Fatty Acid β-Oxidation Is Essential in Leptin-Mediated Oocytes Maturation of Yellow Catfish Pelteobagrus fulvidraco


Abstract
:
1. Introduction
2. Results
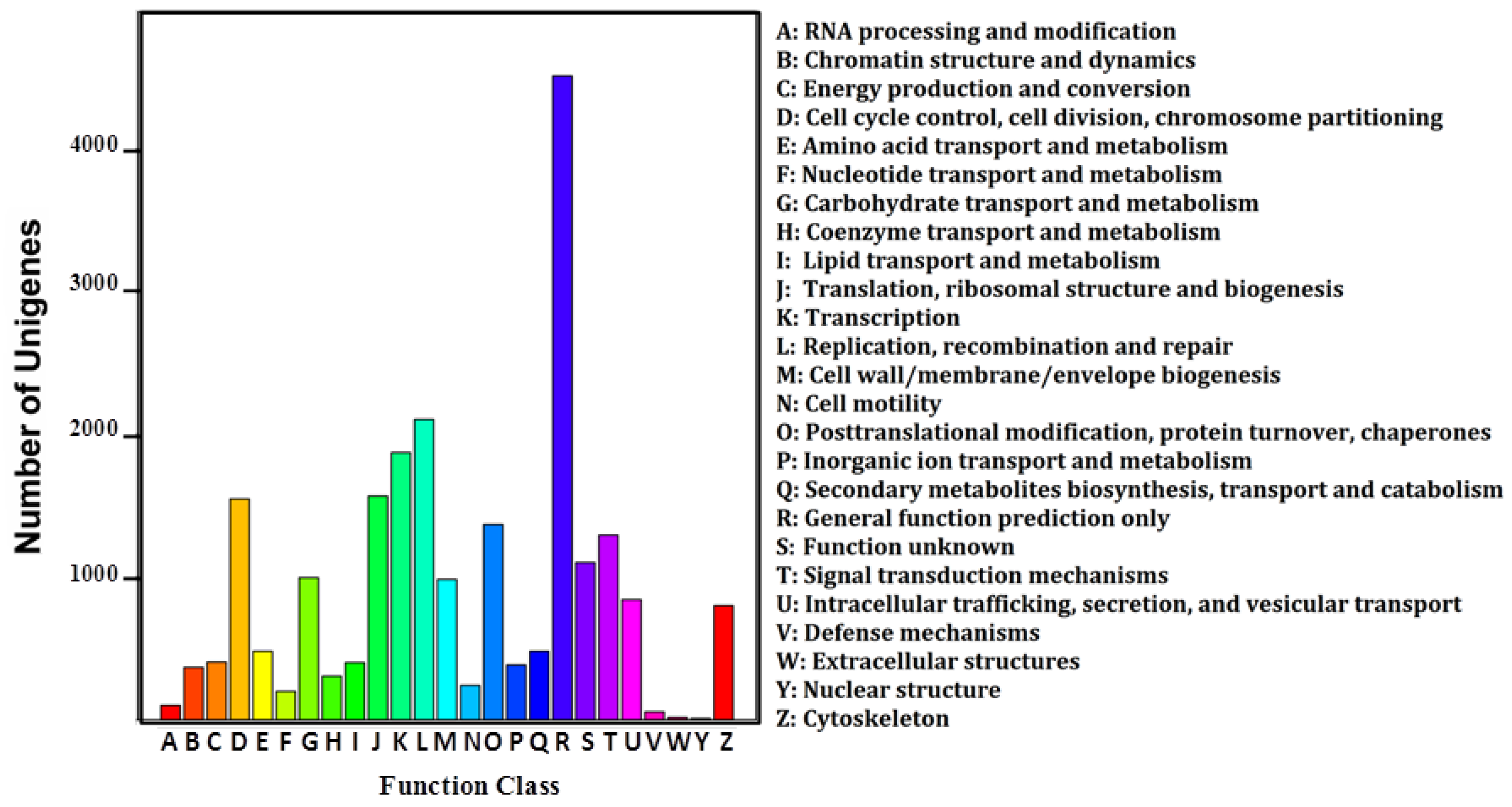
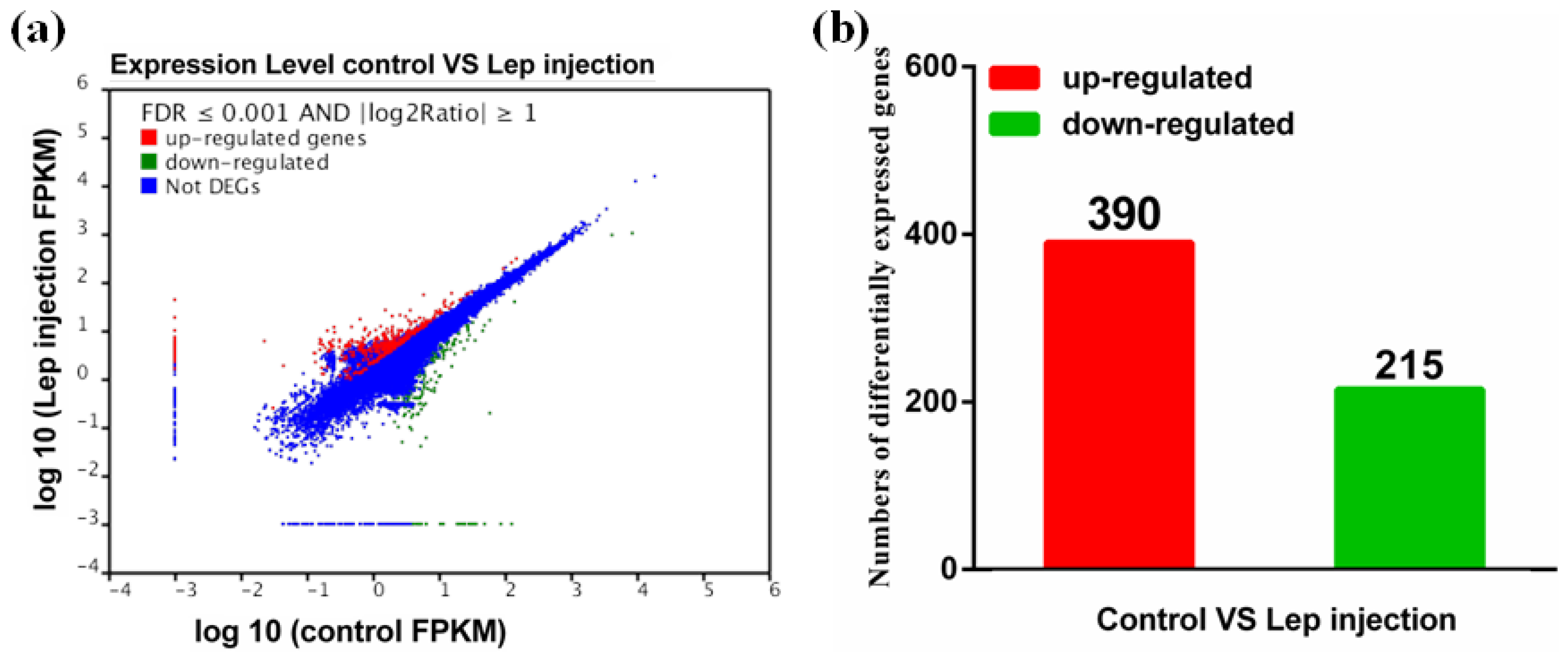
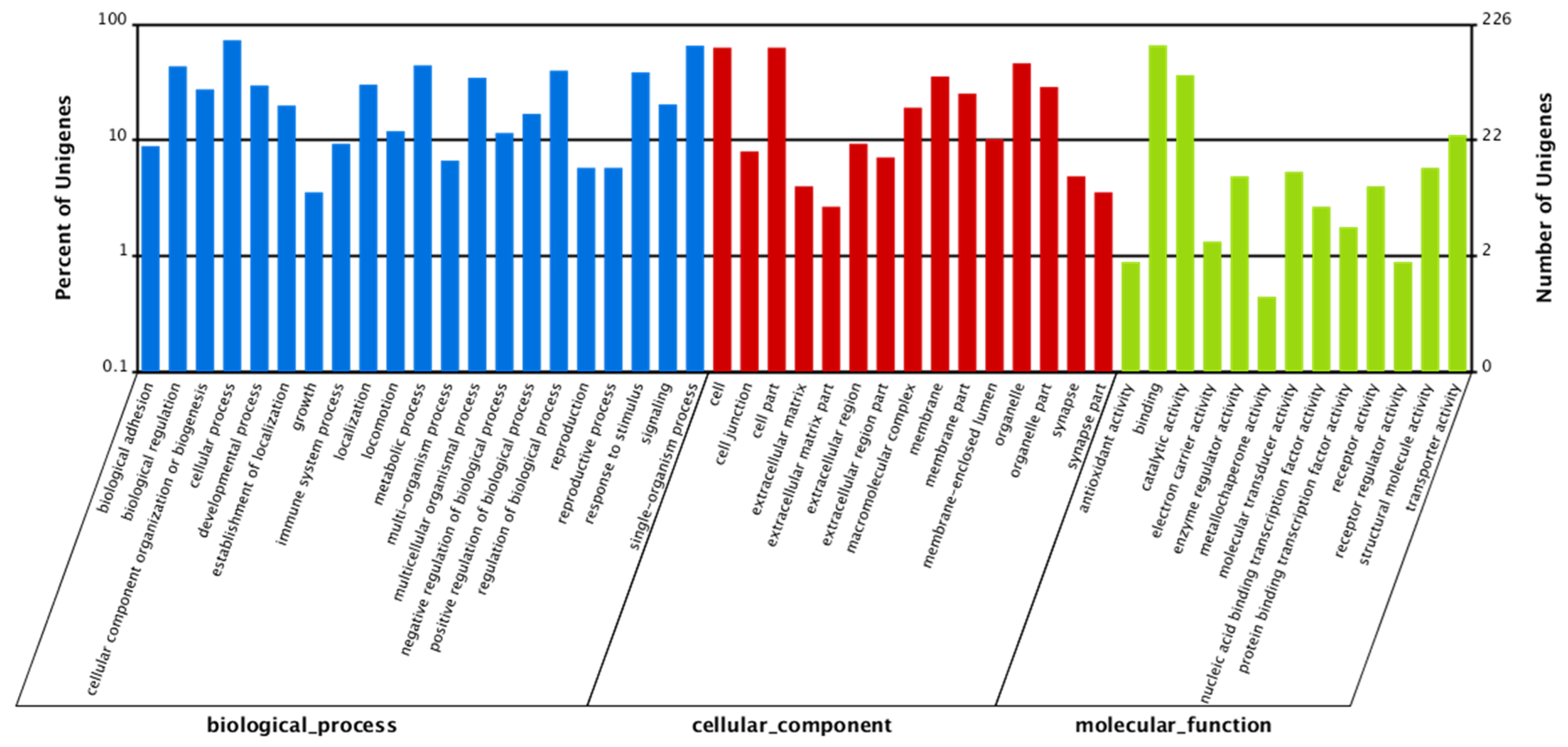
2.1. Ovary Transcriptome of P. fulvidraco Generated by RNA-Seq
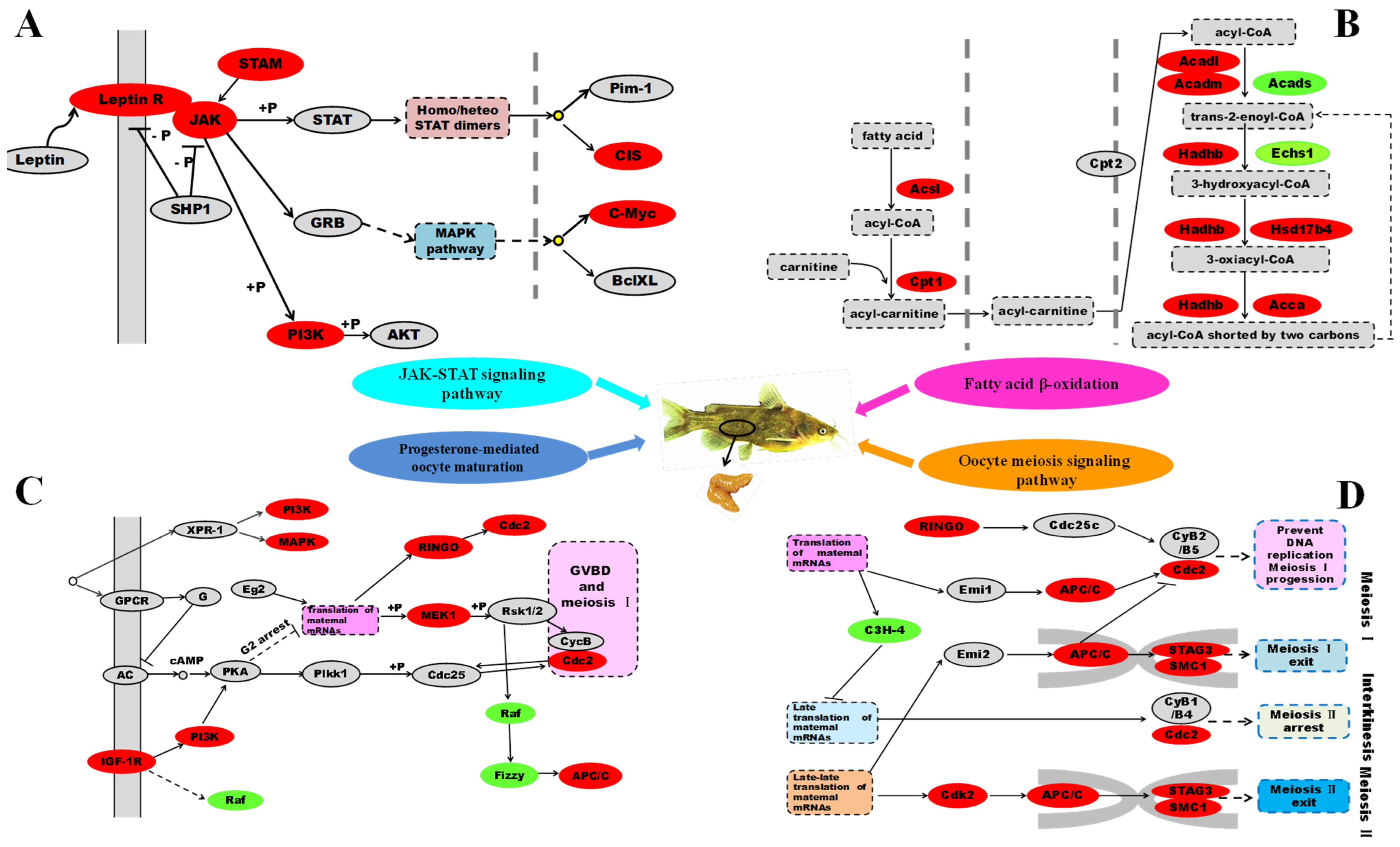
2.2. Leptin Is a Very Important Factor That Links Fatty Acid β-Oxidation with Oocytes Maturation
2.2.1. Differentially Expressed Genes Involved in Signal Transduction
2.2.2. Differentially Expressed Genes Involved in Fatty Acid β-Oxidation
2.2.3. Differentially Expressed Genes Involved in Oocytes Maturation
2.2.4. Differentially Expressed Genes Involved in Other Pathways
2.2.5. Validation of Differential Gene Expression by qPCR
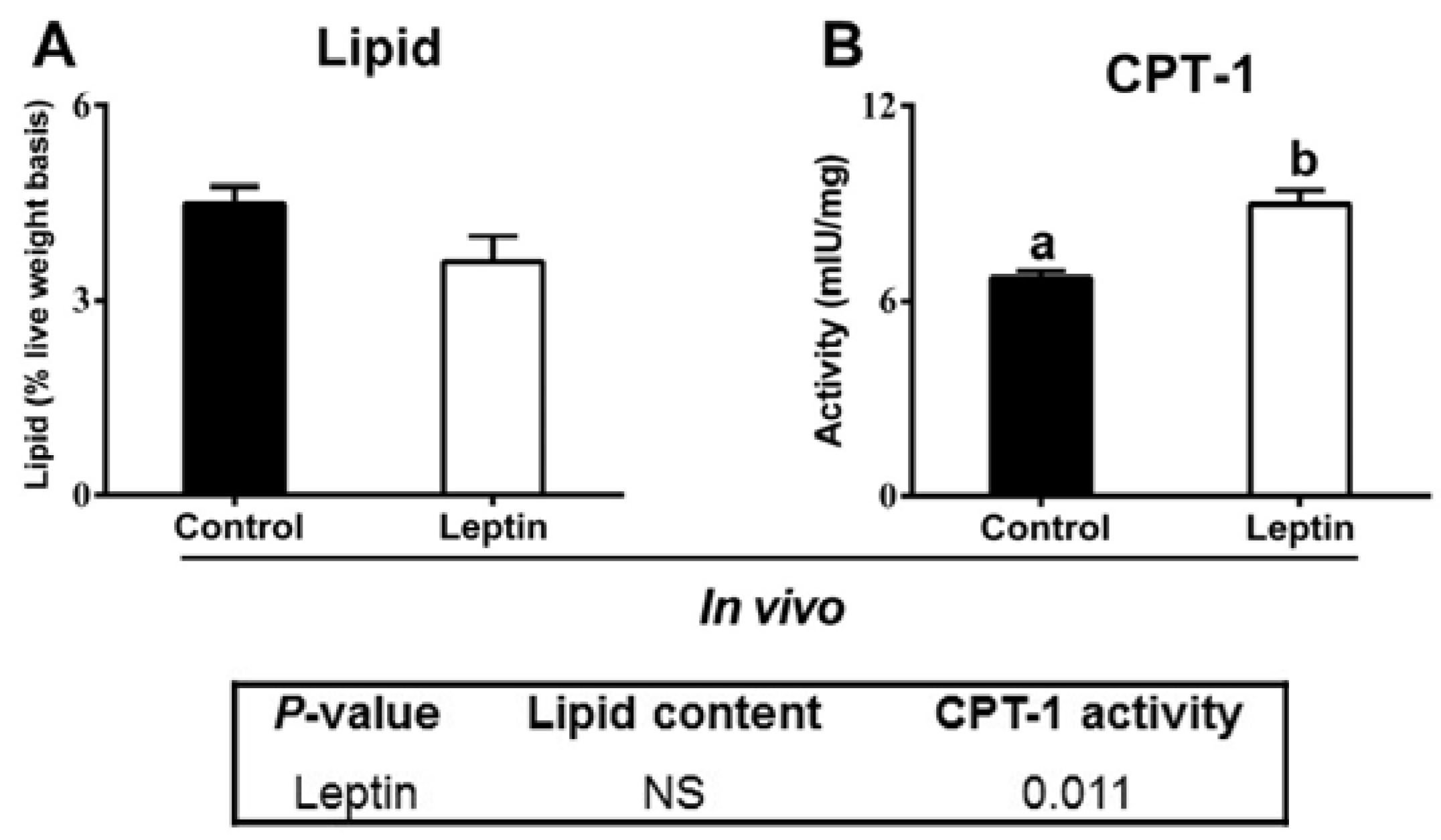
2.2.6. rt-hLEP Injection Activated Ovarian CPT-1 Activity
2.3. Leptin Induced Oocytes Maturation via Activation of JAK-STAT Pathway
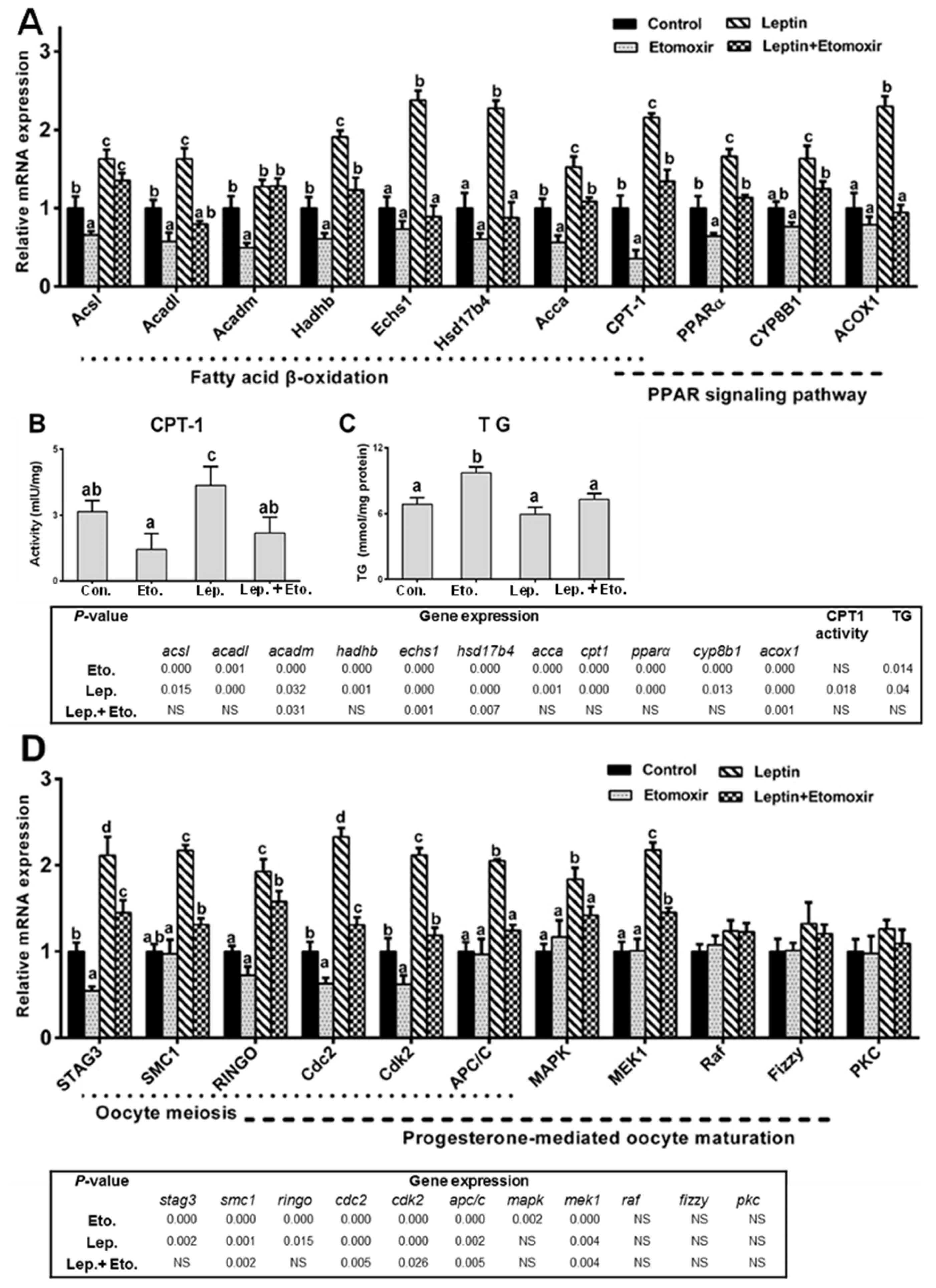
2.4. Fatty Acid β-Oxidation Is Essential for Leptin-Mediated Oocytes Maturation
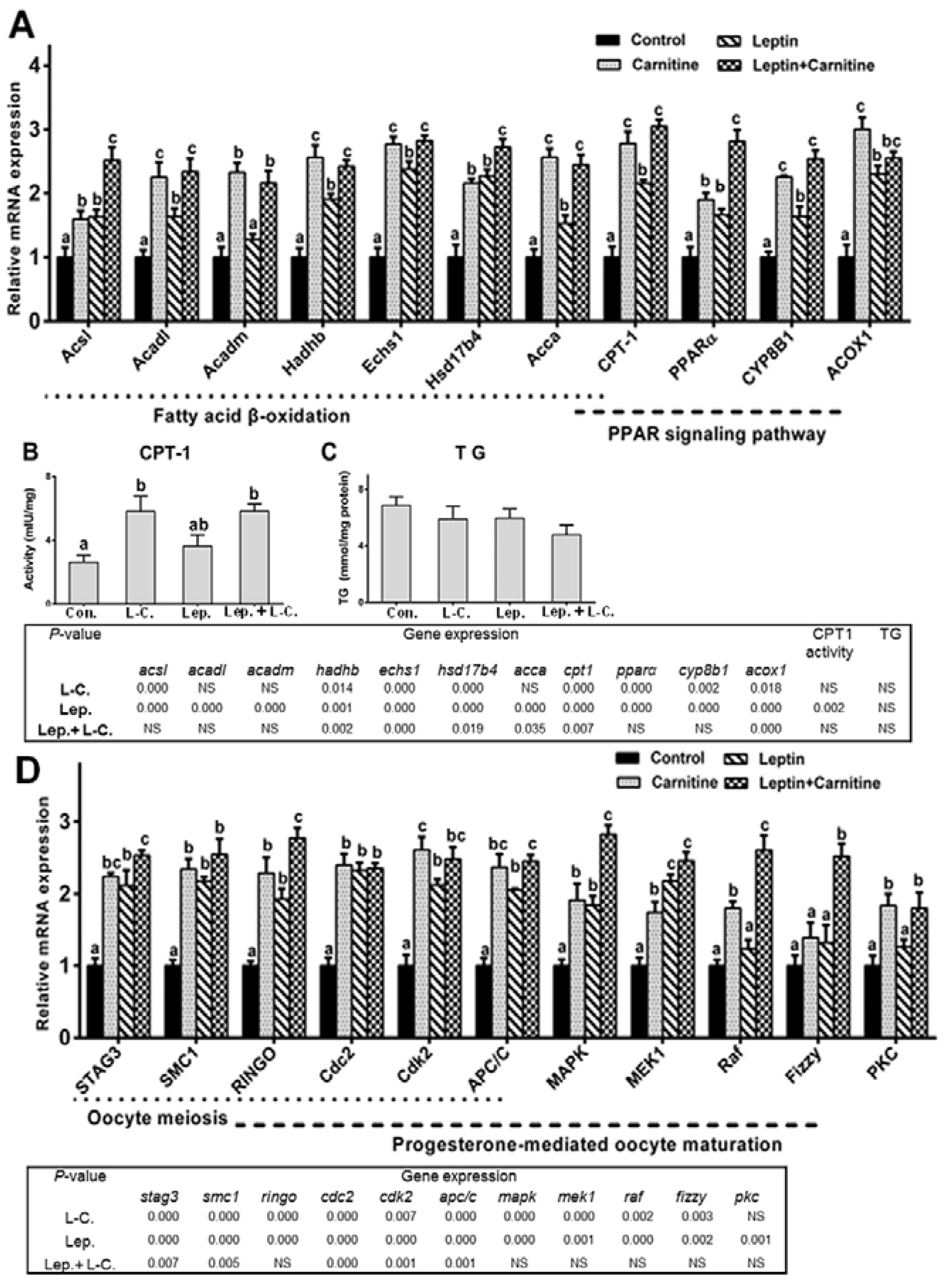
2.5. Fatty Acid β-Oxidation up-Regulation Improved Leptin-Mediated Oocytes Maturation
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Drug Treatment
4.3. Experimental Treatments
4.3.1. Experiment 1: In Vivo Study: Investigating the Role and Molecular Mechanism of Leptin Underlying Fatty Acid β-Oxidation and Oocytes Maturation
4.3.2. Experiment 2: In Vitro Study: Investigating whether Leptin Induced Oocytes Maturation and Its Relationship with JAK-STAT Pathway
4.3.3. Experiment 3: In Vitro Study: Investigating the Potential Role of Fatty Acid β-Oxidation in Leptin-Mediated Oocyte Maturation
4.4. Sample Analysis
4.4.1. Total RNA Extraction, cDNA Library Preparation and Illumina Sequencing
4.4.2. De Novo Assembly and Function Annotation
4.4.3. Identification of Differentially Expressed Genes
4.4.4. Real-Time Quantitative PCR (qPCR) Validation
4.4.5. Determination of Lipid and Triglyceride (TG) Contents, and CPT-1 Activity
4.5. Statistical Analysis
Declarations
Ethics Approval and Consent to Participate
Consent for Publication
Availability of Data and Materials
Authors Contributions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BW | Body weight |
| COC | Cumulus-oocyte complex |
| COG | Clusters of Orthologous Groups |
| CPT | Carnitine palmitoyltransferase |
| DMSO | Dimethyl sulfoxide |
| FDR | False discovery rate |
| FSH | Follicle-stimulating hormone |
| GnRH | Gonadotropin releasing hormone |
| GO | Gene Ontology |
| GV | Germinal vesicle |
| GVBD | Germinal vesicle breakdown |
| JAK | Janus kinase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LepR | Leptin receptor |
| LEP | Leptin |
| LH | Luteinizing hormone |
| MAPK | Mitogen-activated protein kinase |
| NCBI | National Center for Biotechnology Information |
| PBS | Phosphate buffered saline |
| PKC | Protein kinase C |
| PPAR | Peroxisome proliferator-activated receptor |
| qPCR | Real-time quantitative PCR |
| SRA | Short read archive |
| STAT | Signal transducer and activator of transcription |
| TG | Triglyceride |
References
- Paula-Lopes, F.F.; Boelhauve, M.; Habermann, F.A.; Sinowatz, F.; Wolf, E. Leptin promotes meiotic progression and developmental capacity of bovine oocytes via cumulus cell-independent and -dependent mechanisms. Biol. Reprod. 2007, 76, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and β-oxidation. Reproduction 2014, 148, 15–27. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, P.J.; Sturmey, R.G. The role of fatty acids in oocyte and early embryo development. Reprod. Fertil. Dev. 2011, 24, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Hillman, N.; Flynn, T.J. The metabolism of exogenous fatty acids by preimplantation mouse embryos developing in vitro. J. Embryol. Exp. Morphol. 1980, 56, 57–168. [Google Scholar]
- Downs, S.M.; Mosey, J.L.; Klinger, J. Fatty acid oxidation and meiotic resumption in mouse oocytes. Mol. Reprod. Dev. 2009, 76, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Valsangkar, D.; Downs, S.M. A requirement for fatty acid oxidation in the hormone-induced meiotic maturation of mouse oocytes. Biol. Reprod. 2013, 89, 43. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Cashman, K.; Russell, D.L.; Thompson, J.G.; Norman, R.J.; Robker, R.L. β-oxidation is essential for mouse oocyte developmental competence and early embryo development. Biol. Reprod. 2010, 83, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Akison, L.K.; Russell, D.L.; Norman, R.J.; Robker, R.L. Increased β-oxidation and improved oocyte developmental competence in response to L-carnitine during ovarian in vitro follicle development in mice. Biol. Reprod. 2011, 85, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Montjean, D.; Entezami, F.; Lichtblau, I.; Belloc, S.; Gurgan, T.; Menezo, Y. Carnitine content in the follicular fluid and expression of the enzymes involved in β-oxidation in oocytes and cumulus cells. J. Assist. Reprod. Genet. 2012, 291, 221–1225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.; Paglia, D.; Dwan, A.; Beital, M. Increased obese mRNA expression in omental fat cells from massively obese humans. Nat. Med. 1995, 1, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Dohm, G.L.; Fiedorek, F.T., Jr.; Tapscott, E.B.; Coleman, R.A. Leptin directly alters lipid partitioning in skeletal muscle. Diabetes 1997, 46, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Tan, X.Y.; Wu, K.; Zhuo, M.Q.; Song, Y.F.; Chen, Q.L. Regulation and mechanism of leptin on lipid metabolism in ovarian follicle cells from yellow catfish Pelteobagrus fulvidraco. Gen. Comp. Endocrinol. 2015, 222, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Luo, Z.; Zhu, Q.L.; Zheng, J.L.; Tan, X.Y.; Chen, Q.L. Characterization and tissue distribution of leptin, leptin receptor and leptin receptor overlapping transcript genes in yellow catfish (Pelteobagrus fulvidraco). Gen. Comp. Endocrinol. 2013, 182, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ryan, N.K.; Woodhouse, C.M.; Hoek, K.H.V.D.; Gilchrist, R.B.; Armstrong, D.T.; Norman, R.J. Expression of leptin and its receptor in the murine ovary: Possible role in the regulation of oocyte maturation. Biol. Reprod. 2002, 66, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Boelhauve, M.; Sinowatz, F.; Wolf, E.; Paula-Lopes, F.F. Maturation of Bovine Oocytes in the Presence of Leptin Improves Development and Reduces Apoptosis of In Vitro-Produced Blastocysts 1. Biol. Reprod. 2005, 73, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Duggal, P.S.; Ryan, N.K.; Kh, V.D.H.; Ritter, L.J.; Armstrong, D.T.; Magoffin, D.A. Effects of leptin administration and feed restriction on thecal leucocytes in the preovulatory rat ovary and the effects of leptin on meiotic maturation, granulosa cell proliferation, steroid hormone and PGE2 release in culture rat ovarian follicles. Reproduction 2002, 123, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Spicer, L.J.; Francisco, C.C. Adipose obese gene product, leptin, inhibits bovine ovarian thecal cell steroidogenesis. Biol. Reprod. 1998, 58, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.H.; Kimura, M.; Walczewska, A.; Karanth, S.; Mccann, S.M. Role of leptin in hypothalamic–pituitary function. Proc. Natl. Acad. Sci. USA 1997, 94, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Watanobe, H. Leptin directly acts within the hypothalamus to stimulate gonadotropin-releasing hormone secretion in vivo in rats. J. Physiol. 2002, 545, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Peyon, P.; Zanuy, S.; Carrillo, M. Actions of leptin on in vitro luteinizing hormone release in the European sea bass (Dicentrarchus labrax). Biol. Reprod. 2012, 65, 1573–1578. [Google Scholar]
- Weil, C.; Bail, P.Y.L.; Sabin, N.; Gac, F.L. In vitro action of leptin on FSH and LH production in rainbow trout (Onchorynchus mykiss) at different stages of the sexual cycle. Gen. Comp. Endocrinol. 2003, 130, 2–12. [Google Scholar] [CrossRef]
- Saxena, N.K.; Sharma, D.; Ding, X.; Lin, S.; Marra, F.; Merlin, D. Concomitant activation of the JAK/STAT, PI3K/AKT, and ERK signaling is involved in leptin-mediated promotion of invasion and migration of hepatocellular carcinoma cells. Cancer Res. 2007, 67, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Miyazawa, M.; Kamei, A.; Abe, K.; Kojima, T. Ameliorative effects of mulberry (Morus alba L.) leaves on hyperlipidemia in rats fed a high-fat diet: Induction of fatty acid oxidation, inhibition of lipogenesis, and suppression of oxidative stress. Biosci. Biotechnol. Biochem. 2010, 74, 2385–2395. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.A.; Sinclair, G.; Mcintosh, S.; Bamforth, F.; Thompson, R.; Sobol, I.; Osborne, G.; Corriveau, A.; Santos, M.; Hanley, B. Carnitine palmitoyltransferase 1A (CPT1A) P479L prevalence in live newborns in Yukon, Northwest Territories, and Nunavut. Mol. Genet. Metab. 2010, 101, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Kerner, J.; Hoppel, C. Fatty acid import into mitochondria. BBA Mol. Cell Biol. 2000, 1486, 1–17. [Google Scholar] [CrossRef]
- Londraville, R.L.; Duvall, C.S. Murine leptin injections increase intracellular fatty acid-binding protein in green sunfish (Lepomis cyanellus). Gen. Comp. Endocrinol. 2002, 129, 56–62. [Google Scholar] [CrossRef]
- Yamagishi, S.I.; Edelstein, D.; Du, X.L.; Kaneda, Y.; Guzmán, M.; Brownlee, M. Leptin induces mitochondrial superoxide production and monocyte chemoattractant protein-1 expression in aortic endothelial cells by increasing fatty acid oxidation via protein kinase A. J. Biol. Chem. 2001, 276, 25096–25100. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kim, Y.B.; Peroni, O.D.; Fryer, L.G.; Müller, C.; Carling, D. Leptin stimulates fatty-acid oxidation by activating AMP-activated protein kinase. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M.; Flier, J.S. Obesity and the regulation of energy balance. Cell 2001, 104, 531–543. [Google Scholar] [CrossRef]
- Fair, T.; Hyttel, P.; Greve, T. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol. Reprod. Dev. 1995, 42, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, S.H.; Ma, Y.F.; Lou, Y.K.; Wei, H.X.; Sun, X.Z.; Zhao, Y.H.; Li, Y.; Dai, Y.P.; Zhang, L.; et al. Effects of leptin supplementation in in vitro maturation medium on meiotic maturation of oocytes and preimplantation development of parthenogenetic and cloned embryos in pigs. Anim. Reprod. Sci. 2007, 101, 85–96. [Google Scholar]
- Ghilardi, N.; Skoda, R.C. The leptin receptor activates Janus kinase 2 and signals for proliferation in a factor-dependent cell line. Mol. Endocrinol. 1997, 11, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Okimura, Y.; Mizuno, I.; Iida, K.; Takahashi, T.; Kaji, H. Leptin induces mitogenactivated protein kinase-dependent proliferation C3H10T1/2 cells. J. Biol. Chem. 1997, 272, 12897–12900. [Google Scholar] [CrossRef] [PubMed]
- Arias-Álvarez, M.; García-García, R.M.; Torres-Rovira, L.; González-Bulnes, A.; Rebollar, P.G.; Lorenzo, P.L. Influence of leptin on in vitro maturation and steroidogenic secretion of cumulus–oocyte complexes through JAK2/STAT3 and MEK 1/2 pathways in the rabbit model. Reproduction 2010, 139, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.C.; Tian, J.X.; Lv, Y.Y.; Shi, F.X.; Kong, F.D.; Shi, H.; Zhao, L.Y. Leptin induces functional activation of cyclooxygenase-2 through JAK2/STAT3, MAPK/ERK, and PI3K/AKT pathways in human endometrial cancer cells. Cancer Sci. 2009, 100, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.S.; Smift, A.L.; Croteau, N.J.; Ferrick, D.A.; Wu, M. Inhibition of fatty acid oxidation by etomoxir impairs NADPH production and increases reactive oxygen species resulting in ATP depletion and cell death in human glioblastoma cells. Biochim. Biophys. Acta 2011, 1807, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.R.; Hong, Y.M.; Boriack, R.L.; Bennett, M.J. Effect of L-carnitine supplementation on cardiac carnitine palmitoyltransferase activities and plasma carnitine concentrations in adriamycin-treated rats. Pediatr. Res. 2003, 53, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazik, H.; Sharma, R.; Mahfouz, R.; Agarwal, A. L-Carnitine decreases DNA damage and improves the in vitro blastocyst development rate in mouse embryos. Fertil. Steril. 2009, 91, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Mansour, G.; Abdelrazik, H.; Sharma, R.K.; Radwan, E.; Falcone, T.; Agarwal, A. L-Carnitine supplementation reduces oocyte cytoskeleton damage and embryo apoptosis induced by incubation in peritoneal fluid from patients with endometriosis. Fertil. Steril. 2009, 91, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.G.; Reis, A.; Leese, H.J.; McEvoy, T.G. Role of fatty acids in energy provision during oocyte maturation and early embryo development. Reprod. Domest. Anim. 2009, 44, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Baltzeger, D.A.; Reading, B.J.; Douros, J.D.; Borski, R.J. Role for leptin in promoting glucose mobilization during acute hyperosomotic stress in teleost fishes. J. Endocrinol. 2014, 220, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Copeland, D.L.; Duff, R.J.; Liu, Q.; Prokop, J.; Londraville, R.L. Leptin in teleost fishes: An argument for comparative study. Front. Physiol. 2011, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.F.; Wu, K.; Tan, X.Y.; Zhang, L.H.; Zhuo, M.Q.; Pan, Y.X. Effects of recombinant human leptin administration on hepatic lipid metabolism in yellow catfish Pelteobagrus fulvidraco: In vivo and in vitro studies. Gen. Comp. Endocrinol. 2015, 212, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y. Yellow Catfish Leptin: Gene Cloning, Tissues Expression and Its Relationship with Lipid Metabolism. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2013. [Google Scholar]
- Liu, W.B.; Zhang, X.J. Study on the development and annual change in the ovary of Pelteobagrus fulvidraco. J. Nat. Sci. Hunan Norm. Univ. 2003, 26, 73–78, (In Chinese with English Abstract). [Google Scholar]
- Goetz, F.W.; Bergman, H.L. The effects of steroids on final maturation and ovulation of oocytes from brook trout (Salvelinus fontinalis) and yellow perch (Perca flavescens). Biol. Reprod. 1978, 18, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Goetz, F.W.; Theofan, G. In vitro stimulation of germinal vesicle breakdown and ovulation of yellow perch (Perca flavescens) oocytes. Effects of 17α-hydroxy-20β-dihydroprogesterone and prostaglandins. Gen. Comp. Endocrinol. 1979, 37, 273–285. [Google Scholar] [CrossRef]
- Turathum, B.; Saikhun, K.; Sangsuwan, P.; Kitiyanant, Y. Effects of vitrification on nuclear maturation, ultrastructural changes and gene expression of canine oocytes. Reprod. Biol. Endocrinol. 2010, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- De los Reyes, M.; de Lange, J.; Miranda, P.; Palominos, J.; Barros, C. Effect of human chorionic gonadotrophin supplementation during different culture periods on in vitro maturation of canine oocytes. Theriogenology 2005, 64, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kiorpes, T.C.; Hoerr, D.; Ho, W.; Weaner, L.E.; Inman, M.G.; Tutwiler, G.F. Identification of 2-tetradecylglycidyl coenzyme A as the active form of methyl 2-tetradecylglycidate (methyl palmoxirate) and its characterization as an irreversible, active site-directed inhibitor of carnitine palmitoyl transferase A in isolated rat liver mitochondria. J. Biol. Chem. 1984, 259, 9750–9755. [Google Scholar] [PubMed]
- He, S.; Liang, X.F.; Li, L.; Sun, J.; Wen, Z.Y.; Cheng, X.Y. Transcriptome analysis of food habit transition from carnivory to herbivory in a typical vertebrate herbivore, grass carp Ctenopharyngodon idella. BMC Genom. 2015, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2001, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; Garcíagómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; Mccue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Zhao, Y.; Gul, Y.; Li, S.; Wang, W. Cloning, identification and accurate normalization expression analysis of PPARα gene by GeNorm in Megalobrama amblycephala. Fish Shellfish Immunol. 2011, 31, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Tan, X.Y.; Zheng, J.L.; Chen, Q.L.; Liu, C.X. Quantitative dietary zinc requirement of juvenile yellow catfish (Pelteobagrus fulvidraco), and effects on hepatic intermediary metabolism and antioxidant responses. Aquaculture 2011, 319, 150–155. [Google Scholar] [CrossRef]
- Suarez, R.K.; Hochachka, P.W. Preparation and properties of rainbow trout liver mitochondria. J. Comp. Physiol. 1981, 143, 269–273. [Google Scholar]
- Morash, A.J.; Kajimura, M.; McClelland, G.B. Intertissue regulation of carnitine palmitoyltransferase I (CPT I): Mitochondrial membrane properties and gene expression in rainbow trout (Oncorhynchus mykiss). Biochim. Biophys. Acta 2008, 1778, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Bieber, L.L.; Fiol, C. Purification and assay of carnitine acyltransferases. Method Enzymol. 1986, 123, 276–284. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Clean Reads | Total Clean Nucleotides (nt) | Q20 Percentage | N Percentage | GC Percentage |
|---|---|---|---|---|---|
| Control | 50,181,922 | 5,018,192,200 | 97.19% | 0.00% | 48.55% |
| Leptin injection | 41,408,858 | 4,140,885,800 | 97.95% | 0.00% | 48.03% |
| Parameters | Sample | Total Number | Total Length (nt) | Mean Length (nt) | N50 a (nt) |
|---|---|---|---|---|---|
| Contig | Control | 76,153 | 39,303,110 | 516 | 1631 |
| Leptin injection | 75,384 | 38,943,257 | 517 | 1650 | |
| Unigene | Control | 45,002 | 46,286,299 | 1029 | 2217 |
| Leptin injection | 44,503 | 45,719,364 | 1027 | 2232 | |
| All-Unigene | 36,149 | 49,948,406 | 1382 | 2440 |
| Pathways | Up-Regulated Genes | Down-RegulaTed Genes | Pathway ID (p-Value) |
|---|---|---|---|
| Signal Transduction | |||
| JAK-STAT signaling pathway | Leptin R, JAK, STAM, CIS, C-Myc, PI3K | —— | map04630 (0.000242) |
| MAPK signaling pathway | MAPK, MEK1, MEKK, PP3C, HSP72, CASP, IL1, CrkII, C-Myc | Raf, PKC | map04010 (0.000472) |
| AMPK signaling pathway | LeptinR, CPT1, AMPK, PI3K, Acc, IGF-1R | PPARγ | map04152 (0.00180) |
| β-Oxidation | |||
| Fatty acid β-oxidation | CPT1, Acsl, Acadl, Acadm, Hadhb, Echsl, Hsd17b4, Acca | Acads | —— |
| PPAR signaling pathway | CPT1, PPARα, CYP8B1, ACOX1, ACBP | PPARγ, FATCD36, UBC, PDK1 | map03320 (0.00773) |
| Oocytes Maturation | |||
| Progesterone-mediated oocyte maturation | PI3K, MAPK, RINGO, Cdc2, MEK1, IGF-1R, APC/C | Raf, Fizzy | map04914 (0.00309) |
| Oocyte meiosis signaling pathway | RINGO, APC/C, Cdc2, Cdk2, STAG3, SMC1 | C3H-4 | map04114 (0.0164) |
| GnRH signaling pathway | GnRHR, MEKK, CREB, HB-EGF, FSHβ | Raf, PKC | map04912 (0.0242) |
| Cell cycle signaling pathway | APC/C, SMC1, Cdh1, Cdk2, C-Myc | —— | map04110 (0.000123) |
| DEGs | Functions |
|---|---|
| Leptin R | a receptor for the fat cell-specific hormone leptin |
| JAK | a family of intracellular tyrosine kinases that transduce cytokine-mediated signals via the JAK-STAT pathway |
| STAM (signal transducing adaptor molecule) | identified by the rapid tyrosine-phosphorylation of its product in response to cytokine stimulation |
| CIS (cytokine inducible SH2-containing protein) | cytokine-inducible negative regulators of cytokine signaling |
| C-Myc (proto-oncogene C-Myc) | a regulator gene that plays a role in cell cycle progression, apoptosis and cellular transformation |
| PI3K (phosphatidylinositol 3-kinase) | a family of enzymes involved in cellular functions such as cell growth, proliferation, differentiation, motility, survival and intracellular trafficking |
| MEK1 (mitogen-activated protein kinase kinase 1) | stimulates the enzymatic activity of MAP kinases upon activation by a wide variety of extra- and intracellular signals |
| MEKK (mitogen-activated protein kinase kinase kinase) | a serine/threonine kinase that occupies a pivotal role in a network of phosphorylating enzymes integrating cellular responses to a number of mitogenic and metabolic stimuli |
| PP3C (serine/threonine-protein phosphatase 2B catalytic subunit) | involved in a wide range of biologic activities, acting as a Ca2+-dependent modifier of phosphorylation status |
| HSP72 (heat shock 70 kDa protein 1/A) | a member of the heat shock protein 70 family which facilitates the proper folding of newly translated and misfolded proteins, as well as stabilize or degrade mutant proteins |
| CASP (caspase) | a family of protease enzymes playing essential roles in programmed cell death (including apoptosis, pyroptosis and necroptosis) and inflammation |
| IL1 (interleukin 1) | plays a central role in the regulation of immune and inflammatory responses to infections or sterile insults |
| CrkII (proto-oncogene C-crk) | a member of an adapter protein family that binds to several tyrosine-phosphorylated proteins and involved in several signaling pathways, recruiting cytoplasmic proteins in the vicinity of tyrosine kinase through SH2-phosphotyrosine interaction |
| Raf (B-Raf proto-oncogene serine/threonine-protein kinase) | part of the ERK1/2 pathway as a MAP kinase kinase kinase (MAPKKK) that functions downstream of the Ras subfamily of membrane associated GTPases |
| PKC (protein kinase C) | a family of protein kinase enzymes involved in controlling the function of other proteins through the phosphorylation of hydroxyl groups of serine and threonine amino acid residues on these proteins, or a member of this family |
| CPT (carnitine palmitoyltransferase) | a mitochondrial enzyme responsible for the formation of acyl carnitines by catalyzing the transfer of the acyl group of a long-chain fatty acyl-CoA from coenzyme A to l-carnitine |
| AMPK (5′-AMP-activated protein kinase) | an enzyme that plays a role in cellular energy homeostasis |
| Acc (acetyl-CoA carboxylase) | a biotin-dependent enzyme that catalyzes the irreversible carboxylation of acetyl-CoA to produce malonyl-CoA through its two catalytic activities, biotin carboxylase and carboxyltransferase |
| IGF-1R (insulin-like growth factor 1 receptor) | a transmembrane receptor that mediates the effects of IGF-1 |
| PPAR (peroxisome proliferator-activated receptor) | a group of nuclear receptor proteins that function as transcription factors regulating the expression of genes involved in cellular differentiation, development, and metabolism (carbohydrate, lipid, protein), and tumorigenesis of higher organisms |
| Acsl (long-chain acyl-CoA synthetase) | converts free long-chain fatty acids into fatty acyl-CoA esters |
| Acadm (acyl-CoA dehydrogenase) | a class of enzymes that function to catalyze the initial step in each cycle of fatty acid β-oxidation in the mitochondria |
| Hadhb (hydroxyacyl-CoA dehydrogenase, β subunit) | functions in the mitochondrial matrix to catalyze the oxidation of straight-chain 3-hydroxyacyl-CoAs |
| Echs (enoyl-CoA hydratases) | an enzyme that hydrates the double bond between the second and third carbons on acyl-CoA |
| Hsd17b4 (hydroxyacysteroid 17-β dehydrogenase) | a group of alcohol oxidoreductases which catalyse the dehydrogenation of 17-hydroxysteroids in steroidogenesis |
| Acads (acyl-CoA dehydrogenase, short-chain) | an enzyme with systematic name short-chain acyl-CoA: electron-transfer flavoprotein 2,3-oxidoreductase |
| CYP8B1 (sterol 12-alpha-hydroxylase) | A member of the family of oxidoreductases, specifically those acting on paired donors, with O2 as oxidant and incorporation or reduction of oxygen |
| ACO (acyl-CoA oxidase) | A member of the family of oxidoreductases, specifically those acting on the CH-CH group of donor with oxygen as acceptor |
| ACBP (diazepam-binding inhibitor) | encodes diazepam binding inhibitor, a protein that is regulated by hormones and involved in lipid metabolism and the displacement of ß-carbolines and benzodiazepines, which modulate signal transduction at type A γ-aminobutyric acid receptors located in brain synapses |
| FATDC36 (CD36 antigen) | Leucocyte antigens on cell surfaces which recognizes oxidized low density lipoprotein, long chain fatty acids, anionic phospholipids, collagen types I, IV and V, thrombospondin and plasmodium falciparum infected erythrocytes |
| UBC (ubiquitin C) | plays a key role in maintaining cellular ubiquitin levels under stress conditions |
| PDK1 (3-phosphoinositide dependent protein kinase-1) | a master kinase crucial for the activation of AKT/PKB and many other AGC kinases including PKC, S6K, SGK |
| RINGO | a Cdc2 and Cdk2 activator, whose accumulation seems to be required for progesterone-induced oocyte maturation |
| Cdc (cyclin-dependent kinase; Cdk) | a family of protein kinases which are first discovered for their role in regulating the cell cycle |
| Fizzy (fizzy/cell division cycle 20 related 1) | Fizzy directly bind to anaphase-promoting complex and activate its cyclin ubiquitination activity |
| STAG3 (cohesin complex subunit SA-3) | a subunit of the cohesin complex which regulates the cohesion of sister chromatids during cell division |
| SMC1 (structural maintenance of chromosome 1) | A member of the family of proteins required for chromatid cohesion and DNA recombination during meiosis and mitosis |
| C3H-4 (CCCH zinc finger protein C3H-4) | encodes a CCCH-type zinc finger protein that is thought to prevent infection by retroviruses and may function to inhibit viral gene expression and induce an innate immunity to viral infection |
| GnRHR (gonadotropin-releasing hormone receptor) | a member of the seven-transmembrane, G-protein coupled receptor family and responsible for eliciting the actions of LHRH after its release from the hypothalamus |
| CREB (cyclic AMP-dependent transcription factor ATF-4) | a cellular transcription factor |
| HB-EGF (heparin-binding EGF-like growth factor) | play a role in wound healing, cardiac hypertrophy, and heart development and function |
| FSH (follicle stimulating hormone) | a glycoprotein polypeptide hormone which regulates the development, growth, pubertal maturation, and reproductive processes of the body |
| APC/C (anaphase-promoting complex subunit 1) | an E3 ubiquitin ligase that marks target cell cycle proteins for degradation by the 26S proteasome |
| Treatment | No. Oocytes Cultured | Nuclear Status (%) | Mean Diamater of Oocytes (μm) | ||||
|---|---|---|---|---|---|---|---|
| GV | GVBD | M I | M II | GVBD-MII | |||
| Control | 119 | 20.8 ± 2.8 de | 46.7 ± 3.0 c | 28.3 ± 1.0 b | 4.2 ± 1.6 a | 79.2 ± 2.8 b | 103.9 ± 2.8 b |
| AG490 | 120 | 11.7 ± 2.2 bc | 44.2 ± 3.2 bc | 35.0 ± 1.7 c | 9.2 ± 2.5 abc | 88.3 ± 2.2 cd | 115.9 ± 2.3 cd |
| Etomoxir | 119 | 34.2 ± 2.5 f | 44.2 ± 2.8 bc | 17.5 ± 1.6 a | 3.3 ± 2.3 a | 65.8 ± 2.5 a | 89.3 ± 2.0 a |
| Carnitine | 120 | 7.5 ± 2.1 ab | 40.0 ± 3.0 abc | 44.2 ± 3.2 d | 8.3 ± 1.7 abc | 92.5 ± 2.1 d | 122.5 ± 2.5 df |
| Leptin | 116 | 5.8 ± 1.6 ab | 33.3 ± 1.4 a | 40.8 ± 2.1 d | 16.7 ± 4.1 c | 94.2 ± 1.6 d | 128.0 ± 1.6 f |
| Leptin + AG490 | 117 | 15.8 ± 1.6 cd | 43.3 ± 3.0 bc | 32.5 ± 1.6 bc | 5.8 ± 3.7 ab | 84.1 ± 1.6 bc | 107.9 ± 3.8 bc |
| Leptin + Etomoxir | 120 | 24.2 ± 1.6 e | 40.8 ± 2.8 abc | 29.7 ± 1.6 bc | 5.8 ± 3.4 ab | 75.8 ± 1.6 b | 105.4 ± 3.0 b |
| Leptin + Carnitine | 116 | 3.3 ± 1.4 a | 35.8 ± 1.6 ab | 44.2 ± 2.5 d | 13.3 ± 1.4 bc | 96.7 ± 1.4 d | 125.9 ± 3.0 f |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.-F.; Tan, X.-Y.; Pan, Y.-X.; Zhang, L.-H.; Chen, Q.-L. Fatty Acid β-Oxidation Is Essential in Leptin-Mediated Oocytes Maturation of Yellow Catfish Pelteobagrus fulvidraco. Int. J. Mol. Sci. 2018, 19, 1457. https://doi.org/10.3390/ijms19051457
Song Y-F, Tan X-Y, Pan Y-X, Zhang L-H, Chen Q-L. Fatty Acid β-Oxidation Is Essential in Leptin-Mediated Oocytes Maturation of Yellow Catfish Pelteobagrus fulvidraco. International Journal of Molecular Sciences. 2018; 19(5):1457. https://doi.org/10.3390/ijms19051457
Chicago/Turabian StyleSong, Yu-Feng, Xiao-Ying Tan, Ya-Xiong Pan, Li-Han Zhang, and Qi-Liang Chen. 2018. "Fatty Acid β-Oxidation Is Essential in Leptin-Mediated Oocytes Maturation of Yellow Catfish Pelteobagrus fulvidraco" International Journal of Molecular Sciences 19, no. 5: 1457. https://doi.org/10.3390/ijms19051457