Secreted and Tissue miRNAs as Diagnosis Biomarkers of Malignant Pleural Mesothelioma

Abstract
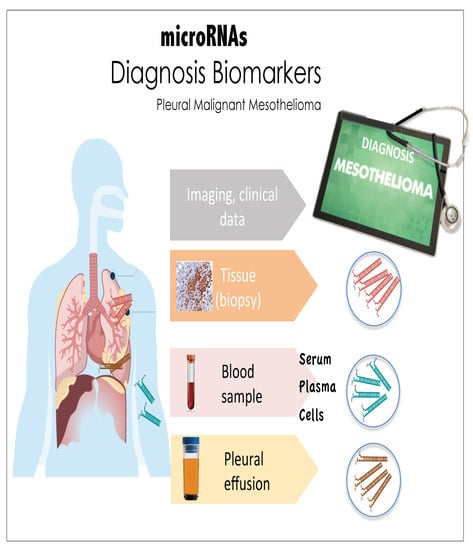
:
1. Introduction
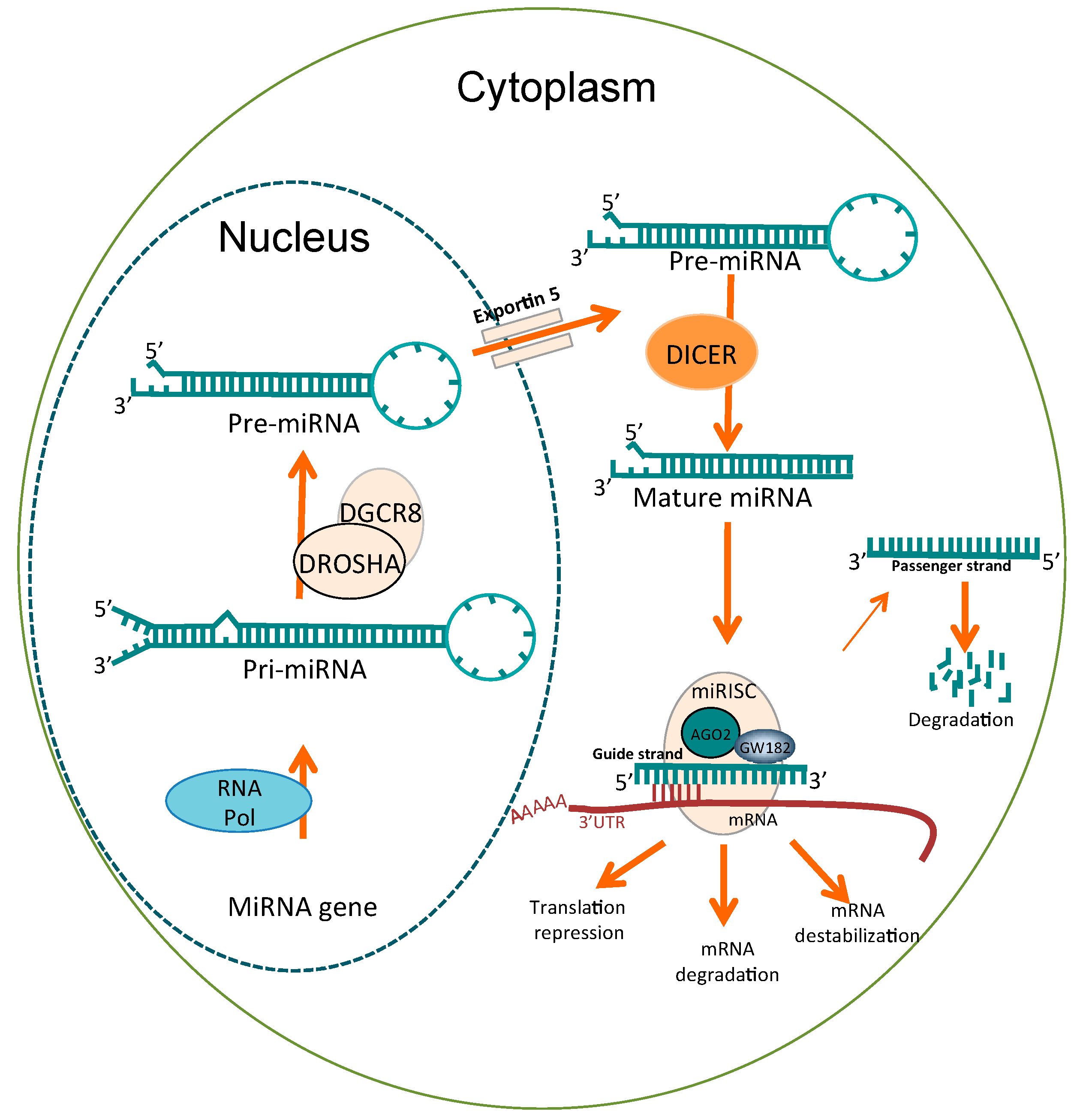
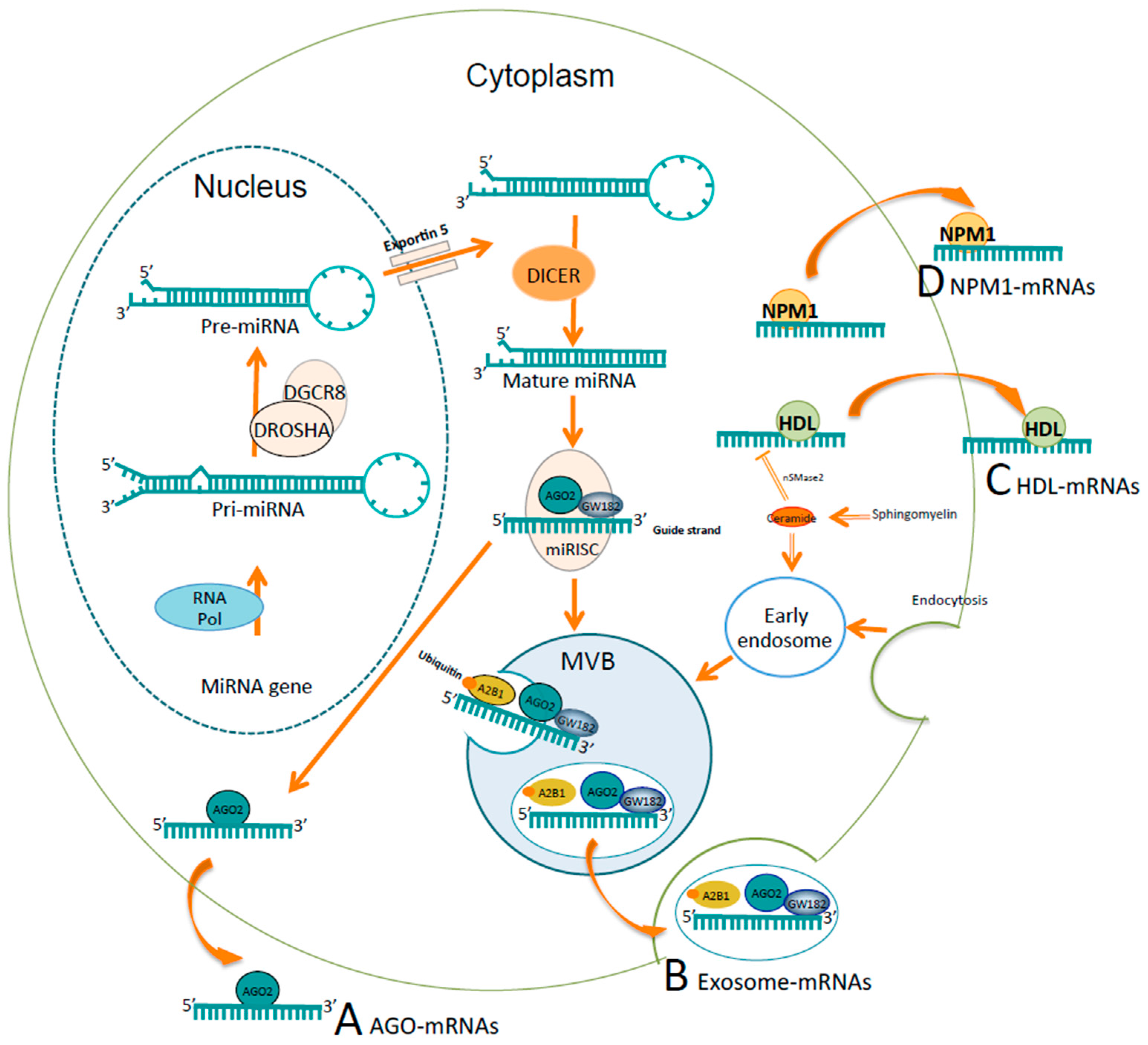
2. MicroRNAs
3. Studies Regarding microRNAs as Biomarkers for MPM
3.1. Studies in Pleural Tissue
3.2. Studies in the Cellular Fraction of Peripheral Blood
3.3. Studies in Serum and Plasma
3.4. Studies in Other Body Fluids
4. Relevant Aspects of the Experimental Designs That May Influence the Accuracy, Consistency and Real Diagnostic Value of Currently Reported Data
4.1. Number of Samples
4.2. Follow up Studies of Promising Candidate miRNAs Biomarkers
4.3. Relevance of Tumor Representativeness in Tissue Samples
4.4. Relevance of the Normal/Healthy Controls Used in These Studies
4.5. Relevance of Proper Normalizers for Quantitative RT-qPCR Analysis (Validation Phase)
4.6. Analysis of Different Histological Subtypes of MPM
4.7. More Than One Biomarker Used for Diagnosis
4.8. Complete and Accurate Reporting
5. miRNAs Associated with Neoplastic Mechanisms of MPM and Their Potential Diagnostic Value
6. Conclusions and Future Perspectives
Author Contributions
Conflicts of Interest
References
- Kaufman, A.J.; Pass, H.I. Current concepts in malignant pleural mesothelioma. Expert Rev. Anticancer Ther. 2008, 8, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.W.; Lake, R.A. Advances in malignant mesothelioma. N. Engl. J. Med. 2005, 353, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, C.; Bianchi, T. Malignant mesothelioma: Global incidence and relationship with asbestos. Ind. Health 2007, 45, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Roggli, V.L.; Sharma, A.; Butnor, K.J.; Sporn, T.; Vollmer, R.T. Malignant mesothelioma and occupational exposure to asbestos: A clinicopathological correlation of 1445 cases. Ultrastruct. Pathol. 2002, 26, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.; Kindler, H.L. Malignant pleural mesothelioma: An update on biomarkers and treatment. Chest 2009, 136, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Delgermaa, V.; Takahashi, K.; Park, E.K.; Le, G.V.; Hara, T.; Sorahan, T. Global mesothelioma deaths reported to the world health organization between 1994 and 2008. Bull. World Health Organ. 2011, 89, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Park, E.K.; Takahashi, K.; Hoshuyama, T.; Cheng, T.J.; Delgermaa, V.; Le, G.V.; Sorahan, T. Global magnitude of reported and unreported mesothelioma. Environ. Health Perspect. 2011, 119, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Addis, B.; Roche, H. Problems in mesothelioma diagnosis. Histopathology 2009, 54, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Inai, K. Pathology of mesothelioma. Environ. Health Prev. Med. 2008, 13, 60–64. [Google Scholar] [CrossRef] [PubMed]
- British Thoracic Society Standards of Care Committee. BTS statement on malignant mesothelioma in the UK, 2007. Thorax 2007, 62 (Suppl. 2), ii1–ii19. [Google Scholar]
- Evans, A.L.; Gleeson, F.V. Radiology in pleural disease: State of the art. Respirology 2004, 9, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Hallifax, R.J.; Talwar, A.; Wrightson, J.M.; Edey, A.; Gleeson, F.V. State-of-the-art: Radiological investigation of pleural disease. Respir. Med. 2017, 124, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Paintal, A.; Raparia, K.; Zakowski, M.F.; Nayar, R. The diagnosis of malignant mesothelioma in effusion cytology: A reappraisal and results of a multi-institution survey. Cancer Cytopathol. 2013, 121, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, D. The cytology of malignant mesothelioma. Cytopathology 2000, 11, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Arif, Q.; Husain, A.N. Malignant mesothelioma diagnosis. Arch. Pathol. Lab. Med. 2015, 139, 978–980. [Google Scholar] [CrossRef] [PubMed]
- Heffner, J.E.; Klein, J.S. Recent advances in the diagnosis and management of malignant pleural effusions. Mayo Clin. Proc. 2008, 83, 235–250. [Google Scholar] [CrossRef]
- Husain, A.N.; Colby, T.V.; Ordonez, N.G.; Krausz, T.; Borczuk, A.; Cagle, P.T.; Chirieac, L.R.; Churg, A.; Galateau-Salle, F.; Gibbs, A.R.; et al. Guidelines for pathologic diagnosis of malignant mesothelioma: A consensus statement from the international mesothelioma interest group. Arch. Pathol. Lab. Med. 2009, 133, 1317–1331. [Google Scholar] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Sherrard, R.; Luehr, S.; Holzkamp, H.; McJunkin, K.; Memar, N.; Conradt, B. miRNAs cooperate in apoptosis regulation during c. Elegans development. Genes Dev. 2017, 31, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Shaw, W.R.; Armisen, J.; Lehrbach, N.J.; Miska, E.A. The conserved mir-51 microRNA family is redundantly required for embryonic development and pharynx attachment in caenorhabditis elegans. Genetics 2010, 185, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Koff, J.L.; Ramachandiran, S.; Bernal-Mizrachi, L. A time to kill: Targeting apoptosis in cancer. Int. J. Mol. Sci. 2015, 16, 2942–2955. [Google Scholar] [CrossRef] [PubMed]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Nosho, K.; Ishigami, K.; Yamamoto, I.; Koide, H.; Kurihara, H.; Mitsuhashi, K.; Shitani, M.; Motoya, M.; Sasaki, S.; et al. MicroRNA-196b is an independent prognostic biomarker in patients with pancreatic cancer. Carcinogenesis 2017, 38, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Kong, X.; Wang, H.; Huang, G.; Ye, C.; He, Z. A novel microRNAs expression signature for hepatocellular carcinoma diagnosis and prognosis. Oncotarget 2017, 8, 8775–8784. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Pan, L.; Niu, Z.; Li, W.; Dang, X.; Wan, L.; Zhang, R.; Yang, S. Identification of microRNAs as potential biomarkers for lung adenocarcinoma using integrating genomics analysis. Oncotarget 2017, 8, 64143–64156. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Tanaka, M.; Yoshida, M.; Umakoshi, M.; Nanjo, H.; Shiraishi, K.; Saito, M.; Kohno, T.; Kuriyama, S.; Konno, H.; et al. The low expression of miR-451 predicts a worse prognosis in non-small cell lung cancer cases. PLoS ONE 2017, 12, e0181270. [Google Scholar] [CrossRef] [PubMed]
- Ahmadinejad, F.; Mowla, S.J.; Honardoost, M.A.; Arjenaki, M.G.; Moazeni-Bistgani, M.; Kheiri, S.; Teimori, H. Lower expression of miR-218 in human breast cancer is associated with lymph node metastases, higher grades, and poorer prognosis. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Rosignolo, F.; Memeo, L.; Monzani, F.; Colarossi, C.; Pecce, V.; Verrienti, A.; Durante, C.; Grani, G.; Lamartina, L.; Forte, S.; et al. MicroRNA-based molecular classification of papillary thyroid carcinoma. Int. J. Oncol. 2017, 50, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Santarelli, L.; Strafella, E.; Staffolani, S.; Amati, M.; Emanuelli, M.; Sartini, D.; Pozzi, V.; Carbonari, D.; Bracci, M.; Pignotti, E.; et al. Association of miR-126 with soluble mesothelin-related peptides, a marker for malignant mesothelioma. PLoS ONE 2011, 6, e18232. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Tang, J.; Yang, H.; Cheng, C.; Lu, S.; Jiang, R.; Sun, B. MTA1 modulated by miR-30e contributes to epithelial-to-mesenchymal transition in hepatocellular carcinoma through an ErbB2-dependent pathway. Oncogene 2017, 36, 3976–3985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, T.; Huang, H.; Jiang, Y.; Yang, L.; Lin, Z.; He, H.; Liu, T.; Wu, B.; Chen, J.; et al. miR-363-3p inhibits tumor growth by targeting PCNA in lung adenocarcinoma. Oncotarget 2017, 8, 20133–20144. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, M.; Josson, S.; Chu, G.C.; Lu, C.L.; Lu, Y.T.; Haga, C.L.; Zhau, H.E.; Liu, C.; Lichterman, J.; Duan, P.; et al. miR-154* and miR-379 in the DLK1-DIO3 microRNA mega-cluster regulate epithelial to mesenchymal transition and bone metastasis of prostate cancer. Clin. Cancer Res. 2014, 20, 6559–6569. [Google Scholar] [CrossRef] [PubMed]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.; Au, G.K.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA 2008, 299, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, S.K.; Kannisto, E.; Knudsen, S.; Yendamuri, S. Evaluation of microRNA expression profiles that may predict recurrence of localized stage I non-small cell lung cancer after surgical resection. Cancer Res. 2010, 70, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Quintero, B. Cell-free microRNAs in blood and other body fluids, as cancer biomarkers. Cell Prolif. 2016, 49, 281–303. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Köberle, V.; Pleli, T.; Schmithals, C.; Augusto Alonso, E.; Haupenthal, J.; Bönig, H.; Peveling-Oberhag, J.; Biondi, R.M.; Zeuzem, S.; Kronenberger, B.; et al. Differential stability of cell-free circulating microRNAs: Implications for their utilization as biomarkers. PLoS ONE. 2013, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. microRNA maturation: Stepwise processing and subcellular localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M. Circulating microRNAs as biomarkers and mediators of cell–cell communication in cancer. Biomedicines 2015, 3, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yi, R.; Cullen, B.R. microRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms. Proc. Natl. Acad. Sci. USA 2003, 100, 9779–9784. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yanez, A.; Novina, C.D. microRNA-repressed mRNAs contain 40 s but not 60 s components. Proc. Natl. Acad. Sci. USA 2008, 105, 5343–5348. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; Van Dongen, S.; Inoue, K.; Enright, A.J.; Schier, A.F. Zebrafish mir-430 promotes deadenylation and clearance of maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Raiborg, C.; Stenmark, H. The escrt machinery in endosomal sorting of ubiquitylated membrane proteins. Nature 2009, 458, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnrnpa2b1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral sphingomyelinase 2 (nsmase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J. Biol. Chem. 2013, 288, 10849–10859. [Google Scholar] [CrossRef] [PubMed]
- Gibbings, D.J.; Ciaudo, C.; Erhardt, M.; Voinnet, O. Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity. Nat. Cell Biol. 2009, 11, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Wurdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, H.; Kosaka, N.; Ochiya, T. Secretory microRNAs as a versatile communication tool. Commun. Integr. Biol. 2010, 3, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. microRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Riwanto, M.; Besler, C.; Knau, A.; Fichtlscherer, S.; Roxe, T.; Zeiher, A.M.; Landmesser, U.; Dimmeler, S. Characterization of levels and cellular transfer of circulating lipoprotein-bound microRNAs. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. Hdl-transferred microRNA-223 regulates icam-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [PubMed]
- Avila-Moreno, F.; Urrea, F.; Ortiz-Quintero, B. microRNAs in diagnosis and prognosis in lung cancer. Rev. Investig. Clin. Organo Hosp. Enferm. Nutr. 2011, 63, 516–535. [Google Scholar]
- Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95.
- Guled, M.; Lahti, L.; Lindholm, P.M.; Salmenkivi, K.; Bagwan, I.; Nicholson, A.G.; Knuutila, S. Cdkn2a, nf2, and jun are dysregulated among other genes by miRNAs in malignant mesothelioma -a miRNA microarray analysis. Genes Chromosom. Cancer 2009, 48, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Gee, G.V.; Koestler, D.C.; Christensen, B.C.; Sugarbaker, D.J.; Ugolini, D.; Ivaldi, G.P.; Resnick, M.B.; Houseman, E.A.; Kelsey, K.T.; Marsit, C.J. Downregulated microRNAs in the differential diagnosis of malignant pleural mesothelioma. Int. J. Cancer 2010, 127, 2859–2869. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, H.; Lebanony, D.; Rosenwald, S.; Cohen, L.; Gibori, H.; Barabash, N.; Ashkenazi, K.; Goren, E.; Meiri, E.; Morgenstern, S.; et al. A diagnostic assay based on microRNA expression accurately identifies malignant pleural mesothelioma. J. Mol. Diagn. 2010, 12, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Nakajima, G.; Gavin, E.; Morris, C.G.; Kudo, K.; Hayashi, K.; Ju, J. Systematic analysis of microRNA expression of RNA extracted from fresh frozen and formalin-fixed paraffin-embedded samples. RNA 2007, 13, 1668–1674. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zheng, M.; Merritt, R.E.; Shrager, J.B.; Wakelee, H.A.; Kratzke, R.A.; Hoang, C.D. Mir-1 induces growth arrest and apoptosis in malignant mesothelioma. Chest 2013, 144, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Pel, M.E.; Kirschner, M.B.; Cheng, Y.Y.; Mugridge, N.; Weiss, J.; Williams, M.; Wright, C.; Edelman, J.J.; Vallely, M.P.; et al. Restoring expression of mir-16: A novel approach to therapy for malignant pleural mesothelioma. Ann. Oncol. 2013, 24, 3128–3135. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Ganci, F.; Canu, V.; Sacconi, A.; Mori, F.; Canino, C.; Korita, E.; Casini, B.; Alessandrini, G.; Cambria, A.; et al. Protumorigenic effects of mir-145 loss in malignant pleural mesothelioma. Oncogene 2014, 33, 5319–5331. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Salazar, E.G.; Salinas-Silva, L.C.; Vazquez-Manriquez, M.E.; Gayosso-Gomez, L.V.; Negrete-Garcia, M.C.; Ramirez-Rodriguez, S.L.; Chavez, R.; Zenteno, E.; Santillan, P.; Kelly-Garcia, J.; et al. Analysis of microRNA expression signatures in malignant pleural mesothelioma, pleural inflammation, and atypical mesothelial hyperplasia reveals common predictive tumorigenesis-related targets. Exp. Mol. Pathol. 2014, 97, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.; Grauslund, M.; Ravn, J.; Sorensen, J.B.; Andersen, C.B.; Santoni-Rugiu, E. Diagnostic potential of mir-126, mir-143, mir-145, and mir-652 in malignant pleural mesothelioma. J. Mol. Diagn. 2014, 16, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Ak, G.; Tomaszek, S.C.; Kosari, F.; Metintas, M.; Jett, J.R.; Metintas, S.; Yildirim, H.; Dundar, E.; Dong, J.; Aubry, M.C.; et al. microRNA and mRNA features of malignant pleural mesothelioma and benign asbestos-related pleural effusion. Biomed. Res. Int. 2015, 2015, 635748. [Google Scholar] [CrossRef] [PubMed]
- Birnie, K.A.; Yip, Y.Y.; Ng, D.C.; Kirschner, M.B.; Reid, G.; Prele, C.M.; Musk, A.W.; Lee, Y.C.; Thompson, P.J.; Mutsaers, S.E.; et al. Loss of mir-223 and jnk signaling contribute to elevated stathmin in malignant pleural mesothelioma. Mol. Cancer Res. 2015, 13, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Cappellesso, R.; Nicole, L.; Caroccia, B.; Guzzardo, V.; Ventura, L.; Fassan, M.; Fassina, A. Young investigator challenge: MicroRNA-21/microRNA-126 profiling as a novel tool for the diagnosis of malignant mesothelioma in pleural effusion cytology. Cancer Cytopathol. 2016, 124, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Cappellesso, R.; Galasso, M.; Nicole, L.; Dabrilli, P.; Volinia, S.; Fassina, A. Mir-130a as a diagnostic marker to differentiate malignant mesothelioma from lung adenocarcinoma in pleural effusion cytology. Cancer Cytopathol. 2017, 125, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.G.; Johnen, G.; Bryk, O.; Jockel, K.H.; Bruning, T. Identification of miRNA-103 in the cellular fraction of human peripheral blood as a potential biomarker for malignant mesothelioma—A pilot study. PLoS ONE 2012, 7, e30221. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.G.; Casjens, S.; Johnen, G.; Bryk, O.; Raiko, I.; Pesch, B.; Kollmeier, J.; Bauer, T.T.; Bruning, T. Combination of mir-103a-3p and mesothelin improves the biomarker performance of malignant mesothelioma diagnosis. PLoS ONE 2014, 9, e114483. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, M.; Staffolani, S.; Nocchi, L.; Neuzil, J.; Strafella, E.; Manzella, N.; Mariotti, L.; Bracci, M.; Valentino, M.; Amati, M.; et al. Clinical significance of circulating mir-126 quantification in malignant mesothelioma patients. Clin. Biochem. 2012, 45, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.B.; Cheng, Y.Y.; Badrian, B.; Kao, S.C.; Creaney, J.; Edelman, J.J.; Armstrong, N.J.; Vallely, M.P.; Musk, A.W.; Robinson, B.W.; et al. Increased circulating mir-625-3p: A potential biomarker for patients with malignant pleural mesothelioma. J. Thorac. Oncol. 2012, 7, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Pigati, L.; Yaddanapudi, S.C.; Iyengar, R.; Kim, D.J.; Hearn, S.A.; Danforth, D.; Hastings, M.L.; Duelli, D.M. Selective release of microRNA species from normal and malignant mammary epithelial cells. PLoS ONE 2010, 5, e13515. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Schafer, L.; Wang, H.; Schmitz, T.; Flender, A.; Schueler, R.; Hammerstingl, C.; Nickenig, G.; Sinning, J.M.; Werner, N. Kinetics of circulating microRNAs in response to cardiac stress in patients with coronary artery disease. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Schulte, C.; Karakas, M.; Zeller, T. microRNAs in cardiovascular disease—Clinical application. Clin. Chem. Lab. Med. 2017, 55, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Gayosso-Gómez, L.V.; Zárraga-Granados, G.; Paredes-Garcia, P.; Falfán-Valencia, R.; Vazquez-Manríquez, M.E.; Martinez-Barrera, L.M.; Castillo-Gonzalez, P.; Rumbo-Nava, U.; Guevara-Gutierrez, R.; Rivera-Bravo, B.; et al. Identification of circulating miRNA profiles that distinguish malignant pleural mesothelioma from lung adenocarcinoma. EXCLI J. 2014, 13, 740–750. [Google Scholar] [PubMed]
- Lamberti, M.; Capasso, R.; Lombardi, A.; Di Domenico, M.; Fiorelli, A.; Feola, A.; Perna, A.F.; Santini, M.; Caraglia, M.; Ingrosso, D. Two different serum miRNA signatures correlate with the clinical outcome and histological subtype in pleural malignant mesothelioma patients. PLoS ONE 2015, 10, e0135331. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Wang, J.; Liew, O.W.; Richards, A.M.; Chen, Y.T. microRNA and heart failure. Int. J. Mol. Sci. 2016, 17, 502. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fan, J.; Yin, Z.; Wang, F.; Chen, C.; Wang, D.W. Identification of cardiac-related circulating microRNA profile in human chronic heart failure. Oncotarget 2016, 7, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Abd-El-Fattah, A.A.; Sadik, N.A.; Shaker, O.G.; Aboulftouh, M.L. Differential microRNAs expression in serum of patients with lung cancer, pulmonary tuberculosis, and pneumonia. Cell Biochem. Biophys. 2013, 67, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, Z.; Wei, W.; Liu, Z.; Fleming, J.; Zhang, S.; Lin, N.; Wang, M.; Chen, M.; Xu, Y.; et al. Identification of serum microRNA biomarkers for tuberculosis using Rna-seq. PLoS ONE 2014, 9, e88909. [Google Scholar] [CrossRef] [PubMed]
- Santarelli, L.; Staffolani, S.; Strafella, E.; Nocchi, L.; Manzella, N.; Grossi, P.; Bracci, M.; Pignotti, E.; Alleva, R.; Borghi, B.; et al. Combined circulating epigenetic markers to improve mesothelin performance in the diagnosis of malignant mesothelioma. Lung Cancer 2015, 90, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Bononi, I.; Comar, M.; Puozzo, A.; Stendardo, M.; Boschetto, P.; Orecchia, S.; Libener, R.; Guaschino, R.; Pietrobon, S.; Ferracin, M.; et al. Circulating microRNAs found dysregulated in ex-exposed asbestos workers and pleural mesothelioma patients as potential new biomarkers. Oncotarget 2016, 7, 82700–82711. [Google Scholar] [CrossRef] [PubMed]
- Cavalleri, T.; Angelici, L.; Favero, C.; Dioni, L.; Mensi, C.; Bareggi, C.; Palleschi, A.; Rimessi, A.; Consonni, D.; Bordini, L.; et al. Plasmatic extracellular vesicle microRNAs in malignant pleural mesothelioma and asbestos-exposed subjects suggest a 2-miRNA signature as potential biomarker of disease. PLoS ONE 2017, 12, e0176680. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.G.; Gawrych, K.; Casjens, S.; Brik, A.; Lehnert, M.; Taeger, D.; Pesch, B.; Kollmeier, J.; Bauer, T.T.; Johnen, G.; et al. Circulating mir-132-3p as a candidate diagnostic biomarker for malignant mesothelioma. Dis. Markers 2017, 2017, 9280170. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S.; Yun, J.; Lim, S.N.; Han, J.H.; Lee, K.H.; Kim, S.T.; Kang, M.H.; Son, S.M.; Lee, Y.M.; Choi, S.Y.; et al. Downregulation of cell-free mir-198 as a diagnostic biomarker for lung adenocarcinoma-associated malignant pleural effusion. Int. J. Cancer 2013, 133, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Micolucci, L.; Akhtar, M.M.; Olivieri, F.; Rippo, M.R.; Procopio, A.D. Diagnostic value of microRNAs in asbestos exposure and malignant mesothelioma: Systematic review and qualitative meta-analysis. Oncotarget 2016, 7, 58606–58637. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.D.; Rao, M.; Xi, S.; Inchauste, S.; Mani, H.; Fetsch, P.; Filie, A.; Zhang, M.; Hong, J.A.; Walker, R.L.; et al. Polycomb repressor complex-2 is a novel target for mesothelioma therapy. Clin. Cancer Res. 2012, 18, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Peltier, H.J.; Latham, G.J. Normalization of microRNA expression levels in quantitative rt-pcr assays: Identification of suitable reference RNA targets in normal and cancerous human solid tissues. RNA 2008, 14, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative rt-pcr data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1–research0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The miqe guidelines: Minimum information for publication of quantitative real-time pcr experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Rutjes, A.W.; Reitsma, J.B.; Di Nisio, M.; Smidt, N.; van Rijn, J.C.; Bossuyt, P.M. Evidence of bias and variation in diagnostic accuracy studies. CMAJ 2006, 174, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Lijmer, J.G.; Mol, B.W.; Heisterkamp, S.; Bonsel, G.J.; Prins, M.H.; van der Meulen, J.H.; Bossuyt, P.M. Empirical evidence of design-related bias in studies of diagnostic tests. JAMA 1999, 282, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Bossuyt, P.M.; Reitsma, J.B.; Bruns, D.E.; Gatsonis, C.A.; Glasziou, P.P.; Irwig, L.M.; Lijmer, J.G.; Moher, D.; Rennie, D.; de Vet, H.C. Towards complete and accurate reporting of studies of diagnostic accuracy: The stard initiative. BMJ 2003, 326, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; McShane, L.M.; Sauerbrei, W.; Taube, S.E. Reporting recommendations for tumor marker prognostic studies (remark): Explanation and elaboration. BMC Med. 2012, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gotzsche, P.C.; Vandenbroucke, J.P. The strengthening the reporting of observational studies in epidemiology (strobe) statement: Guidelines for reporting observational studies. PLoS Med. 2007, 4, e296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Studies in | Sample Source | Study Design and Sample Size | Assay (and Number of miRNAs Analyzed) | miRNAs Differentially Expressed in MPM | ROC Analysis | Study Aim | Selection of Endogenous Stable Normalizer (Validation Phase) | Reference |
|---|---|---|---|---|---|---|---|---|
| Pleural tissue (frozen) | Discovery: 17 MPM pleural tissue vs. 1 total RNA from normal human pericardium | Microarray (723) | let-7b*↑, MiR-1228*↑, miR-195*↑, miR-30b*↑, miR-32*↑, miR-345↑, miR-483-3p↑, miR-584↑, miR-595↑, miR-615-3p↑, and miR-885-3p↑. let-7e*↓, miR-144*↓, miR-203↓, miR-340*↓, miR-34a*↓, miR-423↓, miR-582↓, miR-7-1*↓ and miR-9↓ | Not performed (NP) | Oncogenic mechanisms | NA | [58] | |
| Tissue | Pleural and lung tissue | Discovery: 15 MPM pleural tissues vs. 10 AD pleural tissues. Validation: 100 MPM pleural tissues vs. 32 AD lung tissues | Microarray (2564), RT-qPCR (7) | RT-qPCR: miR-200c↓, miR-200b↓, miR-203↓, miR-141↓, miR-429↓ and miR-205↓ | Specificity and sensitivity >80% | Diagnosis | NP. Use of RNU44 and RNU48 | [59] |
| Pleural and several tumor tissue (FFPE) | Discovery: 7 MPM pleural tissues vs. 97 epithelial carcinomas. Validation: (1) 32 pleura tissues MPM vs. 113 epithelial carcinomas. (2) 16 MPM pleural tissues vs. 23 epithelial carcinomas. (3) 14 pleural tissues MPM vs. 46 epithelial carcinomas | Microarray (747), RT-qPCR (3) | RT-qPCR: miR-193a-3p↑, miR-200c↓ and miR-192↓ | Specificity 94%, sensitivity 100% | Diagnosis | NP. Use of U6 snoRNA | [60] | |
| Pleural tissue (frozen), pleural tissue (FFPE). | Discovery: 10 MPM vs. 5 Healthy controls (frozen). Validation: 27 MPM vs. 27 adjacent normal pleural tissues (FFPE) | PCRArray (88). RT-qPCR (3) | miR-126↓ | NP | Diagnosis | NP. Use of U6 small nuclear RNA | [29] | |
| Pleural tissue (frozen) | Discovery: 25 MPM vs. 6 normal parietal pleura (patients without cancer). Validation: Same cohort? Not specified | Microarray (1145), RT-qPCR (4) | RT-qPCR: miR-206↓, miR-1↓, miR-483-5p↓ and miR-155*↑ | NP | Oncogenic mechanisms | NP. Use of RNU44 | [62] | |
| Pleural tissue (FFPE). Also MPM cell lines. | Discovery: [74]. Validation: 60 MPM vs. 23 normal pleural tissues | RT-qPCR (4) | miR-16↓, miR15a↓, miR-15b↓, and miR-195↓ | NP | New therapy targets | NP. Use of RNU6B | [63] | |
| Pleural tissue (fresh, frozen & FFPE). Peritoneal tissue (frozen and FFPE). | Discovery: 29 MPM pleural tissues vs. 12 peritoneal mesothelial cysts (FFPE). Validation: (1) 6 MPM pleural tissues vs. 14 benign pleural tissues (Fresh). (2) 36 pleural tissues MPM vs. 36 peritoneal mesothelium (frozen) | Microarray (887), RT-qPCR (1) | RT-qPCR: miR-145↓ | NP | Oncogenic mechanisms | NP. Use of RNU6B and RNU49 | [64] | |
| Pleural tissue (FFPE) | Discovery: 5 MPM pleural tissues vs. 5 non-cancerous/non-inflammatory pleural tissues vs. 4 pleural chronic inflammation tissues vs. 5 mesothelial hyperplasia tissue | PCRArray (667) | miR-517b-3p↓, miR-627↓, miR-766-3p↓, miR-101-3p↓, miR-501-3p↓, miR-212-3p↓, miR-596↓, miR-145-5p↓, miR-671-3p↓, miR-181a-5p↓, miR-18a-3p↓, miR-30e-3p↑, miR-34a-3p↑, miR-622↑, let-7-g-5p↑, miR-196b-5p↑, miR-135b-5p↑, miR-18a-5p↑, miR-302b-3p↑ | NP | Oncogenic mechanisms | Normalization factor: the global mean expression value | [65] | |
| Pleural tissue (FFPE) | Discovery: 5 preoperative pleural tissues with MPM (before Cth = DB) and 5 pleural tissues MPM (after Cth = MPMc) vs. 5 non-neoplastic pleura tissues (after Cth = NNP). Validation: 40 MPMc vs. 12 DB vs. 14 NNP | PCRArray (742), RT-qPCR (4) | RT-qPCR: miR-126↓*, miR-143↓, miR-145↓, miR-652↓ | Specificity and sensitivity close to or >80% | Diagnosis | NP. Use of snord49A | [66] | |
| Pleural tissue (frozen) | Discovery: 18 MPM pleural tissues vs. 6 pleural tissues from benign asbestos-related pleural effusion patients (BAPE) (tissue with unspecific pleuritis/fibrosis) | PCRArray (384) | miR-484↑, miR-320↑, let-7a↑, miR-125a-5p↑ | Specificity and sensitivity close to or >80% | Diagnosis | NP. Use of U6 snoRNA | [67] | |
| Pleural tissue (FFPE) and cells from pleural effusion. | Discovery and Validation: 17 MPM pleural tissues vs. 6 normal pleural tissues patients without cancer undergoing cardiac or aortic surgery. Cells from pleural effusion of 26 MPM patients vs. 10 benign pleural diseases | RT-qPCR (1) | miR-223↓ | NP | Oncogenic mechanisms | NP. Use of RNU6B for tumor and RNU48, RNU44, or SNOR202 for cells | [68] | |
| Cell lines, pleural tissue (FFPE), pleural citology | Discovery: 2 MPM cell lines vs. 1 mesothelium cell line. Validation: (1) 51 MPM pleural tissues vs. 40 benign/reactive pleurae. (2) 29 MPM cytologic specimens vs. 24 reactive mesothelial cells | RT-qPCR (15) | miR-19a↑, miR-19b↑, miR-25↑, miR-21↑, miR-126↓ | Specificity and sensitivity >80% | Diagnosis | NP. Use of RNA U6B | [69] | |
| Pleural tissue (FFPE), pleural citology | Discovery: Bionformatic analysis 3 database. Validation: 41 epithelioid MPM vs. 40 AD and 26 cytologic specimen epitheloid MPM vs. 26 AD | RT-qPCR (6) | miR-130a↑(histological and cytological specimens) | Specificity 67% and sensitivity 77% | Differential diagnosis MPM vs. AD | NP. Use of RNU6B | [70] | |
| Peripheral blood | Cellular fraction of peripheral blood | Discovery: 23 MPM vs. 17 asbestos-exposed controls (AE). Validation: 23 MPM vs. 17 AE vs. 25 healthy controls | Microarray (328), RT-qPCR (2) | RT-qPCR: miR-103↓ | Specificity and sensitivity >80% | Diagnosis | Yes: miR-125a | [71] |
| Cellular fraction of peripheral blood | Discovery: [71].Validation: 43 MPM vs. 52 asbestos-exposed controls | RT-qPCR (1) | miR-103a-3p↓ (plus mesothelin↑ in plasma) | Specificity and sensitivity >80% | Diagnosis | Yes: miR-125a | [72] | |
| Serum and plasma | Serum | Discovery: In pleural tissue (miR-126↓ same paper). Validation: 44 MPM vs. 196 asbestos-exposed controls vs. 50 Healthy controls | RT-qPCR (1) | miR-126↓ | Specificity 74% and sensitivity 73%. | Diagnosis | NP. Use of U6 snoRNA | [29] |
| Serum | Pre-Validation: [29]. Validation: 45 MPM vs. 20 NSCLC vs. 56 healthy controls | RT-qPCR (1) | miR-126↓ | Specificity 60% and sensitivity 80% | Diagnosis | Yes: U6 snoRNA and use of exogenous control cel-miR-39 | [73] | |
| Plasma, serum. Also pleural tissue (FFPE) | Discovery: 5 MPM (plasma) vs. 3 healthy controls (HC). Validation: 15 MPM (plasma) vs. 14 HC. Validation serum: 30 MPM (serum) vs. 10 asbestosis (serum). Validation tissue: 18 MPM pleural tissues vs. 7 pericardial tissues | Microarray (854), RT-qPCR | RT-qPCR: miR-625-3p↑ (plasma & serum). miR-625-3p↑, miR-29c*↓, miR-16↓, miR-196b↓, miR-26a-2-3p↓ and miR-1914-3p↓ (tissue) | Specificity & sensitivity close to or >80 % (plasma & serum miR-625-3p) | Diagnosis | Only SD of Cq range values without specified clearly which samples were used. Previous work (plasma): miR-16 | [74] | |
| Serum | Discovery: 11 MPM (epithelial) vs. 45 healthy controls vs. 36 AD | Deep sequencing (Ilumina) | MPM vs. control: miR-4791↑, miR-185-5p↑, miR-96-5p↑, miR-1271-5p, miR-1292-5p↑, miR-409-5p↑ y miR-92b-5p↑ | NP | Diagnosis | NA | [79] | |
| Serum | Discovery: 14 MPM vs. 10 non-cancer related effusions patients. Validation: Not specified | PCRArray (384), RT-qPCR (7) | RT-qPCR: miR-101↑, miR-25↑, miR-26b↑, miR-335↑, miR-29a↑, miR-516↑, miR-433↑, miR-191↓, miR-223↓ | Not performed | Prognosis | NP. Use of miR-16 | [80] | |
| Serum | Discovery: [29]. Validation 1: 45 MPM vs. 99 asbestos-exposed subjects (AE) vs. 44 healthy subjects. Validation 2:18 MPM vs. 50 (AE) vs. 20 healthy controls and 42 lung cancer (LC) patients | RT-qPCR (1) | Combination SMRPs↑, miR-126↓ and Met-TM↑ | AUC of 0.857 (95% CI, 0.767–0.927) | Diagnosis | NP. Use of U6 snoRNA. Use of exogenous control cel-miR-39 | [85] | |
| Serum | Discovery: 10 MPM vs. 10 asbestos-exposed subjects (AE) vs. 10 healthy controls (HC). Validation: 20 MPM vs. 15 AE vs. 14 HC | Microarray (1201), RT-qPCR (3) | RT-qPCR: miR-197-3p↑, miR-1281↑, miR-32-3p↑ (MPM vs. HS and MPM vs. AE) | AUC 95% CI, 0.5398-0.8959 (miR-197-3p) | Diagnosis | Yes: miR-3665 | [86] | |
| Plasma (exosomal fraction) | Discovery: 23 MPM vs. 19 past asbestos-exposed subjects. Validation: Same samples minus 4 | OpenArray (754). RT-qPCR (20) | 2-miRNAs signatures: miR-103a-3p↓, miR-30e-3p↓ | Specificity 80% and sensitivity 95.5% | Diagnosis | RNU48. It is not clear | [87] | |
| Plasma | Discovery: 21 MPM vs. 21 asbestos-exposed controls. Validation: 22 MPM vs. 44 asbestos-exposed controls | PCRArray (377), RT-qPCR (2) | RT-qPCR: miR-132-3p↓ | Specificity 61% and sensitivity 86% | Diagnosis | Yes. Use of miR-146b-5p. Another untested normalizer was used too | [88] |
| miRNA | Number of Studies | Sample Source | Comparative Analysis Design | References |
|---|---|---|---|---|
| miR-200c↓ | 2 | Pleural tissue | (1) MPM vs. AD. (2) MPM vs. epithelial carcinoma | [59,60] |
| miR-126↓ | 3 | Pleural tissue | (1) MPM vs. normal pleura. (2) MPM vs. normal pleura (with Cth). (3) MPM vs. benign/reactive pleurae | [29,66,69] |
| miR-145↓ | 3 | Pleural tissue | (1) MPM vs. benign pleural tissue. (2) MPM vs. normal pleura. (3) MPM vs. normal pleura (with Cth) | [64,65,66] |
| miR-16↓ | 2 | Pleural tissue | (1) MPM vs. pericardial tissues. (2) MPM vs. normal pleural tissue | [63,74] |
| miR-103↓ | 2 | Cellular fraction of peripheral blood | Two subsequent studies: MPM vs. asbestos-exposed controls | [71,72] |
| miR-126↓ | 3 | Serum | Three subsequent studies: (a) MPM vs. asbestos-exposed controls vs. healthy controls. (b) MPM vs. Healthy controls. (c) MPM vs. asbestos-exposed controls vs. healthy controls | [29,73,85] |
| miRNA with Deregulated Expression in MPM | Potential Function | Biological Effect of Experimental Manipulation of miRNA Expression | Other Effects | Reference |
|---|---|---|---|---|
| miR-16↓ (tissue) | Tumor suppressor | Restoring miR-16: growth inhibition, cell cycle arrest in G0/G1, increased apoptosis and reduced colony formation in MPM cell lines | Correlation with downregulation of Bcl2, CCND1. Administration of miR-16-containing minicells to xenografted mice inhibited tumor growth | [63] |
| miR-1↓ (tissue) | Tumor suppressor | Restoring miR-1: cell cycle arrest, increased apoptosis. | Correlation with upregulation of p53, BAX, p16/21; and downregulation of Bcl2 and survivin | [62] |
| miR-145↓ (tissue) | Tumor suppressor | Restoring miR-145: reduction of proliferation and migration of two out of three transfected MPM cell lines | Xenotransplant (transfected MPM cell line): inhibition of tumor growth in 6 of 8 treated mice compared to controls | [64] |
| miR-223↓ (tissue) | Tumor suppressor | Over-expression of miR-223: reduction of two MPM cell lines motility. | STMN1 levels were reduced and tubulin acetylation was induced | [68] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Rivera, V.; Negrete-García, M.C.; Ávila-Moreno, F.; Ortiz-Quintero, B. Secreted and Tissue miRNAs as Diagnosis Biomarkers of Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2018, 19, 595. https://doi.org/10.3390/ijms19020595
Martínez-Rivera V, Negrete-García MC, Ávila-Moreno F, Ortiz-Quintero B. Secreted and Tissue miRNAs as Diagnosis Biomarkers of Malignant Pleural Mesothelioma. International Journal of Molecular Sciences. 2018; 19(2):595. https://doi.org/10.3390/ijms19020595
Chicago/Turabian StyleMartínez-Rivera, Vanessa, María Cristina Negrete-García, Federico Ávila-Moreno, and Blanca Ortiz-Quintero. 2018. "Secreted and Tissue miRNAs as Diagnosis Biomarkers of Malignant Pleural Mesothelioma" International Journal of Molecular Sciences 19, no. 2: 595. https://doi.org/10.3390/ijms19020595