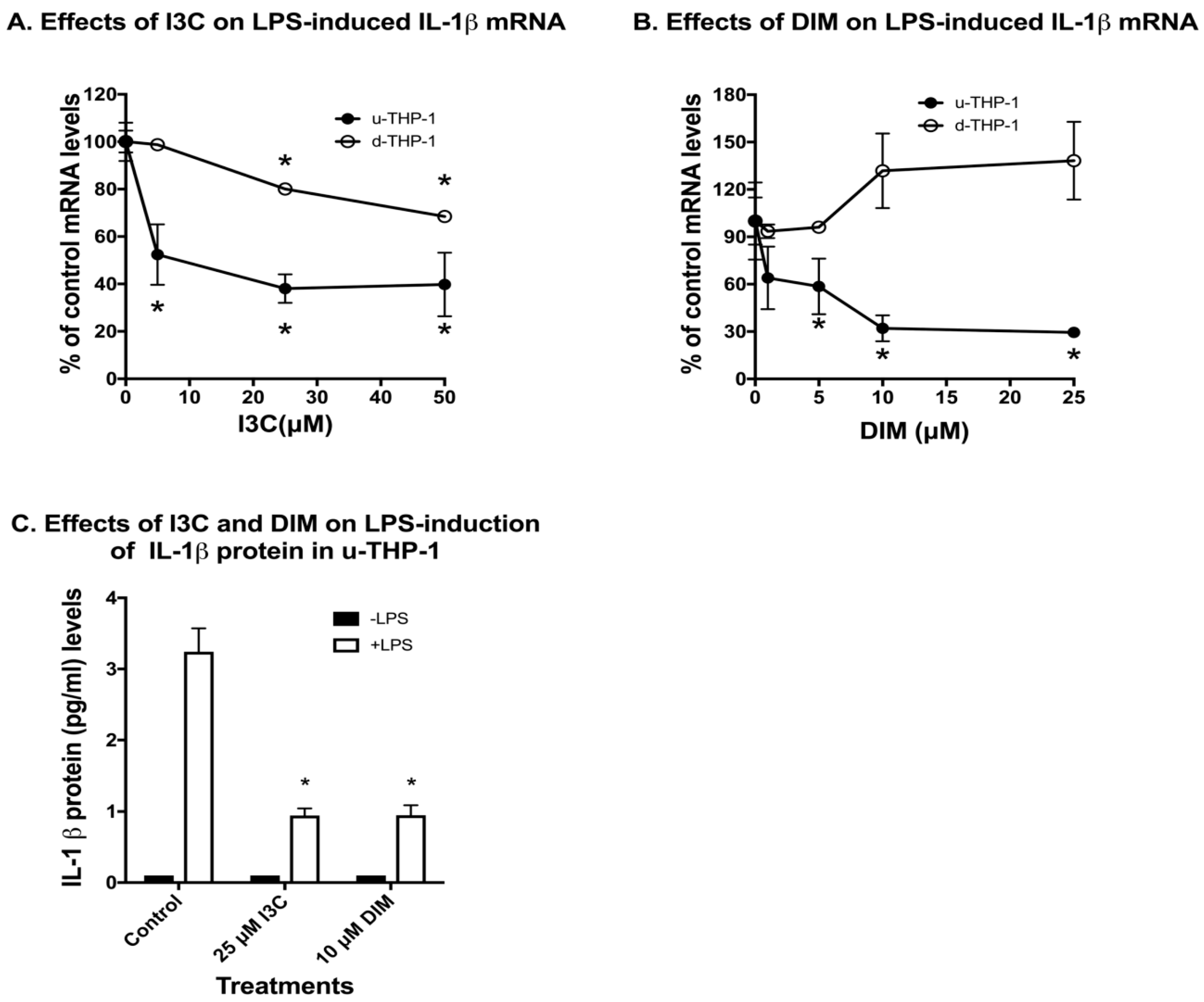
2.1. Effects of Indole-3-Carbinol (I3C) and 3,3′-Diindolylmethane (DIM) on Lipopolysaccharide (LPS) Induction of IL-1β in THP-1 Monocytic Cells
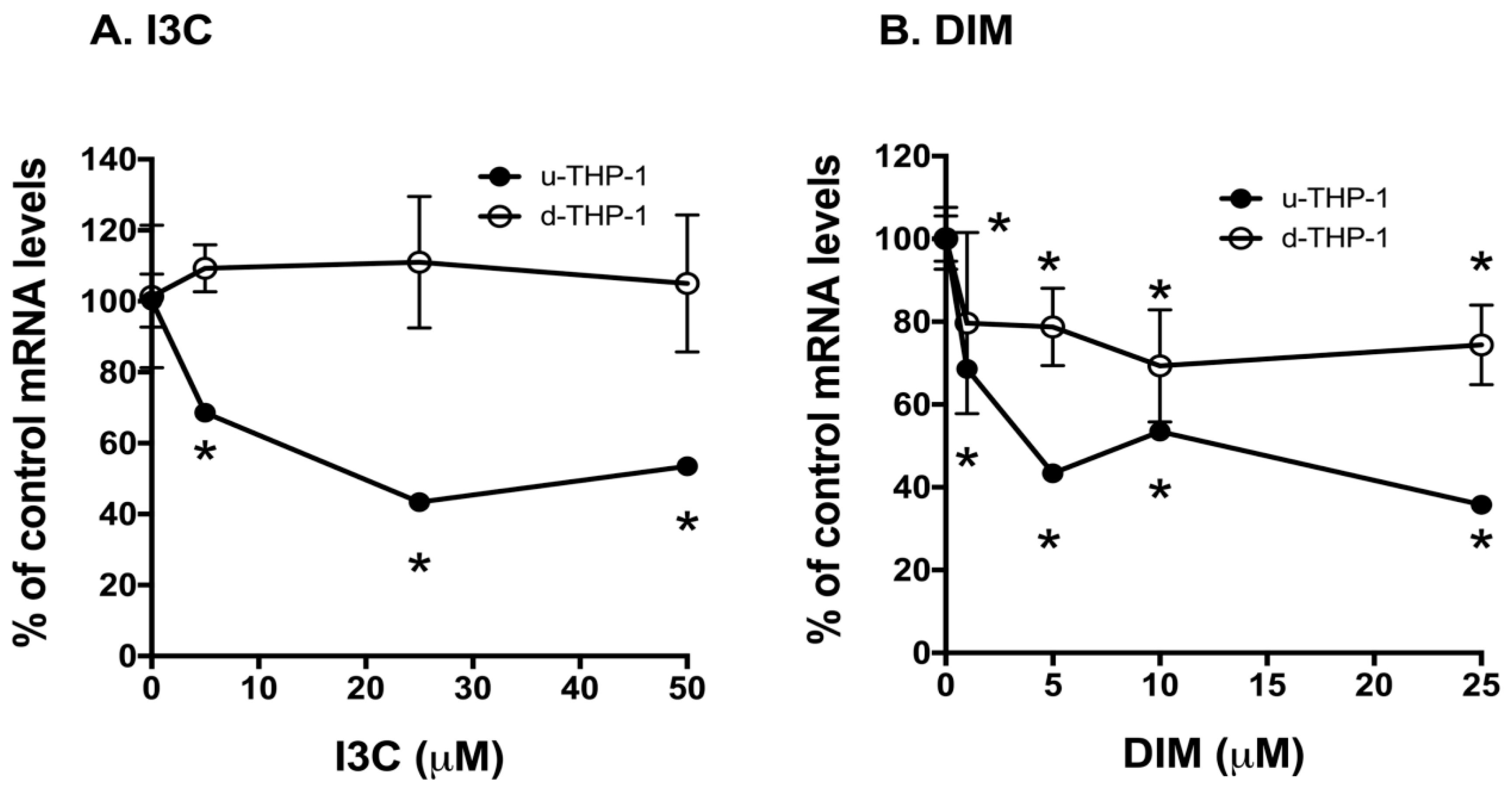
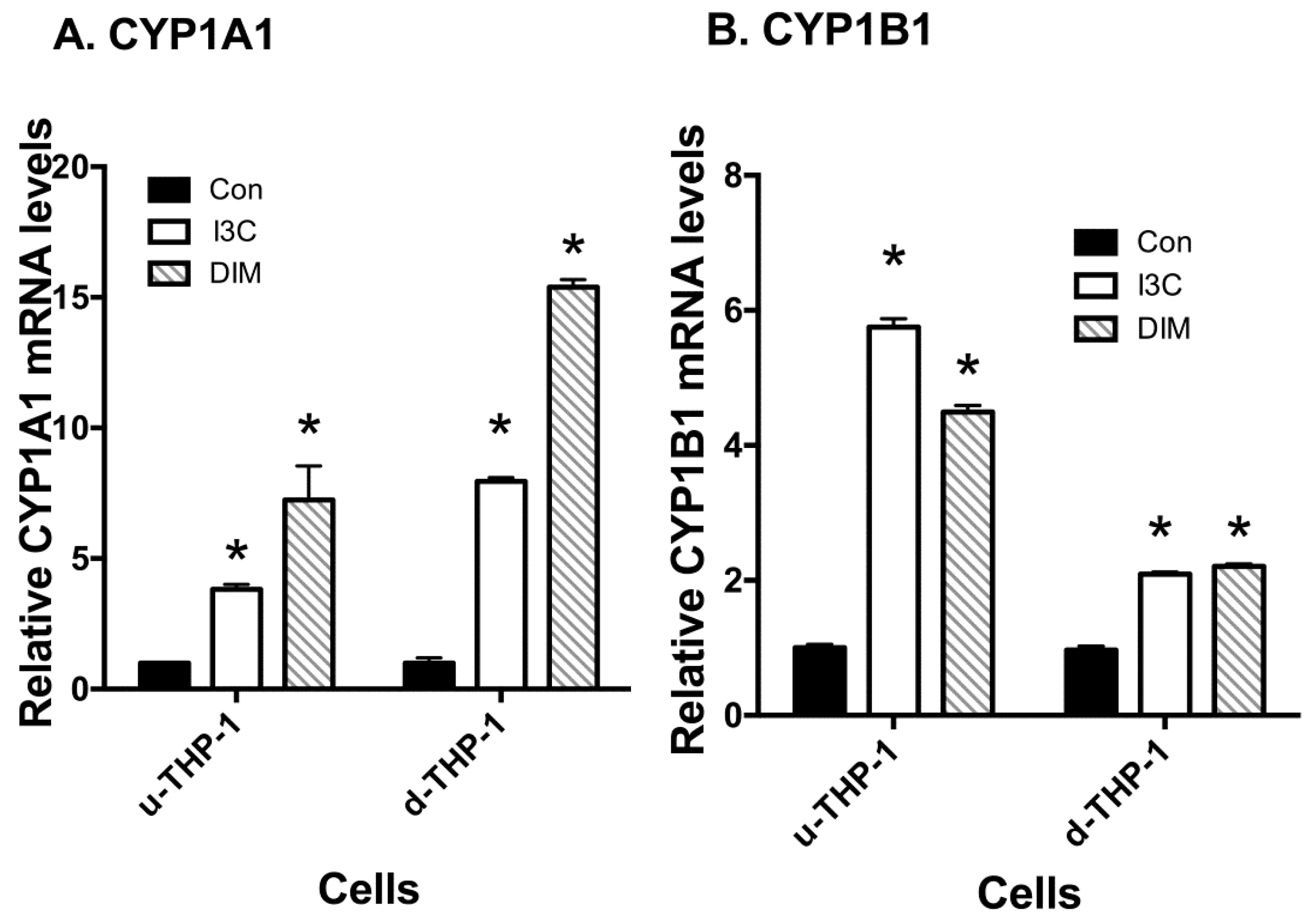
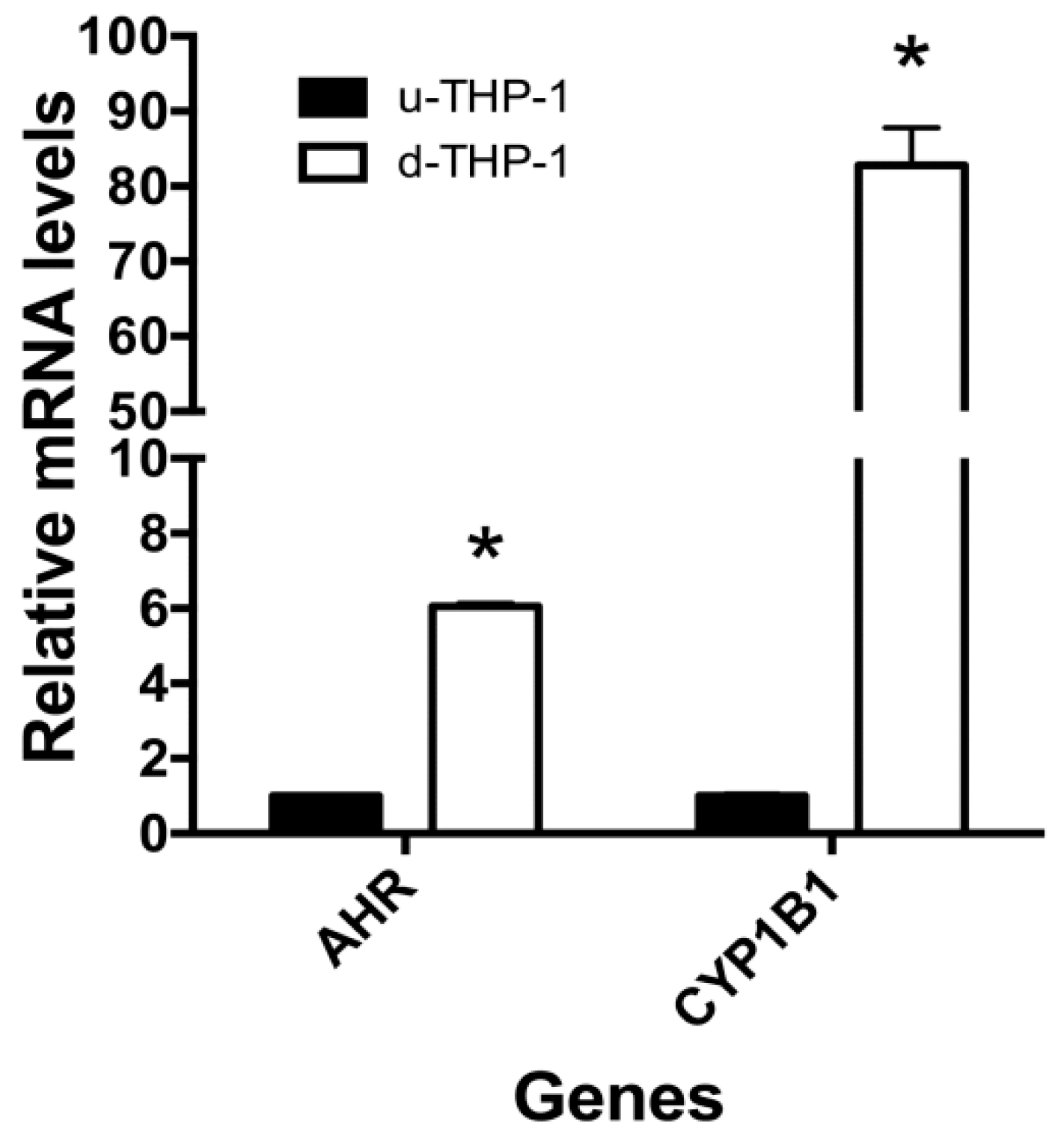
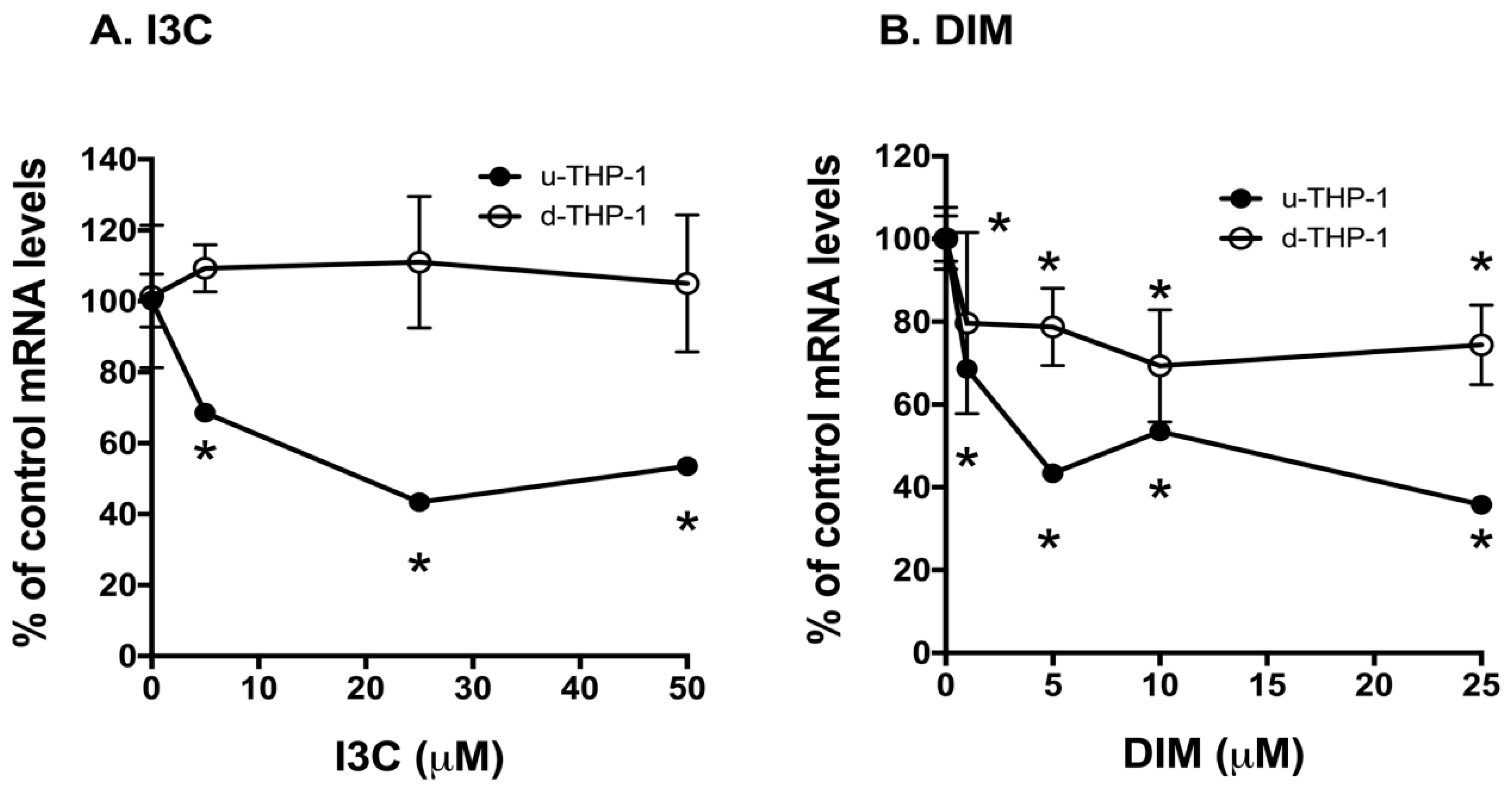
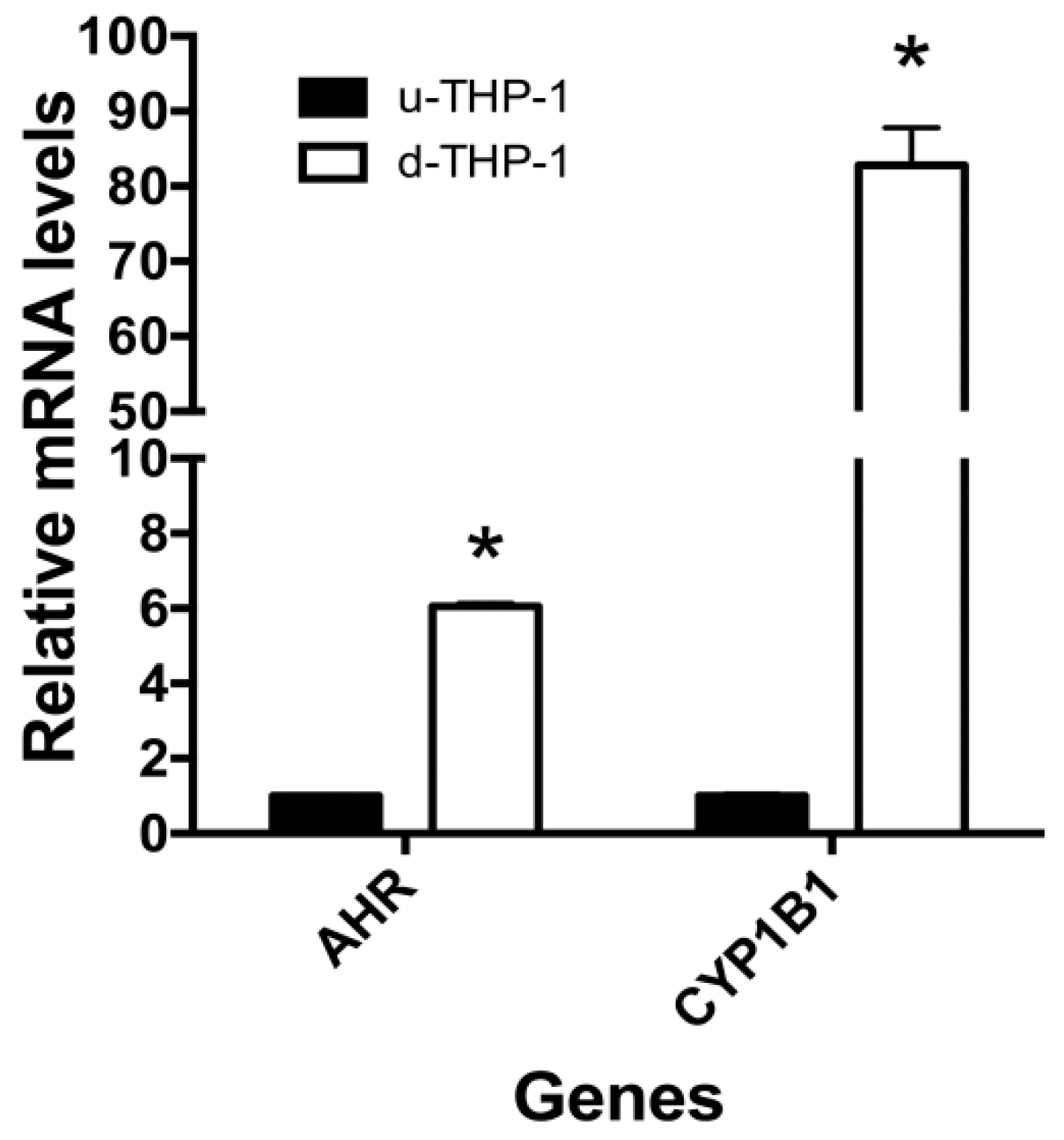
2.2. Effects of I3C and DIM on CD84 and Aryl Hydrocarbon (AHR)-Responsive Gene CYP1A1/B1 mRNA Levels in THP-1 Cells
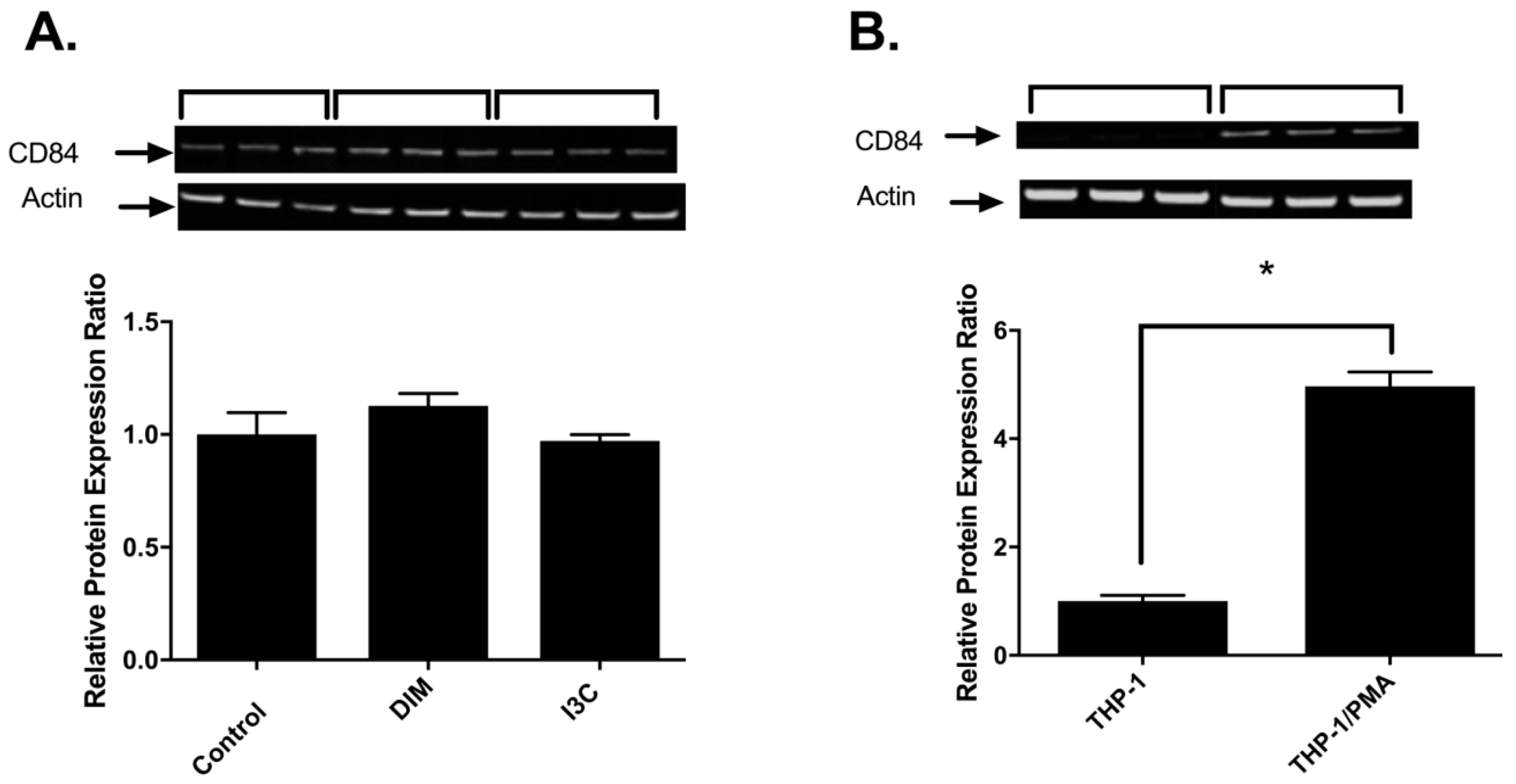
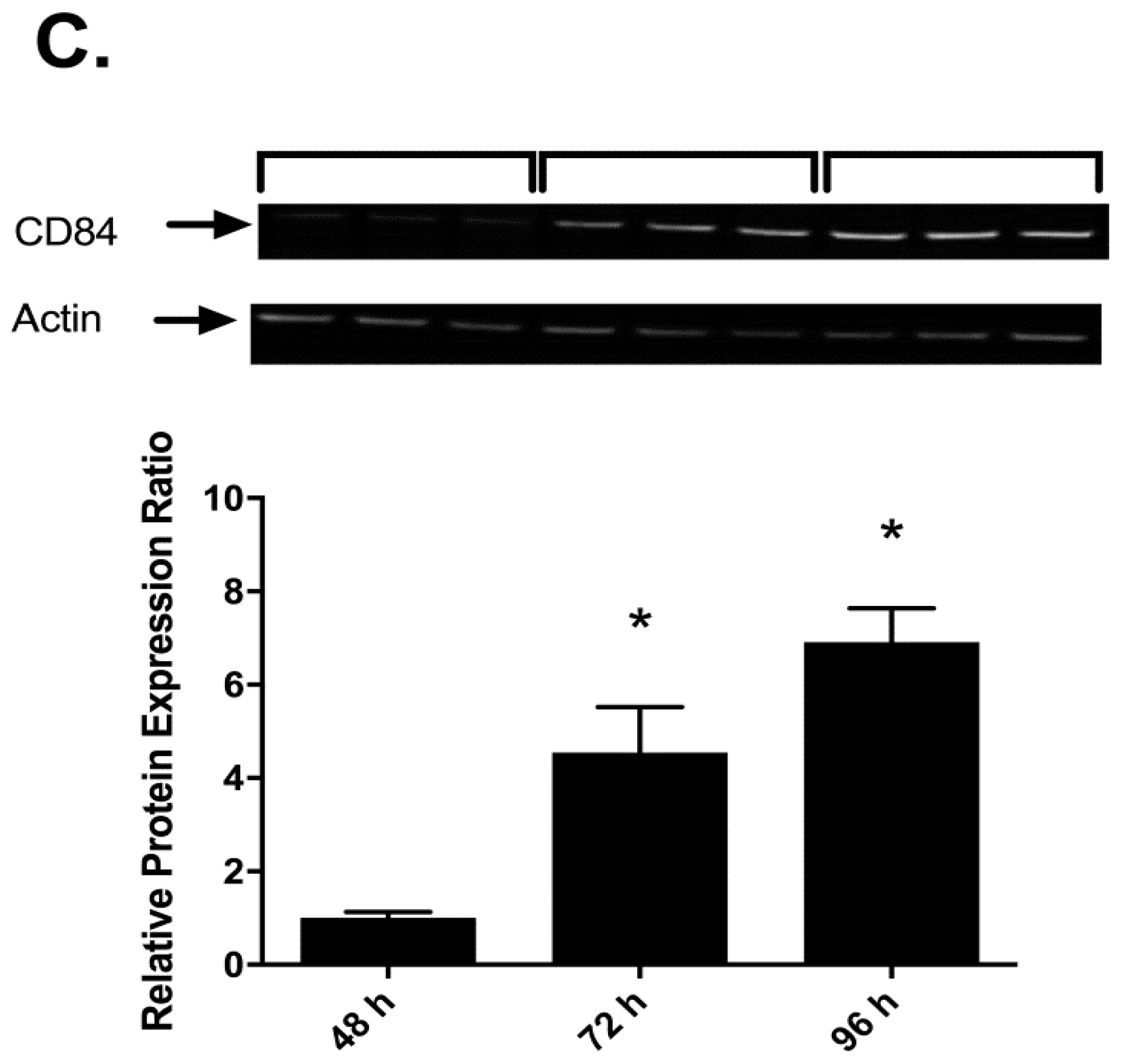
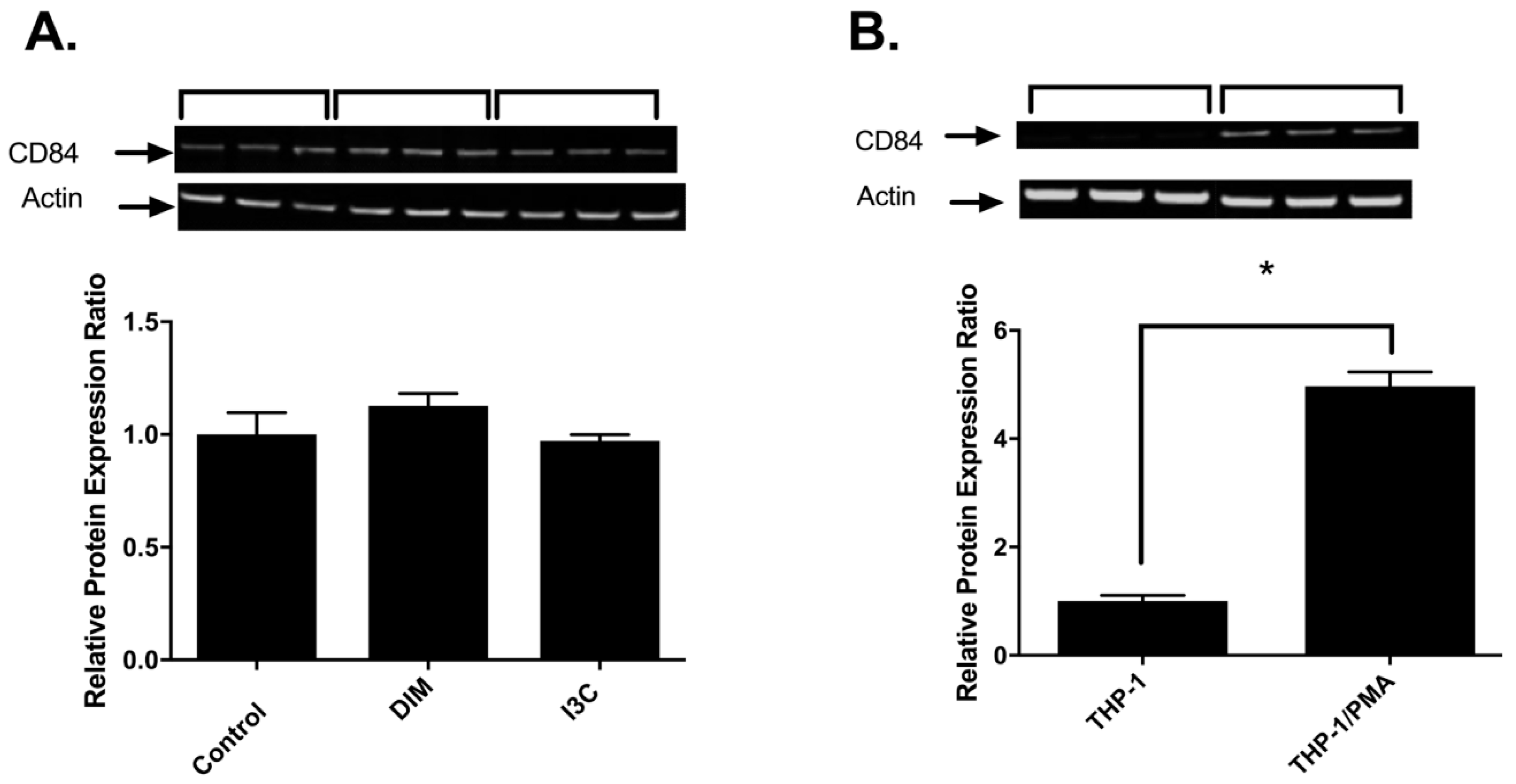
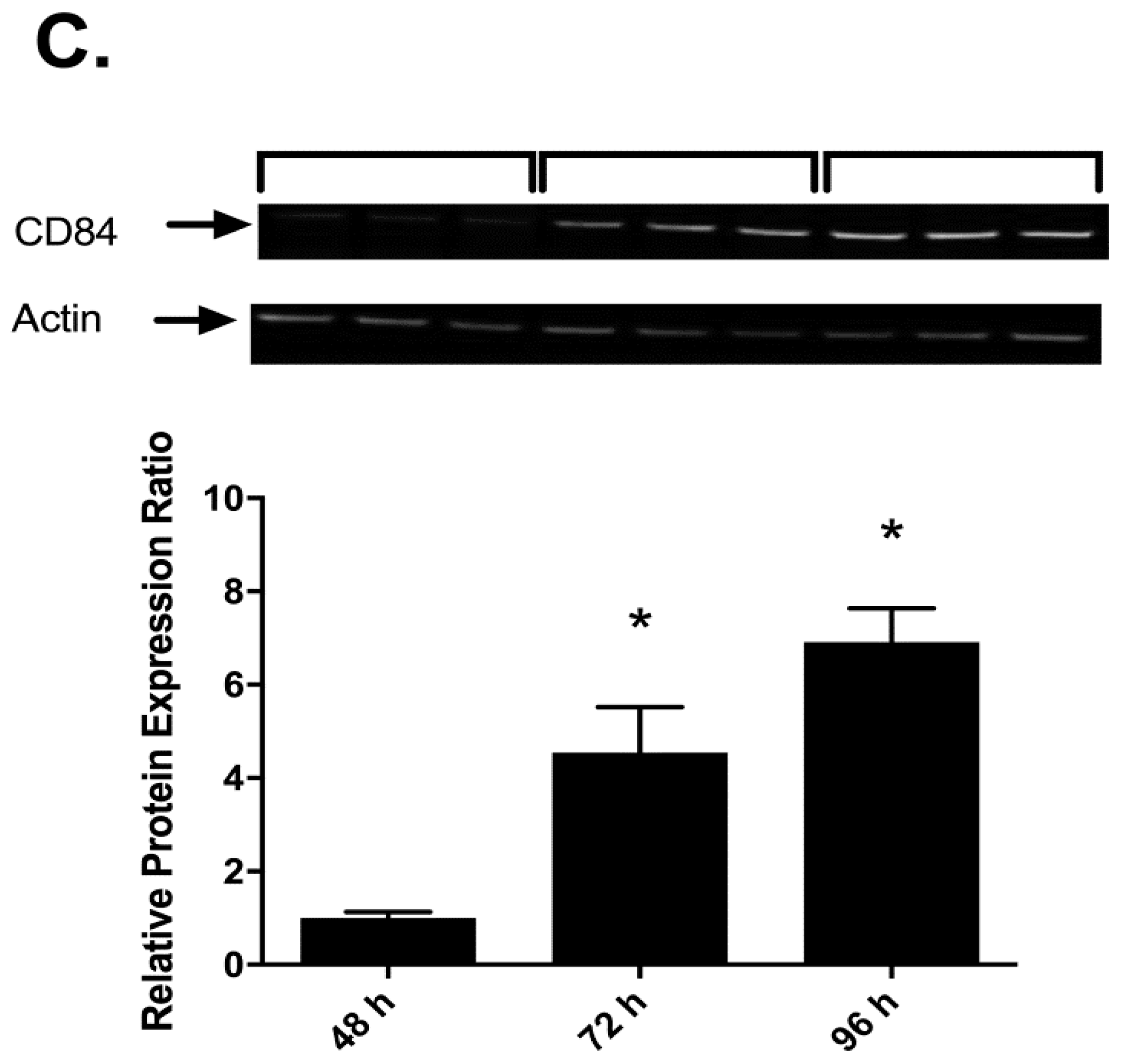
2.3. Effects of I3C, DIM and PMA on CD84 Protein in THP-1 Cells
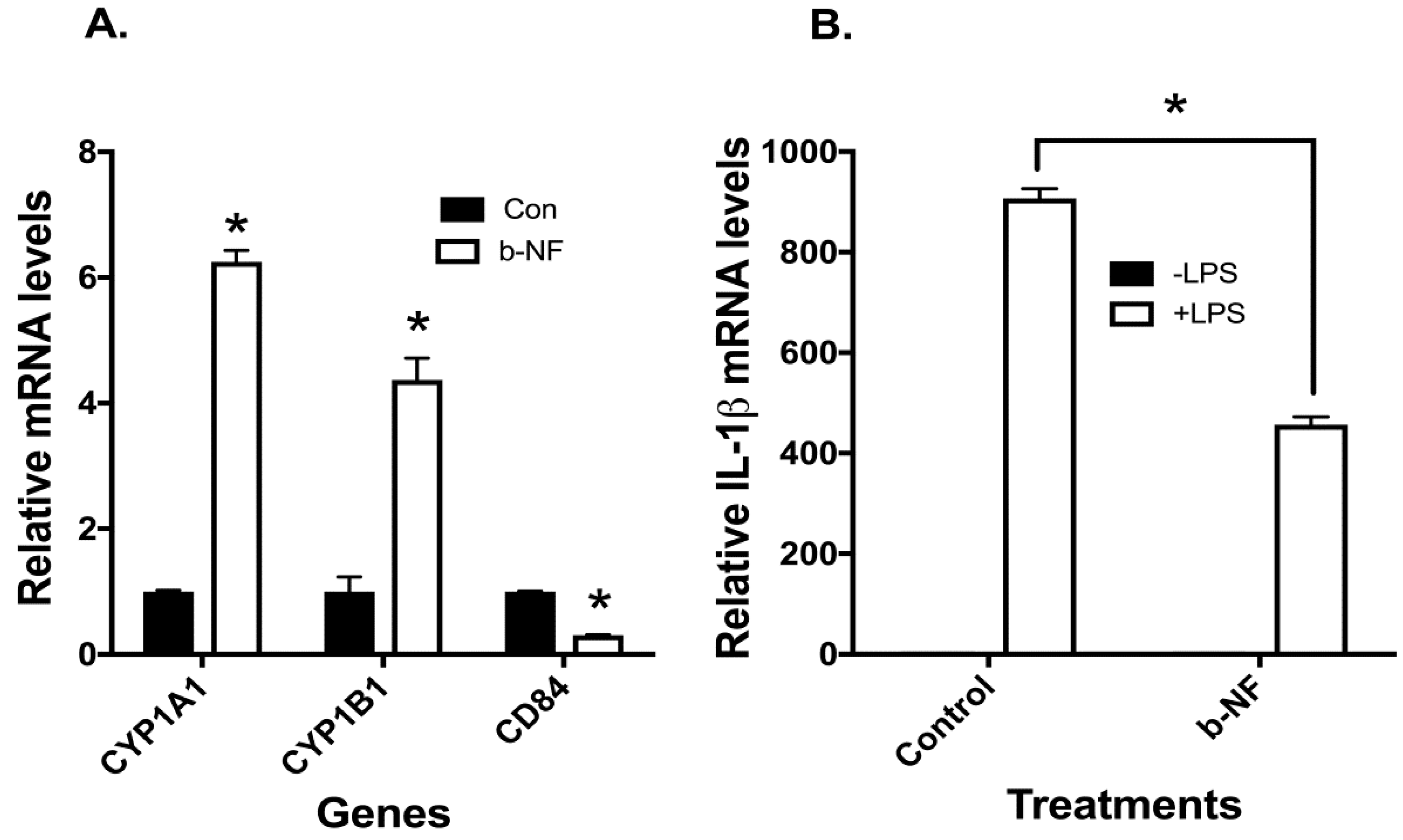
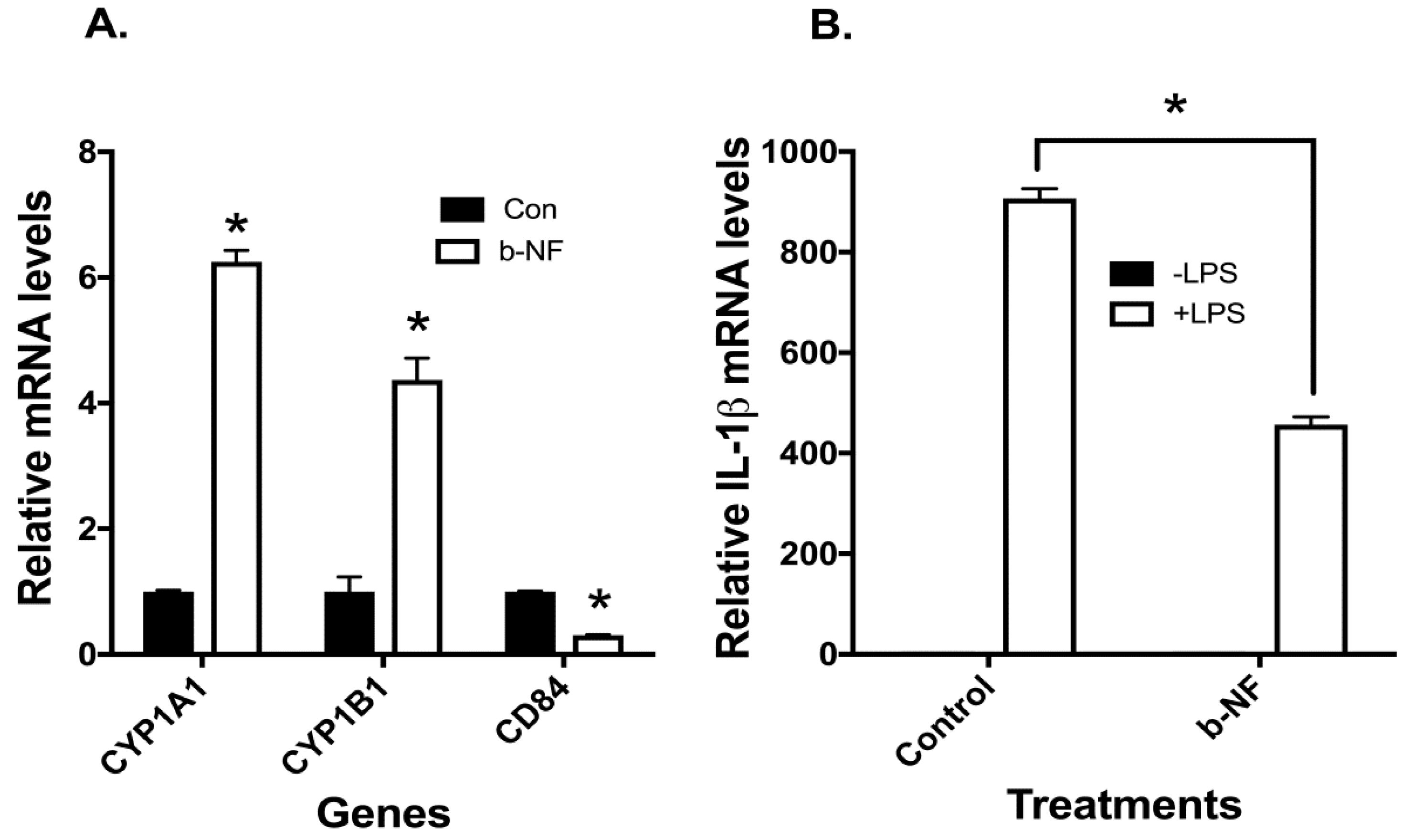
2.4. Effects of AHR Agonist beta-Naphthoflavone (b-NF) on CYP1A1/B1, CD84, and LPS-Induced IL-1β mRNA in u-THP-1 Cells
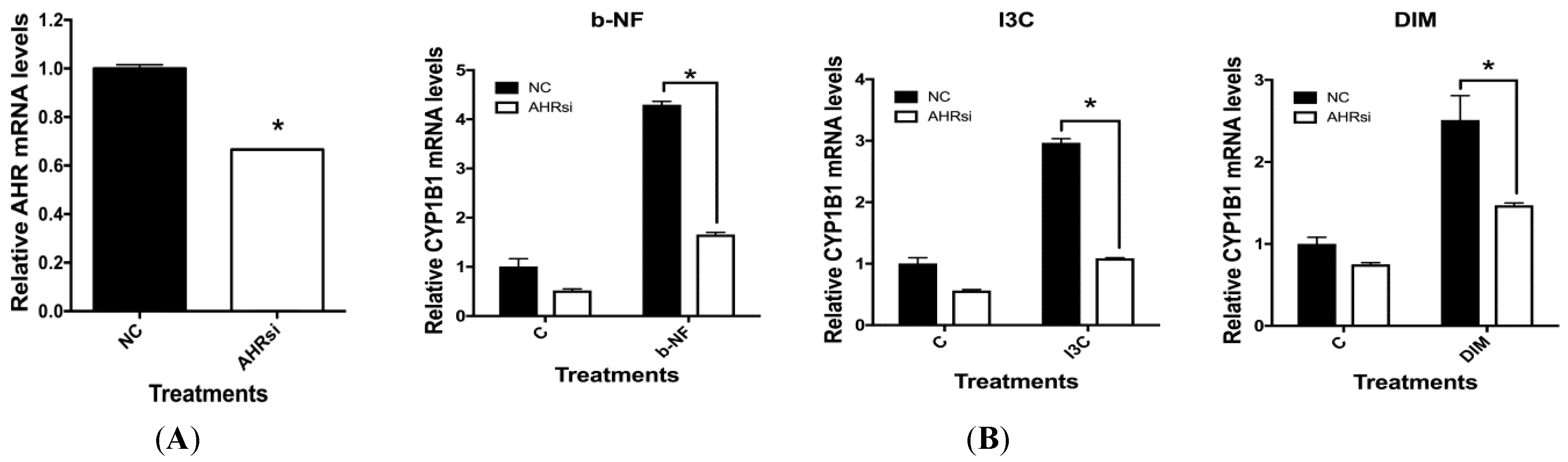
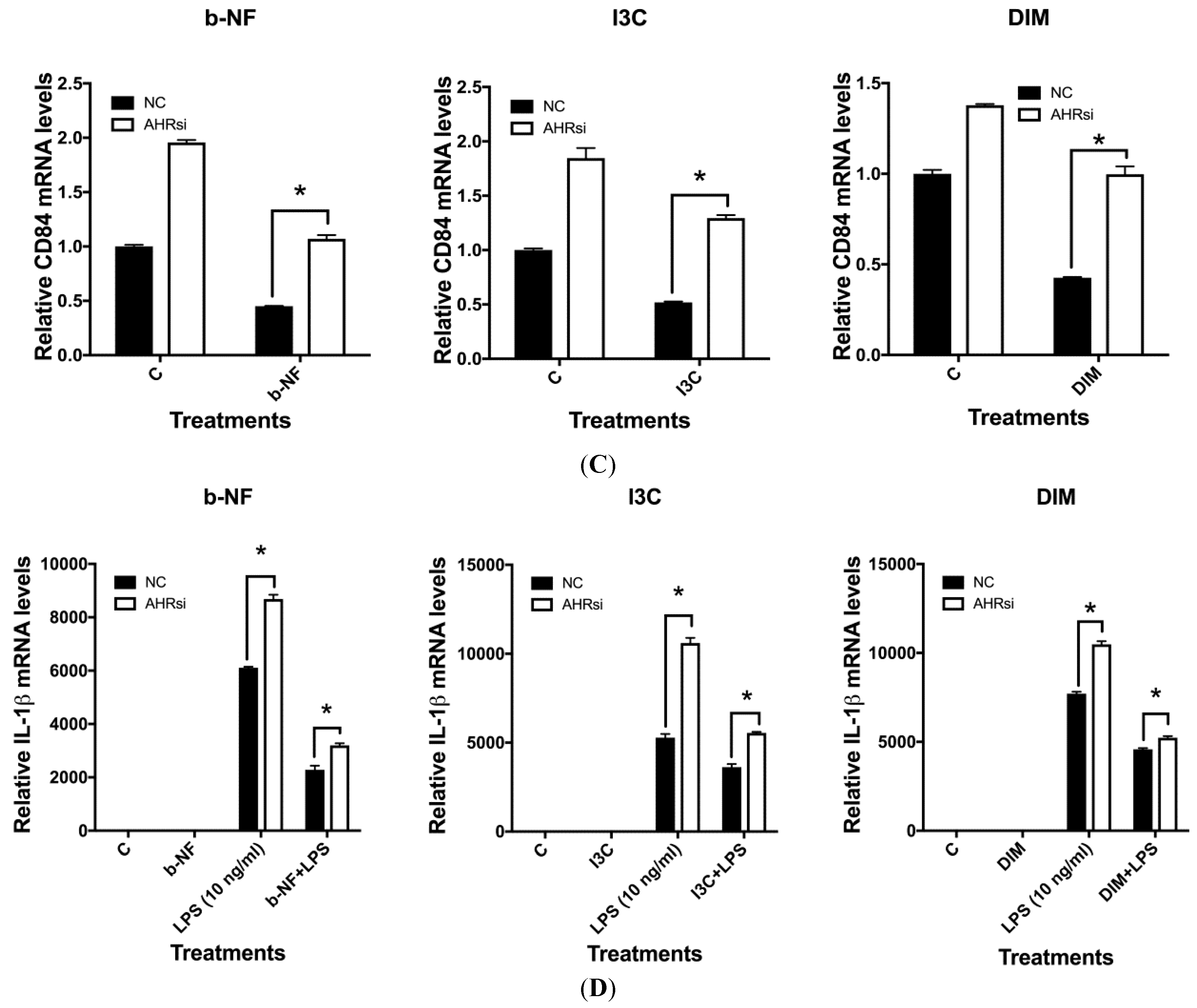
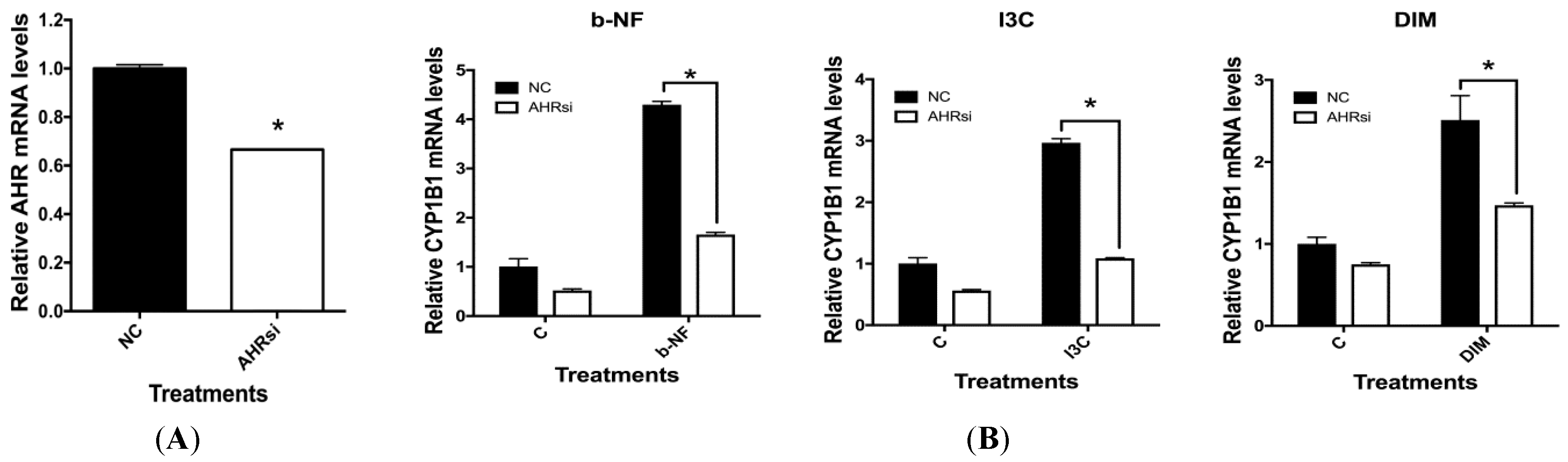
2.5. Effects of AHR siRNA on I3C, DIM and b-NF-Mediated Changes in IL-1β, CD84 and CYP1B1 mRNA
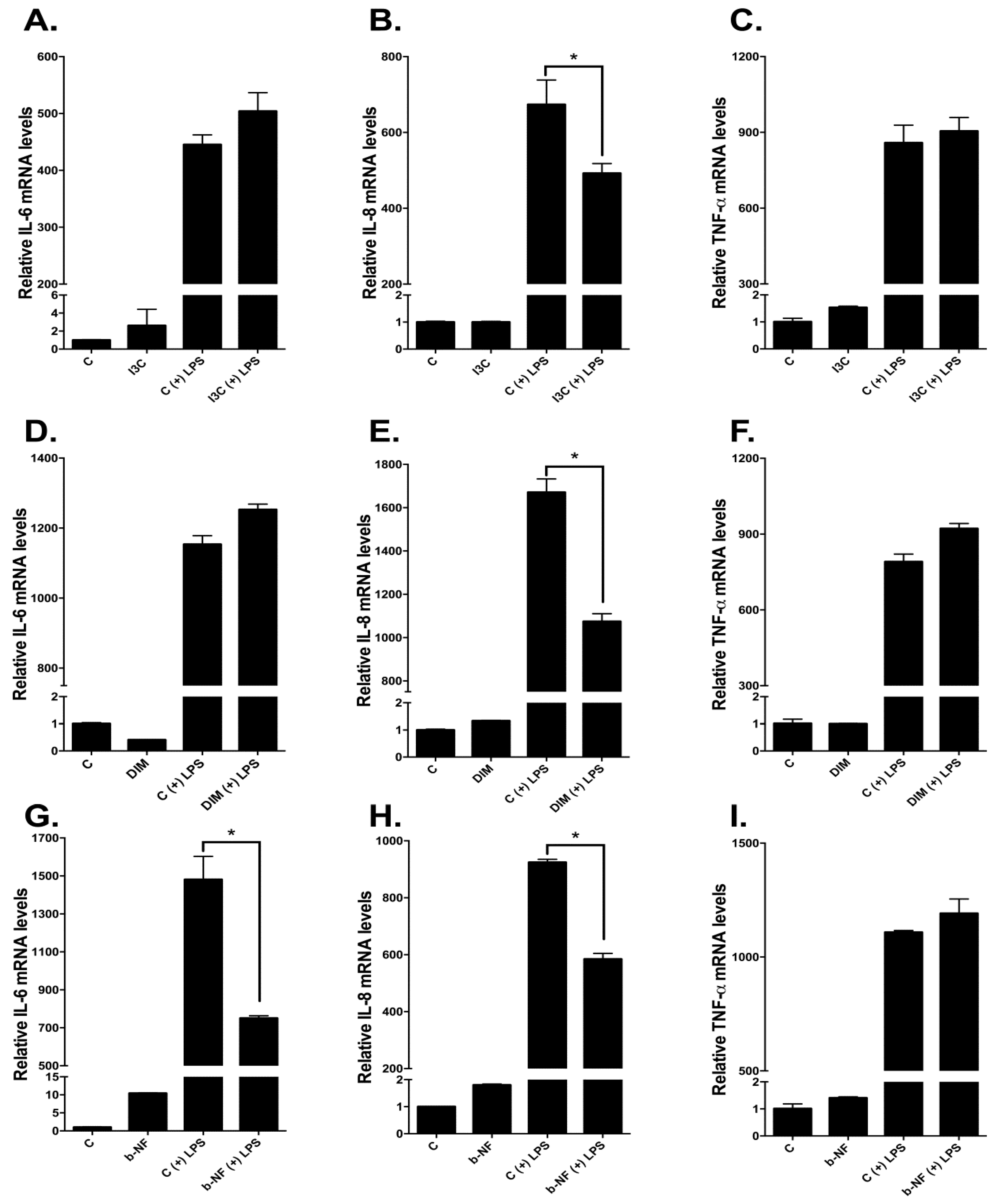
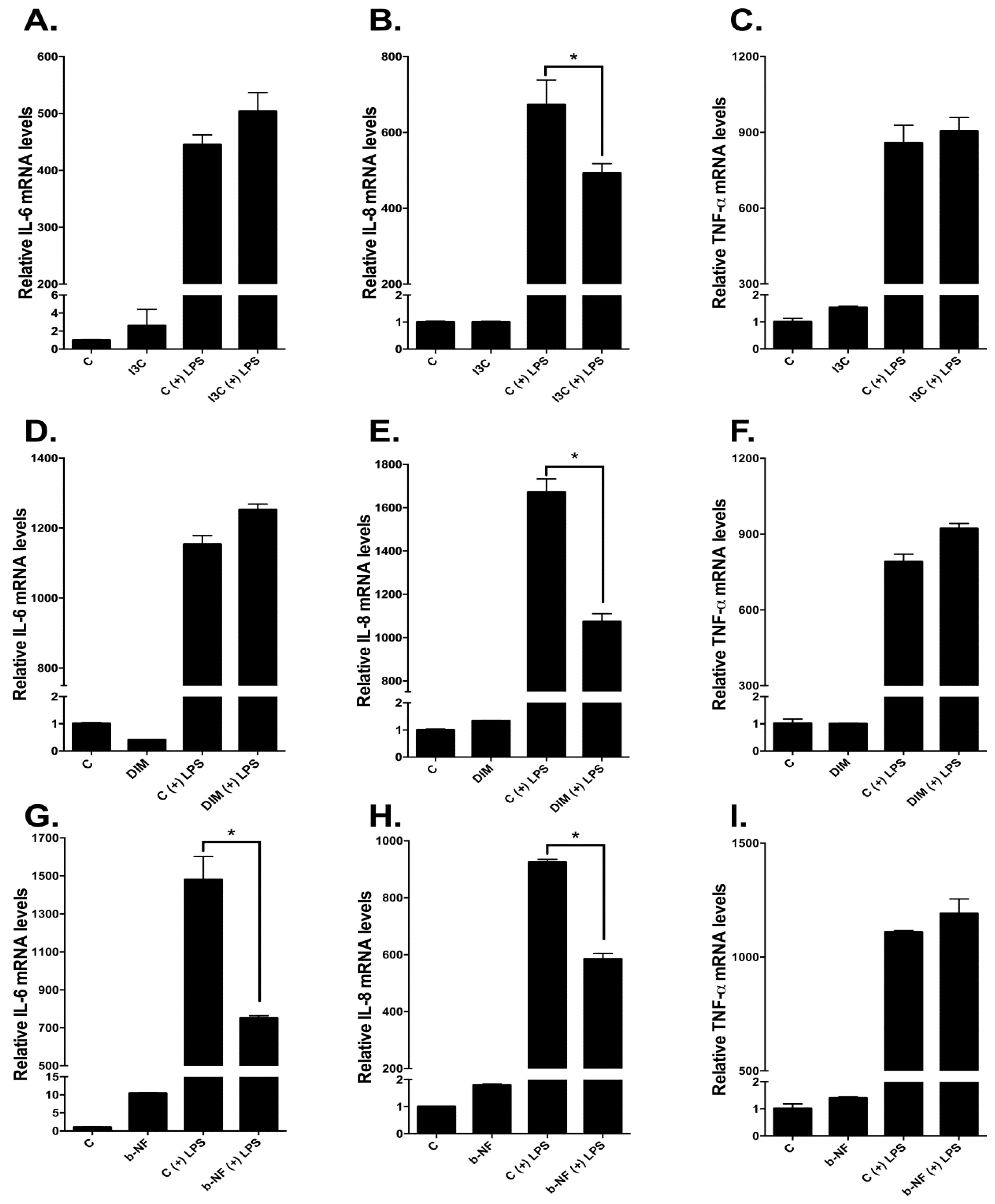
2.6. Effects of I3C, DIM, and b-NF on Other LPS-Inducible Cytokines
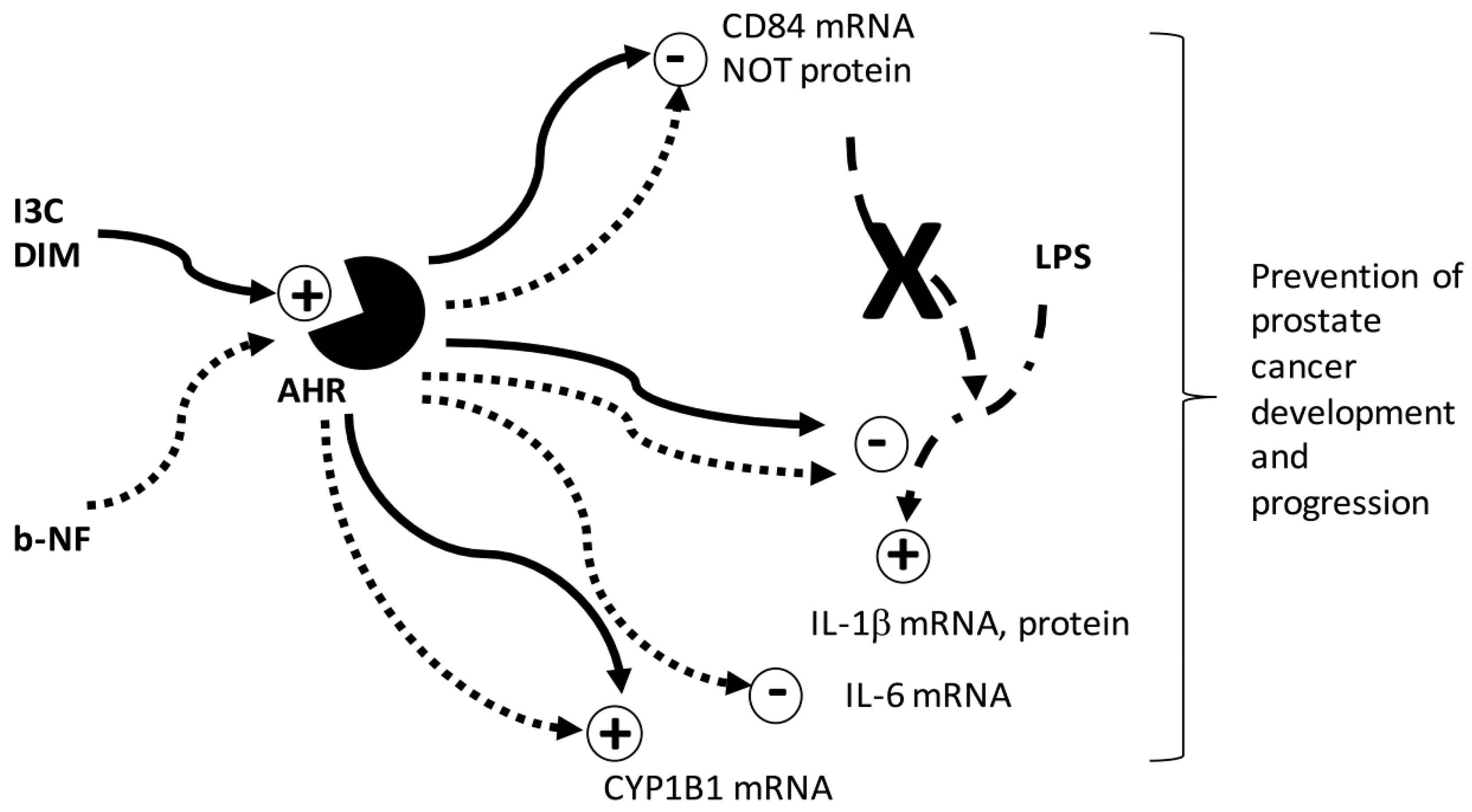
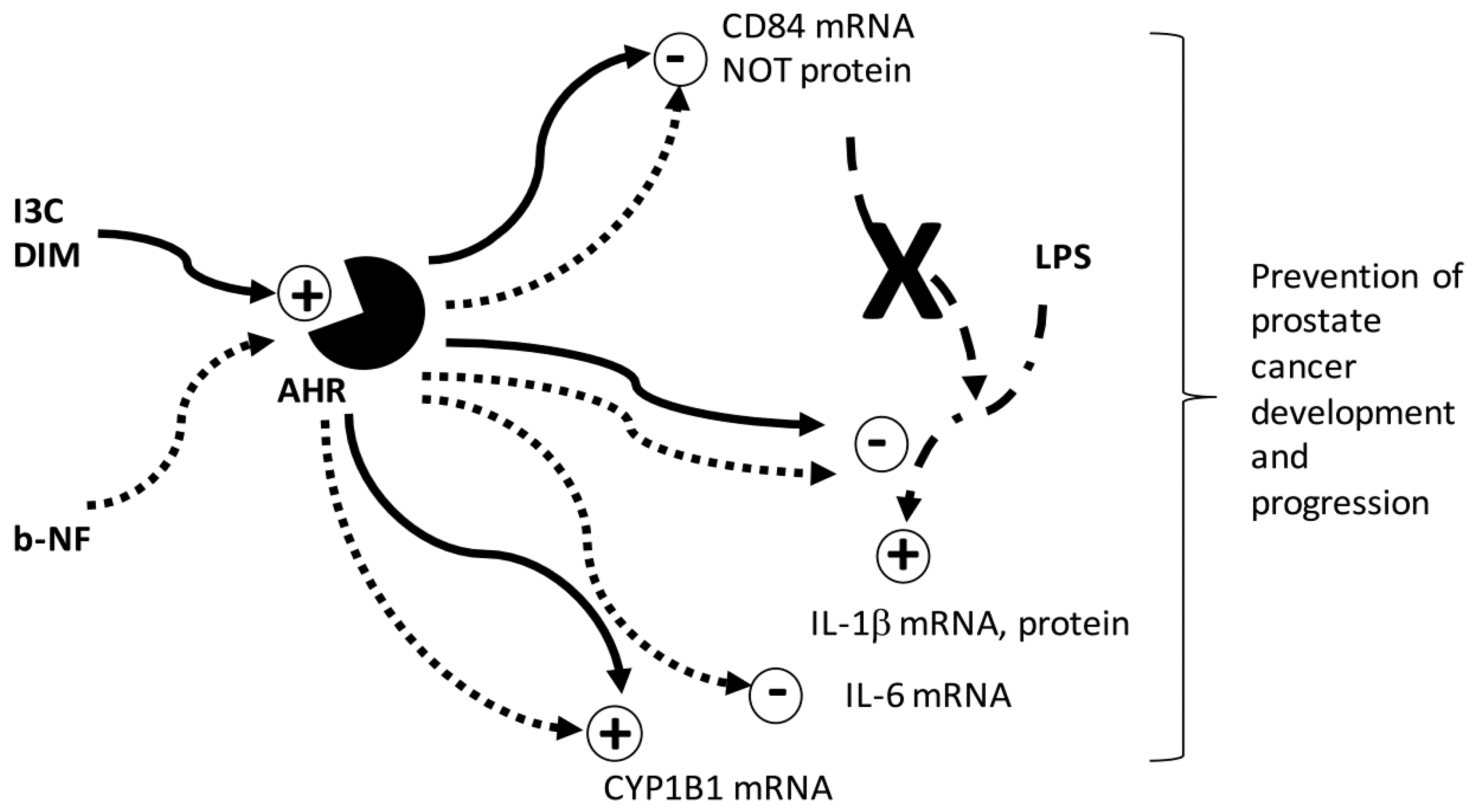
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. THP-1 Cell Differentiation
4.3. Cell Treatments
4.4. Total RNA Isolation, Reverse Transcription (RT)-PCR and Gene Expression Analysis
4.5. Determining Effects of I3C and DIM on IL-1β Protein Secretion by THP-1
4.6. Effects of AHR siRNA on I3C, DIM and β-Naphthoflavone-Mediated Changes
4.7. Western Blot Analysis of CD84 Protein Levels
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AHR | aryl hydrocarbon |
| b-NF | beta-naphthoflavone |
| CCL2/MCP-1 | C-C Motif Chemokine Ligand 2/Monocyte Chemoattractant Protein-1 |
| DIM | 3,3′-diindolylmethane |
| u-THP-1 | undifferentiated THP-1 cells |
| d-THP-1 | differentiated THP-1 cells |
| I3C | indole-3-carbinol |
| IL-1β | interleukin-1β |
| LPS | lipopolysaccharide |
| PMA | phorbol 12-myristate 13-acetate |
| SLAM | signaling lymphocyte activation molecule |
| TLR | toll-like receptor |
| TNF-α | tumor necrosis factor-α |
References
- Barron, D.A.; Rowley, D.R. The reactive stroma microenvironment and prostate cancer progression. Endocr. Relat. Cancer 2012, 19, R187–R204. [Google Scholar] [CrossRef] [PubMed]
- Northcott, J.M.; Northey, J.J.; Barnes, J.M.; Weaver, V.M. Fighting the force: Potential of homeobox genes for tumor microenvironment regulation. Biochim. Biophys. Acta 2015, 1855, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.L.; Chu, G.C.; Chung, L.W. Regulation of prostate cancer progression by the tumor microenvironment. Cancer Lett. 2016, 380, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Stark, T.; Livas, L.; Kyprianou, N. Inflammation in prostate cancer progression and therapeutic targeting. Transl. Androl. Urol. 2015, 4, 455–463. [Google Scholar] [PubMed]
- Serbina, N.V.; Jia, T.; Hohl, T.M.; Pamer, E.G. Monocyte-mediated defense against microbial pathogens. Annu. Rev. Immunol. 2008, 26, 421–452. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Helming, L.; Gordon, S. Alternative activation of macrophages: An immunologic functional perspective. Annu. Rev. Immunol. 2009, 27, 451–483. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.; Eubank, T.D.; Doseff, A.I. Monocytes and macrophages regulate immunity through dynamic networks of survival and cell death. J. Innate Immun. 2010, 2, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Sun, T.; Yu, X.H.; Yang, Y.X.; Yeo, A.E. The effects of TLR activation on T-cell development and differentiation. Clin. Dev. Immunol. 2012, 2012, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ding, J.L. The molecular mechanisms of TLR-signaling cooperation in cytokine regulation. Immunol. Cell Biol. 2016, 94, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.S.; Ho, B.; Leung, B.P.; Ding, J.L. TLR cross-talk confers specificity to innate immunity. Int. Rev. Immunol. 2014, 33, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Fletcher, A.; Niu, Y.; Wang, T.T.; Yu, L. Characterization of lipopolysaccharide-stimulated cytokine expression in macrophages and monocytes. Inflamm. Res. 2012, 61, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Shchors, K.; Evan, G. Tumor angiogenesis: Cause or consequence of cancer? Cancer Res. 2007, 67, 7059–7061. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Koh, Y.; Kim, D.W.; Ahn, Y.O.; Kim, T.M.; Han, S.W.; Oh, D.Y.; Lee, S.H.; Im, S.A.; Kim, T.Y.; et al. Clinical implications of VEGF, TGF-β1, and IL-1β in patients with advanced non-small cell lung cancer. Cancer Res. Treat. 2013, 45, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Sintes, J.; Romero, X.; de Salort, J.; Terhorst, C.; Engel, P. Mouse CD84 is a pan-leukocyte cell-surface molecule that modulates LPS-induced cytokine secretion by macrophages. J. Leukoc. Biol. 2010, 88, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.; Hirtreiter, C.; Rehli, M.; Rehm, A.; Kunz-Schughart, L.A.; Andreesen, R.; Hennemann, B. CD84 expression on human hematopoietic progenitor cells. Exp. Hematol. 2003, 31, 798–805. [Google Scholar] [CrossRef]
- Quintana, F.J.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 2013, 65, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Hanieh, H.; Nakahama, T.; Kishimoto, T. The roles of aryl hydrocarbon receptor in immune responses. Int. Immunol. 2013, 25, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Naka, T.; Nakahama, T.; Chinen, I.; Masuda, K.; Nohara, K.; Fujii-Kuriyama, Y.; Kishimoto, T. Aryl hydrocarbon receptor in combination with Stat1 regulates LPS-induced inflammatory responses. J. Exp. Med. 2009, 206, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Sekine, H.; Mimura, J.; Oshima, M.; Okawa, H.; Kanno, J.; Igarashi, K.; Gonzalez, F.J.; Ikuta, T.; Kawajiri, K.; Fujii-Kuriyama, Y. Hypersensitivity of aryl hydrocarbon receptor-deficient mice to lipopolysaccharide induced septic shock. Mol. Cell. Biol. 2009, 29, 6391–6400. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Kimura, A.; Hanieh, H.; Nguyen, N.T.; Nakahama, T.; Chinen, I.; Otoyo, Y.; Murotani, T.; Yamatodani, A.; Kishimoto, T. Aryl hydrocarbon receptor negatively regulates LPS-induced IL-6 production through suppression of histamine production in macrophages. Int. Immunol. 2011, 23, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, D.T.; Goldbohm, R.A.; van Poppel, G.; Verhagen, H.; van den Brandt, P.A. Epidemiological studies on brassica vegetables and cancer risk. Cancer Epidemiol. Biomark. Prev. 1996, 5, 733–748. [Google Scholar]
- Hong, C.; Kim, H.A.; Firestone, G.L.; Bjeldanes, L.F. 3,3’-Diindolylmethane (DIM) induces a G(1) cell cycle arrest in human breast cancer cells that is accompanied by Sp1-mediated activation of p21(WAF1/CIP1) expression. Carcinogenesis 2002, 23, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; McDougal, A.; Wang, F.; Safe, S. Aryl hydrocarbon receptor-mediated antiestrogenic and antitumorigenic activity of diindolylmethane. Carcinogenesis 1998, 19, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Schoene, N.W.; Milner, J.A.; Kim, Y.S. Broccoli-derived phytochemicals indole-3-carbinol and 3,3’-diindolylmethane exerts concentration-dependent pleiotropic effects on prostate cancer cells: Comparison with other cancer preventive phytochemicals. Mol. Carcinog. 2012, 51, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Rogan, E.G. The natural chemopreventive compound Indole-3-carbinol: State of the science. In Vivo 2006, 20, 221–228. [Google Scholar] [PubMed]
- Wu, T.Y.; Saw, C.L.; Khor, T.O.; Pung, D.; Boyanapalli, S.S.; Kong, A.N. In vivo pharmacodynamics of indole-3-carbinol in the inhibition of prostate cancer in transgenic adenocarcinoma of mouse prostate (TRAMP) mice: Involvement of Nrf2 and cell cycle/apoptosis signaling pathways. Mol. Carcinog. 2012, 51, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Beaver, L.M.; Yu, T.W.; Sokolowski, E.I.; Williams, D.E.; Dashwood, R.H.; Ho, E. 3,3’-Diindolylmethane, but not indole-3-carbinol, inhibits histone deacetylase activity in prostate cancer cells. Toxicol. Appl. Pharmacol. 2012, 263, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Feng, Z.; Narod, S.A. Multiple therapeutic and preventive effects of 3,3’-diindolylmethane on cancers including prostate cancer and high grade prostatic intraepithelial neoplasia. J. Biomed. Res. 2014, 28, 339–348. [Google Scholar] [PubMed]
- Kim, E.K.; Kim, Y.S.; Milner, J.A.; Wang, T.T. Indole-3-carbinol and 3’,3’-diindolylmethane modulate androgen’s effect on C-C chemokine ligand 2 and monocyte attraction to prostate cancer cells. Cancer Prev. Res. (Phila) 2013, 6, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, S.; Bilan, P.J.; Pillon, N.J.; Klip, A. Contracting C2C12 myotubes release CCL2 in an NF-κB-dependent manner to induce monocyte chemoattraction. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E160–E170. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Wang, T. The suppressive effects of Indole-3-carbinol (I3C)/3,3’ diindolylmethane (DIM) on LPS-induced IL1 release may be associated with changes in CD84 in THP-1 monocytes. FASEB J. 2015, 29 (Suppl. S1), 593–595. [Google Scholar]
- Wang, C.; Xu, C.X.; Bu, Y.; Bottum, K.M.; Tischkau, S.A. Beta-naphthoflavone (DB06732) mediates estrogen receptor-positive breast cancer cell cycle arrest through AhR-dependent regulation of PI3K/AKT and MAPK/ERK signaling. Carcinogenesis 2014, 35, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Molecular biology of the Ah receptor and its role in carcinogenesis. Toxicol. Lett. 2001, 120, 1–7. [Google Scholar] [CrossRef]
- Köhle, C.; Bock, K.W. Activation of coupled Ah receptor and Nrf2 gene batteries by dietary phytochemicals in relation to chemoprevention. Biochem. Pharmacol. 2006, 72, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed]
- Aalinkeel, R.; Nair, B.; Chen, C.K.; Mahajan, S.D.; Reynolds, J.L.; Zhang, H.; Sun, H.; Sykes, D.E.; Chadha, K.C.; Turowski, S.G.; et al. Nanotherapy silencing the interleukin-8 gene produces regression of prostate cancer by inhibition of angiogenesis. Immunology 2016, 148, 387–406. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.T.Y.; Pham, Q.; Kim, Y.S. Elucidating the Role of CD84 and AHR in Modulation of LPS-Induced Cytokines Production by Cruciferous Vegetable-Derived Compounds Indole-3-Carbinol and 3,3′-Diindolylmethane. Int. J. Mol. Sci. 2018, 19, 339. https://doi.org/10.3390/ijms19020339
Wang TTY, Pham Q, Kim YS. Elucidating the Role of CD84 and AHR in Modulation of LPS-Induced Cytokines Production by Cruciferous Vegetable-Derived Compounds Indole-3-Carbinol and 3,3′-Diindolylmethane. International Journal of Molecular Sciences. 2018; 19(2):339. https://doi.org/10.3390/ijms19020339
Chicago/Turabian StyleWang, Thomas T. Y., Quynhchi Pham, and Young S. Kim. 2018. "Elucidating the Role of CD84 and AHR in Modulation of LPS-Induced Cytokines Production by Cruciferous Vegetable-Derived Compounds Indole-3-Carbinol and 3,3′-Diindolylmethane" International Journal of Molecular Sciences 19, no. 2: 339. https://doi.org/10.3390/ijms19020339