Ethanol Extract of Oldenlandia diffusa Herba Attenuates Scopolamine-Induced Cognitive Impairments in Mice via Activation of BDNF, P-CREB and Inhibition of Acetylcholinesterase


 ,
,
Abstract
:
1. Introduction
2. Results
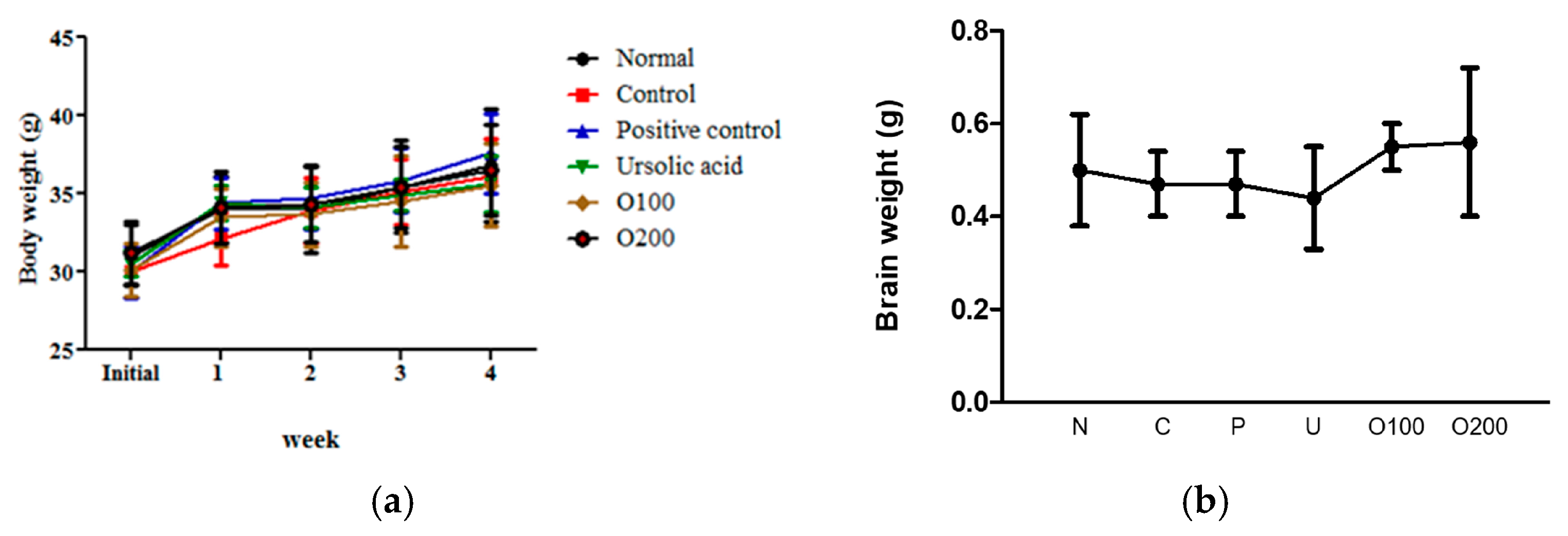
2.1. ODH Treatment Did Not Significantly Affect Body and Brain Weight Loss on ICR Mice
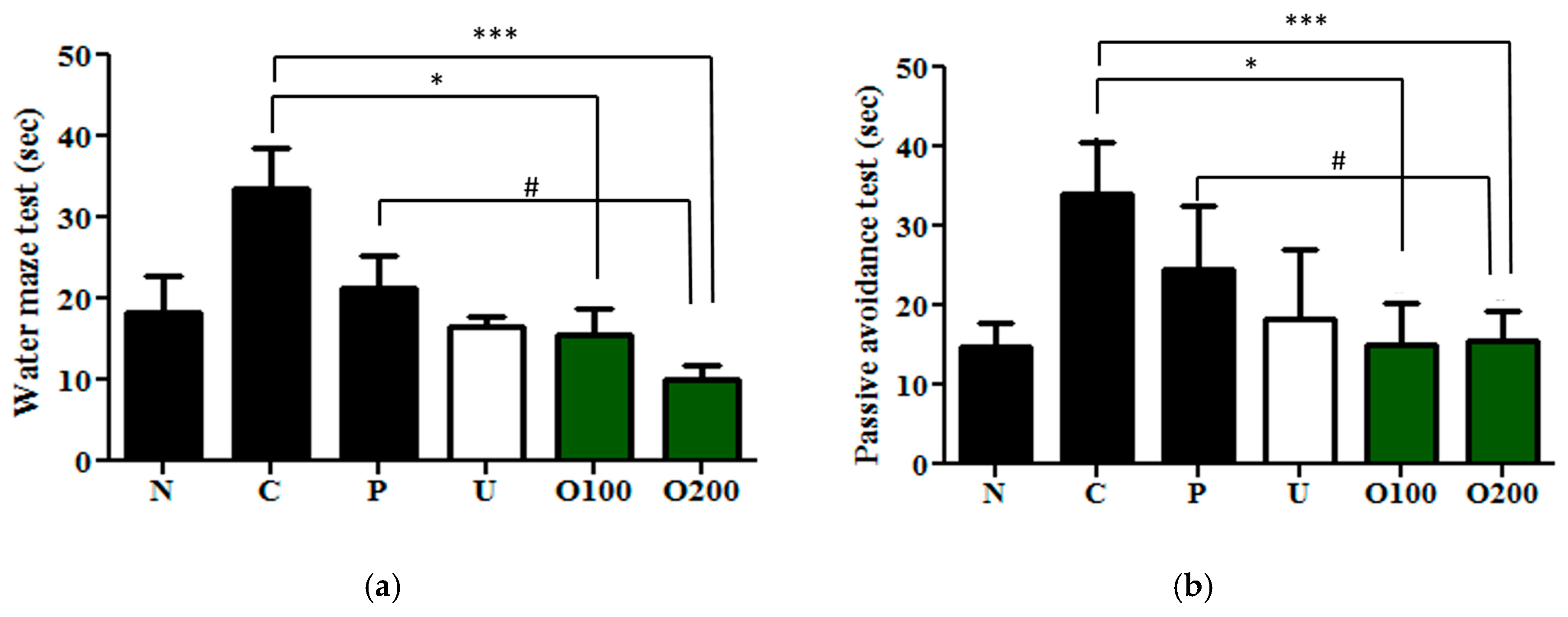
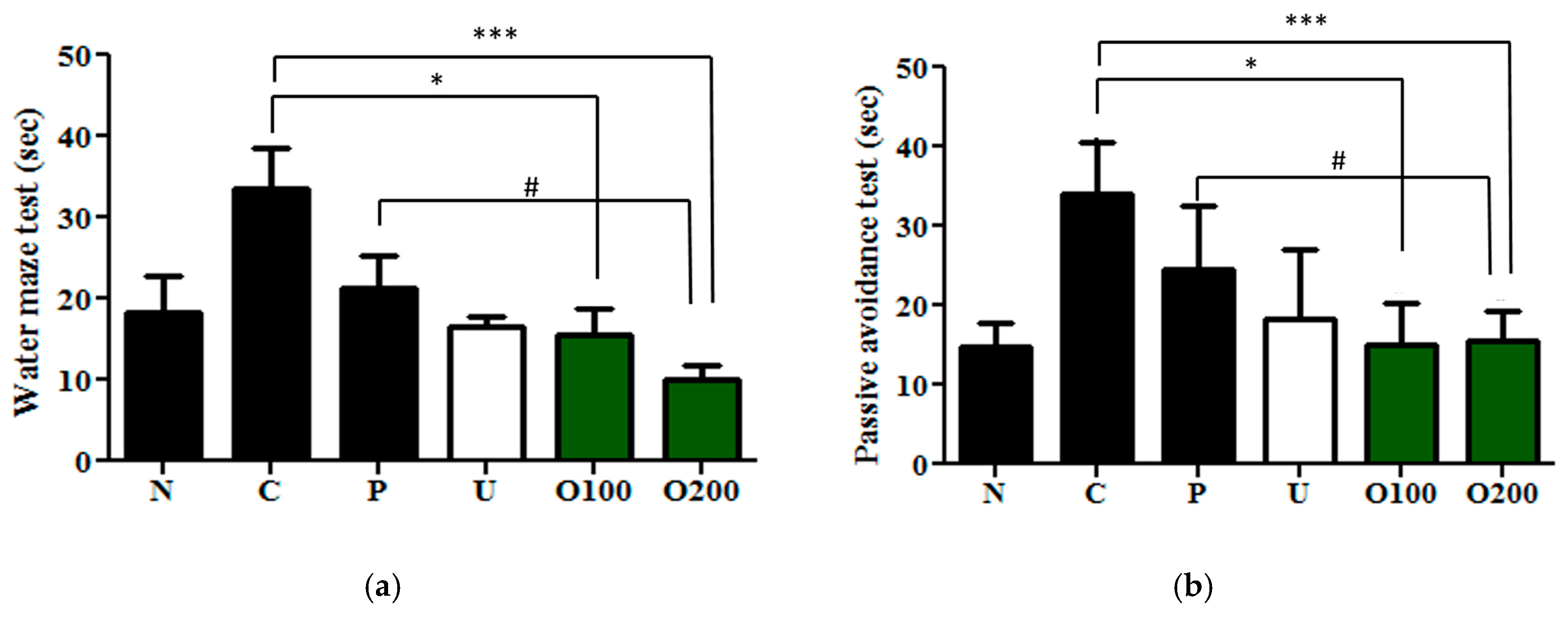
2.2. ODH Treatment Increased Learning and Memory Ability of ICR Mice
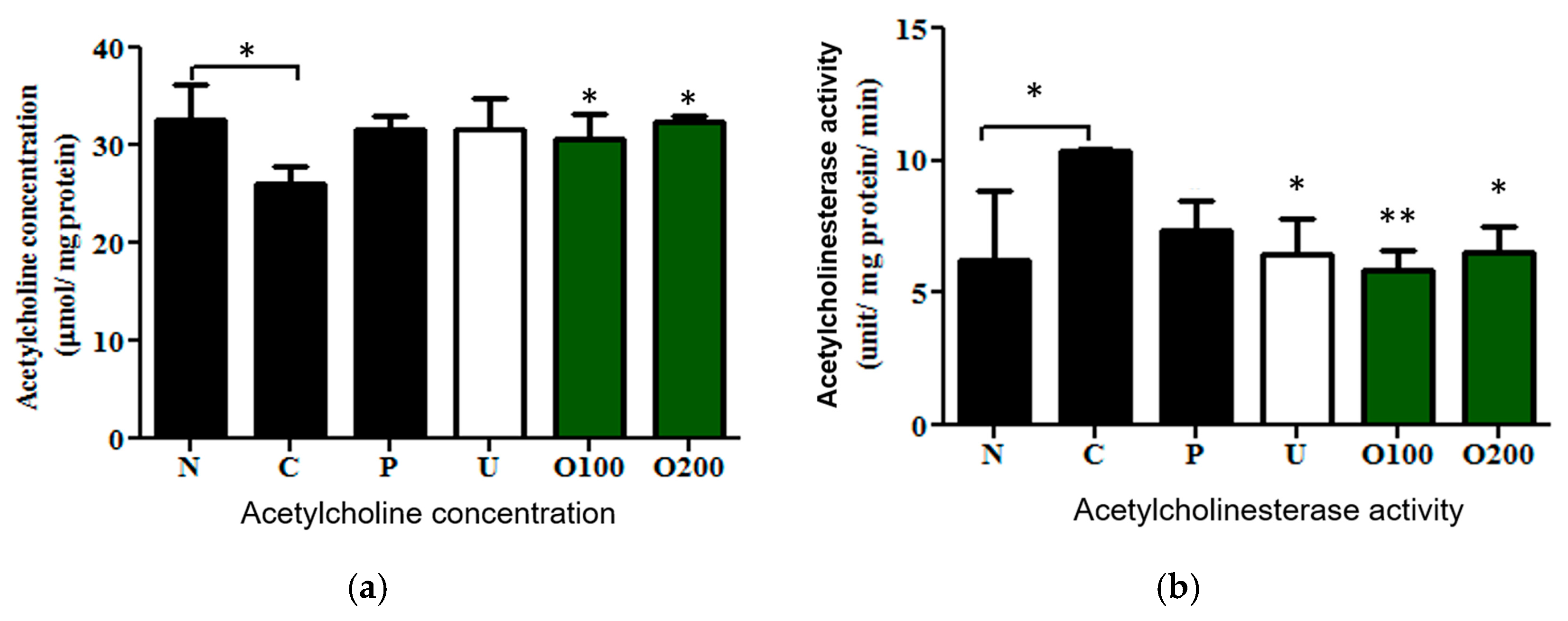
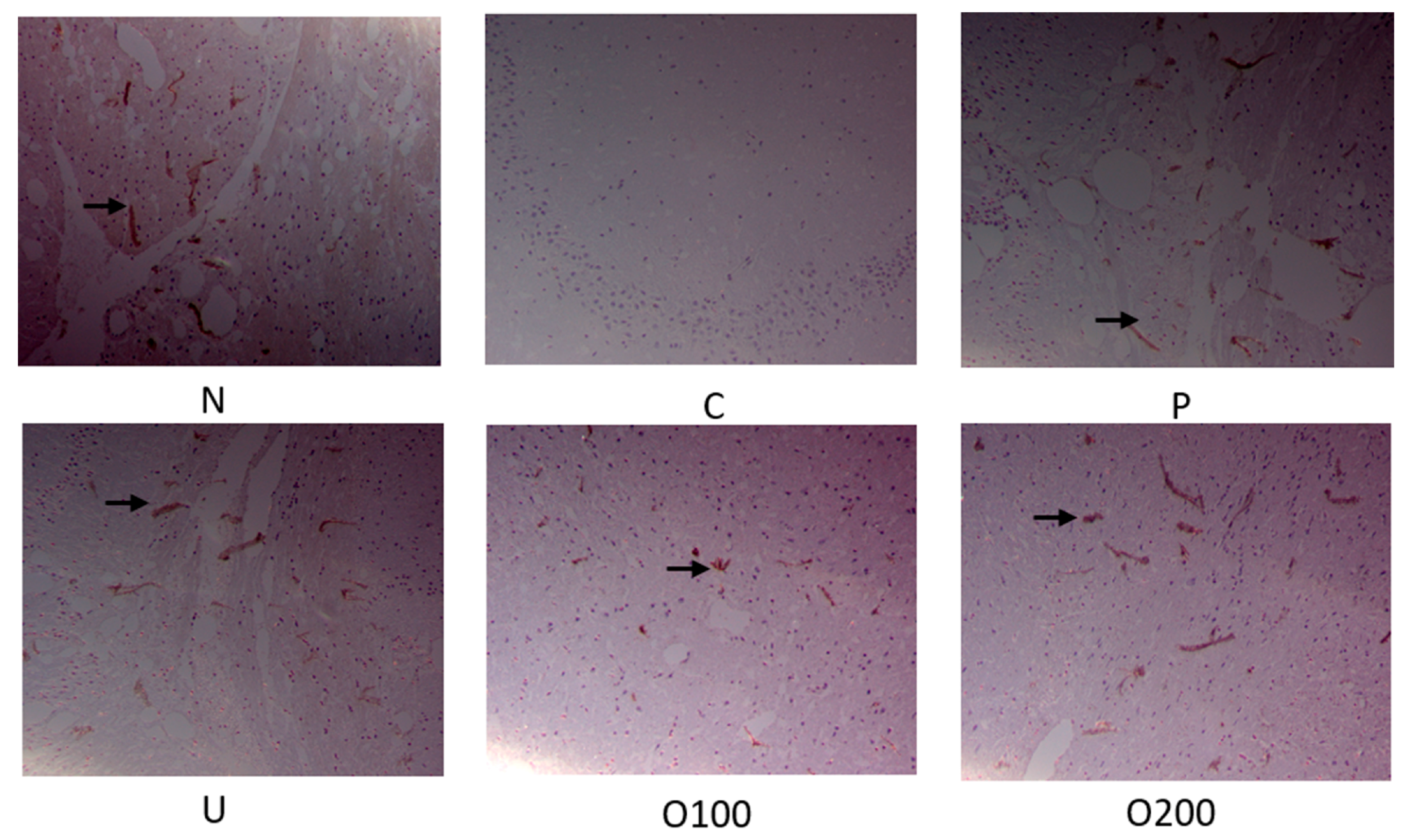
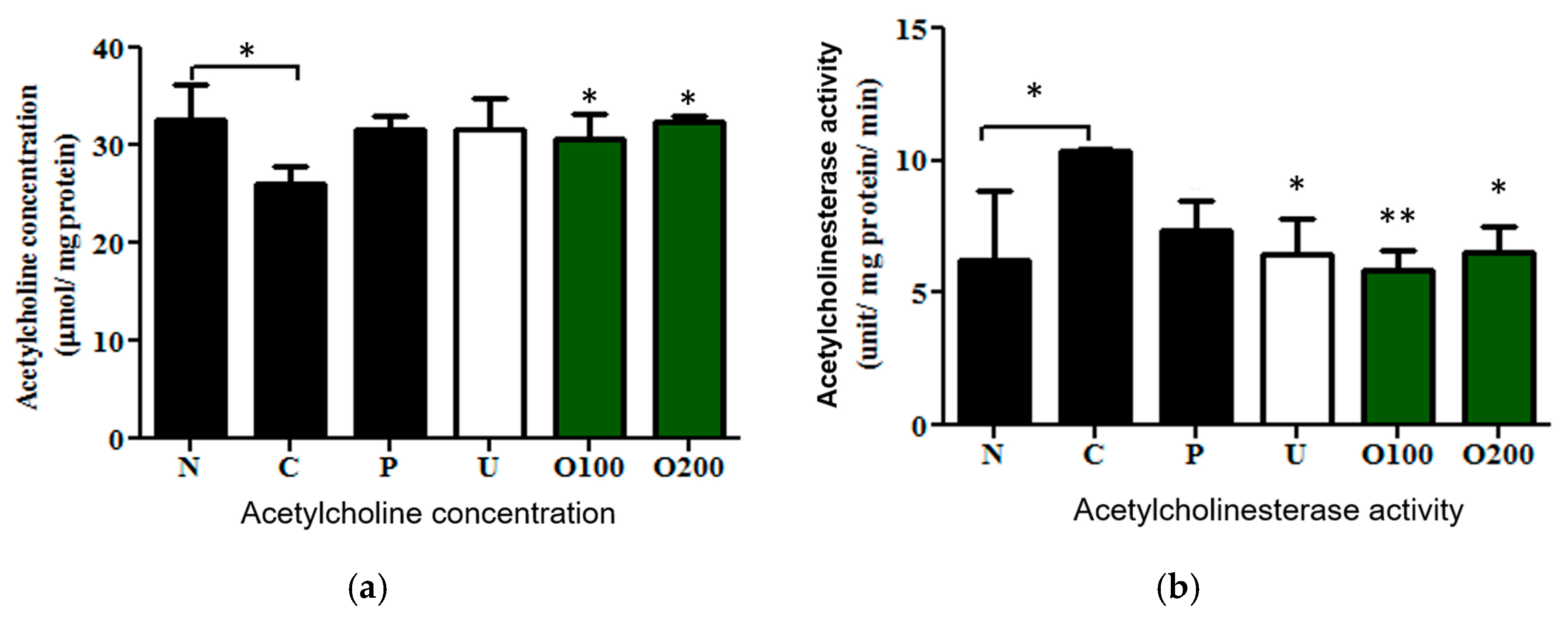
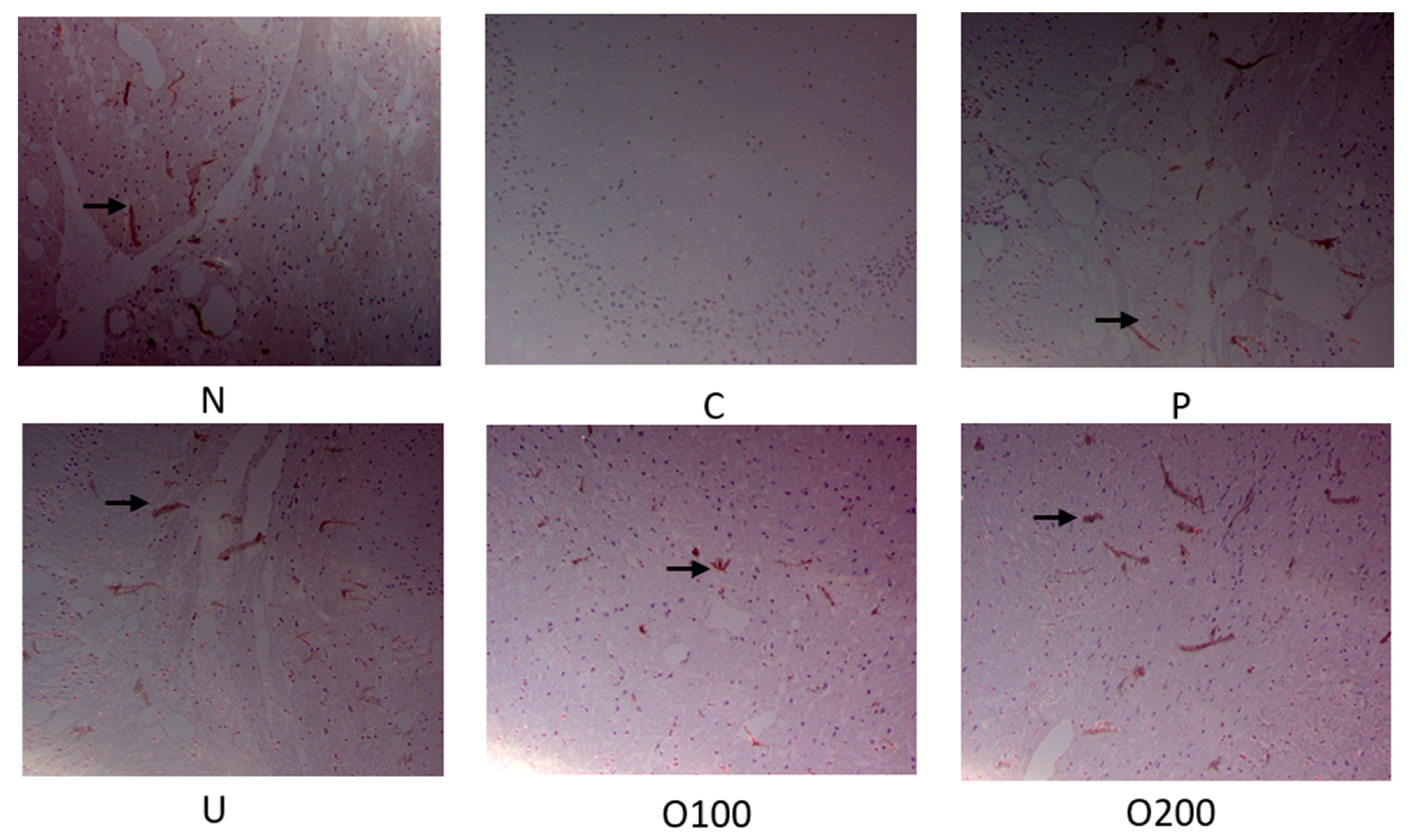
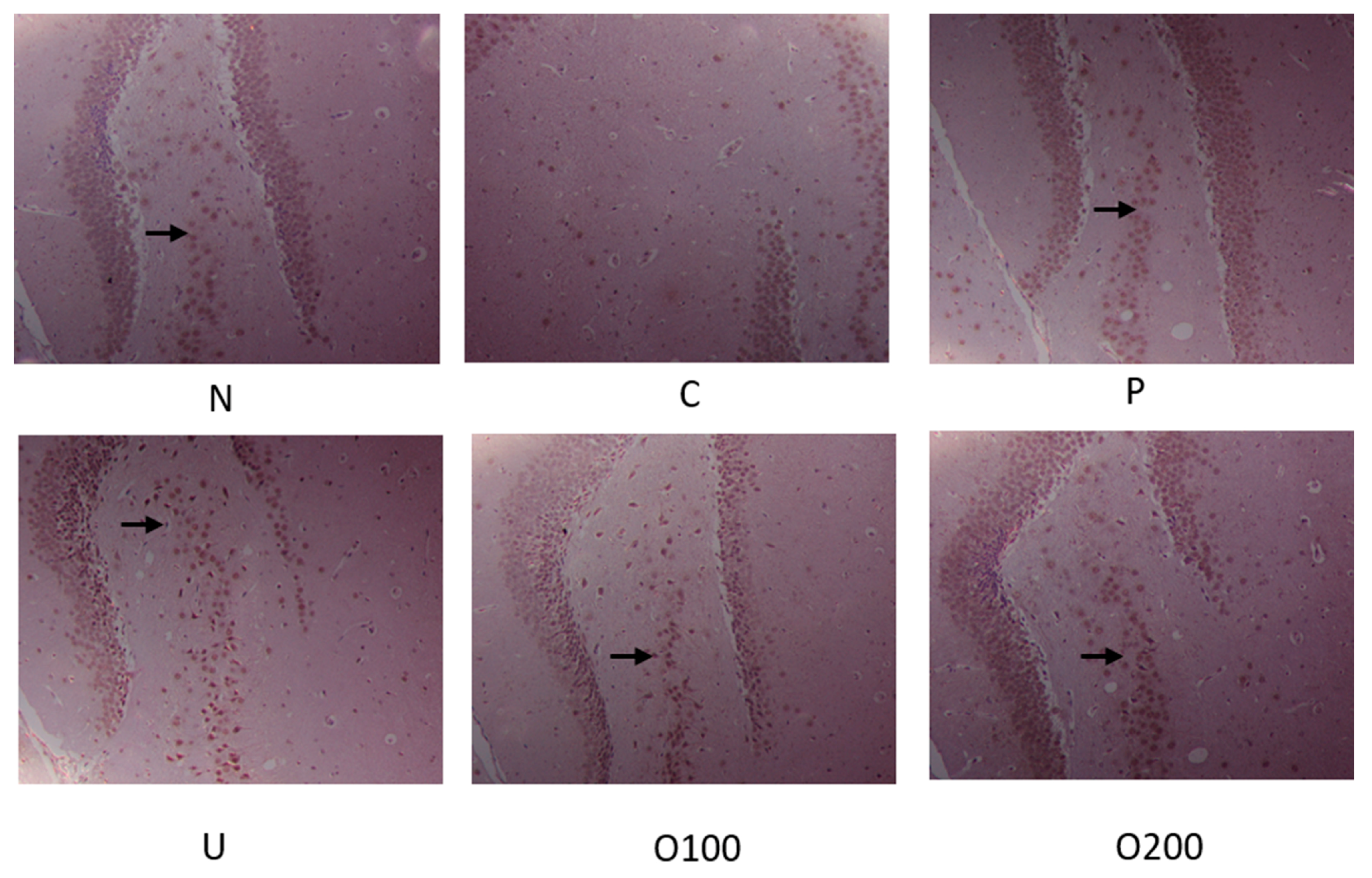
2.3. ODH Regulated Ach Concentration and AChE Activity in Scopolamine Treated Murine Brain
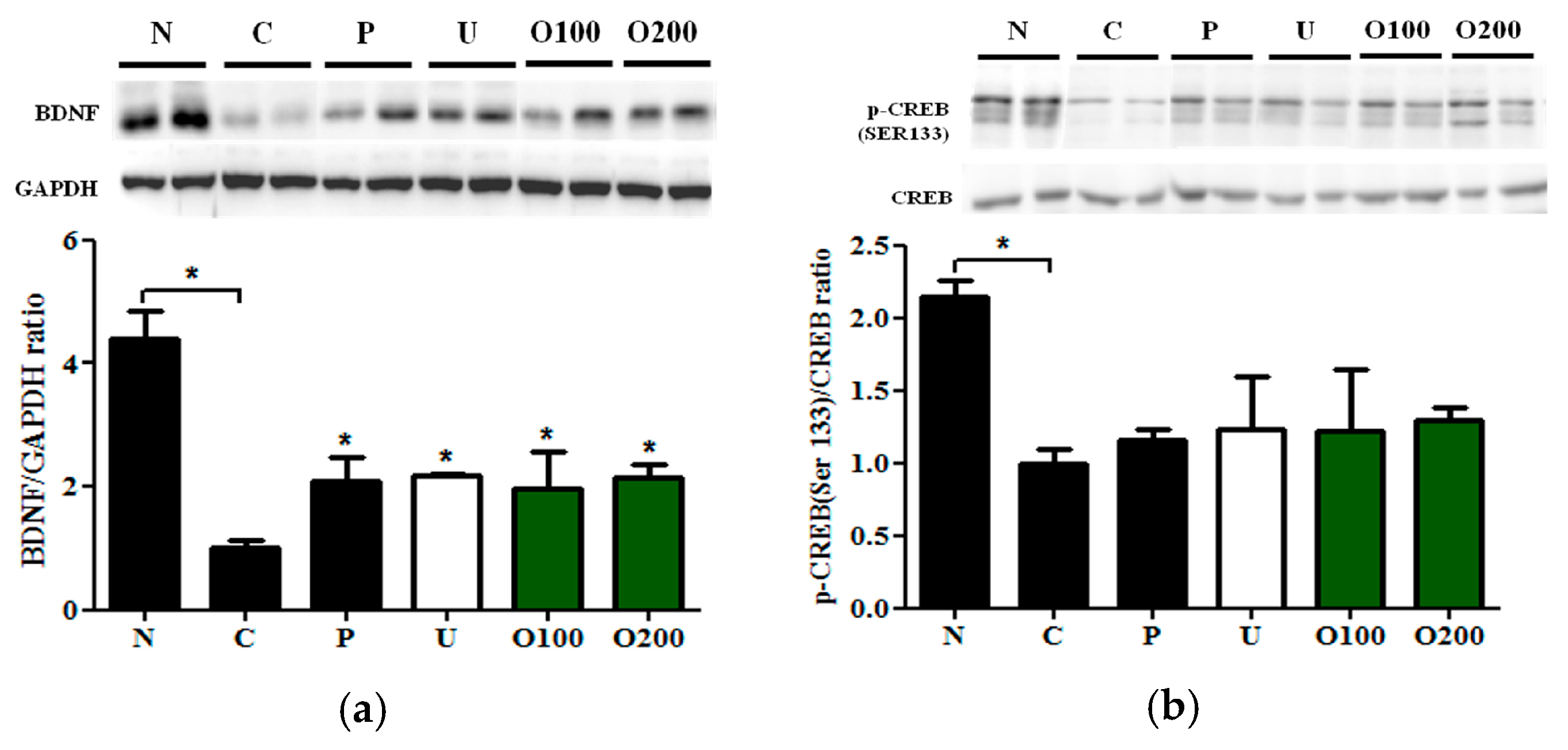
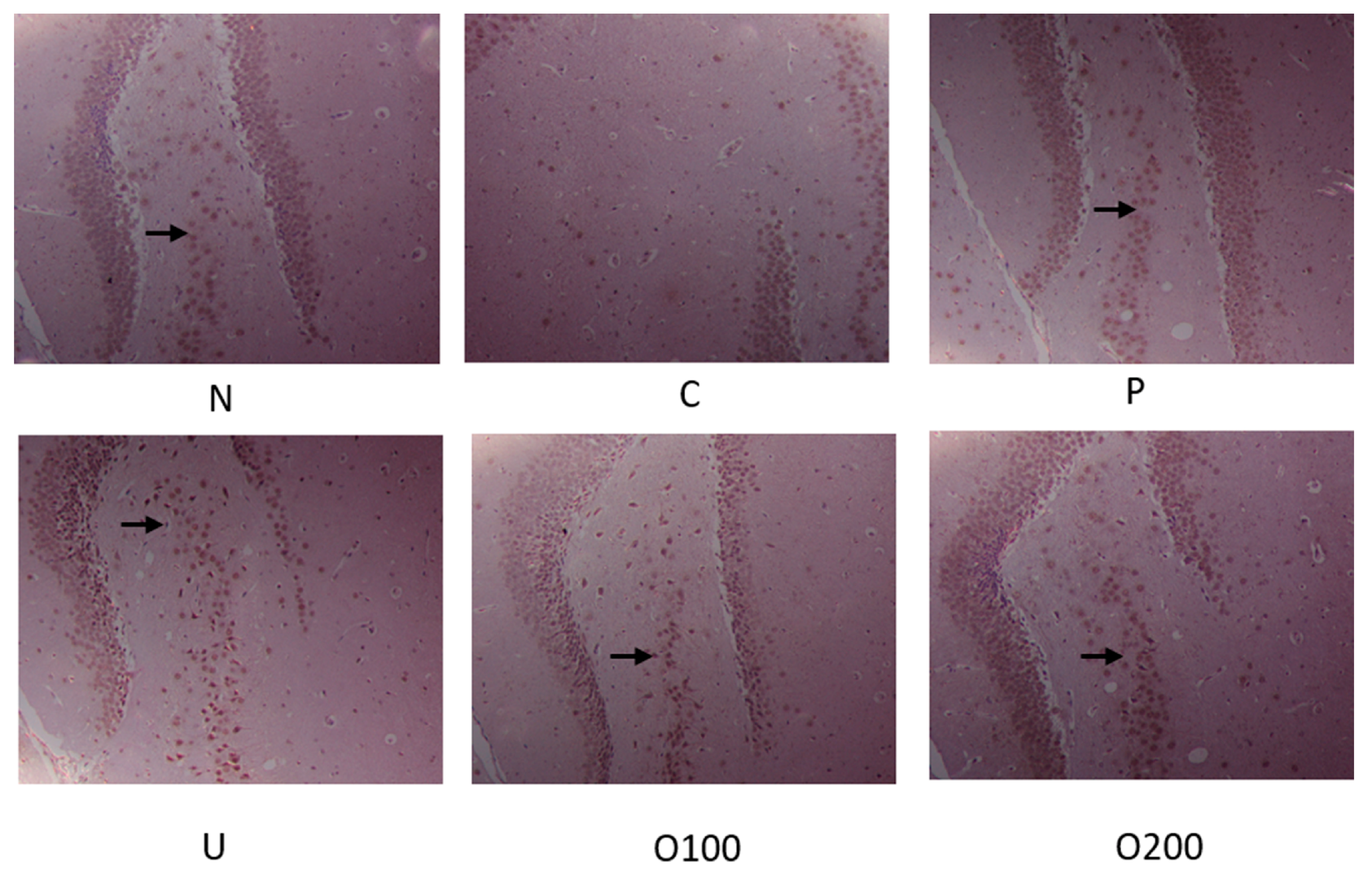
2.4. ODH Increased the Expression of BDNF and Phosphorylated CREB Protein Expression in Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
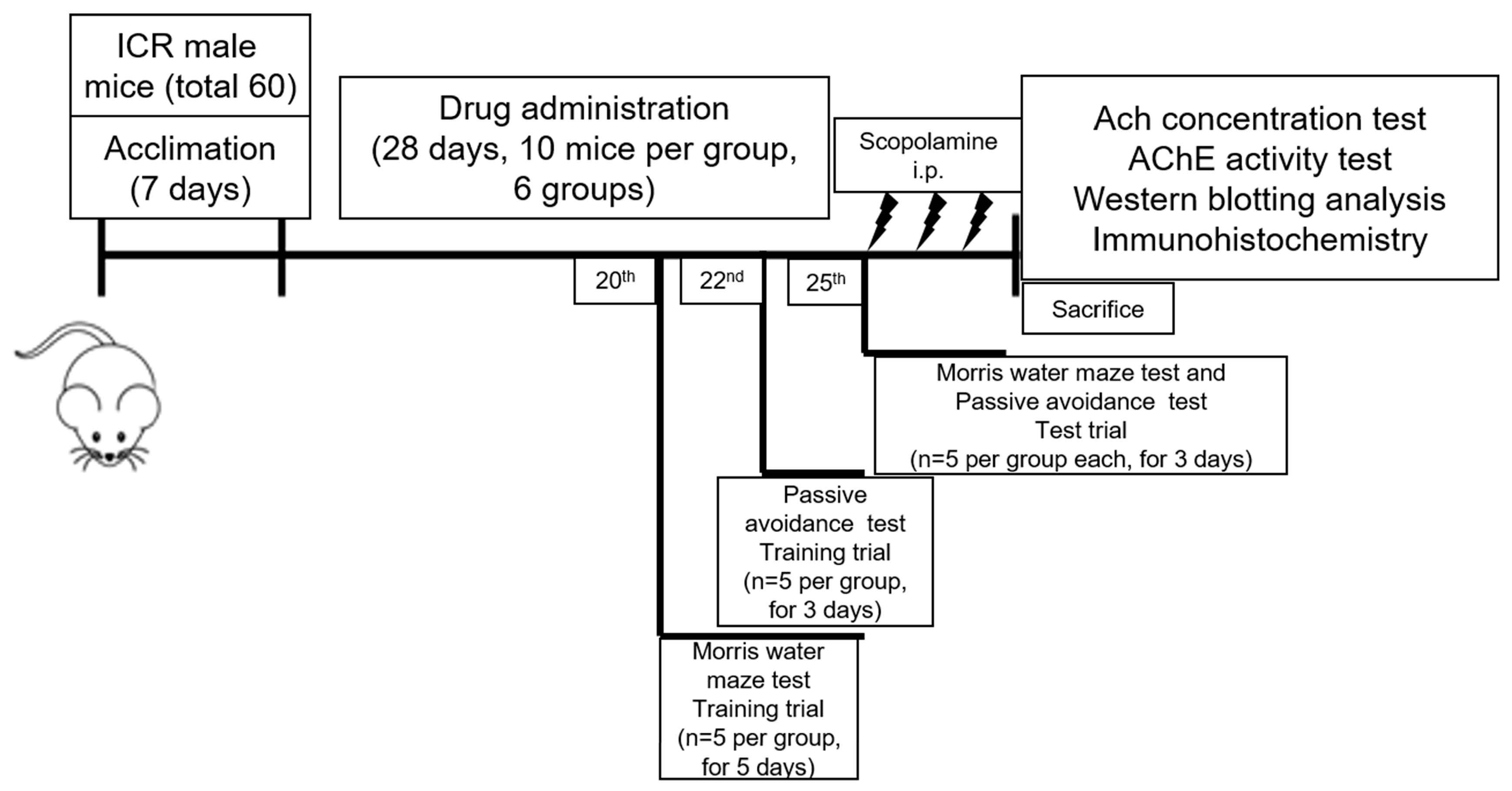
4.2. Animals
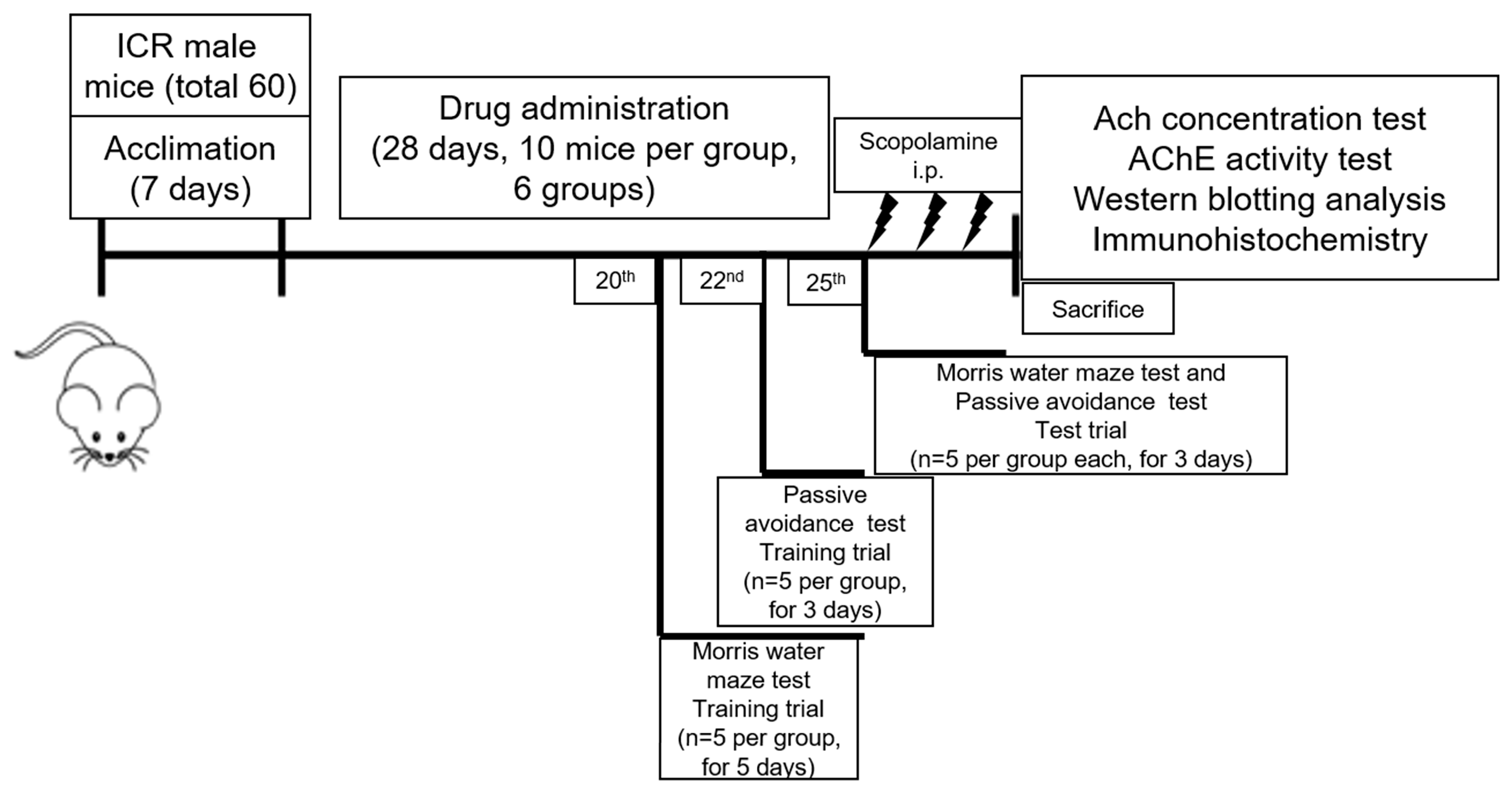
4.3. Drug Treatment
4.4. Morris Water Maze Test
4.5. Passive Avoidance Test
4.6. Tissue Preparation for Biochemical Analysis
4.7. Measurement of Acetylcholine Concentration
4.8. Measurement of Acetylcholinesterase Activity
4.9. Western Blot Analysis
4.10. Immunohistochemistry Staining Analysis
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Xian, Y.F.; Ip, S.P.; Mao, Q.Q.; Su, Z.R.; Chen, J.N.; Lai, X.P.; Lin, Z.X. Honokiol improves learning and memory impairments induced by scopolamine in mice. Eur. J. Pharmacol. 2015, 760, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.E.; Yaffe, K. The projected effect of risk factor reduction on Alzheimer’s disease prevalence. Lancet Neurol. 2011, 10, 819–828. [Google Scholar] [CrossRef]
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of Alzheimer’s disease. Alzheimers Dement. 2007, 3, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Tincer, G.; Mashkaryan, V.; Bhattarai, P.; Kizil, C. Neural stem/progenitor cells in Alzheimer’s disease. Yale J. Biol. Med. 2016, 89, 23–35. [Google Scholar] [PubMed]
- Arbor, S.C.; LaFontaine, M.; Cumbay, M. Amyloid-β Alzheimer targets-protein processing, lipid rafts, and amyloid-β pores. Yale J. Biol. Med. 2016, 89, 5–21. [Google Scholar] [PubMed]
- Sugisaki, E.; Fukushima, Y.; Fujii, S.; Yamazaki, Y.; Aihara, T. The effect of coactivation of muscarinic and nicotinic acetylcholine receptors on LTD in the hippocampal CA1 network. Brain Res. 2016, 1649, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Provensi, G.; Costa, A.; Passani, M.B.; Blandina, P. Donepezil, an acetylcholine esterase inhibitor, and ABT-239, a histamine H3 receptor antagonist/inverse agonist, require the integrity of brain histamine system to exert biochemical and procognitive effects in the mouse. Neuropharmacology 2016, 109, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol. 2006, 16, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Micheau, J.; Marighetto, A. Acetylcholine and memory: A long, complex and chaotic but still living relationship. Behav. Brain Res. 2010, 221, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Shen, J.; Meng, Z.; Yang, X.; Li, F. Neuroprotective effects of polygalacic acid on scopolamine-induced memory deficits in mice. Phytomedicine 2016, 23, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; He, X.H.; Yuan, R.; Wei, B.J.; Chen, Z.; Dong, J.X.; Wang, J. Sesquiterpenes and a monoterpenoid with acetylcholinesterase (AchE) inhibitory activity from valeriana officinalis var. Latiofolia in vitro and in vivo. Fitoterapia 2016, 110, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Altar, C.A.; Cai, N.; Bliven, T.; Juhasz, M.; Conner, J.M.; Acheson, A.L.; Lindsay, R.M.; Wiegand, S.J. Anterograde transport of brain-derived neurotrophic factor and its role in the brain. Nature 1997, 389, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Schuman, E.M. Long-lasting neurotrophin-induced enhancement of synaptic transmission in the adult hippocampus. Science 1995, 267, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Hock, C.; Heese, K.; Hulette, C.; Rosenberg, C.; Otten, U. Region-specific neurotrophin imbalances in Alzheimer disease: Decreased levels of brain-derived neurotrophic factor and increased levels of nerve growth factor in hippocampus and cortical areas. Arch. Neurol. 2000, 57, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Merrill, D.A.; Coppola, G.; Tsukada, S.; Schroeder, B.E.; Shaked, G.M.; Wang, L.; Blesch, A.; Kim, A.; Conner, J.M.; et al. Neuroprotective effects of brain-derived neurotrophic factor in rodent and primate models of Alzheimer’s disease. Nat. Med. 2009, 15, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.J.; Kogan, J.H.; Frankland, P.W.; Kida, S. CREB and memory. Annu. Rev. Neurosci. 1998, 21, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Motaghinejad, M.; Motevalian, M.; Fatima, S.; Faraji, F.; Mozaffari, S. The neuroprotective effect of curcumin against nicotine-induced neurotoxicity is mediated by CREB-BDNF signaling pathway. Neurochem. Res. 2017, 42, 2921–2932. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.W.; Wu, M.S.; Guo, J.D. Effects of microRNA-10a on synapse remodeling in hippocampal neurons and neuronal cell proliferation and apoptosis through the BDNF signaling pathway in a rat model of Alzheimer’s disease. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, H.G.; Lee, H.W.; Han, J.M.; Lee, S.K.; Kim, D.W.; Saravanakumar, A.; Son, C.G. Hippocampal memory enhancing activity of pine needle extract against scopolamine-induced amnesia in a mouse model. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Jackson, P.A.; Forster, J.; Khan, J.; Grothe, T.; Perrinjaquet-Moccetti, T.; Haskell-Ramsay, C.F. Acute effects of a wild green-oat (Avena sativa) extract on cognitive function in middle-aged adults: A double-blind, placebo-controlled, within-subjects trial. Nutr. Neurosci. 2015, 20, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.S.; Jin, D.E.; Park, S.K.; Park, C.H.; Seung, T.W.; Bae, D.W.; Kim, D.O.; Heo, H.J. Antiamnesic effect of actinidia arguta extract intake in a mouse model of tmt-induced learning and memory dysfunction. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Chung, W.S.; Kim, S.S.; Ko, S.G.; Um, J.Y. Antiinflammatory effect of Oldenlandia diffusa and its constituent, hentriacontane, through suppression of caspase-1 activation in mouse peritoneal macrophages. Phytother. Res. 2011, 25, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.H.; Chen, M.B.; Ji, C.; Li, W.T.; Wei, M.X.; Wu, M.H. Aqueous Oldenlandia diffusa extracts inhibits colorectal cancer cells via activating AMP-activated protein kinase signalings. Oncotarget 2016, 7, 45889–45900. [Google Scholar] [CrossRef] [PubMed]
- Sunwoo, Y.Y.; Lee, J.H.; Jung, H.Y.; Jung, Y.J.; Park, M.S.; Chung, Y.A.; Maeng, L.S.; Han, Y.M.; Shin, H.S.; Lee, J.; et al. Oldenlandia diffusa promotes antiproliferative and apoptotic effects in a rat hepatocellular carcinoma with liver cirrhosis. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Barone, I.; Gelsomino, L.; Giordano, C.; Bonofiglio, D.; Statti, G.; Menichini, F.; Catalano, S.; Ando, S. Oldenlandia diffusa extracts exert antiproliferative and apoptotic effects on human breast cancer cells through ERalpha/Sp1-mediated p53 activation. J. Cell. Physiol. 2012, 227, 3363–3372. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.O.; Ban, J.Y.; Kim, J.Y.; Ju, H.S.; Lee, I.S.; Song, K.S.; Bae, K.; Seong, Y.H. Anti-ischemic activities of aralia cordata and its active component, oleanolic acid. Arch. Pharm. Res. 2009, 32, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.K.; Heo, H.J.; Kim, E.K.; Kim, H.K.; Huh, T.L.; Lim, Y.; Kim, S.K.; Shin, D.H. Inhibitory effect of ursolic acid purified from Origanum majorana L. on the acetylcholinesterase. Mol. Cells 2001, 11, 137–143. [Google Scholar] [PubMed]
- Kim, M.Y.; Kim, K.N.; Cho, H.M.; Lee, D.J.; Cho, D.Y. Reference intervals for plasma amyloid β in korean adults without cognitive impairment. Ann. Lab. Med. 2016, 36, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Risner, M.E.; Saunders, A.M.; Altman, J.F.; Ormandy, G.C.; Craft, S.; Foley, I.M.; Zvartau-Hind, M.E.; Hosford, D.A.; Roses, A.D. Efficacy of rosiglitazone in a genetically defined population with mild-to-moderate Alzheimer’s disease. Pharmacogenom. J. 2006, 6, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Boublay, N.; Schott, A.M.; Krolak-Salmon, P. Neuroimaging correlates of neuropsychiatric symptoms in Alzheimer’s disease: A review of 20 years of research. Eur. J. Neurol. 2016, 23, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.M.; Yeon, B.K.; Cho, S.J.; Suh, Y.H. Stem cell therapy for Alzheimer’s disease: A review of recent clinical trials. J. Alzheimers Dis. 2016, 54, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.M.; Takahashi, P.Y.; Yu Ballard, A.C. Active vaccines for Alzheimer disease treatment. J. Am. Med. Dir. Assoc. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Galimberti, D.; Scarpini, E. Old and new acetylcholinesterase inhibitors for Alzheimer’s disease. Expert Opin. Investig. Drugs 2016, 25, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xue, W.; Chu, S.F.; Wang, Z.Z.; Li, C.J.; Jiang, Y.N.; Luo, L.M.; Luo, P.; Li, G.; Zhang, D.M.; et al. Polygalasaponin XXXII, a triterpenoid saponin from polygalae radix, attenuates scopolamine-induced cognitive impairments in mice. Acta Pharmacol. Sin. 2016, 37, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Dabaghian, F.; Khademian, S.; Azadi, A.; Zarshenas, M. Fingerprints, pharmaceutical and radical scavenging activity evaluation of an Alzheimer-targeted herbal preparation. Iran. J. Med. Sci. 2016, 41, S6. [Google Scholar] [PubMed]
- Szczepanik, J.; Nugent, A.C.; Drevets, W.C.; Khanna, A.; Zarate, C.A., Jr.; Furey, M.L. Amygdala response to explicit sad face stimuli at baseline predicts antidepressant treatment response to scopolamine in major depressive disorder. Psychiatry Res. 2016, 254, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Tabari, S.S.; Babri, S.; Mirzaie, F.; Farajdokht, F.; Mohaddes, G. Enduring amnesia induced by ICV scopolamine is reversed by sesame oil in male rats. Acta Cir. Bras. 2016, 31, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Jiang, Z.; Fong, D.W.; Zhao, Z. Determination of oleanolic acid and ursolic acid in Oldenlandia diffusa and its substitute using high performance liquid chromatography. J. Food Drug Anal. 2009, 17, 69–77. [Google Scholar]
- Lu, J.; Wu, D.M.; Zheng, Y.L.; Hu, B.; Cheng, W.; Zhang, Z.F.; Shan, Q. Ursolic acid improves high fat diet-induced cognitive impairments by blocking endoplasmic reticulum stress and IκB kinase β /nuclear factor-κB-mediated inflammatory pathways in mice. Brain Behav. Immun. 2011, 25, 1658–1667. [Google Scholar] [CrossRef] [PubMed]
- Leal, G.; Comprido, D.; Duarte, C.B. BDNF-induced local protein synthesis and synaptic plasticity. Neuropharmacology 2014, 76, 639–656. [Google Scholar] [CrossRef] [PubMed]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef]
- Kwon, O.S.; Jeong, M.S.; Kim, B.; Kim, S.H. Antiangiogenic effect of ethanol extract of vigna angularis via inhibition of phosphorylation of VEGFR2, Erk, and Akt. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Woo, M.J.; Park, C.S.; Lee, S.H.; Kim, J.S.; Kim, B.; An, S.; Kim, S.H. Hovenia dulcis extract reduces lipid accumulation in oleic acid-induced steatosis of hep G2 cells via activation of AMPK and PPARalpha/CPT-1 pathway and in acute hyperlipidemia mouse model. Phytother. Res. 2017, 31, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Qizilbash, N.; Birks, J.; Lopez Arrieta, J.; Lewington, S.; Szeto, S. Tacrine for Alzheimer’s disease. Cochrane Database Syst. Rev. 2000. [Google Scholar] [CrossRef]
- Wilkinson, K.; Boyd, J.D.; Glicksman, M.; Moore, K.J.; El Khoury, J. A high content drug screen identifies ursolic acid as an inhibitor of amyloid β protein interactions with its receptor CD36. J. Biol. Chem. 2011, 286, 34914–34922. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Van der Zee, E.A.; Biemans, B.A.; Gerkema, M.P.; Daan, S. Habituation to a test apparatus during associative learning is sufficient to enhance muscarinic acetylcholine receptor-immunoreactivity in rat suprachiasmatic nucleus. J. Neurosci. Res. 2004, 78, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Hestrin, S. The reaction of acetylcholine and other carboxylic acid derivatives with hydroxylamine, and its analytical application. J. Biol. Chem. 1949, 180, 249–261. [Google Scholar] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Group | Dose (mg/kg) | Design | Number | |
|---|---|---|---|---|---|
| 1 | Normal | Saline | 10 | ||
| 2 | Control | Scopolamine 3 mg/kg i.p. 1 | 10 | ||
| 3 | Positive control | 2 | tacrine. | 10 | |
| 4 | Positive control | 2 | Ursolic acid | 10 | |
| 5 | ODH | 100 | O100 | 10 | |
| 6 | 200 | O200 | 10 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.E.; Song, H.-S.; Park, M.N.; Kim, S.-H.; Shim, B.-S.; Kim, B. Ethanol Extract of Oldenlandia diffusa Herba Attenuates Scopolamine-Induced Cognitive Impairments in Mice via Activation of BDNF, P-CREB and Inhibition of Acetylcholinesterase. Int. J. Mol. Sci. 2018, 19, 363. https://doi.org/10.3390/ijms19020363
Lee JE, Song H-S, Park MN, Kim S-H, Shim B-S, Kim B. Ethanol Extract of Oldenlandia diffusa Herba Attenuates Scopolamine-Induced Cognitive Impairments in Mice via Activation of BDNF, P-CREB and Inhibition of Acetylcholinesterase. International Journal of Molecular Sciences. 2018; 19(2):363. https://doi.org/10.3390/ijms19020363
Chicago/Turabian StyleLee, Jung Eun, Hyo-Sook Song, Moon Nyeo Park, Sung-Hoon Kim, Bum-Sang Shim, and Bonglee Kim. 2018. "Ethanol Extract of Oldenlandia diffusa Herba Attenuates Scopolamine-Induced Cognitive Impairments in Mice via Activation of BDNF, P-CREB and Inhibition of Acetylcholinesterase" International Journal of Molecular Sciences 19, no. 2: 363. https://doi.org/10.3390/ijms19020363