Anti-Cancerous Effect of Inonotus taiwanensis Polysaccharide Extract on Human Acute Monocytic Leukemia Cells through ROS-Independent Intrinsic Mitochondrial Pathway

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
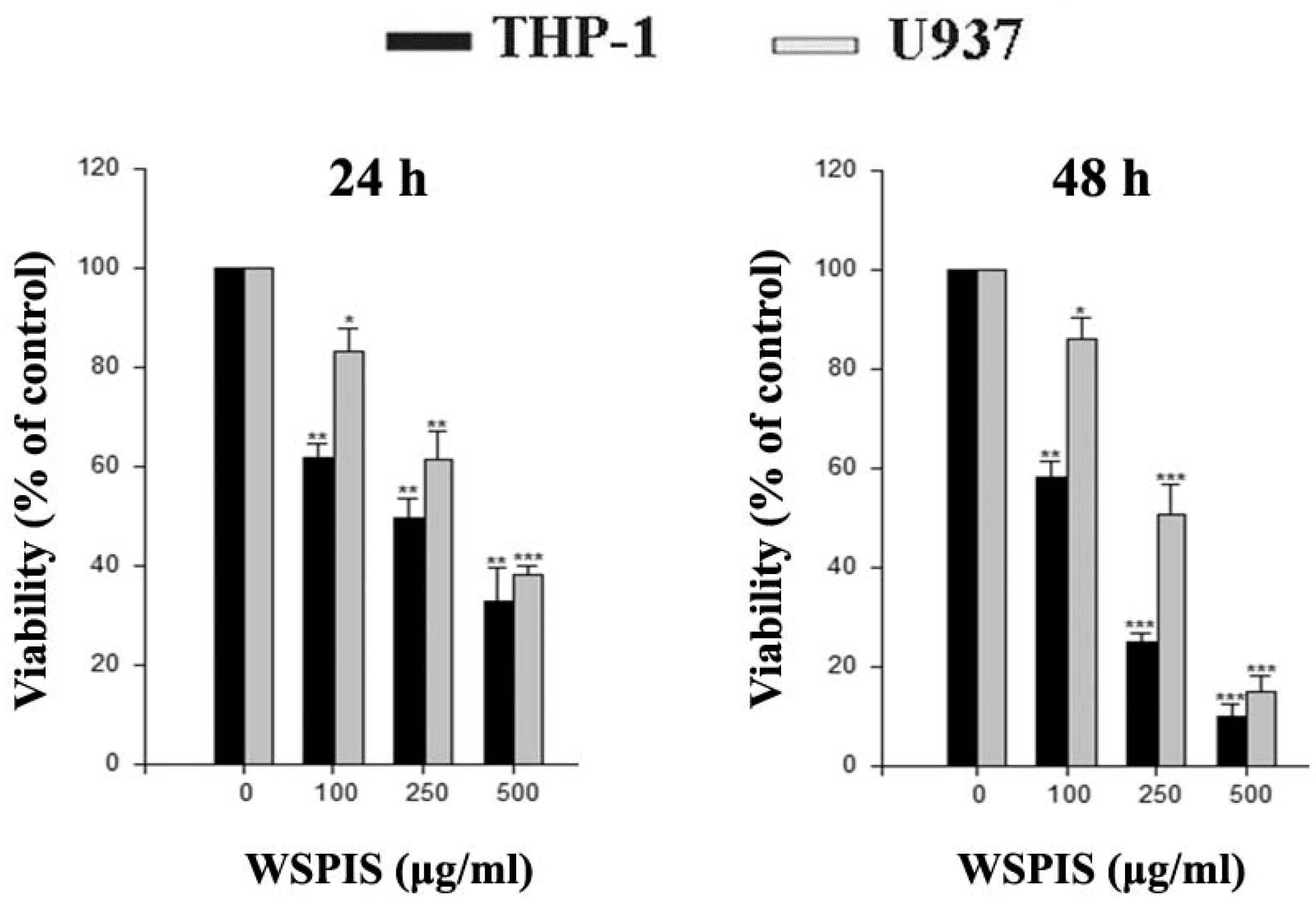
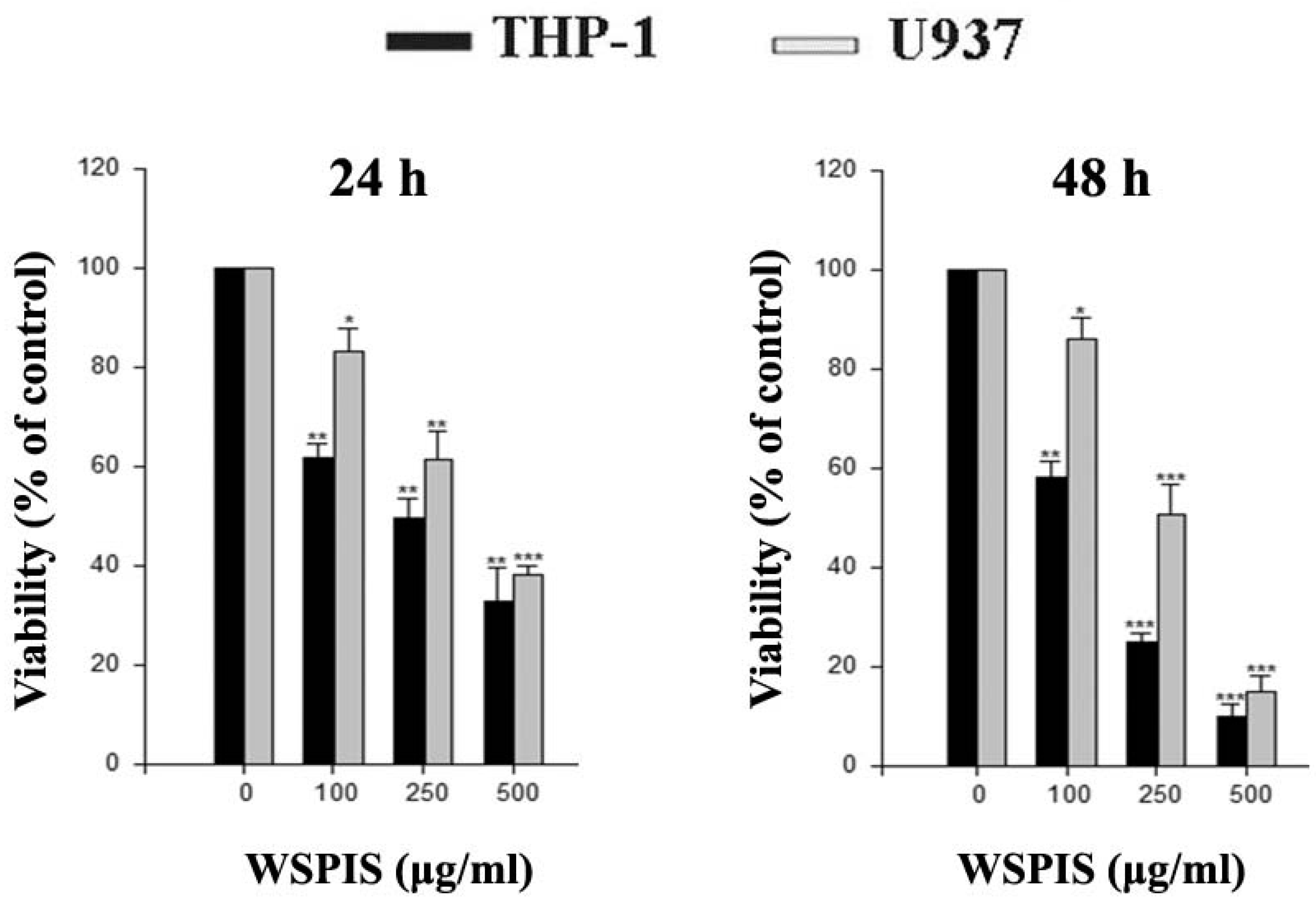
2.1. Anti-Proliferative Activity of WSPIS in Human Monocytic Leukemia Cells
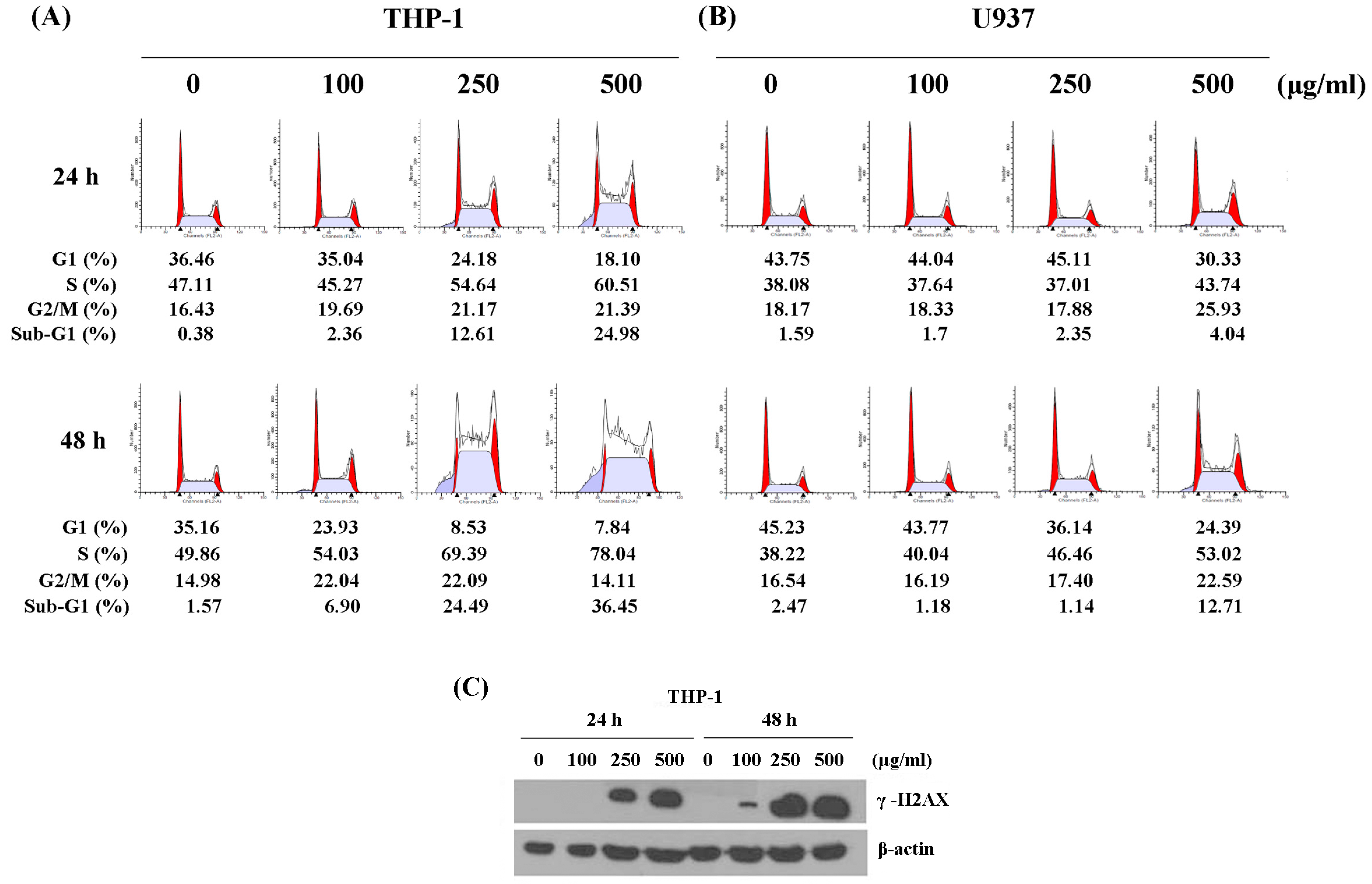
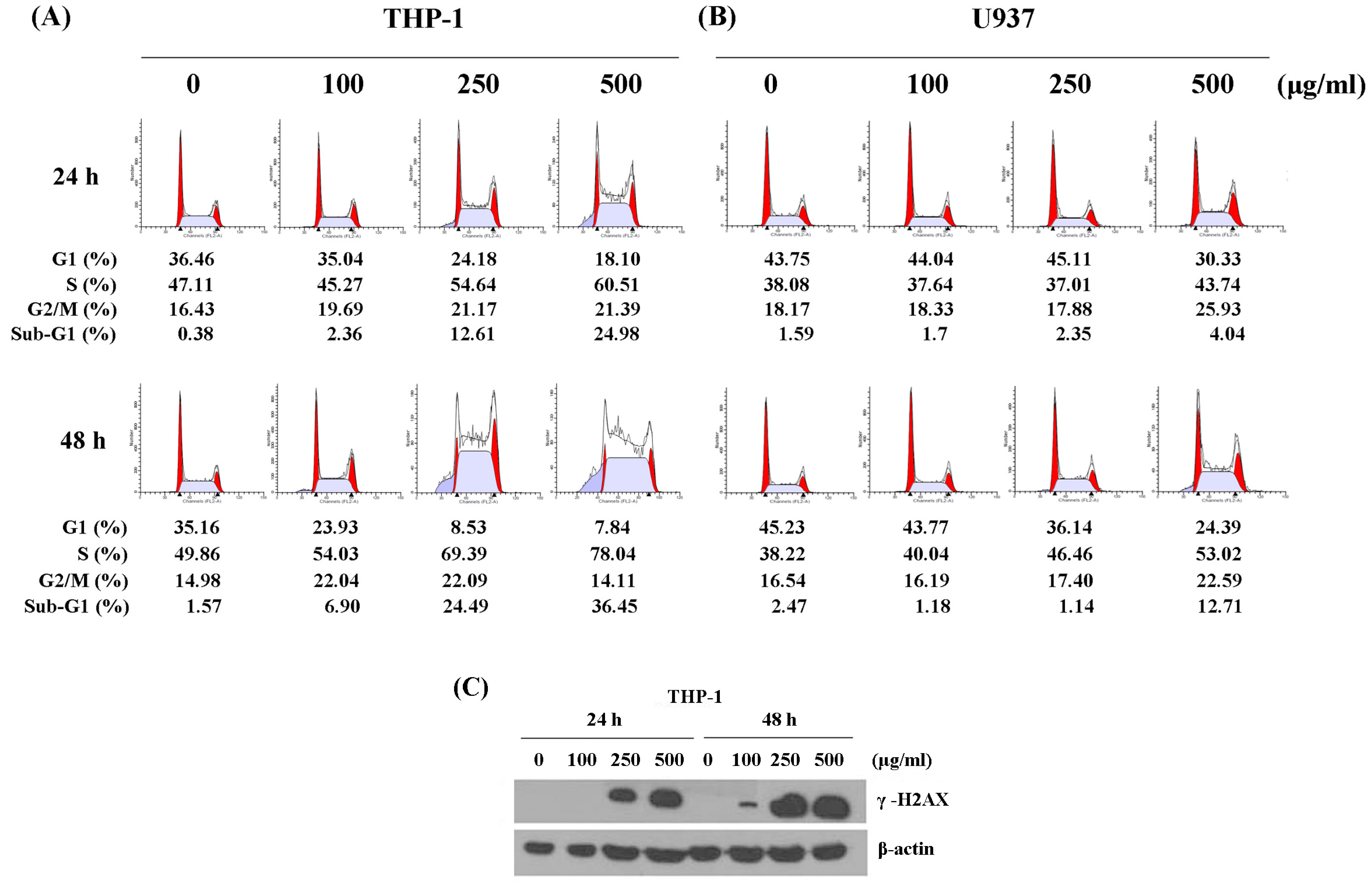
2.2. WSPIS Caused DNA Damage and Delayed Cell Cycle Progression
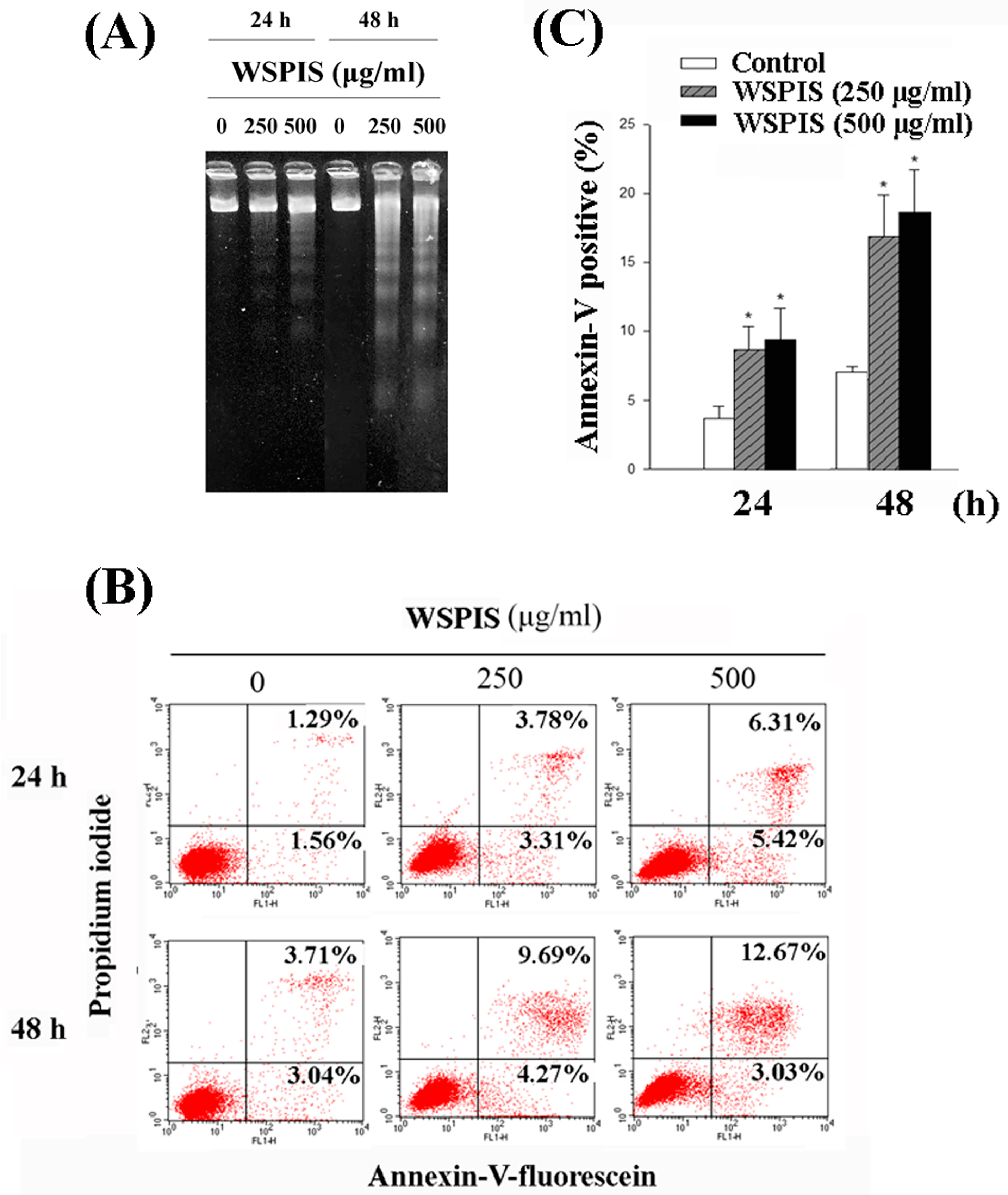
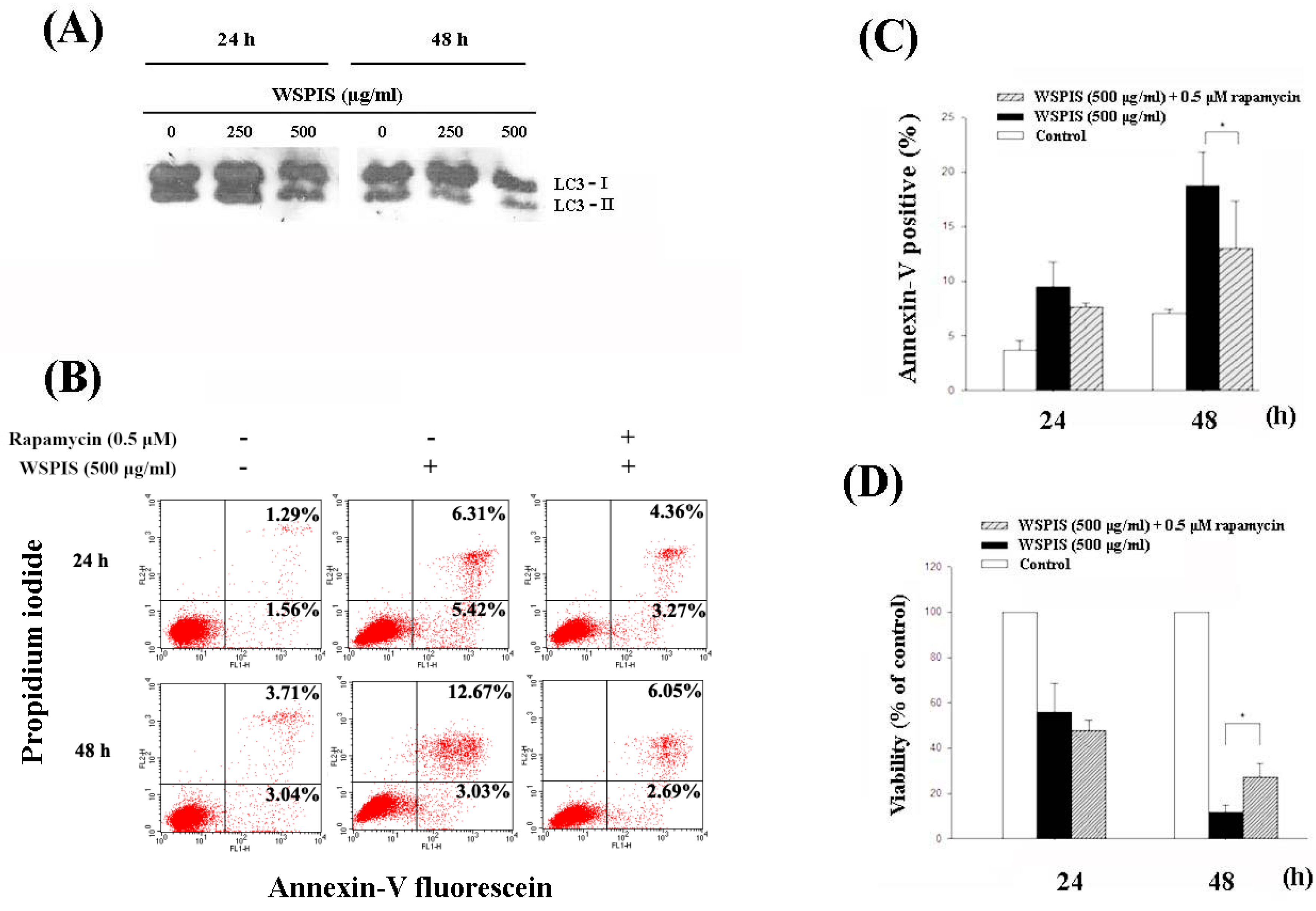
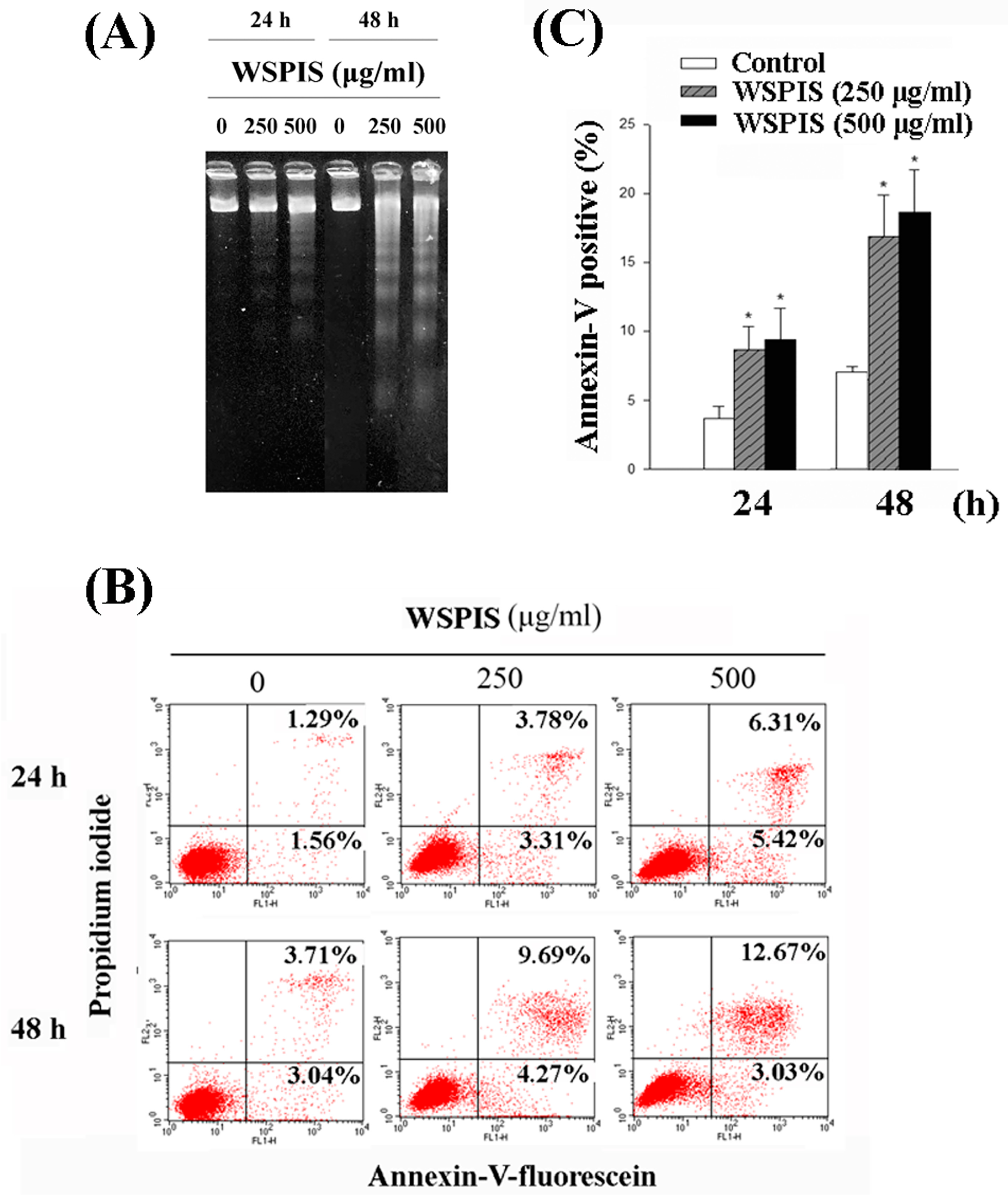
2.3. WSPIS Induced Apoptosis in THP-1 Cells
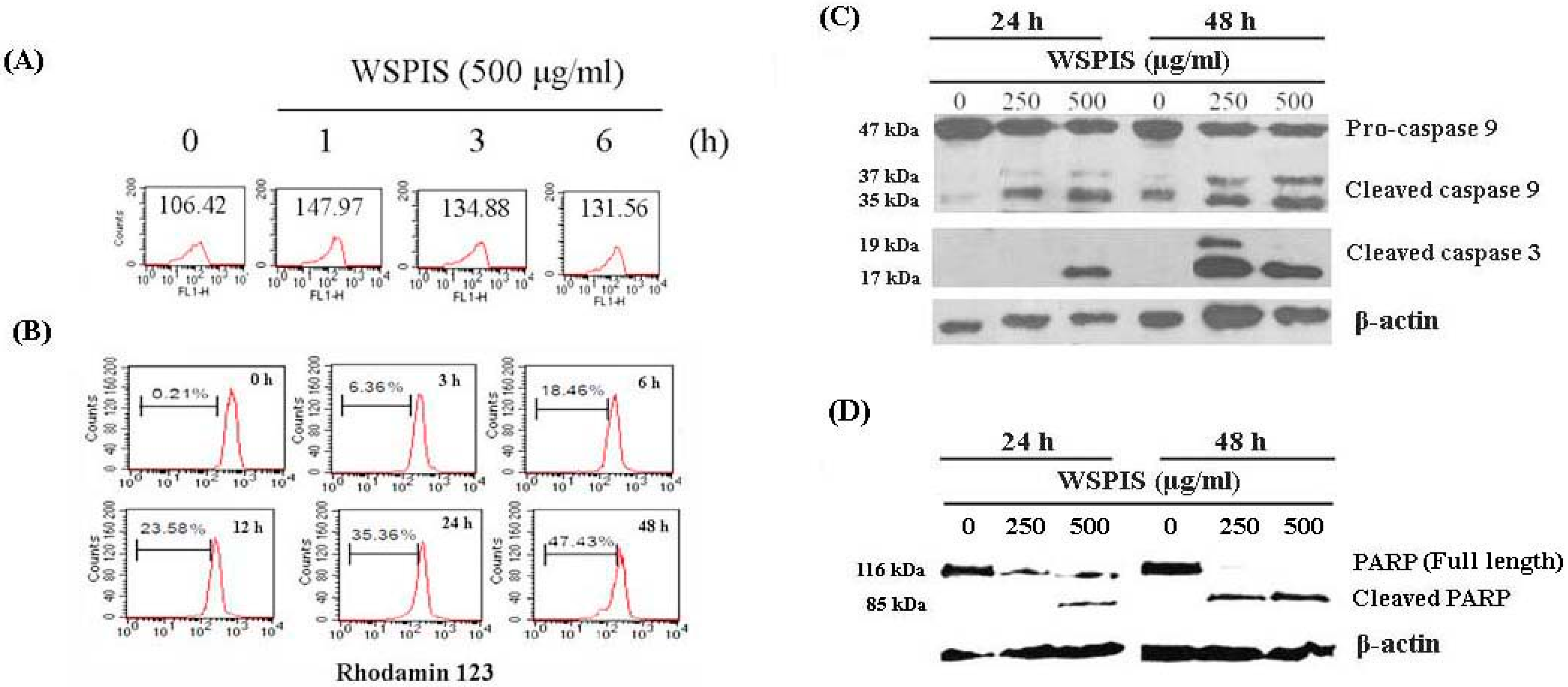
2.4. WSPIS Induced Reactive Oxygen Species (ROS) Production and Activated Mitochondrial Apoptotic Pathway
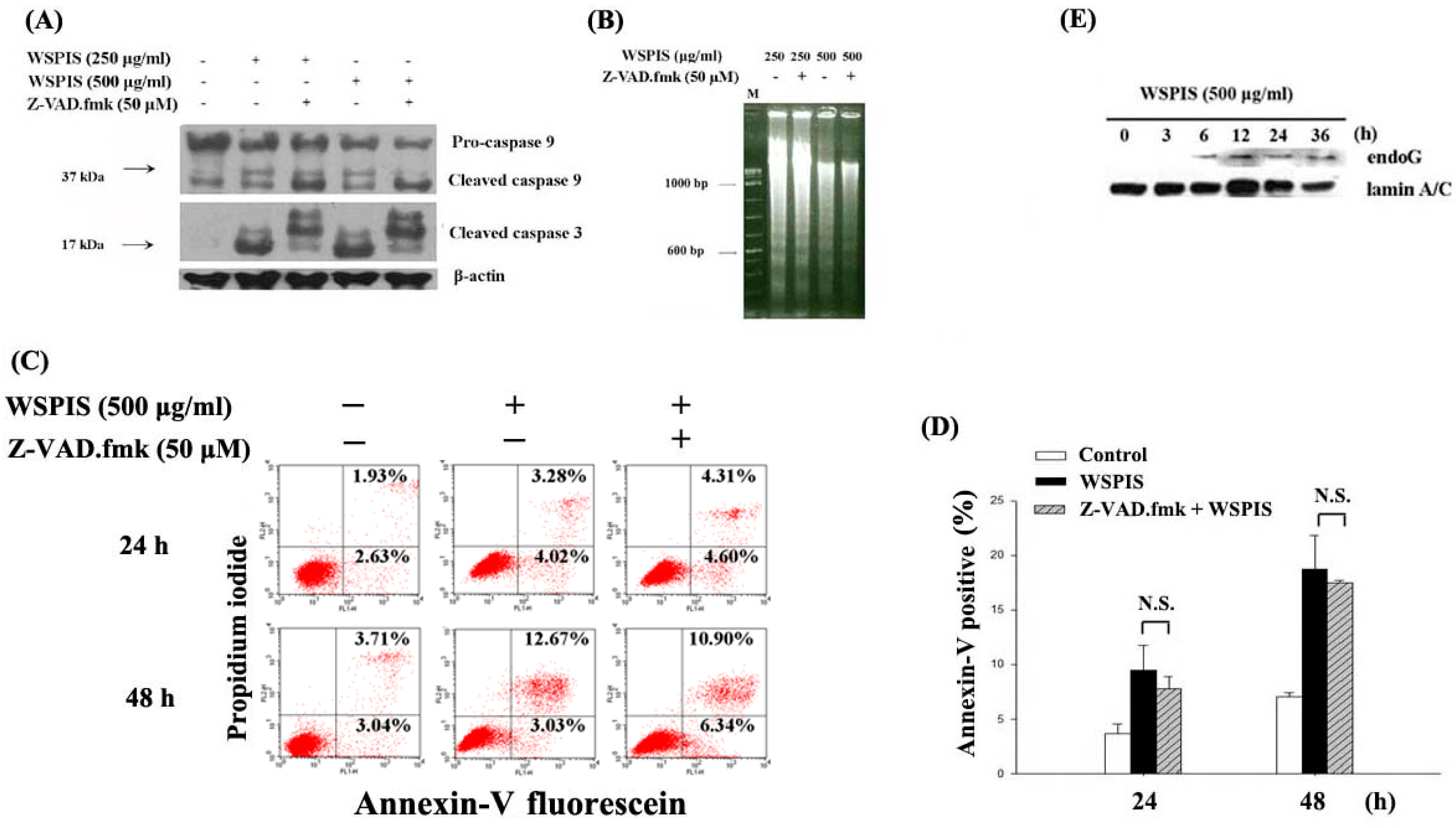
2.5. WSPIS Induced Caspase-Independent but EndoG-Mediated Apoptosis
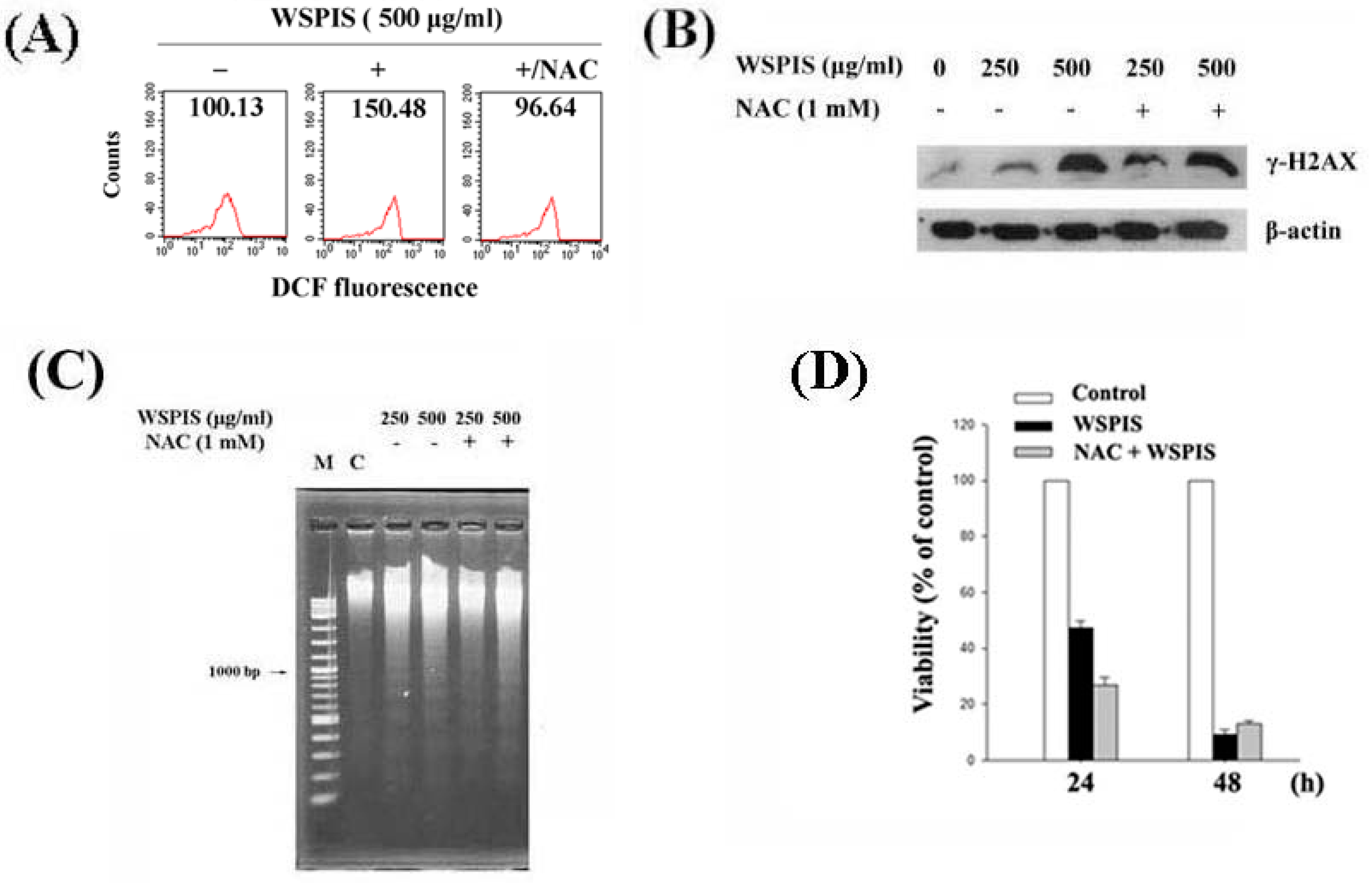
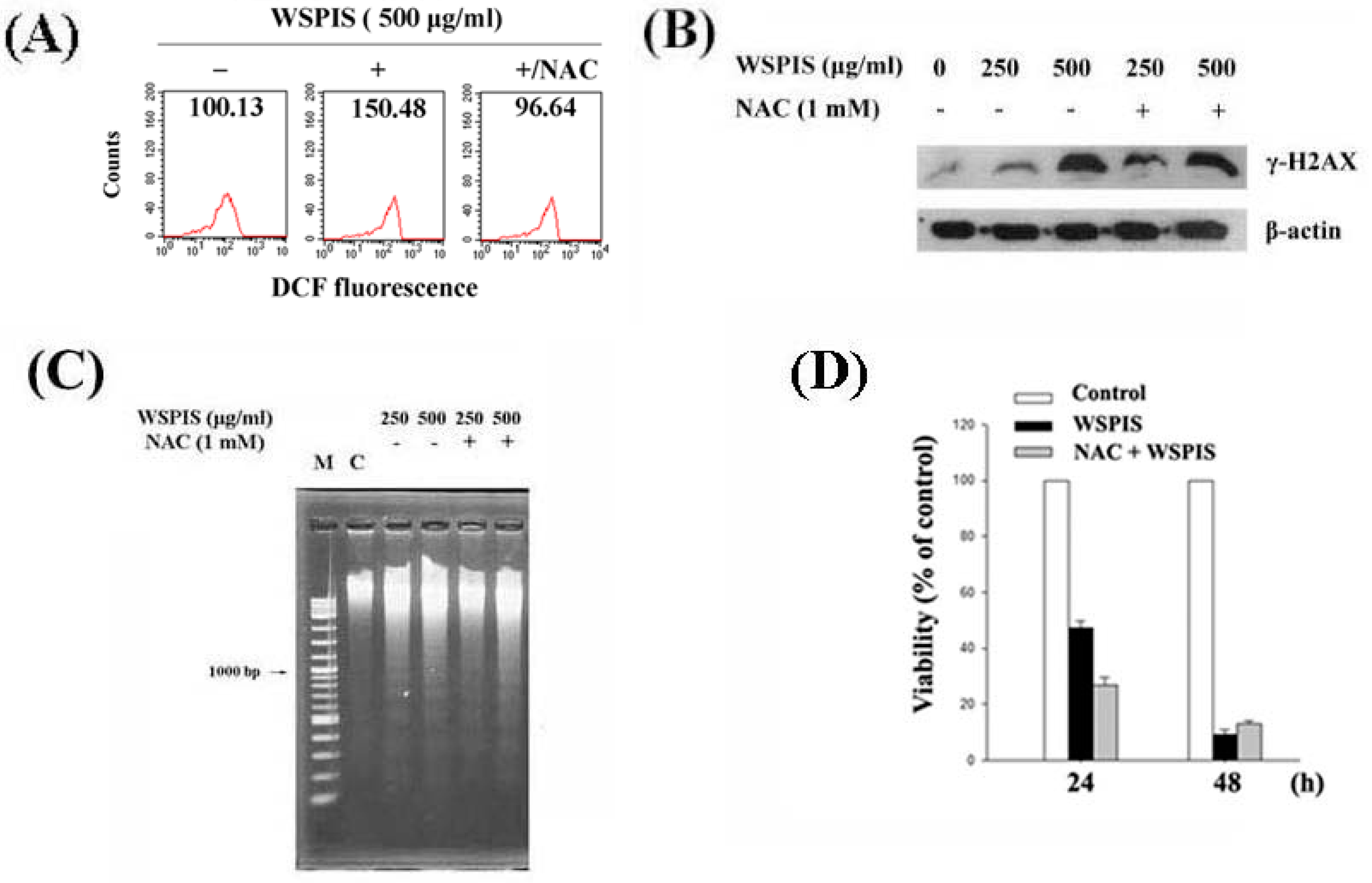
2.6. WSPIS-Induced Apoptosis through the ROS-Independent Pathway
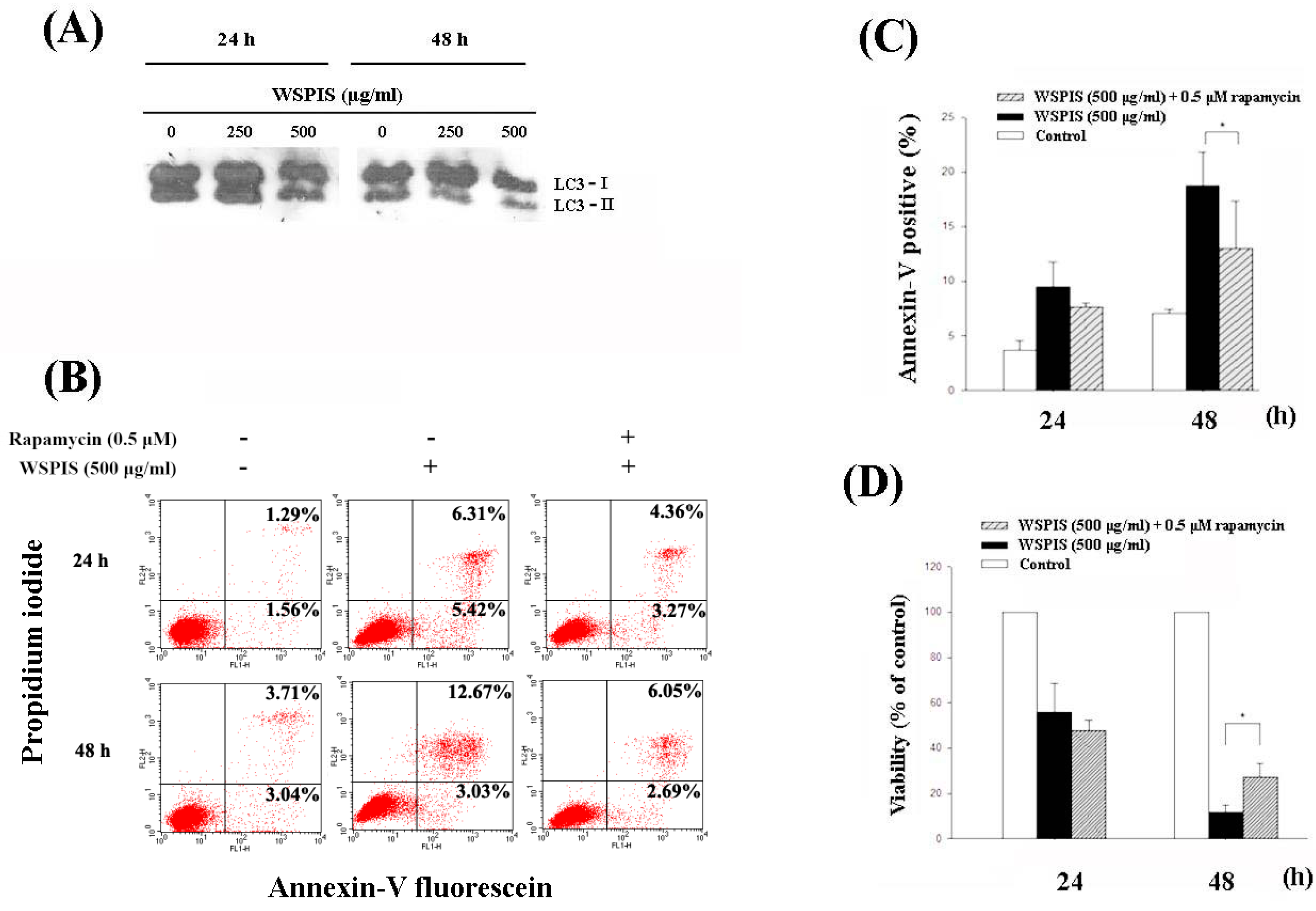
2.7. WSPIS Inhibited Autophagy in THP-1 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. WSPIS Polysaccharide Extract Preparation and Total Carbohydrate Measurement
4.3. Cell Culture and Treatment with WSPIS
4.4. Cell Viability Assay
4.5. Flow Cytometry Analysis of DNA Content
4.6. Agarose Gel Electrophoresis of DNA Fragmentation
4.7. Annexin-V/Propidium Iodide (PI) Staining
4.8. Measurement of Mitochondrial Membrane Potential (MMP) by Flow Cytometry
4.9. Cell Lysate Preparation and Immunoblotting
4.10. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deschler, B.; Lubbert, M. Acute myeloid leukemia: Epidemiology and etiology. Cancer 2006, 107, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Micallef, I.N.; Rohatiner, A.Z.; Carter, M.; Boyle, M.; Slater, S.; Amess, J.A.; Lister, T.A. Long-term outcome of patients surviving for more than ten years following treatment for acute leukaemia. Br. J. Haematol. 2001, 113, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Debatin, K.M.; Poncet, D.; Kroemer, G. Chemotherapy: Targeting the mitochondrial cell death pathway. Oncogene 2002, 21, 8786–8803. [Google Scholar] [CrossRef] [PubMed]
- Susin, S.A.; Daugas, E.; Ravagnan, L.; Samejima, K.; Zamzami, N.; Loeffler, M.; Costantini, P.; Ferri, K.F.; Irinopoulou, T.; Prévost, M.C.; et al. Two distinct pathways leading to nuclear apoptosis. J. Exp. Med. 2000, 192, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Shimamoto, N. Involvement of endonuclease G in nucleosomal DNA fragmentation under sustained endogenous oxidative stress. J. Biol. Chem. 2006, 281, 6726–6733. [Google Scholar] [CrossRef] [PubMed]
- Chihara, G.; Maeda, Y.; Hamuro, J.; Sasaki, T.; Fukuoka, F. Inhibition of mouse sarcoma 180 by polysaccharides from Lentinus edodes (Berk.) sing. Nature 1969, 222, 687–688. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Yoshida, R.; Kanada, Y.; Fukuda, Y.; Yagyu, T.; Inagaki, K.; Kondo, T.; Kurita, N.; Suzuki, M.; Kanayama, N.; et al. Suppressing effects of daily oral supplementation of beta-glucan extracted from Agaricus blazei Murill on spontaneous and peritoneal disseminated metastasis in mouse model. J. Cancer Res. Clin. Oncol. 2005, 131, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, H.; Koike, A.; Saji, S.; Ogawa, N.; Sakamoto, J. Efficacy of immunechemotherapy as adjuvant treatment after curative resection of gastric cancer. Study group of immuno-chemotherapy with PSK for gastric cancer. Lancet 1994, 343, 1122–1126. [Google Scholar] [CrossRef]
- Shin, J.Y.; Song, J.Y.; Yun, Y.S.; Yang, H.O.; Rhee, D.K.; Pyo, S. Immunostimulating effects of acidic polysaccharides extract of Panax ginseng on macrophage function. Immunopharmacol. Immunotoxicol. 2002, 24, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, L.; Krizan, J.; Sima, P.; Stakheev, D.; Caja, F.; Rajsiglova, L.; Horak, V.; Saieh, M. Immunostimulatory properties and antitumor activities of glucans. Int. J. Oncol. 2013, 43, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Lin, Y.T.; Chern, C.L.; Ke, S.T. Inonotus taiwanensis sp. nov. (Basidiomycota) from Taiwan. Mycoscience 2017. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX phosphorylation: A marker for DNA damage. Methods Mol. Biol. 2012, 920, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, B.; Moreau, K.; Jahreiss, L.; Puri, C.; Rubinsztein, D.C. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nat. Cell Biol. 2010, 12, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Ralph, S.J.; Rodríguez-Enríquez, S.; Neuzil, J.; Saavedra, E.; Moreno-Sánchez, R. The causes of cancer revisited: “mitochondrial malignancy” and ROS-induced oncogenic transformation–why mitochondria are targets for cancer therapy. Mol. Aspects Med. 2010, 31, 145–170. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.; Grieco, D. New insights on oxidative stress in cancer. Curr. Opin. Drug. Discov. Dev. 2009, 12, 240–245. [Google Scholar]
- Amaravadi, R.K.; Lippincott-Schwartz, J.; Yin, X.M.; Weiss, W.A.; Takebe, N.; Timmer, W.; DiPaola, R.S.; Lotze, M.T.; White, E. Principles and current strategies for targeting autophagy for cancer treatment. Clin. Cancer Res. 2011, 17, 654–666. [Google Scholar] [CrossRef] [PubMed]
- White, E.; DiPaola, R.S. The double-edged sword of autophagy modulation in cancer. Clin. Cancer Res. 2009, 15, 5308–5316. [Google Scholar] [CrossRef] [PubMed]
- Carew, J.S.; Kelly, K.R.; Nawrocki, S.T. Autophagy as a target for cancer therapy: New developments. Cancer Manag. Res. 2012, 4, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Z.; Cheng, J.T.; Yiin, S.J.; Chen, C.Y.; Chen, C.H.; Wu, M.J.; Chern, C.L. Isoobtusilactone A induces both caspase-dependent and -independent apoptosis in Hep G2 cells. Food Chem. Toxicol. 2008, 46, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Tai, C.J.; Cheng, J.T.; Zheng, J.J.; Chen, Y.Z.; Liu, T.Z.; Yiin, S.J.; Chern, C.L. 6-Dehydrogingerdione sensitizes human hepatoblastoma Hep G2 cells to TRAIL-induced apoptosis via reactive oxygen species-mediated increase of DR5. J. Agric. Food Chem. 2010, 58, 5604–5611. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, T.-L.; Wang, T.-Y.; Lee, C.-H.; Yiin, S.-J.; Ho, C.-T.; Wu, S.-H.; You, H.-L.; Chern, C.-L. Anti-Cancerous Effect of Inonotus taiwanensis Polysaccharide Extract on Human Acute Monocytic Leukemia Cells through ROS-Independent Intrinsic Mitochondrial Pathway. Int. J. Mol. Sci. 2018, 19, 393. https://doi.org/10.3390/ijms19020393
Chao T-L, Wang T-Y, Lee C-H, Yiin S-J, Ho C-T, Wu S-H, You H-L, Chern C-L. Anti-Cancerous Effect of Inonotus taiwanensis Polysaccharide Extract on Human Acute Monocytic Leukemia Cells through ROS-Independent Intrinsic Mitochondrial Pathway. International Journal of Molecular Sciences. 2018; 19(2):393. https://doi.org/10.3390/ijms19020393
Chicago/Turabian StyleChao, Tsai-Ling, Ting-Yin Wang, Chin-Huei Lee, Shuenn-Jiun Yiin, Chun-Te Ho, Sheng-Hua Wu, Huey-Ling You, and Chi-Liang Chern. 2018. "Anti-Cancerous Effect of Inonotus taiwanensis Polysaccharide Extract on Human Acute Monocytic Leukemia Cells through ROS-Independent Intrinsic Mitochondrial Pathway" International Journal of Molecular Sciences 19, no. 2: 393. https://doi.org/10.3390/ijms19020393