Comparison of Tumor- and Bone Marrow-Derived Mesenchymal Stromal/Stem Cells from Patients with High-Grade Osteosarcoma

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of Nine Patients with High-Grade OS and Sample Processing
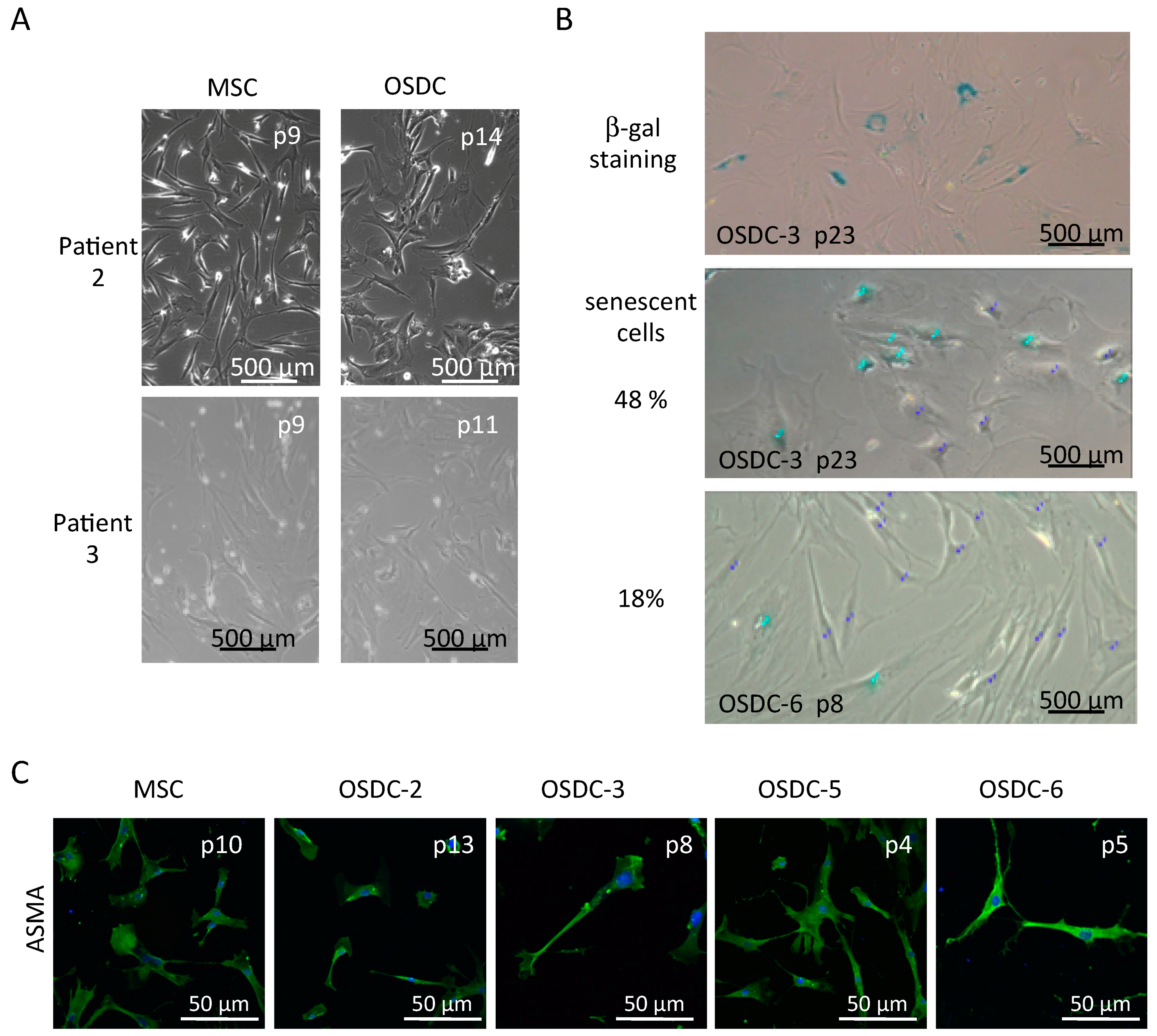
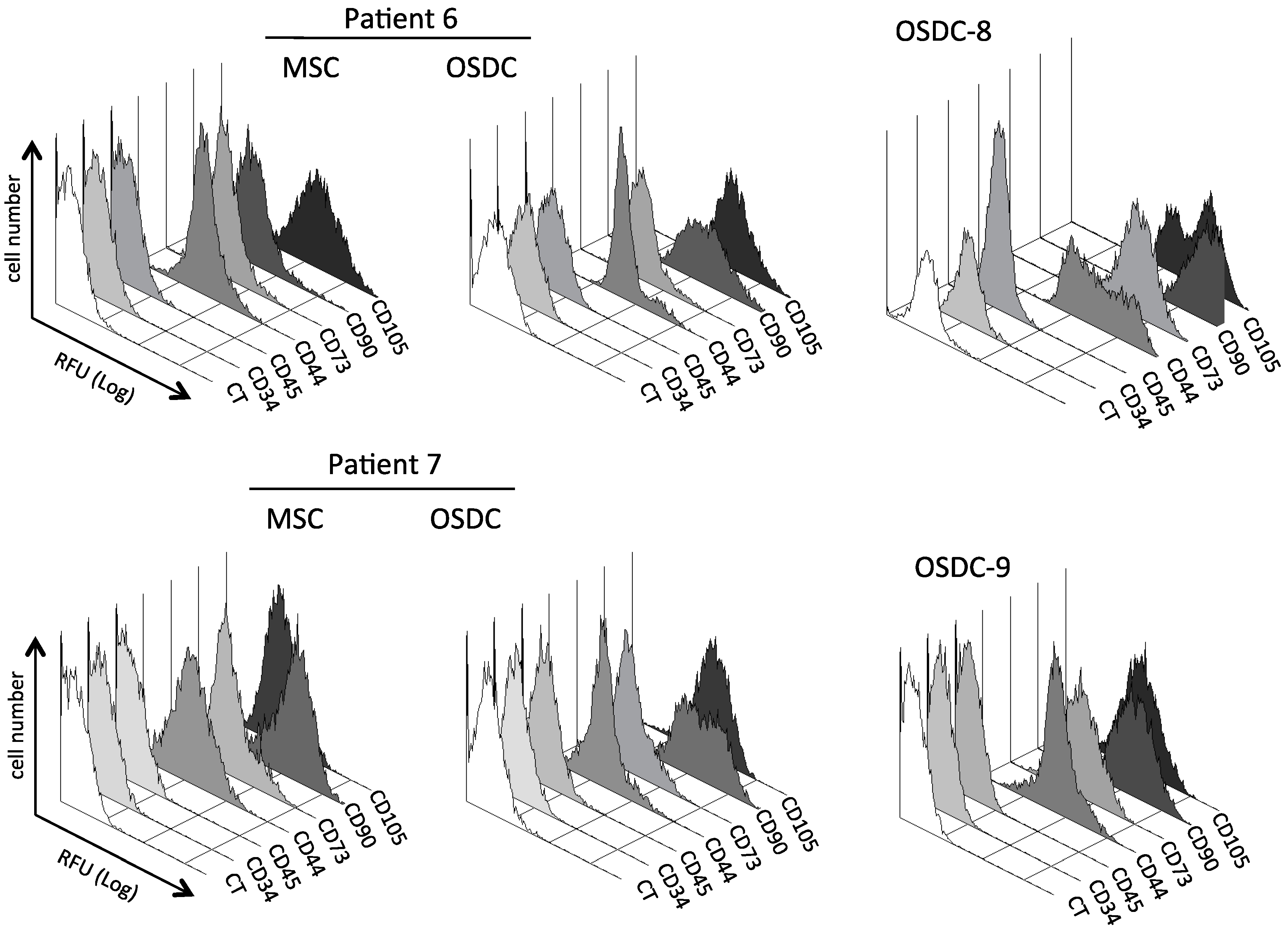
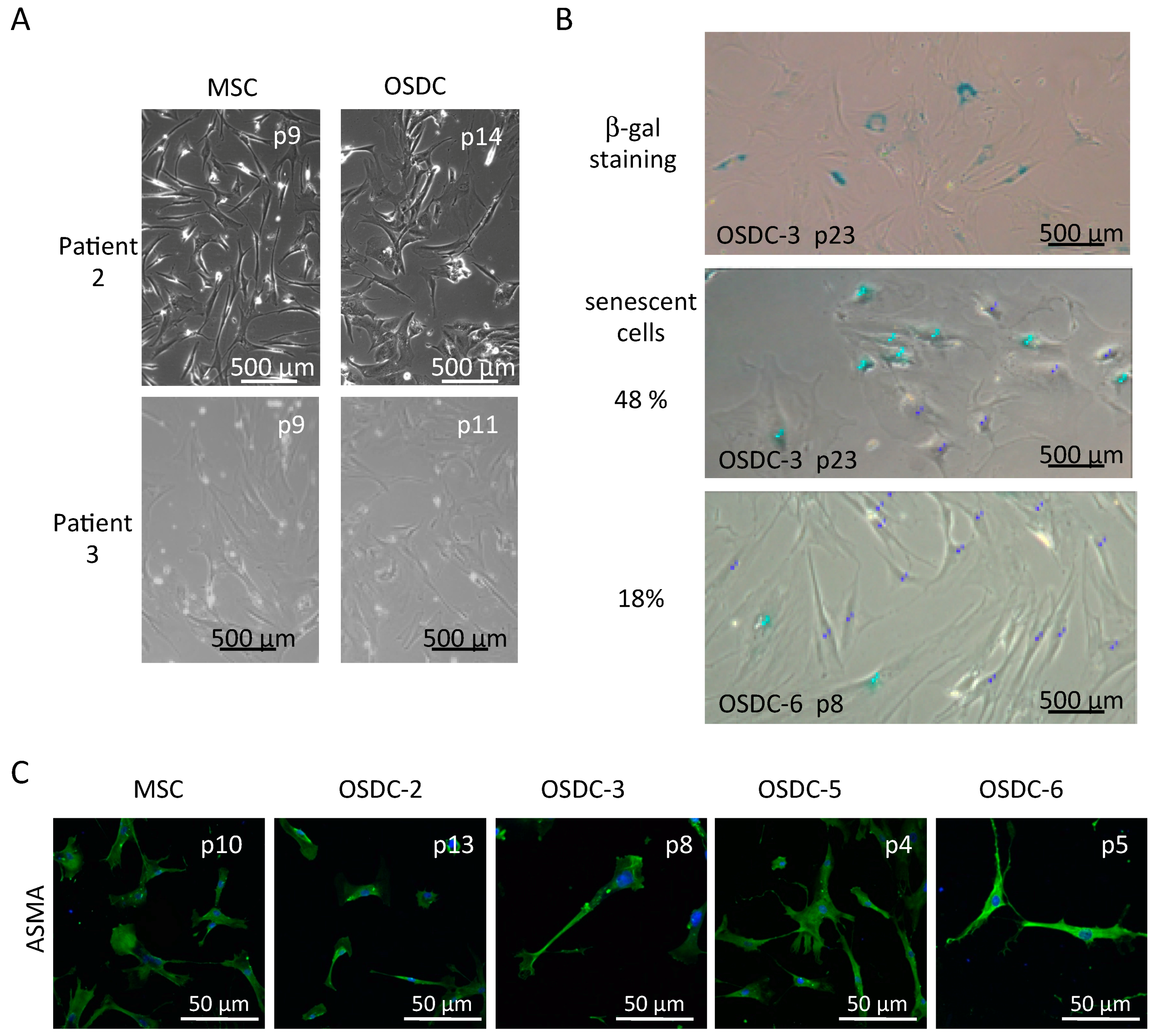
2.2. Similarities between Osteosarcoma Derived Cells (OSDC) and Mesenchymal Stromal/Stem Cells (MSC) Derived from the Bone Marrow of Either OS Patients, or Healthy Donors
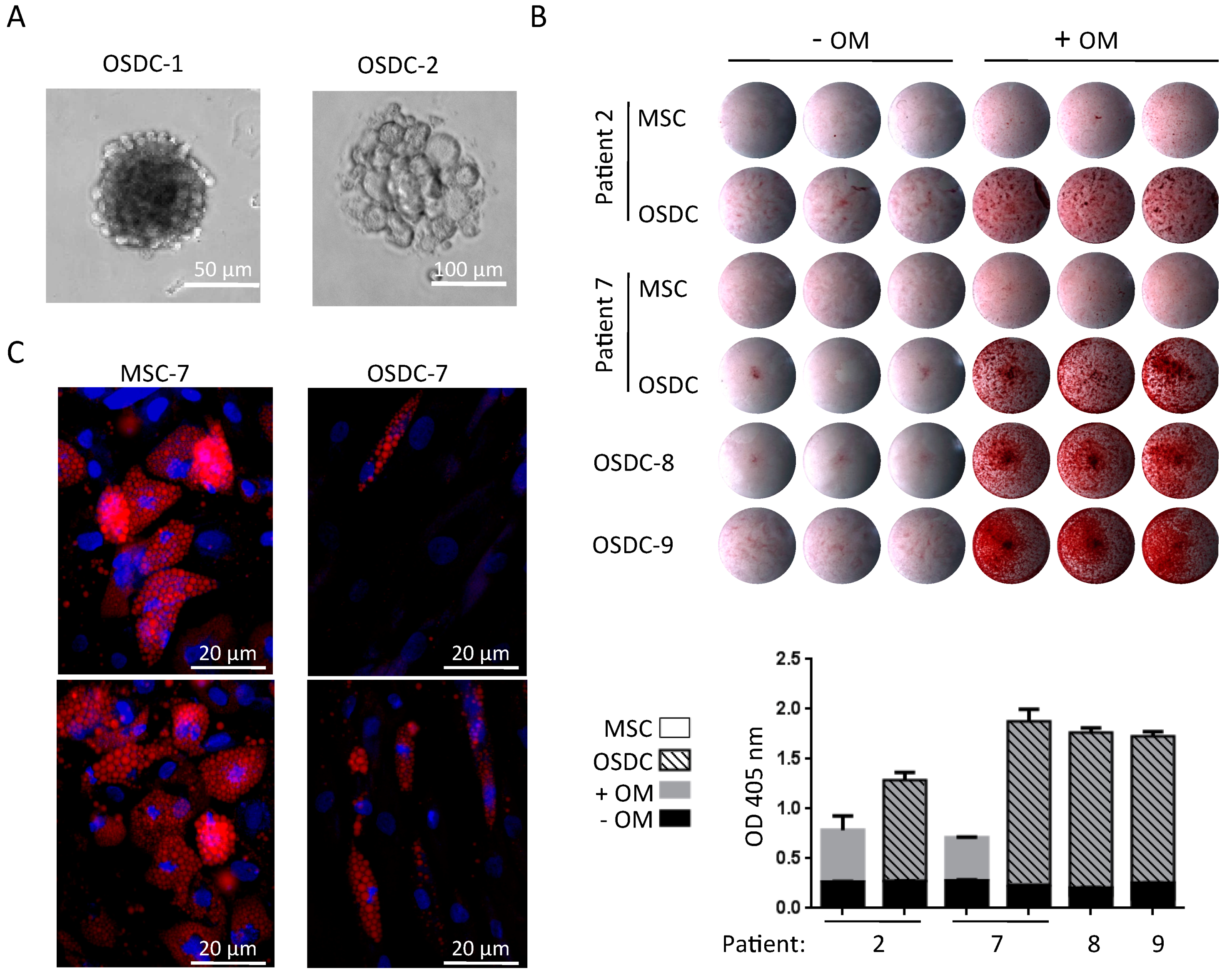
2.3. Differences in Stemness Properties between OSDC and MSC
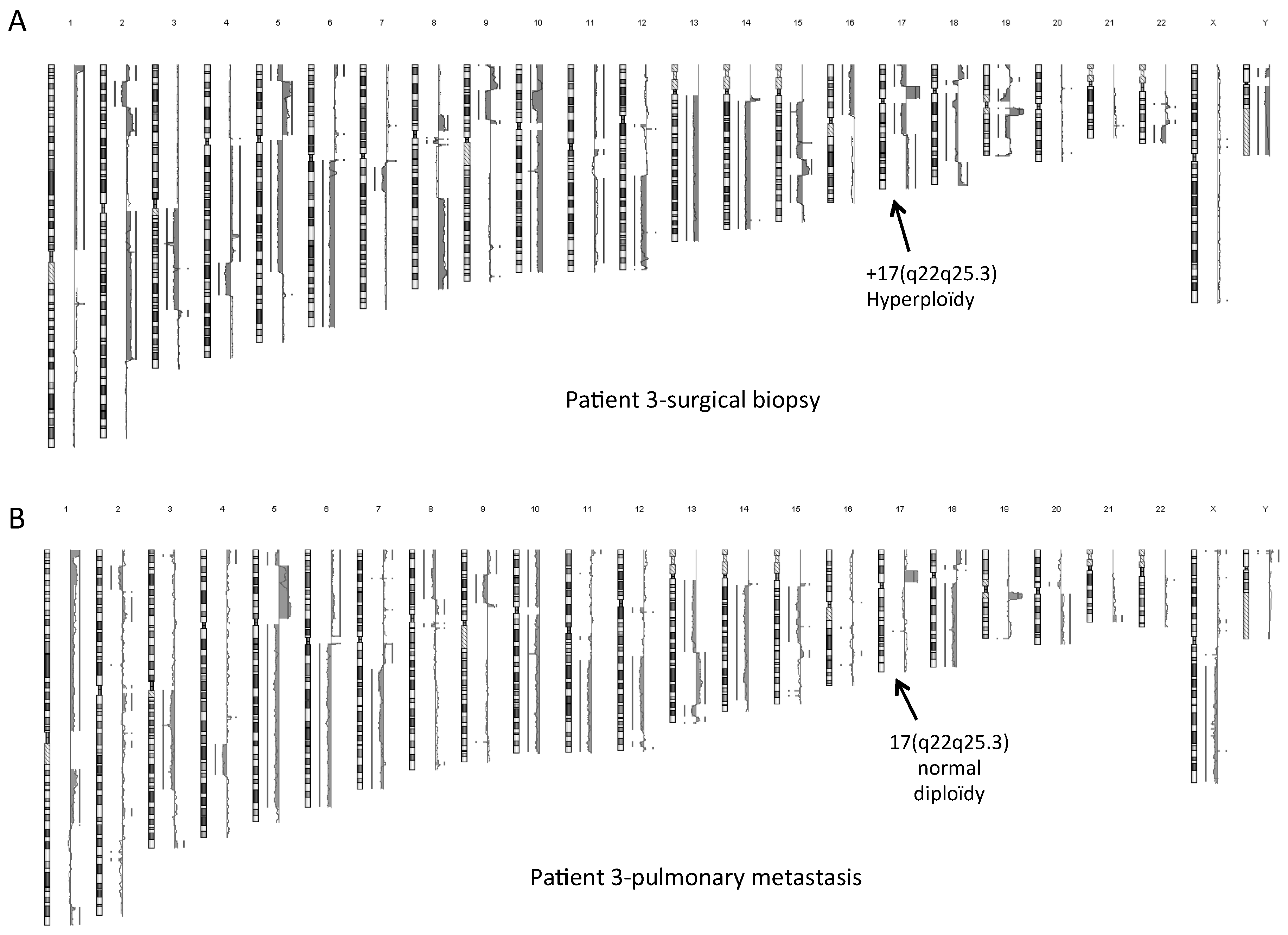
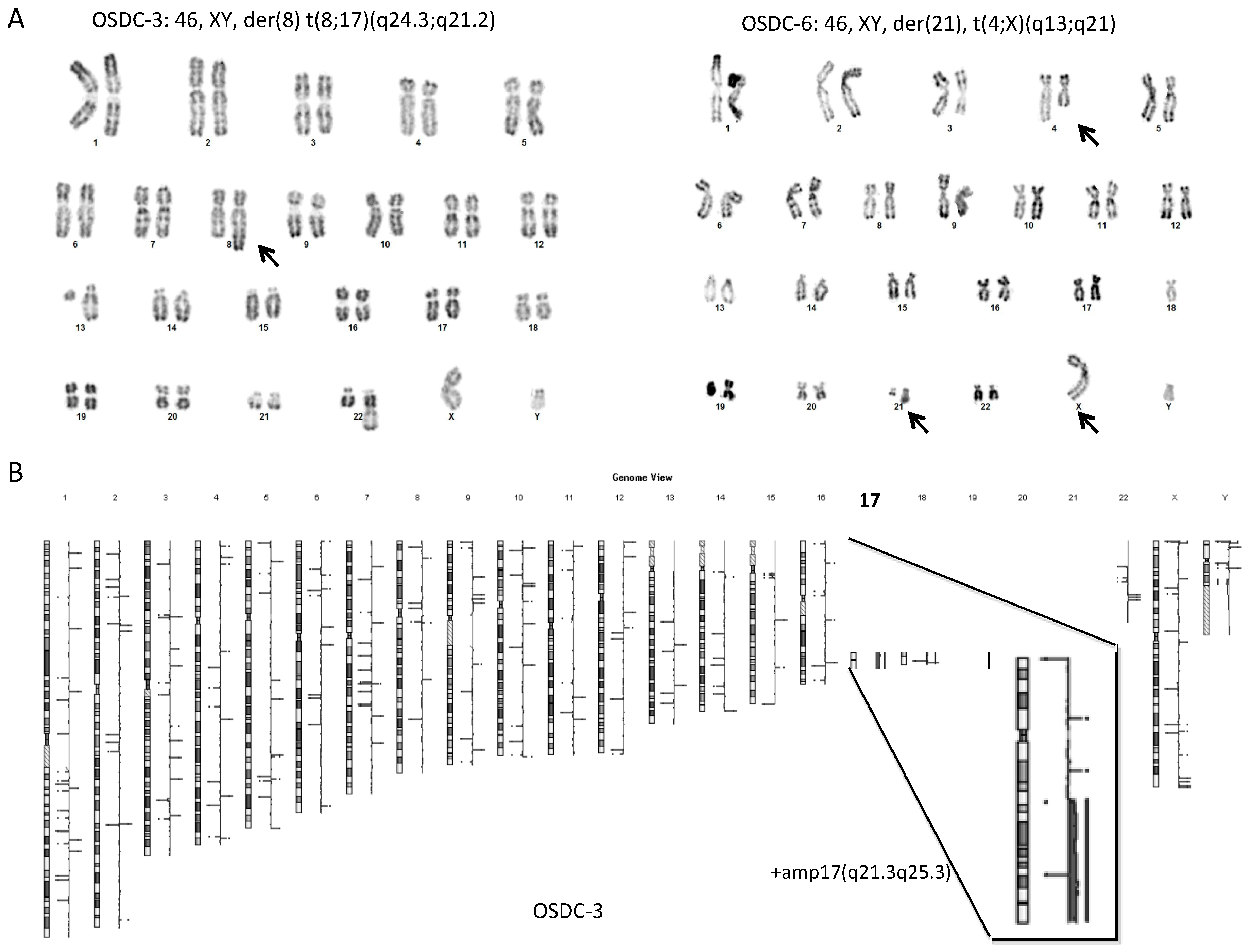
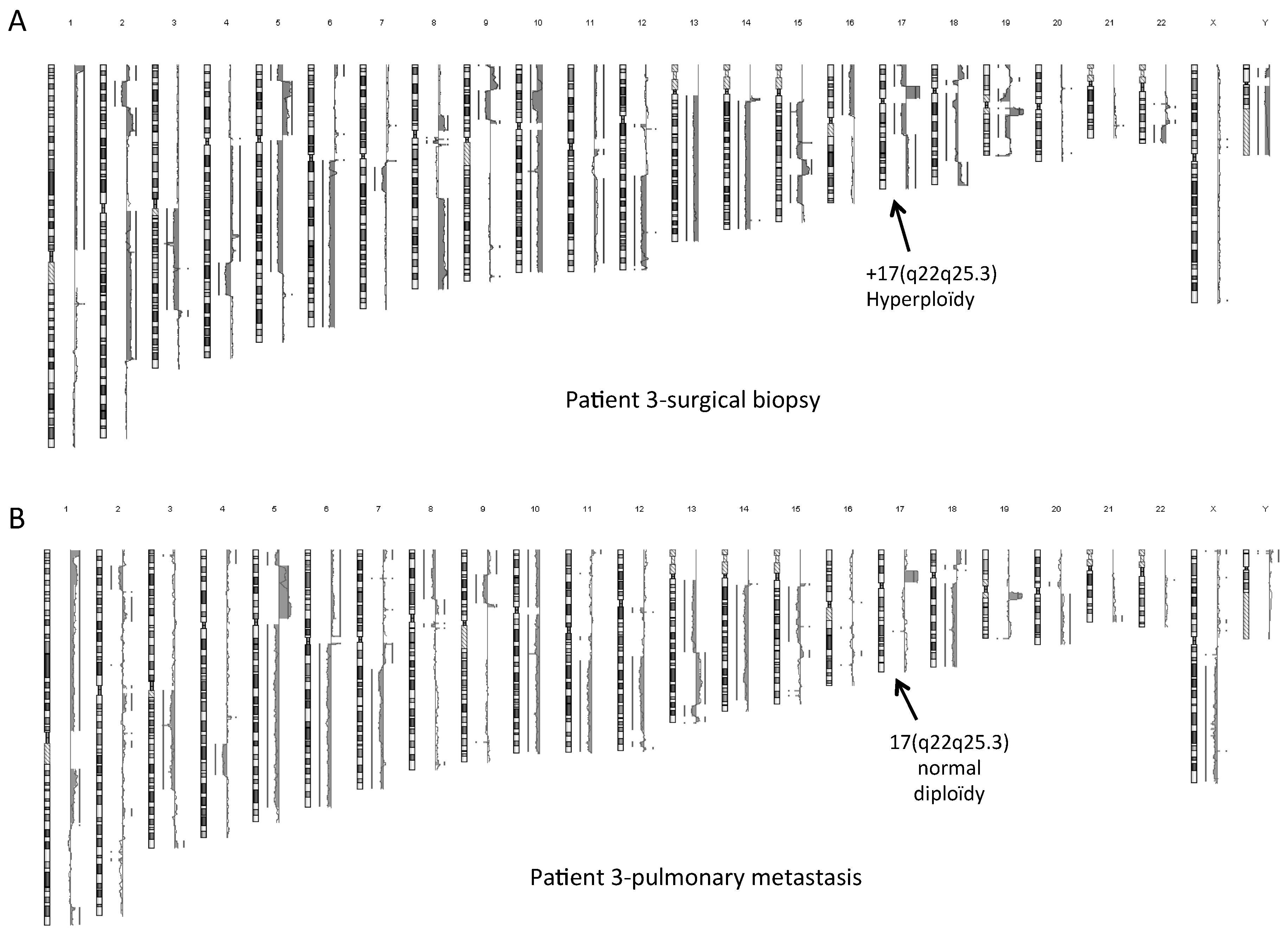
2.4. Normal Karyotype or Focused Genetic Aberrations in OSDC Compared to Primary and Secondary OS
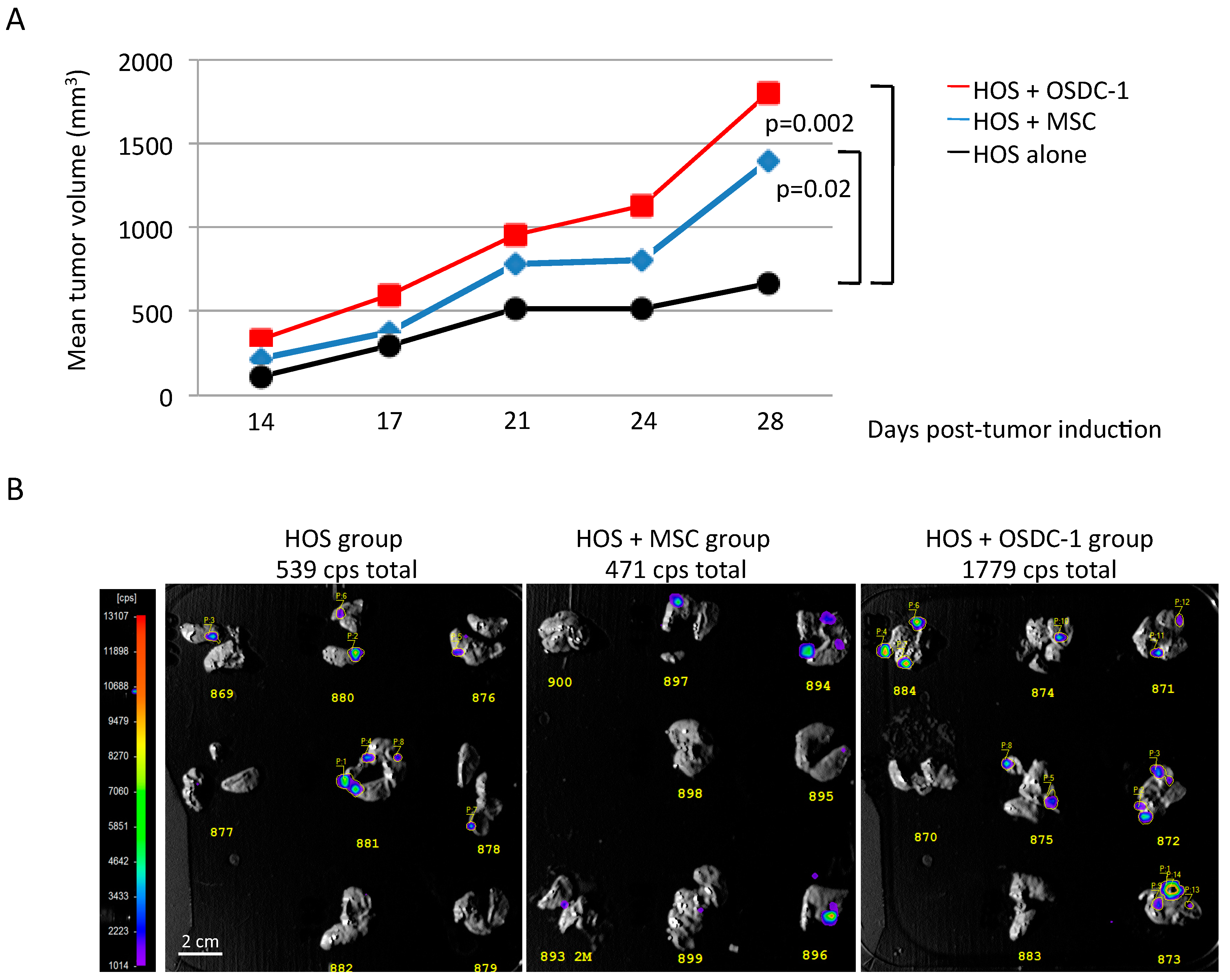
2.5. OSDC Are Not Tumorigenic but Support OS Progression
3. Discussion
4. Materials and Methods
4.1. Ethics Approval Statement
4.2. Primary Cell Cultures
4.3. Flow Cytometry Analysis
4.4. Fluorescent Microscopy Analysis
4.5. Osteogenic and Adipogenic Differentiation
4.6. Genetic Analysis
4.7. Cell Senescence Analysis
4.8. Condition Medium and Multiplex Analysis
4.9. MNNG-HOS-Induced Osteosarcoma in Nude Mice
4.10. Histology Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| OS | Osteosarcoma |
| OSDC | Osteosarcoma Derived Cells |
| MSC | Mesenchymal Stem Cells |
| CSC | Cancer Stem Cells |
| CAF | Cancer associated fibroblast |
References
- Dorfman, H.D.; Czerniak, B. Bone cancers. Cancer 1995, 75, 203–210. [Google Scholar] [CrossRef]
- Rosenberg, E.; Cleton-Jansen, A.-M.; de Pinieux, G. Conventional osteosarcoma. In WHO Classification of Tumours of Soft Tissue and Bone; Fletcher, C.D.M., Bridge, J.A., Hogendoorn, P.C.W., Mertens, F., Eds.; International Agency for Research on Cancer: Lyon, France, 2013; pp. 282–288. [Google Scholar]
- Ottaviani, G.; Jaffe, N. The epidemiology of osteosarcoma. Cancer Treat. Res. 2009, 152, 3–13. [Google Scholar] [PubMed]
- Mialou, V.; Philip, T.; Kalifa, C.; Perol, D.; Gentet, J.-C.; Marec-Berard, P.; Pacquement, H.; Chastagner, P.; Defaschelles, A.-S.; Hartmann, O. Metastatic osteosarcoma at diagnosis: Prognostic factors and long-term outcome—The French pediatric experience. Cancer 2005, 104, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Anninga, J.K.; Gelderblom, H.; Fiocco, M.; Kroep, J.R.; Taminiau, A.H.M.; Hogendoorn, P.C.W.; Egeler, R.M. Chemotherapeutic adjuvant treatment for osteosarcoma: Where do we stand? Eur. J. Cancer 2011, 47, 2431–2445. [Google Scholar] [CrossRef] [PubMed]
- Longhi, A.; Errani, C.; de Paolis, M.; Mercuri, M.; Bacci, G. Primary bone osteosarcoma in the pediatric age: State of the art. Cancer Treat. Rev. 2006, 32, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Palmerini, E. Adjuvant and neoadjuvant combination chemotherapy for osteogenic sarcoma. Curr. Opin. Oncol. 2007, 19, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.C.; Carney, S.C.; Ahlmann, E.R.; Hendifar, A.; Chawla, S.; Fedenko, A.; Angeles, C.; Menendez, L.R. A meta-analysis of osteosarcoma outcomes in the modern medical era. Sarcoma 2012, 2012, 704872. [Google Scholar] [CrossRef] [PubMed]
- Trama, A.; Botta, L.; Foschi, R.; Ferrari, A.; Stiller, C.; Desandes, E.; Maule, M.M.; Merletti, F.; Gatta, G. Survival of European adolescents and young adults diagnosed with cancer in 2000–07: Population-based data from EUROCARE-5. Lancet Oncol. 2016, 17, 896–906. [Google Scholar] [CrossRef]
- Dahlin, D.C.; Coventry, M.B. Osteogenic sarcoma. A study of six hundred cases. J. Bone Joint Surg. Am. 1967, 49, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.W.; Squire, J.A.; Zielenska, M. The Genetics of Osteosarcoma. Sarcoma 2012, 2012, 627254. [Google Scholar] [CrossRef] [PubMed]
- Gillette, J.M.; Gibbs, C.P.; Nielsen-Preiss, S.M. Establishment and characterization of OS 99-1, a cell line derived from a highly aggressive primary human osteosarcoma. In Vitro Cell. Dev. Biol. Anim. 2008, 44, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Brune, J.C.; Tormin, A.; Johansson, M.C.; Rissler, P.; Brosjö, O.; Löfvenberg, R.; von Steyern, F.V.; Mertens, F.; Rydholm, A.; Scheding, S. Mesenchymal stromal cells from primary osteosarcoma are non-malignant and strikingly similar to their bone marrow counterparts. Int. J. Cancer 2011, 129, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Arndt, C.A.; Crist, W.M. Common musculoskeletal tumors of childhood and adolescence. N. Engl. J. Med. 1999, 341, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, F.; Binh, M.B.N.; Bouvier, C.; Gomez-Brouchet, A.; Larousserie, F.; Muret, A.; de Louis-Brennetot, C.; Aurias, A.; Coindre, J.-M.; Guillou, L.; et al. MDM2 and CDK4 immunohistochemistry is a valuable tool in the differential diagnosis of low-grade osteosarcomas and other primary fibro-osseous lesions of the bone. Mod. Pathol. 2011, 24, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Forment, J.V.; Kaidi, A.; Jackson, S.P. Chromothripsis and cancer: Causes and consequences of chromosome shattering. Nat. Rev. Cancer 2012, 12, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Basu-Roy, U.; Basilico, C.; Mansukhani, A. Perspectives on cancer stem cells in osteosarcoma. Cancer Lett. 2013, 338, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Abarrategi, A.; Tornin, J.; Martinez-Cruzado, L.; Hamilton, A.; Martinez-Campos, E.; Rodrigo, J.P.; González, M.V.; Baldini, N.; Garcia-Castro, J.; Rodriguez, R.; et al. Osteosarcoma: Cells-of-Origin, Cancer Stem Cells, and Targeted Therapies. Stem Cells Int. 2016, 2016, 3631764. [Google Scholar] [CrossRef] [PubMed]
- Cleton-Jansen, A.-M.; Anninga, J.K.; Briaire de Bruijn, I.H.; Romeo, S.; Oosting, J.; Egeler, R.M.; Gelderblom, H.; Taminiau, A.H.M.; Hogendoorn, P.C.W.; et al. Profiling of high-grade central osteosarcoma and its putative progenitor cells identifies tumourigenic pathways. Br. J. Cancer 2009, 101, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Deschaseaux, F.; Pontikoglou, C.; Sensébé, L. Bone regeneration: The stem/progenitor cells point of view. J. Cell. Mol. Med. 2010, 14, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Mohseny, A.B.; Hogendoorn, P.C.W.; Cleton-Jansen, A.-M. Mesenchymal stem cell transformation and sarcoma genesis. Clin. Sarcoma Res. 2013, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Wu, P.-K.; Chen, P.C.-H.; Lee, C.-W.; Chen, W.-M.; Hung, S.-C. Generation of Osteosarcomas from a Combination of Rb Silencing and c-Myc Overexpression in Human Mesenchymal Stem Cells. Stem Cells Transl. Med. 2017, 6, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Skoda, J.; Nunukova, A.; Loja, T.; Zambo, I.; Neradil, J.; Mudry, P.; Zitterbart, K.; Hermanova, M.; Hampl, A.; Sterba, J.; et al. Cancer stem cell markers in pediatric sarcomas: Sox2 is associated with tumorigenicity in immunodeficient mice. Tumor Biol. 2016, 37, 9535–9548. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, C.P.; Kukekov, V.G.; Reith, J.D.; Tchigrinova, O.; Suslov, O.N.; Scott, E.W.; Ghivizzani, S.C.; Ignatova, T.N.; Steindler, D.A.; et al. Stem-Like Cells in Bone Sarcomas: Implications for Tumorigenesis. Neoplasia 2005, 7, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.S.; Agarwal, N.; Wood, B.M.; Porretta, C.; Ruiz, B.; Pochampally, R.R.; Iwakuma, T. CD117 and Stro-1 Identify Osteosarcoma Tumor-Initiating Cells Associated with Metastasis and Drug Resistance. Cancer Res. 2010, 70, 4602–4612. [Google Scholar] [CrossRef] [PubMed]
- Buchsbaum, R.J.; Oh, S.Y. Breast Cancer-Associated Fibroblasts: Where We Are and Where We Need to Go. Cancers 2016, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Melchor, L.; Alvarez, S.; Honrado, E.; Palacios, J.; Barroso, A.; Díez, O.; Osorio, A.; Benítez, J. The accumulation of specific amplifications characterizes two different genomic pathways of evolution of familial breast tumors. Clin. Cancer Res. 2005, 11 Pt 1, 8577–8584. [Google Scholar] [CrossRef] [PubMed]
- Pandita, A.; Zielenska, M.; Thorner, P.; Bayani, J.; Godbout, R.; Greenberg, M.; Squire, J.A. Application of comparative genomic hybridization, spectral karyotyping, and microarray analysis in the identification of subtype-specific patterns of genomic changes in rhabdomyosarcoma. Neoplasia 1999, 1, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, M.; Boulware, K.S.; Wood, R.D. The ERCC1 and ERCC4 (XPF) genes and gene products. Gene 2015, 569, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.H.; Lim, W.K.; Ishak, N.D.B.; Li, S.-T.; Goh, W.L.; Tan, G.S.; Lim, K.H.; Teo, M.; Young, C.N.C.; Malik, S.; et al. Germline Mutations in Cancer Predisposition Genes are Frequent in Sporadic Sarcomas. Sci. Rep. 2017, 7, 10660. [Google Scholar] [CrossRef] [PubMed]
- Bayani, J.; Zielenska, M.; Pandita, A.; Al-Romaih, K.; Karaskova, J.; Harrison, K.; Bridge, J.A.; Sorensen, P.; Thorner, P.; Squire, J.A.; et al. Spectral karyotyping identifies recurrent complex rearrangements of chromosomes 8, 17, and 20 in osteosarcomas. Genes. Chromosomes Cancer 2003, 36, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Avril, P.; Le Nail, L.-R.; Brennan, M.Á.; Rosset, P.; De Pinieux, G.; Layrolle, P.; Heymann, D.; Perrot, P.; Trichet, V. Mesenchymal stem cells increase proliferation but do not change quiescent state of osteosarcoma cells: Potential implications according to the tumor resection status. J. Bone Oncol. 2016, 5, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, J.; Escriou, V.; Perrot, P.; Picarda, G.; Charrier, C.; Scherman, D.; Heymann, D.; Rédini, F.; Trichet, V. Advantages of bioluminescence imaging to follow siRNA or chemotherapeutic treatments in osteosarcoma preclinical models. Cancer Gene Ther. 2010, 17, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Perrot, P.; Rousseau, J.; Bouffaut, A.-L.; Rédini, F.; Cassagnau, E.; Deschaseaux, F.; Heymann, M.-F.; Heymann, D.; Duteille, F.; Trichet, V.; et al. Safety Concern between Autologous Fat Graft, Mesenchymal Stem Cell and Osteosarcoma Recurrence. PLoS ONE 2010, 5, e10999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brullé, L.; Vandamme, M.; Riès, D.; Martel, E.; Robert, E.; Lerondel, S.; Trichet, V.; Richard, S.; Pouvesle, J.-M.; Le Pape, A. Effects of a Non Thermal Plasma Treatment Alone or in Combination with Gemcitabine in a MIA PaCa2-luc Orthotopic Pancreatic Carcinoma Model. PLoS ONE 2012, 7, e52653. [Google Scholar] [CrossRef] [PubMed]
- Hauben, E.I.; Weeden, S.; Pringle, J.; van Marck, E.A.; Hogendoorn, P.C.W. Does the histological subtype of high-grade central osteosarcoma influence the response to treatment with chemotherapy and does it affect overall survival? A study on 570 patients of two consecutive trials of the European Osteosarcoma Intergroup. Eur. J. Cancer 2002, 38, 1218–1225. [Google Scholar] [CrossRef]
- Billiau, A.; Edy, V.G.; Heremans, H.; Van Damme, J.; Desmyter, J.; Georgiades, J.A.; De Somer, P. Human interferon: Mass production in a newly established cell line, MG-63. Antimicrob. Agents Chemother. 1977, 12, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Yousefzadeh, M.J.; Faridounnia, M.; Chong, J.X.; Hisama, F.M.; Hudgins, L.; Mercado, G.; Wade, E.A.; Barghouthy, A.S.; Lee, L.; et al. ERCC4 variants identified in a cohort of patients with segmental progeroid syndromes. Hum. Mutat. 2018, 39, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Kovac, M.; Blattmann, C.; Ribi, S.; Smida, J.; Mueller, N.S.; Engert, F.; Castro-Giner, F.; Weischenfeldt, J.; Kovacova, M.; Krieg, A.; et al. Exome sequencing of osteosarcoma reveals mutation signatures reminiscent of BRCA deficiency. Nat. Commun. 2015, 6, 8940. [Google Scholar] [CrossRef] [PubMed]
- Overholtzer, M.; Rao, P.H.; Favis, R.; Lu, X.-Y.; Elowitz, M.B.; Barany, F.; Ladanyi, M.; Gorlick, R.; Levine, A.J. The presence of p53 mutations in human osteosarcomas correlates with high levels of genomic instability. Proc. Natl. Acad. Sci. USA 2003, 100, 11547–11552. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Rao, P.H.; Favis, R.; Lu, X.-Y.; Elowitz, M.B.; Barany, F.; Ladanyi, M.; Gorlick, R.; Levine, A.J.; et al. Germline TP53 variants and susceptibility to osteosarcoma. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef] [PubMed]
- Hochane, M.; Trichet, V.; Pecqueur, C.; Avril, P.; Oliver, L.; Denis, J.; Brion, R.; Amiaud, J.; Pineau, A.; Naveilhan, P.; et al. Low-Dose Pesticide Mixture Induces Senescence in Normal Mesenchymal Stem Cells (MSC) and Promotes Tumorigenic Phenotype in Premalignant MSC. Stem Cells 2017, 35, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Liu, Y.; Akiyama, K.; Chen, C.; Qu, C.; Jin, Y.; Shi, S. IFN-γ and TNF-α Synergistically Induce Mesenchymal Stem Cell Impairment and Tumorigenesis via NFκB Signaling. Stem Cells 2013, 31, 1383–1395. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.S.; Park, D.K.; Arnstein, P.; Huebner, R.J.; Weisburger, E.K.; Nelson-Rees, W.A. Transformation of human cells in culture by N-methyl-N′-nitro-N-nitrosoguanidine. Nature 1975, 256, 751–753. [Google Scholar] [CrossRef] [PubMed]
- Pantic, I. Cancer stem cell hypotheses: Impact on modern molecular physiology and pharmacology research. J. Biosci. 2011, 36, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.Á.; Renaud, A.; Amiaud, J.; Rojewski, M.T.; Schrezenmeier, H.; Heymann, D.; Trichet, V.; Layrolle, P. Pre-clinical studies of bone regeneration with human bone marrow stromal cells and biphasic calcium phosphate. Stem Cell Res. Ther. 2014, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- Correa, D.; Somoza, R.A.; Lin, P.; Schiemann, W.P.; Caplan, A.I. Mesenchymal stem cells regulate melanoma cancer cells extravasation to bone and liver at their perivascular niche: MSC/pericytes regulate bone and liver melanoma invasion. Int. J. Cancer 2016, 138, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Mesenchymal Stem Cells. Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Bian, Z.; Fan, Q.; Li, G.; Tang, T. Human mesenchymal stem cells (hMSCs) target osteosarcoma and promote its growth and pulmonary metastasis. Cancer Lett. 2009, 281, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, E.L.; Dembinski, J.L.; Sasser, A.K.; Watson, K.; Klopp, A.; Hall, B.; Andreeff, M.; Marini, F. Mesenchymal Stem Cell Transition to Tumor-Associated Fibroblasts Contributes to Fibrovascular Network Expansion and Tumor Progression. PLoS ONE 2009, 4, e4992. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Saidou, J.; Watabe, K. Cancer associated fibroblasts (CAFs) in tumor microenvironment. Front. Biosci. Landmark Ed. 2010, 15, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Blau, O.; Baldus, C.D.; Hofmann, W.-K.; Thiel, G.; Nolte, F.; Burmeister, T.; Türkmen, S.; Benlasfer, O.; Schümann, E.; Sindram, A.; et al. Mesenchymal stromal cells of myelodysplastic syndrome and acute myeloid leukemia patients have distinct genetic abnormalities compared with leukemic blasts. Blood 2011, 118, 5583–5592. [Google Scholar] [CrossRef] [PubMed]
- Kurose, K.; Gilley, K.; Matsumoto, S.; Watson, P.H.; Zhou, X.-P.; Eng, C. Frequent somatic mutations in PTEN and TP53 are mutually exclusive in the stroma of breast carcinomas. Nat. Genet. 2002, 32, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Marycz, K.; Tomaszewski, K.A.; Marędziak, M.; Śmieszek, A. The Effect of Age on Osteogenic and Adipogenic Differentiation Potential of Human Adipose Derived Stromal Stem Cells (hASCs) and the Impact of Stress Factors in the Course of the Differentiation Process. Oxid. Med. Cell. Longev. 2015, 2015, 309169. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Brouchet, A.; Illac, C.; Gilhodes, J.; Bouvier, C.; Aubert, S.; Guinebretiere, J.-M.; Marie, B.; Larousserie, F.; Entz-Werlé, N.; de Pinieux, G.; et al. CD163-positive tumor-associated macrophages and CD8-positive cytotoxic lymphocytes are powerful diagnostic markers for the therapeutic stratification of osteosarcoma patients: An immunohistochemical analysis of the biopsies fromthe French OS2006 phase 3 trial. OncoImmunology 2017, 6, e1331193. [Google Scholar] [PubMed]
- Abumaree, M.; al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive Properties of Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2012, 8, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Studeny, M.; Marini, F.C.; Dembinski, J.L.; Zompetta, C.; Cabreira-Hansen, M.; Bekele, B.N.; Champlin, R.E.; Andreeff, M. Mesenchymal Stem Cells: Potential Precursors for Tumor Stroma and Targeted-Delivery Vehicles for Anticancer Agents. JNCI J. Natl. Cancer Inst. 2004, 96, 1593–1603. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Gong, C.; Peng, H.; Fang, H.; Zhou, J.; Li, J.; Chen, S.; Zheng, H. High CCL5 expression is associated with osteosarcoma metastasis and poor prognosis of patients with osteosarcoma. Mol. Med. Rep. 2017, 16, 6953–6957. [Google Scholar] [CrossRef] [PubMed]
- Aggoune, D.; Sorel, N.; Bonnet, M.-L.; Goujon, J.-M.; Tarte, K.; Hérault, O.; Domenech, J.; Réa, D.; Legros, L.; Johnson-Ansa, H. Bone marrow mesenchymal stromal cell (MSC) gene profiling in chronic myeloid leukemia (CML) patients at diagnosis and in deep molecular response induced by tyrosine kinase inhibitors (TKIs). Leuk. Res. 2017, 60, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.Á.; Renaud, A.; Gamblin, A.; D’Arros, C.; Nedellec, S.; Trichet, V.; Layrolle, P. 3D cell culture and osteogenic differentiation of human bone marrow stromal cells plated onto jet-sprayed or electrospun micro-fiber scaffolds. Biomed. Mater. 2015, 10, 45019. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Year of diagnosis | 2007 | 2010 | 2010 | 2014 | 2014 | 2015 | 2015 | 2015 | 2015 |
| Age | 21 | 18 | 16 | 23 | 30 | 36 | 18 | 14 | 14 |
| Gender | F | M | M | M | F | M | M | M | M |
| Histological subtype | F | OB, CH, F | OB | OB | CH | OB, CH | OB, CH, F | OB, T | OB |
| Tumor site | proximal humerus | Distal femur | Distal femur | Distal tibia | pelvis | Distal femur | proximal tibia | proximal fibula | proximal fibula |
| Limb salvage | Yes | No | Yes | No | Yes | Yes | Yes | Yes | Yes |
| Metastasis at diagnosis | Yes | No | No | No | No | No | Yes | No | n.d. |
| Response to NAPC | GR | PR | GR | PR | PR | GR | GR | PR | PR |
| % residual tumor cells | 6% | 55% | 7% | 20% | 30% | 8% | <1% | 11% | 52% |
| Local recurrence | No | No | No | No | No | No | No | No | No |
| Metastasis progression | Yes | Yes | Yes | Yes | Yes | No | No | No | No |
| site | lung | lung | lung | lung | lung | ||||
| Outcomes | DOD | DOD | AWD | DOD | AWD | NED | NED | NED | NED |
| Patient’s OSDC | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Time of tumor sample collection for primary cell lines | post-NAPC | post-NAPC | post-NAPC | post-NAPC | pre-NAPC | post-NAPC | post-NAPC | post-NAPC | post-NAPC |
| Sample storage before dissociation | in culture medium P/S at 4 °C for 12 h | in SVF with 10% DMSO at −80 °C for 1 to 4 weeks | in MACS Tissue Storage Solution at 4 °C for 24 to 72 h | ||||||
| Enzymatic dissociation | No | in Collagenase I (InVitrogen) during 1 to 4 h at 37 °C | in MACS human tumor dissociation kit (Miltenyi Biotech) for 1 h at 37 °C | ||||||
| Sphere rate (10−5) | n.d. | 65 | 0 | 3.3 | 0.67 | 5.5 | 15.8 | n.d. | 47 |
| Patient’s MSC | n.a. | 2 | 3 | 4 | 5 | 6 | 7 | n.a. | n.a. |
| Time of BM aspirates | n.a. | pre-NAPC | post-NAPC | post-NAPC | pre-NAPC | pre-NAPC | pre-NAPC | n.a. | n.a. |
| Sphere rate (10−5) | n.a. | 0 | 0.13 | n.a. | n.a. | 0 | 0 | n.a. | n.a. |
| 9 AD-OSDC | 6 AD-MSC | 6 SP-OSDC | ||||
|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | |
| CD34 | 1 | 0 | 1 | 0 | 2 | 1 |
| CD45 | 1 | 0 | 1 | 1 | 1 | 0 |
| CD44 | 128 | 99 | 144 | 100 | 210 | 34 |
| CD73 | 44 | 32 | 62 | 41 | 91 | 17 |
| CD90 | 274 | 224 | 208 | 164 | 474 | 65 |
| CD105 | 81 | 35 | 86 | 56 | 51 | 14 |
| Patient’s OSDC | 1 | 2 | 3 | 4 | 5 | 7 | 9 |
|---|---|---|---|---|---|---|---|
| HOS alone | 5/8 | 2/8 | 2/8 | 5/8 | 5/8 | 6/8 | 1/8 |
| HOS + OSDC | 6/8 | 5/8 | 5/8 | 7/8 | 7/8 | 6/8 | 2/8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Nail, L.-R.; Brennan, M.; Rosset, P.; Deschaseaux, F.; Piloquet, P.; Pichon, O.; Le Caignec, C.; Crenn, V.; Layrolle, P.; Hérault, O.; et al. Comparison of Tumor- and Bone Marrow-Derived Mesenchymal Stromal/Stem Cells from Patients with High-Grade Osteosarcoma. Int. J. Mol. Sci. 2018, 19, 707. https://doi.org/10.3390/ijms19030707
Le Nail L-R, Brennan M, Rosset P, Deschaseaux F, Piloquet P, Pichon O, Le Caignec C, Crenn V, Layrolle P, Hérault O, et al. Comparison of Tumor- and Bone Marrow-Derived Mesenchymal Stromal/Stem Cells from Patients with High-Grade Osteosarcoma. International Journal of Molecular Sciences. 2018; 19(3):707. https://doi.org/10.3390/ijms19030707
Chicago/Turabian StyleLe Nail, Louis-Romée, Meadhbh Brennan, Philippe Rosset, Frédéric Deschaseaux, Philippe Piloquet, Olivier Pichon, Cédric Le Caignec, Vincent Crenn, Pierre Layrolle, Olivier Hérault, and et al. 2018. "Comparison of Tumor- and Bone Marrow-Derived Mesenchymal Stromal/Stem Cells from Patients with High-Grade Osteosarcoma" International Journal of Molecular Sciences 19, no. 3: 707. https://doi.org/10.3390/ijms19030707