The Connexin Mimetic Peptide Gap27 and Cx43-Knockdown Reveal Differential Roles for Connexin43 in Wound Closure Events in Skin Model Systems


Abstract
:1. Introduction
2. Results
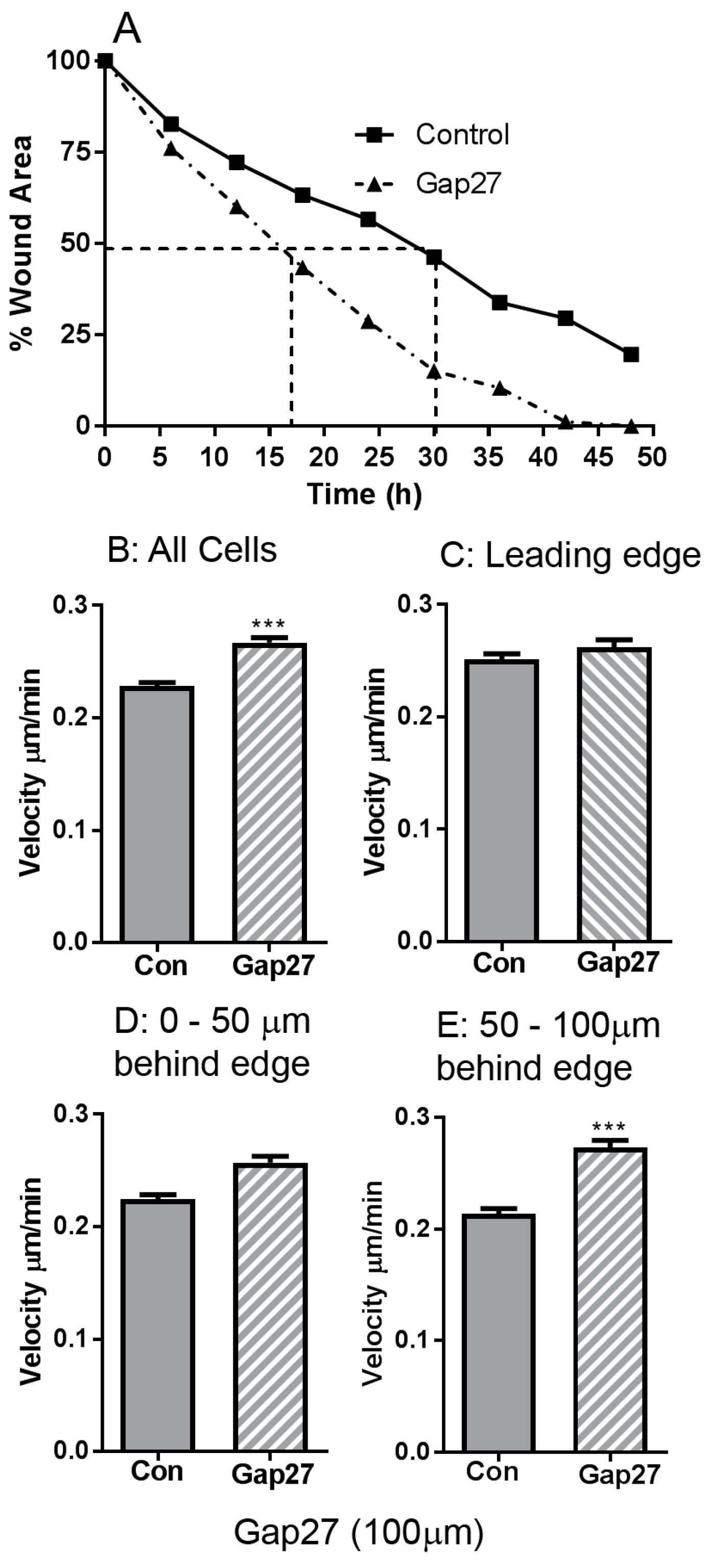
2.1. The Impact of Gap27 on Cell Migration Rates in Juvenile Foreskin Fibroblasts
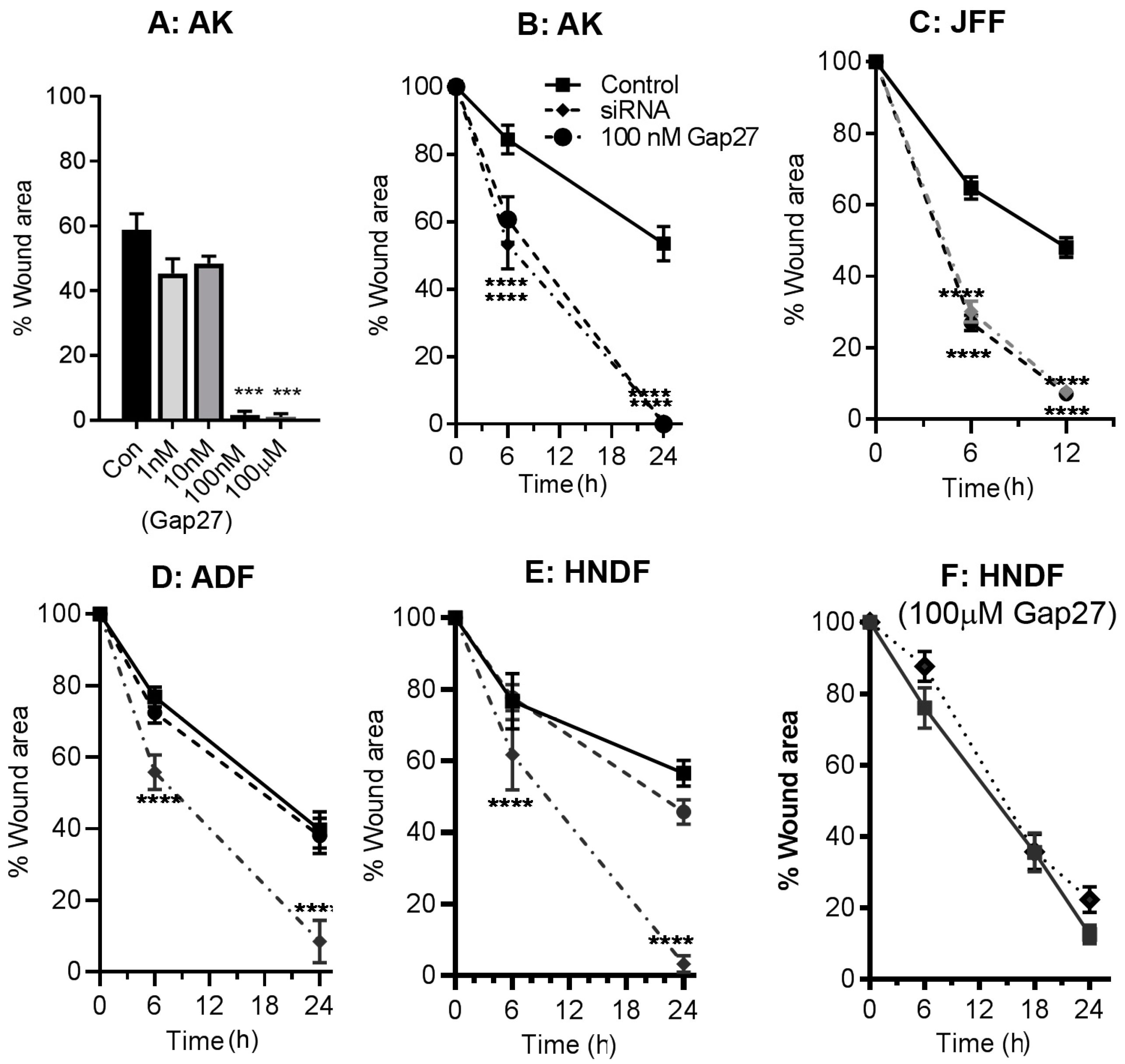
2.2. Impact of Gap27 and SiRNA Targeted to Cx43 on Cell Migration in Skin Model Systems
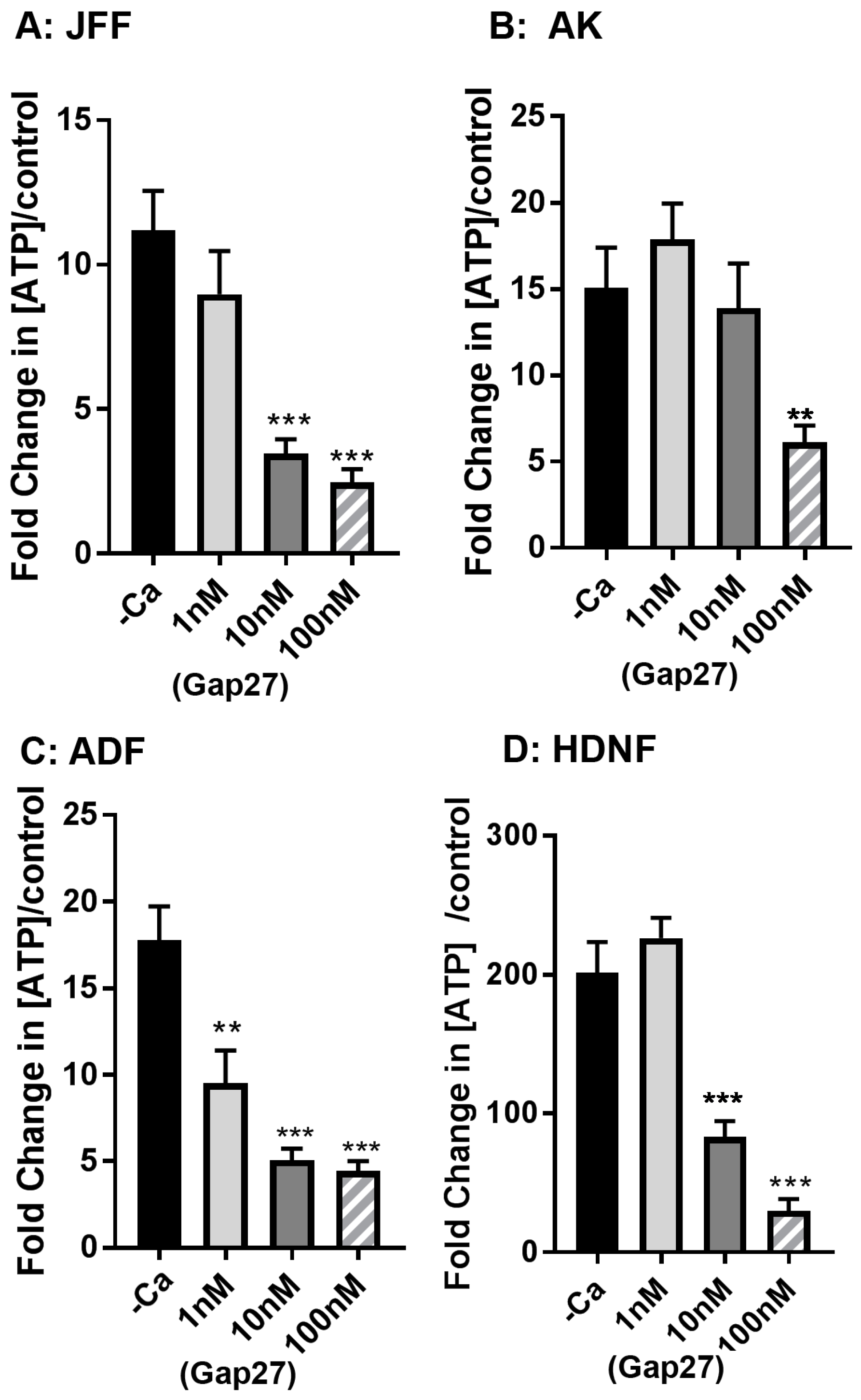
2.3. Gap27 Attenuates Hemichannel Signalling at Lower Doses than Gap Junction Coupling
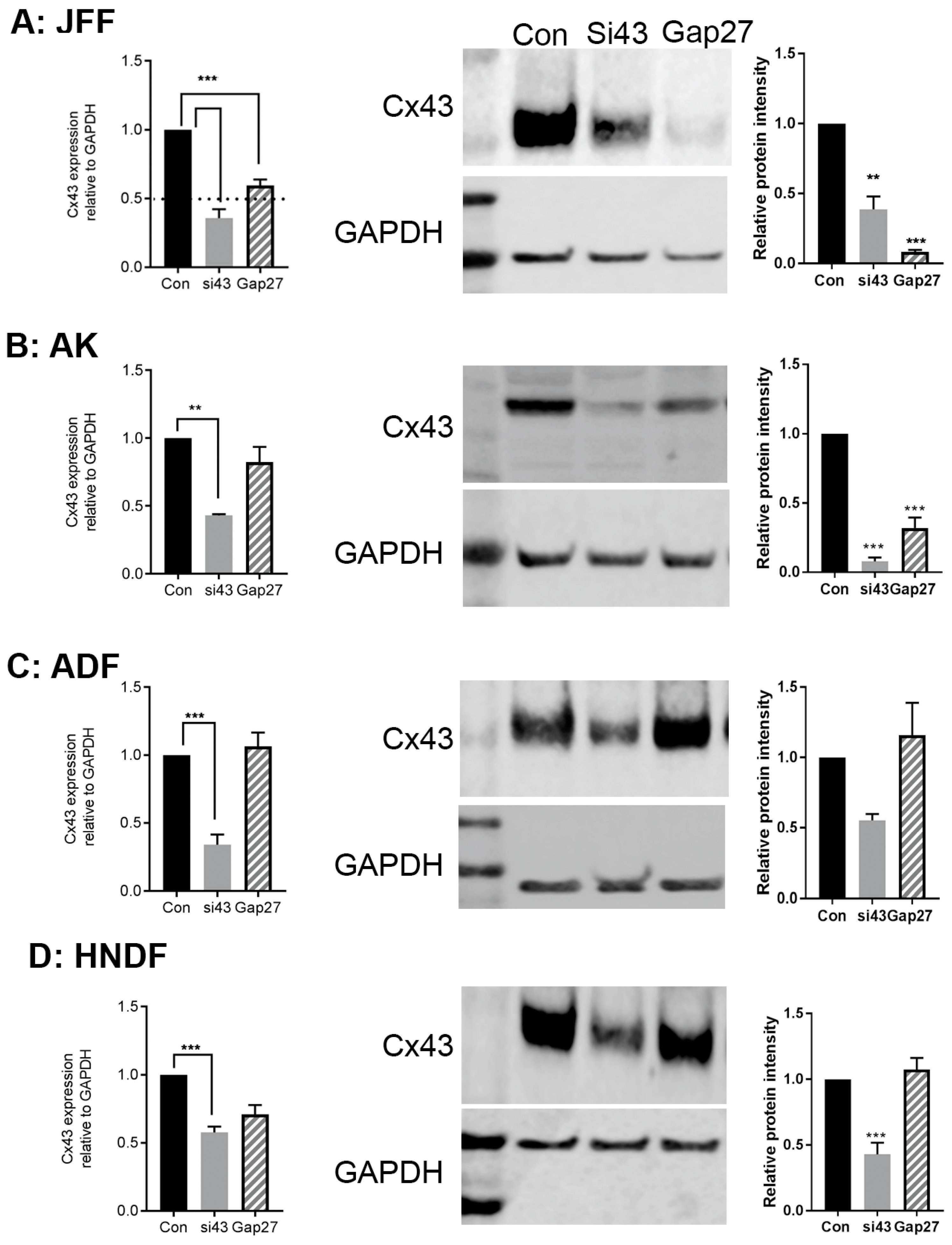
2.4. The Impact of Gap27 and SiRNA Targeted to Cx43 on Gene and Protein Expression Profiles in Skin Model Systems
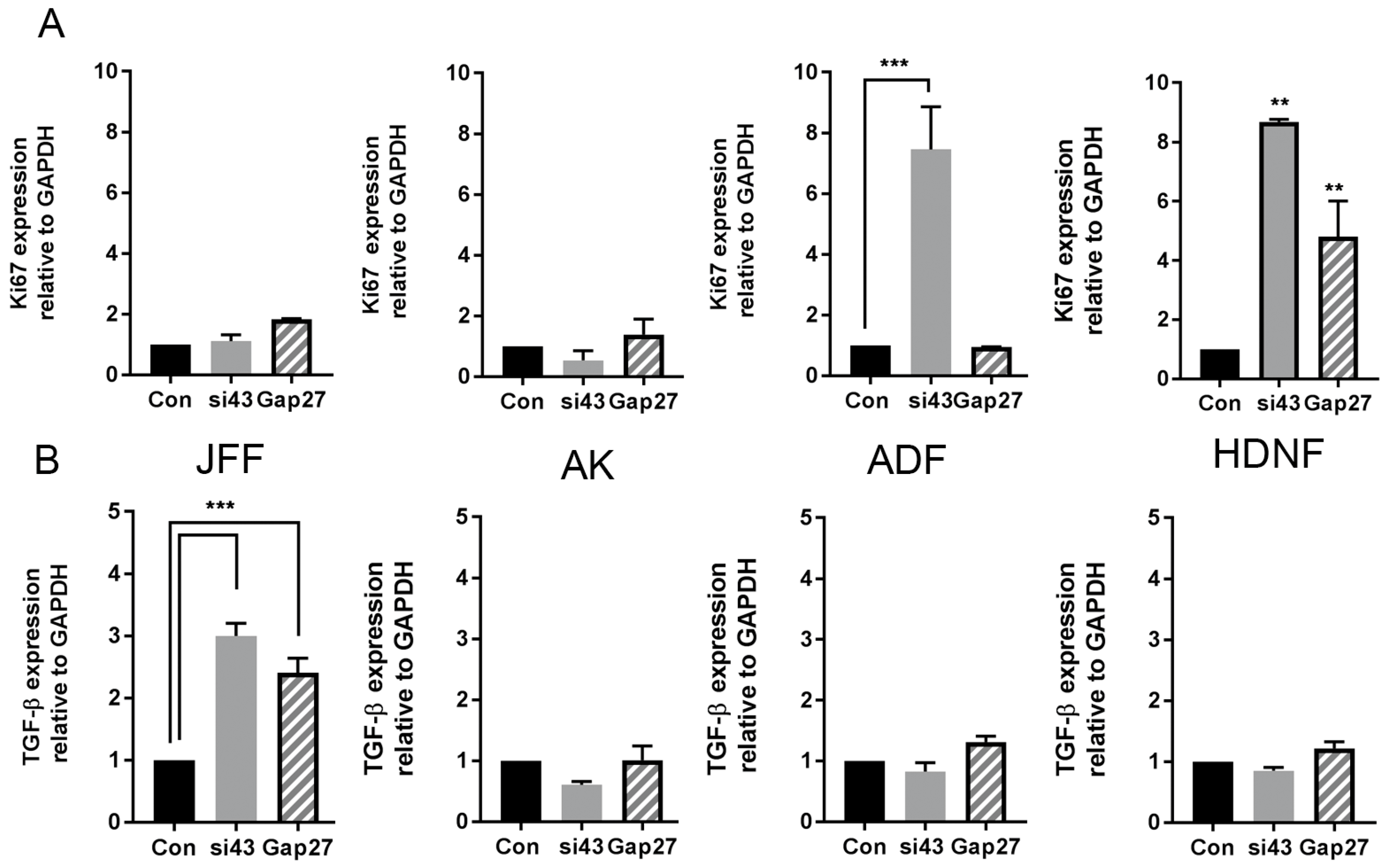
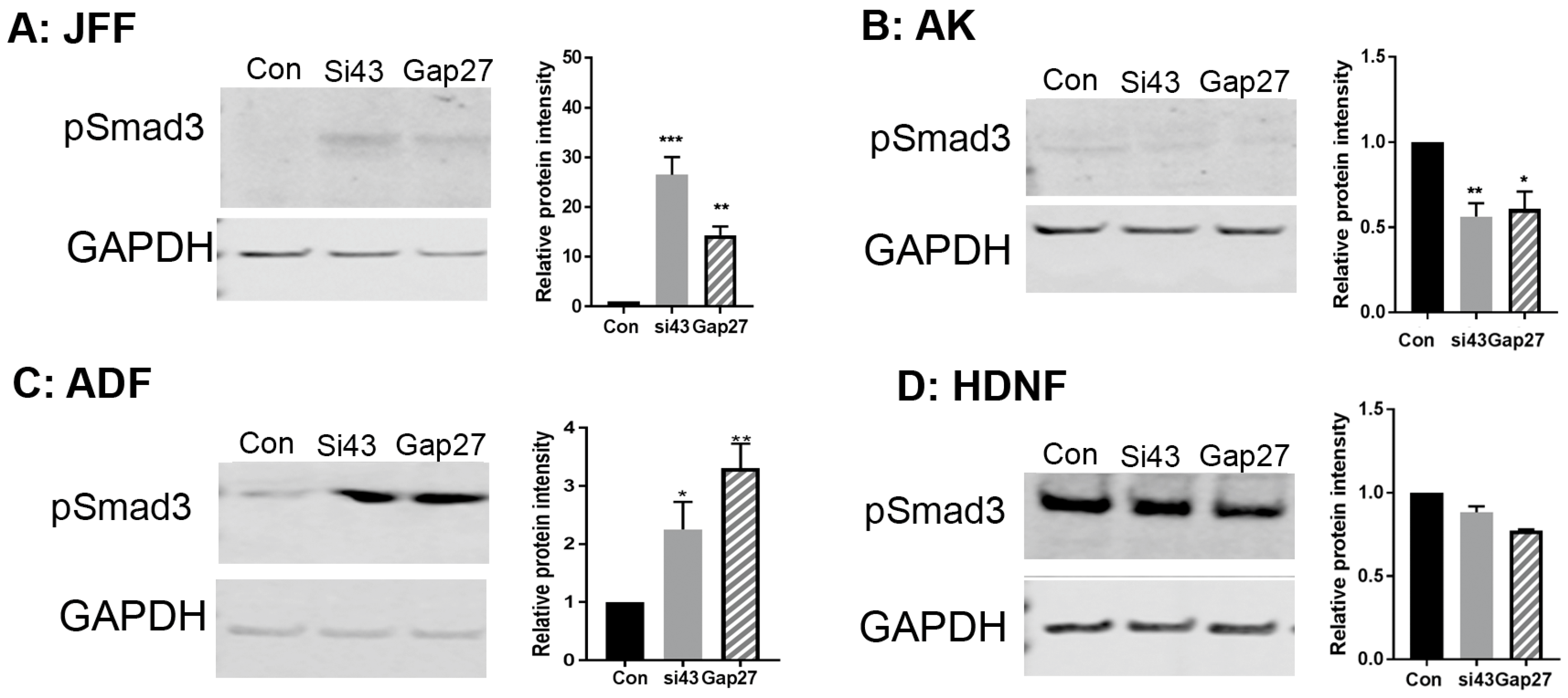
2.5. The Impact of Gap27 and SiRNA Targeted to Cx43 on Cell Proliferation, TGF-β1 and SMAD3 Signalling Pathways
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Inhibition of Connexin Mediated Communication
4.3. Knockdown of Cx43 Expression by siRNA
4.4. Hemichannel Functionality Assays
4.5. Scrape Wound Assays and Time-Lapse Microscopic Analysis of Cell Migration
4.6. RNA Extraction and Real Time PCR
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Cx | connexin |
| CMP | connexin mimetic peptide |
| CMC | connexin mediated communication |
| JFF | human juvenile foreskin derived fibroblasts |
| HNDF | human neonatal dermal fibroblasts |
| AF | adult dermal derived fibroblasts |
| AK | adult keratinocytes |
| SFM | serum free media |
References
- Evans, W.H.; Martin, P.E. Gap junctions: Structure and function (Review). Mol. Membr. Biol. 2002, 19, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.H.; Bultynck, G.; Leybaert, L. Manipulating connexin communication channels: Use of peptidomimetics and the translational outputs. J. Mem. Biol. 2012, 245, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.H.; Leybaert, L. Mimetic peptides as blockers of connexin channel-facilitated intercellular communication. Cell. Commun. Adhes. 2007, 14, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Willebrords, J.; Maes, M.; Crespo Yanguas, S.; Vinken, M. Inhibitors of connexin and pannexin channels as potential therapeutics. Pharmacol. Ther. 2017, 180, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.T.; Evans, W.H.; Griffith, T.M. Peptides homologous to extracellular loop motifs of connexin 43 reversibly abolish rhythmic contractile activity in rabbit arteries. J. Physiol. 1997, 503 Pt 1, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.T.; Martin, P.E.; Evans, W.H.; Randall, M.D.; Griffith, T.M. The endothelial component of cannabinoid-induced relaxation in rabbit mesenteric artery depends on gap junctional communication. J. Physiol. 1999, 520, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Dora, K.A.; Martin, P.E.; Chaytor, A.T.; Evans, W.H.; Garland, C.J.; Griffith, T.M. Role of heterocellular Gap junctional communication in endothelium-dependent smooth muscle hyperpolarization: inhibition by a connexin-mimetic peptide. Biochem. Biophys. Res. Commun. 1999, 254, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, I.R.; Chaytor, A.T.; Evans, W.H.; Griffith, T.M. Nitric oxide-independent relaxations to acetylcholine and A23187 involve different routes of heterocellular communication. Role of Gap junctions and phospholipase A2. Circ. Res. 1999, 84, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Griffith, T.M.; Chaytor, A.T.; Edwards, D.H. The obligatory link: Role of gap junctional communication in endothelium-dependent smooth muscle hyperpolarization. Pharmacol. Res. 2004, 49, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Straub, A.C.; Zeigler, A.C.; Isakson, B.E. The myoendothelial junction: connections that deliver the message. Physiology 2014, 29, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Hallett, M.B.; Leybaert, L.; Martin, P.E.; Evans, W.H. Perturbing plasma membrane hemichannels attenuates calcium signalling in cardiac cells and HeLa cells expressing connexins. Eur. J. Cell. Biol. 2009, 88, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Boitano, S.; Evans, W.H. Connexin mimetic peptides reversibly inhibit Ca2+ signaling through gap junctions in airway cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L623–L630. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.J.; Nicholson, L.F.; Feng, S.X.; Vis, J.C.; Green, C.R. A novel method of organotypic brain slice culture using connexin-specific antisense oligodeoxynucleotides to improve neuronal survival. Brain Res. 2010, 1353, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Orta, E.; Errington, R.J.; Evans, W.H. Gap junction intercellular communication during lymphocyte transendothelial migration. Cell. Biol. Int. 2002, 26, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Orta, E.; Evans, W.H. Gap junctions and connexins: potential contributors to the immunological synapse. J. Leukoc. Biol. 2002, 72, 636–642. [Google Scholar] [PubMed]
- Kameritsch, P.; Pogoda, K.; Pohl, U. Channel-independent influence of connexin 43 on cell migration. BBA-Bioenergetics 2012, 1818, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.M.; Houdek, P.; Husing, B.; Kaiser, C.; Moll, I. Connexins 26, 30, and 43: Differences among spontaneous, chronic, and accelerated human wound healing. J. Investig. Dermatol. 2004, 122, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.L.; Thrasivoulou, C.; Phillips, A.R. Connexins in wound healing; perspectives in diabetic patients. BBA-Bioenergetics 2012, 1818, 2068–2075. [Google Scholar] [CrossRef] [PubMed]
- Kretz, M.; Euwens, C.; Hombach, S.; Eckardt, D.; Teubner, B.; Traub, O.; Willecke, K.; Ott, T. Altered connexin expression and wound healing in the epidermis of connexin-deficient mice. J. Cell. Sci. 2003, 116 Pt 16, 3443–3452. [Google Scholar] [CrossRef] [PubMed]
- Mori, R.; Power, K.T.; Wang, C.M.; Martin, P.; Becker, D.L. Acute downregulation of connexin43 at wound sites leads to a reduced inflammatory response, enhanced keratinocyte proliferation and wound fibroblast migration. J. Cell. Sci. 2006, 119 Pt 24, 5193–5203. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Naranjo, A.; Cormie, P.; Serrano, A.E.; Hu, R.; O′Neill, S.; Wang, C.M.; Thrasivoulou, C.; Power, K.T.; White, A.; Serena, T.; et al. Targeting Cx43 and N-cadherin, which are abnormally upregulated in venous leg ulcers, influences migration, adhesion and activation of Rho GTPases. PLoS ONE 2012, 7, e37374. [Google Scholar] [CrossRef] [PubMed]
- Pollok, S.; Pfeiffer, A.C.; Lobmann, R.; Wright, C.S.; Moll, I.; Martin, P.E.; Brandner, J.M. Connexin 43 mimetic peptide Gap27 reveals potential differences in the role of Cx43 in wound repair between diabetic and non-diabetic cells. J. Cell. Mol. Med. 2011, 15, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.S.; Pollok, S.; Flint, D.J.; Brandner, J.M.; Martin, P.E. The connexin mimetic peptide Gap27 increases human dermal fibroblast migration in hyperglycemic and hyperinsulinemic conditions in vitro. J. Cell. Physiol. 2012, 227, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.S.; van Steensel, M.A.; Hodgins, M.B.; Martin, P.E. Connexin mimetic peptides improve cell migration rates of human epidermal keratinocytes and dermal fibroblasts in vitro. Wound Repair Regen. 2009, 17, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Ghatnekar, G.S.; Grek, C.L.; Armstrong, D.G.; Desai, S.C.; Gourdie, R.G. The effect of a connexin43-based Peptide on the healing of chronic venous leg ulcers: a multicenter, randomized trial. J. Investig. Dermatol. 2015, 135, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Coutinho, P.; Frank, S.; Franke, S.; Law, L.Y.; Martin, P.; Green, C.R.; Becker, D.L. Targeting connexin43 expression accelerates the rate of wound repair. Curr. Biol. 2003, 13, 1697–1703. [Google Scholar] [CrossRef] [PubMed]
- Lorraine, C. The role of connexins in skin wound healing events. Ph.D. Thesis, Glasgow Caledonian University, Glasgow, UK, available through British Library Electronic Theses Online System. 2015. [Google Scholar]
- Wright, J.A.; Richards, T.; Becker, D.L. Connexins and diabetes. Cardiol. Res. Pract. 2012, 2012, 496904. [Google Scholar] [CrossRef] [PubMed]
- Tarzemany, R.; Jiang, G.; Larjava, H.; Hakkinen, L. Expression and function of connexin 43 in human gingival wound healing and fibroblasts. PLoS ONE 2015, 10, e0115524. [Google Scholar] [CrossRef] [PubMed]
- Riding, A.; Pullar, C.E. ATP Release and P2 Y Receptor Signaling are Essential for Keratinocyte Galvanotaxis. J. Cell. Physiol. 2016, 231, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Grupcheva, C.N.; Laux, W.T.; Rupenthal, I.D.; McGhee, J.; McGhee, C.N.; Green, C.R. Improved corneal wound healing through modulation of gap junction communication using connexin43-specific antisense oligodeoxynucleotides. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Ormonde, S.; Chou, C.Y.; Goold, L.; Petsoglou, C.; Al-Taie, R.; Sherwin, T.; McGhee, C.N.; Green, C.R. Regulation of connexin43 gap junction protein triggers vascular recovery and healing in human ocular persistent epithelial defect wounds. J. Memb. Biol. 2012, 245, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ghatnekar, G.S.; O′Quinn, M.P.; Jourdan, L.J.; Gurjarpadhye, A.A.; Draughn, R.L.; Gourdie, R.G. Connexin43 carboxyl-terminal peptides reduce scar progenitor and promote regenerative healing following skin wounding. Regen. Med. 2009, 4, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.; Bryant, Z.J.; Ghatnekar, G.; Singh, U.P.; Gourdie, R.G.; Potts, J.D. A synthetic connexin 43 mimetic peptide augments corneal wound healing. Exp. Eye Res. 2013, 115, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Soder, B.L.; Propst, J.T.; Brooks, T.M.; Goodwin, R.L.; Friedman, H.I.; Yost, M.J.; Gourdie, R.G. The connexin43 carboxyl-terminal peptide ACT1 modulates the biological response to silicone implants. Plast. Reconstr. Surg. 2009, 123, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Danesh-Meyer, H.V.; Kerr, N.M.; Zhang, J.; Eady, E.K.; O′Carroll, S.J.; Nicholson, L.F.; Johnson, C.S.; Green, C.R. Connexin43 mimetic peptide reduces vascular leak and retinal ganglion cell death following retinal ischaemia. Brain 2012, 135 Pt 2, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.X.; Mat Nor, M.N.; Danesh-Meyer, H.V.; Vessey, K.A.; Fletcher, E.L.; O′Carroll, S.J.; Acosta, M.L.; Green, C.R. Connexin43 Mimetic Peptide Improves Retinal Function and Reduces Inflammation in a Light-Damaged Albino Rat Model. Investig. Ophthalmol. Vis.Sci. 2016, 57, 3961–3973. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Nguyen, T.; Tonkin, R.S.; Lees, J.G.; Warren, C.; O′Carroll, S.J.; Nicholson, L.F.B.; Green, C.R.; Moalem-Taylor, G.; Gorrie, C.A. Characterisation of Peptide5 systemic administration for treating traumatic spinal cord injured rats. Exp. Brain Res. 2017, 235, 3033–3048. [Google Scholar] [CrossRef] [PubMed]
- Elbadawy, H.M.; Mirabelli, P.; Xeroudaki, M.; Parekh, M.; Bertolin, M.; Breda, C.; Cagini, C.; Ponzin, D.; Lagali, N.; Ferrari, S. Effect of connexin43 inhibition by the mimetic peptide Gap27 on corneal wound healing, inflammation and neovascularization. Br. J. Pharmacol. 2016, 173, 2880–2893. [Google Scholar] [CrossRef] [PubMed]
- Tarzemany, R.; Jiang, G.; Jiang, J.X.; Larjava, H.; Hakkinen, L. Connexin43 Hemichannels Regulate the Expression of Wound Healing-Associated Genes in Human Gingival Fibroblasts. Sci. Rep. 2017, 7, 14157. [Google Scholar] [CrossRef] [PubMed]
- Abudara, V.; Bechberger, J.; Freitas-Andrade, M.; De Bock, M.; Wang, N.; Bultynck, G.; Naus, C.C.; Leybaert, L.; Giaume, C. The connexin43 mimetic peptide Gap19 inhibits hemichannels without altering gap junctional communication in astrocytes. Front. Cell. Neurosci. 2014, 8, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyyathurai, J.; Wang, N.; D′Hondt, C.; Jiang, J.X.; Leybaert, L.; Bultynck, G. The SH3—Binding domain of Cx43 participates in loop/tail interactions critical for Cx43—hemichannel activity. CMLS 2017. [Google Scholar] [CrossRef] [PubMed]
- Glass, B.J.; Hu, R.G.; Phillips, A.R.; Becker, D.L. The action of mimetic peptides on connexins protects fibroblasts from the negative effects of ischemia reperfusion. Biol. Open 2015, 4, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Kandyba, E.E.; Hodgins, M.B.; Martin, P.E. A murine living skin equivalent amenable to live-cell imaging: Analysis of the roles of connexins in the epidermis. J. Investig. Dermatol. 2008, 128, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; Esseltine, J.L.; Shao, Q.; Jabs, E.W.; Sampson, J.; Auranen, M.; Bai, D.; Laird, D.W. Specific functional pathologies of Cx43 mutations associated with oculodentodigital dysplasia. Mol. Biol. Cell. 2016, 27, 2172–2185. [Google Scholar] [CrossRef] [PubMed]
- Hills, C.E.; Siamantouras, E.; Smith, S.W.; Cockwell, P.; Liu, K.K.; Squires, P.E. TGFbeta modulates cell-to-cell communication in early epithelial-to-mesenchymal transition. Diabetologia 2012, 55, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Postlethwaite, A.E.; Keski-Oja, J.; Moses, H.L.; Kang, A.H. Stimulation of the chemotactic migration of human fibroblasts by transforming growth factor beta. J. Exp. Med. 1987, 165, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Nakagami, T.; Tanaka, H.; Hitomi, T.; Takamatsu, T. Cx43 mediates TGF-beta signaling through competitive Smads binding to microtubules. Mol. Biol. Cell. 2007, 18, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Leivonen, S.K.; Lazaridis, K.; Decock, J.; Chantry, A.; Edwards, D.R.; Kahari, V.M. TGF-β-elicited induction of tissue inhibitor of metalloproteinases (TIMP)-3 expression in fibroblasts involves complex interplay between Smad3, p38α, and ERK1/2. PLoS ONE 2013, 8, e57474. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Bi, J.; Owen, G.R.; Chen, W.; Rokka, A.; Koivisto, L.; Heino, J.; Hakkinen, L.; Larjava, H. Keratinocyte Microvesicles Regulate the Expression of Multiple Genes in Dermal Fibroblasts. J. Investig. Dermatol. 2015, 135, 3051–3059. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.S.; Berends, R.F.; Flint, D.J.; Martin, P.E. Cell motility in models of wounded human skin is improved by Gap27 despite raised glucose, insulin and IGFBP-5. Exp. Cell. Res. 2013, 319, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, S.R.; Best, A.K.; Wright, C.S.; Isakson, B.E.; Errington, R.J.; Martin, P.E. Enhanced connexin 43 expression delays intra-mitotic duration and cell cycle traverse independently of gap junction channel function. J. Cell. Biochem. 2010, 110, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Leithe, E.; Rivedal, E. Ubiquitination and down-regulation of gap junction protein connexin-43 in response to 12-O-tetradecanoylphorbol 13-acetate treatment. J. Biol. Chem. 2004, 279, 50089–50096. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Treatment | HC | Migration | Cx43 Protein | Proliferation | TGF-β1 | pSmad3 |
|---|---|---|---|---|---|---|---|
| JFF | SiRNA | ND | ↑ | ↓ | NE | ↑ | ↑ |
| Gap27 | ↓ | ↑ | ↓ | NE | ↑ | ↑ | |
| AK | SiRNA | ND | ↑ | ↓ | NE | NE | NE |
| Gap27 | ↓ | ↑ | ↓ | NE | NE | NE | |
| ADF | SiRNA | ND | ↑ | ↓ | ↑ | ↑ | ↑ |
| Gap27 | ↓ | NE | NE | NE | NE | ↑ | |
| HDNF | SiRNA | ND | ↑ | ↓ | ↑ | NE | NE |
| Gap27 | ↓ | NE | NE | ↑ | NE | NE |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faniku, C.; O’Shaughnessy, E.; Lorraine, C.; Johnstone, S.R.; Graham, A.; Greenhough, S.; Martin, P.E.M. The Connexin Mimetic Peptide Gap27 and Cx43-Knockdown Reveal Differential Roles for Connexin43 in Wound Closure Events in Skin Model Systems. Int. J. Mol. Sci. 2018, 19, 604. https://doi.org/10.3390/ijms19020604
Faniku C, O’Shaughnessy E, Lorraine C, Johnstone SR, Graham A, Greenhough S, Martin PEM. The Connexin Mimetic Peptide Gap27 and Cx43-Knockdown Reveal Differential Roles for Connexin43 in Wound Closure Events in Skin Model Systems. International Journal of Molecular Sciences. 2018; 19(2):604. https://doi.org/10.3390/ijms19020604
Chicago/Turabian StyleFaniku, Chrysovalantou, Erin O’Shaughnessy, Claire Lorraine, Scott R. Johnstone, Annette Graham, Sebastian Greenhough, and Patricia E.M. Martin. 2018. "The Connexin Mimetic Peptide Gap27 and Cx43-Knockdown Reveal Differential Roles for Connexin43 in Wound Closure Events in Skin Model Systems" International Journal of Molecular Sciences 19, no. 2: 604. https://doi.org/10.3390/ijms19020604