Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen

 , ,
, ,
Abstract
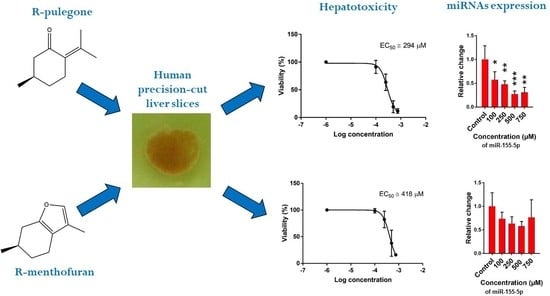
:
1. Introduction
2. Results
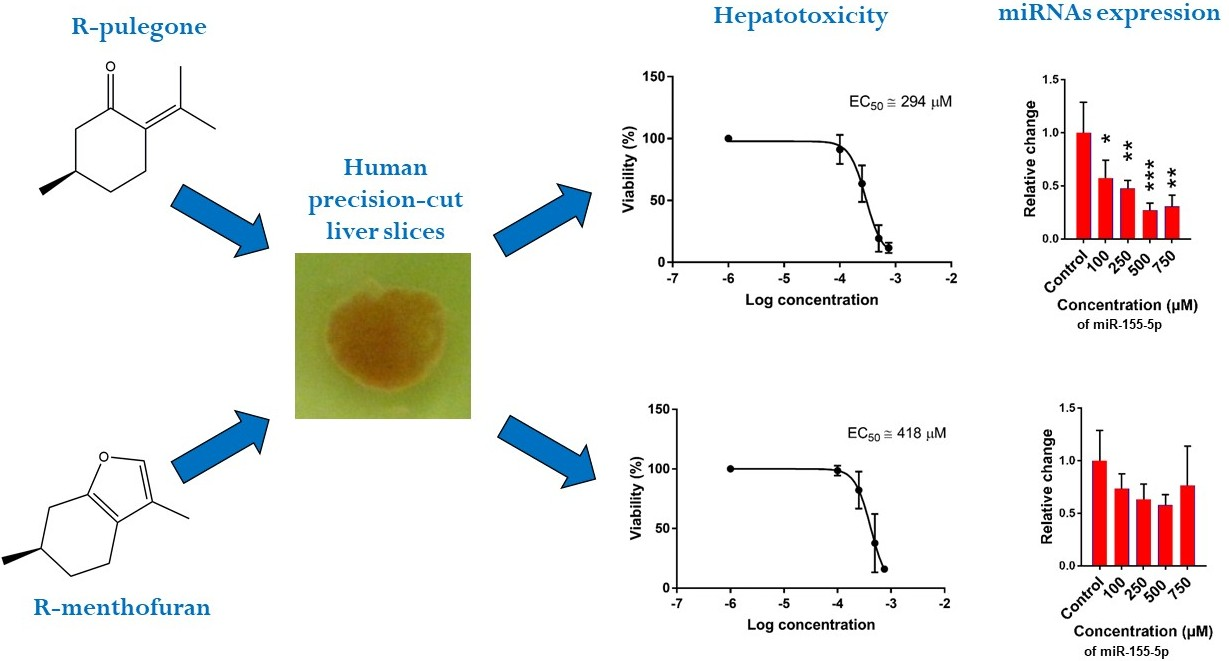
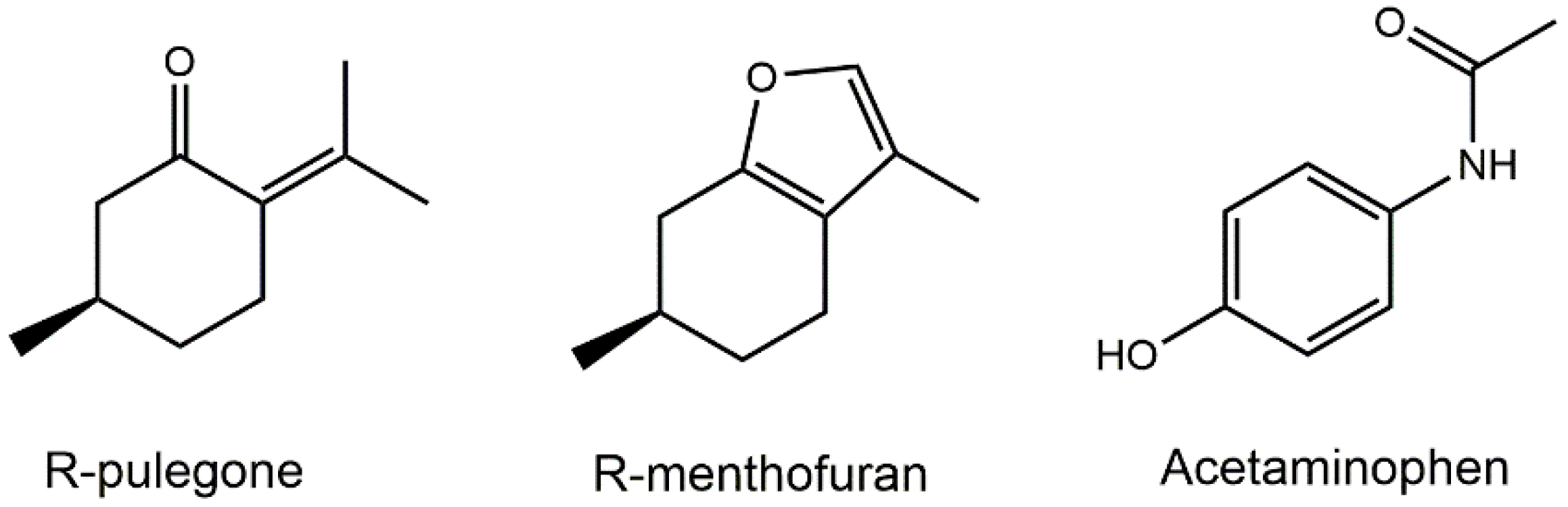
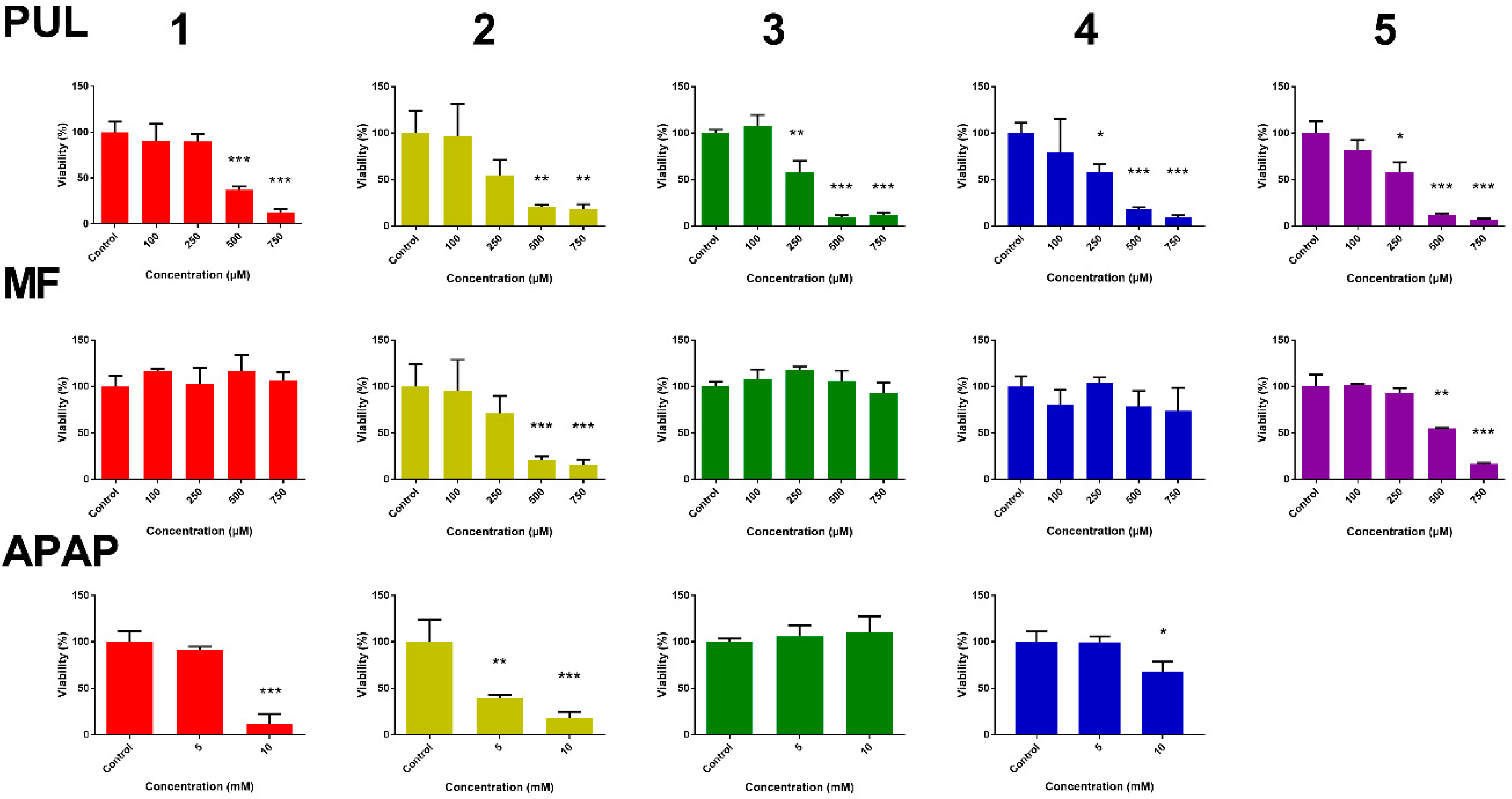
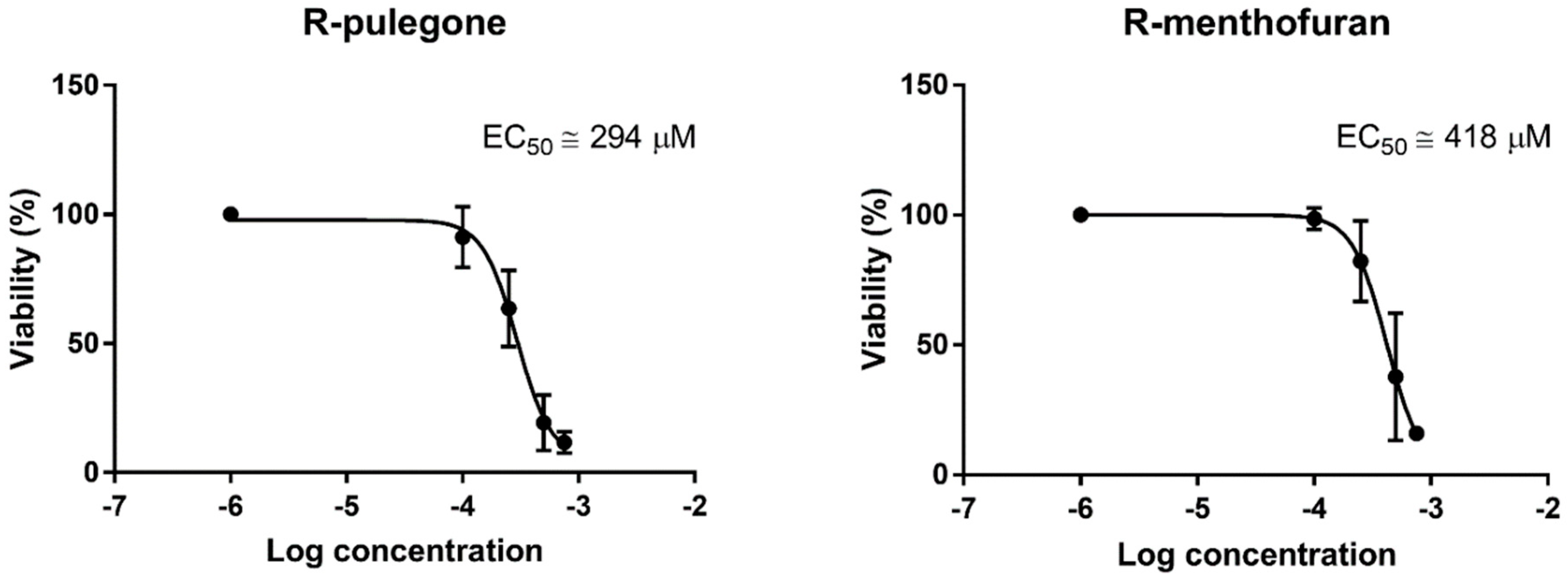
2.1. Hepatotoxicity of R-pulegone, R-menthofuran and Acetaminophen
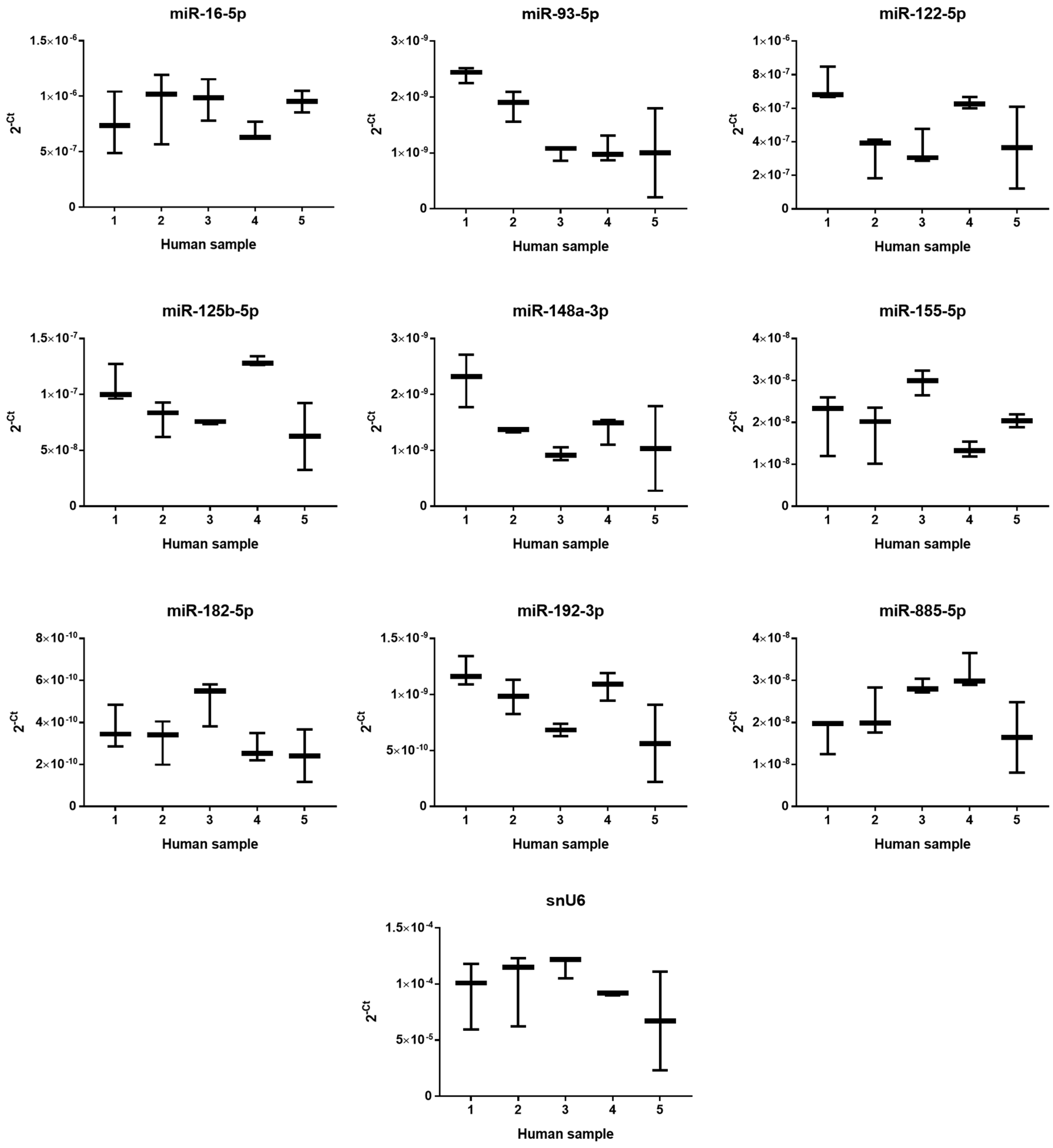
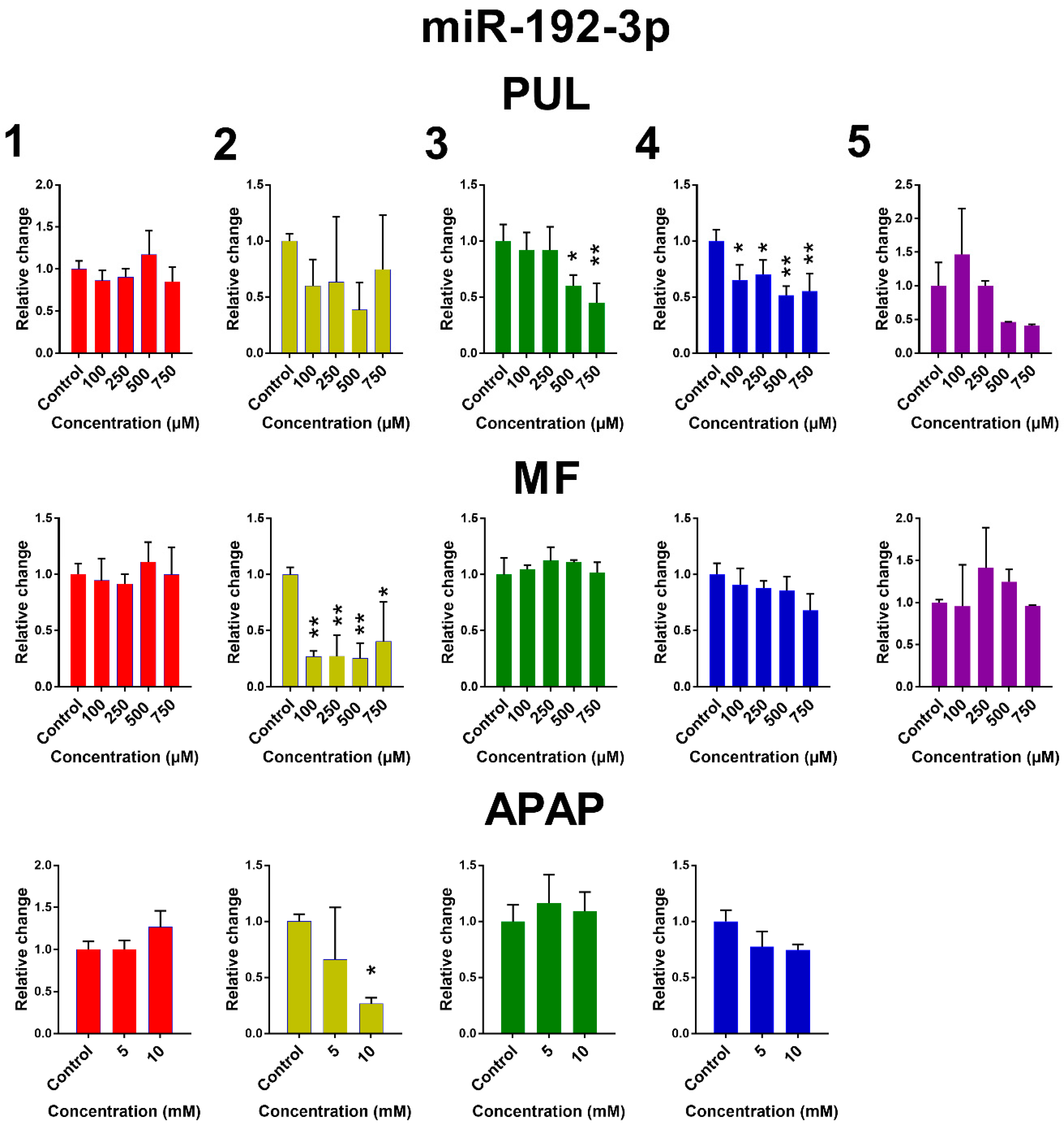
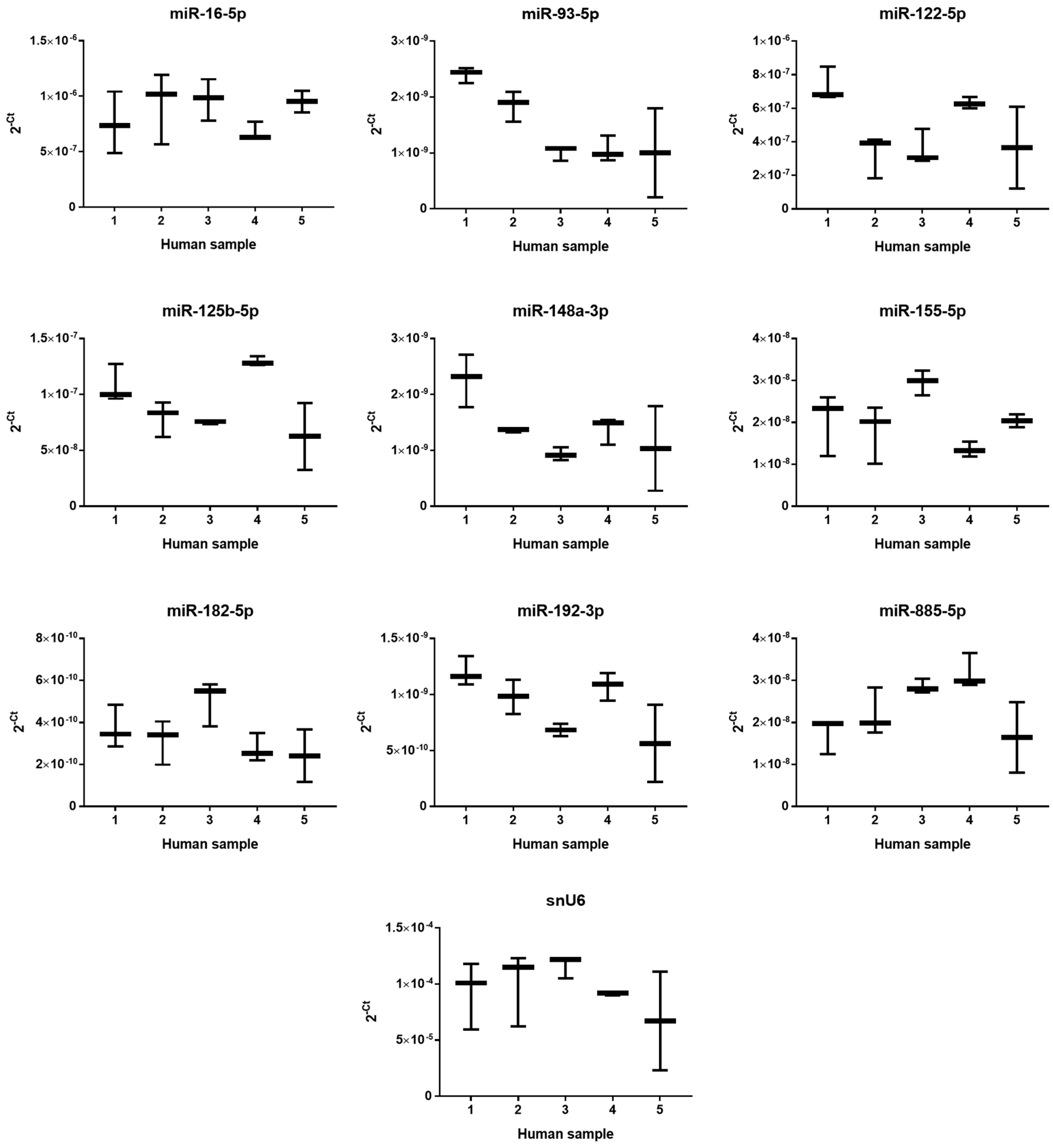
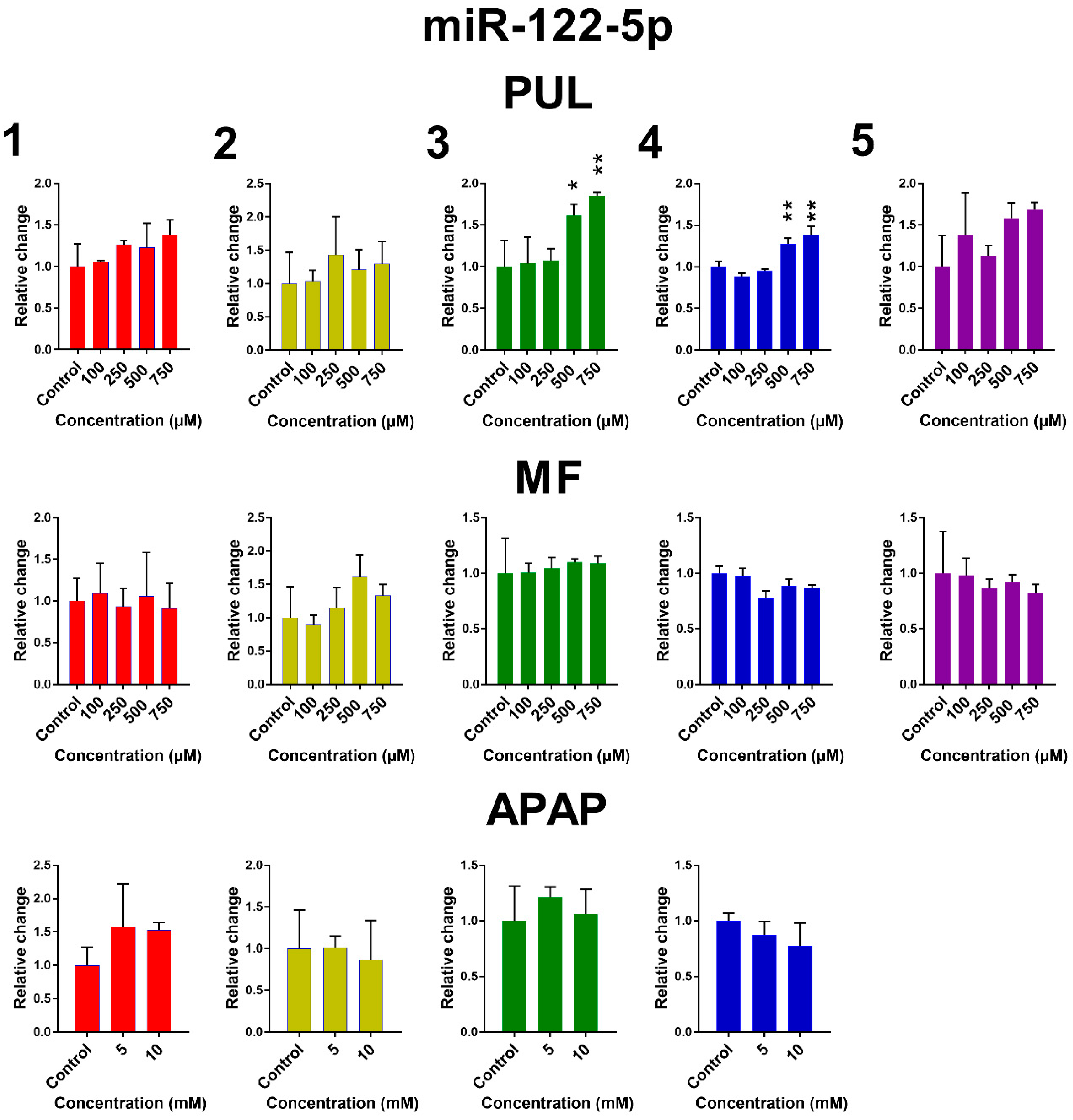
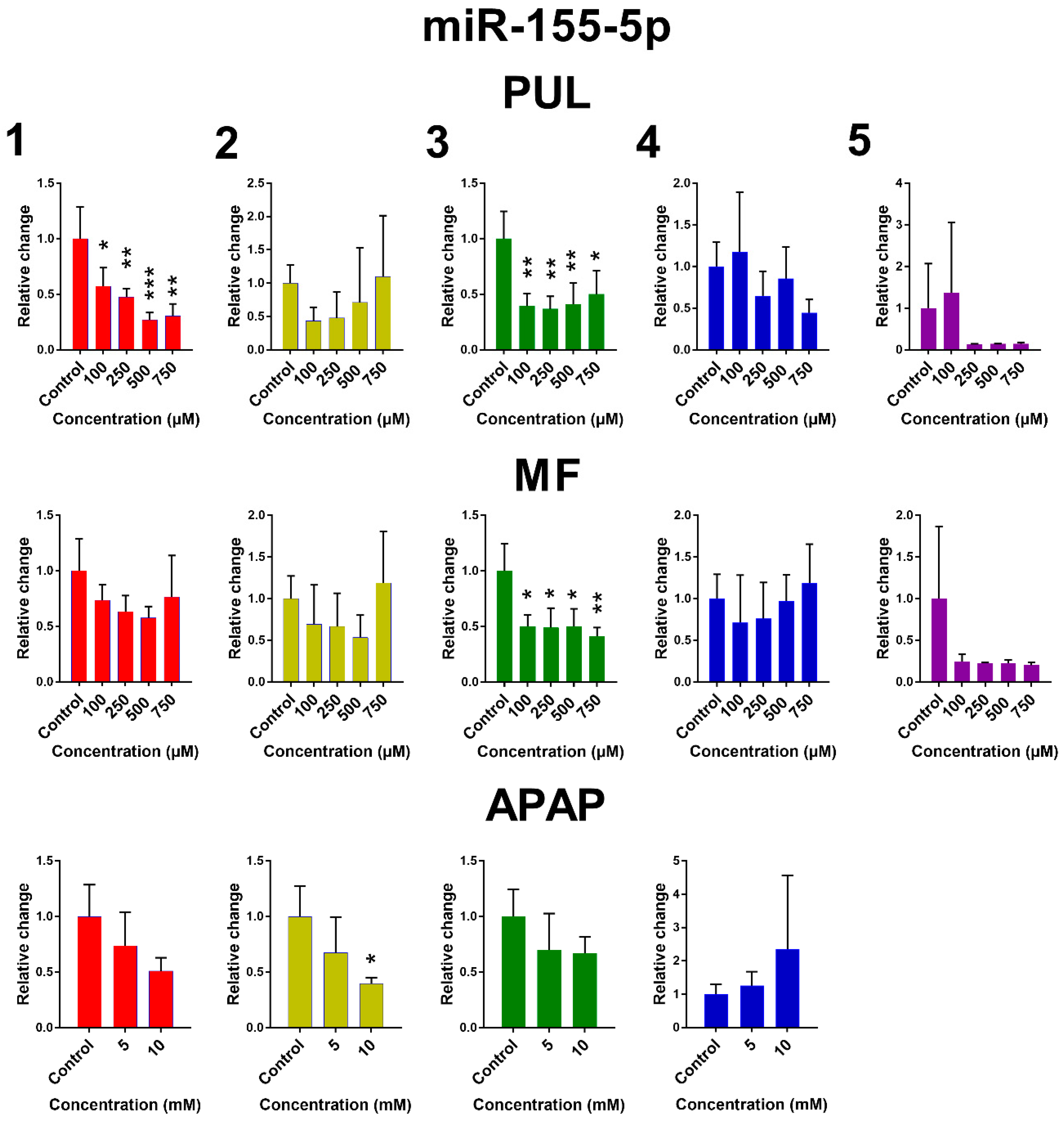
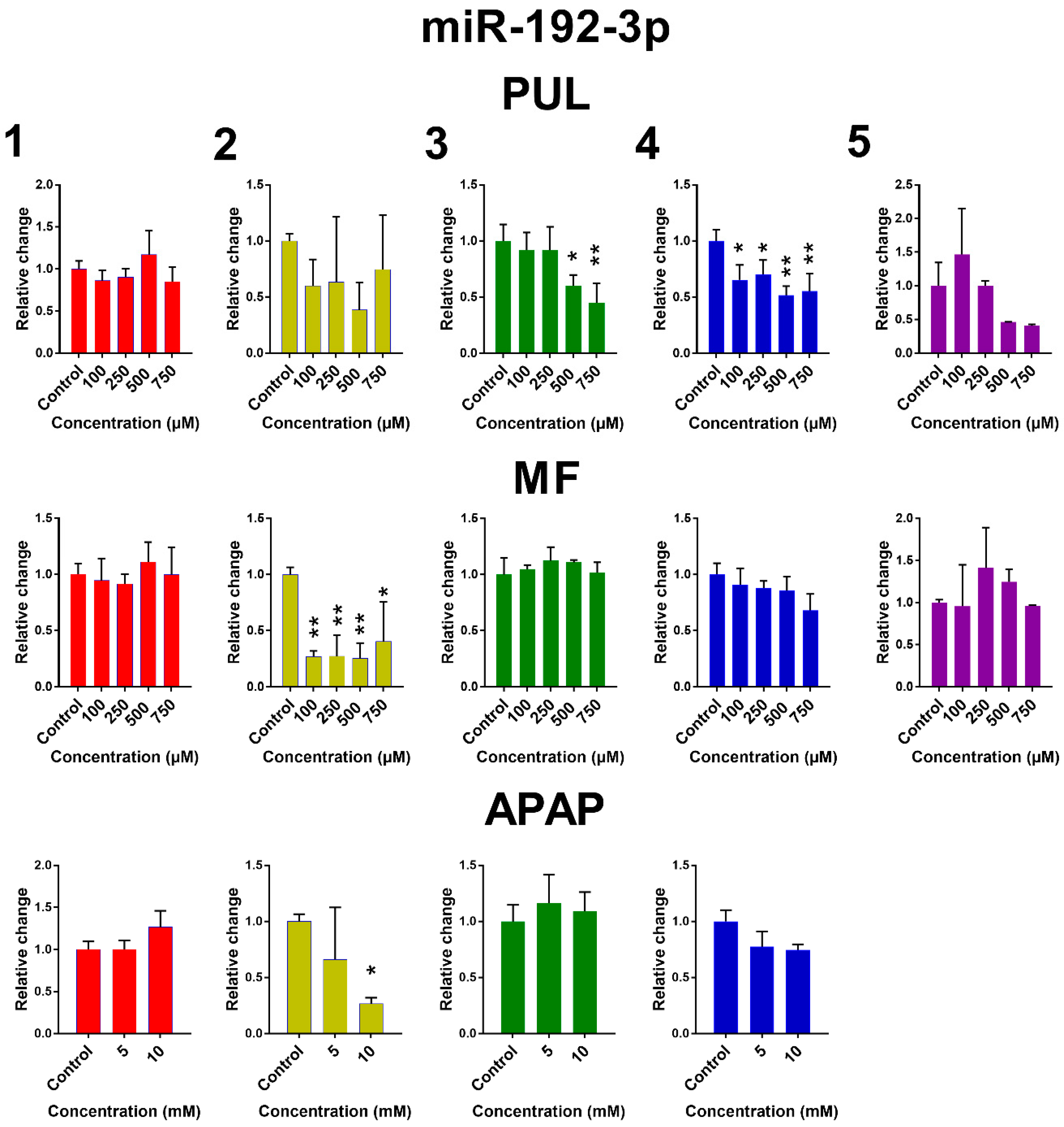
2.2. The Effect of PUL, MF and APAP on the Expression of Selected miRNAs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Ethics Statement
4.3. Human Liver
4.4. Precision-Cut Liver Slice Preparation and Experimental Treatment
4.5. ATP Determination
4.6. Gene Expression
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| APAP | acetaminophen |
| ATP | adenosine triphosphate |
| DILI | drug-induced liver injury |
| EC50 | half-maximal effective concentration |
| MF | R-menthofuran |
| miRNA | microRNA |
| PCLS | precision-cut liver slices |
| PUL | R-pulegone |
| qPCR | quantitative polymerase chain reaction |
References
- Gordon, P.; Khojasteh, S.C. A decades-long investigation of acute metabolism-based hepatotoxicity by herbal constituents: A case study of pennyroyal oil. Drug Metab. Rev. 2015, 47, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Phua, D.H.; Zosel, A.; Heard, K. Dietary supplements and herbal medicine toxicities-when to anticipate them and how to manage them. Int. J. Emerg. Med. 2009, 2, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Gordon, W.P.; Forte, A.J.; McMurtry, R.J.; Gal, J.; Nelson, S.D. Hepatotoxicity and Pulmonary Toxicity of Pennyroyal Oil and Its Constituent Terpenes in the Mouse. Toxicol. Appl. Pharmacol. 1982, 65, 413–424. [Google Scholar] [CrossRef]
- Moorthy, B.; Madyastha, P.; Madyastha, K.M. Hepatotoxicity of pulegone in rats—Its effects on microsomal-enzymes, invivo. Toxicology 1989, 55, 327–337. [Google Scholar] [CrossRef]
- Gordon, W.P.; Huitric, A.C.; Seth, C.L.; McClanahan, R.H.; Nelson, S.D. The Metabolism of the Abortifacient Terpene, (R)-(+)-Pulegone, to a Proximate Toxin, Menthofuran. Drug Metab. Dispos. 1987, 15, 589–594. [Google Scholar] [PubMed]
- Olinga, P.; Schuppan, D. Precision-cut liver slices: A tool to model the liver ex vivo. J. Hepatol. 2013, 58, 1252–1253. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.X.; Ewing, L.E.; Koturbash, I.; Gurley, B.J.; Miousse, I.R. MicroRNAs as biomarkers for liver injury: Current knowledge, challenges and future prospects. Food Chem. Toxicol. 2017, 110, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Antoine, D.J.; Dear, J.W.; Lewis, P.S.; Platt, V.; Coyle, J.; Masson, M.; Thanacoody, R.H.; Gray, A.J.; Webb, D.J.; Moggs, J.G.; et al. Mechanistic biomarkers provide early and sensitive detection of acetaminophen-induced acute liver injury at first presentation to hospital. Hepatology 2013, 58, 777–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Guo, A.; Xu, J.; Yao, J.; Chen, H.; Wang, F.; Zhu, C. Evaluation of a combinational use of serum microRNAs as biomarkers for liver diseases. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Shi, Q.; Mattes, W.B.; Mendrick, D.L.; Yang, X. Translating extracellular microRNA into clinical biomarkers for drug-induced toxicity: From high-throughput profiling to validation. Biomark. Med. 2015, 9, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, A.E.; Cisilotto, J.; Creczynski-Pasa, T.B.; de Lucca Schiavon, L. Circulating miRNAs in nontumoral liver diseases. Pharmacol. Res. 2018, 128, 274–287. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EMA). Public Statement on the Use of Herbal Medicinal Products Containing Pulegone and Menthofuran; EMA: Canary Wharf, UK, 2016.
- Anderson, I.B.; Mullen, W.H.; Meeker, J.E.; KhojastehBakht, S.C.; Oishi, S.; Nelson, S.D.; Blanc, P.D. Pennyroyal toxicity: Measurement of toxic metabolite levels in two cases and review of the literature. Ann. Intern. Med. 1996, 124, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Thomassen, D.; Slattery, J.T.; Nelson, S.D. Contribution of Menthofuran to the hepatotoxicity of Pulegone—Assessment based on matched area under the curve and on matched time course. J. Pharmacol. Exp. Ther. 1988, 244, 825–829. [Google Scholar] [PubMed]
- Mizutani, T.; Nomura, H.; Nakanishi, K.; Fujita, S. Effects of Drug-Metabolism Modifiers on Pulegone-Induced Hepatotoxicity in Mice. Res. Commun. Chem. Pathol. Pharmacol. 1987, 58, 75–83. [Google Scholar] [PubMed]
- Madyastha, K.M.; Raj, C.P. Effects of menthofuran, a monoterpene furan on rat liver microsomal enzymes, in vivo. Toxicology 1994, 89, 119–125. [Google Scholar] [CrossRef]
- Loosen, S.H.; Schueller, F.; Trautwein, C.; Roy, S.; Roderburg, C. Role of circulating microRNAs in liver diseases. World J. Hepatol. 2017, 9, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Khojasteh-Bakht, S.C.; Chen, W.Q.; Koenigs, L.L.; Peter, R.M.; Nelson, S.D. Metabolism of (R)-(+)-pulegone and (R)-(+)-menthofuran by human liver cytochrome P-450s: Evidence for formation of a furan epoxide. Drug Metab. Dispos. 1999, 27, 574–580. [Google Scholar] [PubMed]
- Khojasteh, S.C.; Oishi, S.; Nelson, S.D. Metabolism and Toxicity of Menthofuran in Rat Liver Slices and in Rats. Chem. Res. Toxicol. 2010, 23, 1824–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, L.T.; Chan, D.; King, R.G. Investigation of the effects of peppermint oil and valerian on rat liver and cultured human liver cells. Clin. Exp. Pharmacol. Physiol. 2003, 30, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Lassila, T.; Rousu, T.; Mattila, S.; Chesne, C.; Pelkonen, O.; Turpeinen, M.; Tolonen, A. Formation of GSH-trapped reactive metabolites in human liver microsomes, S9 fraction, HepaRG-cells, and human hepatocytes. J. Pharm. Biomed. Anal. 2015, 115, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Jemnitz, K.; Veres, Z.; Monostory, K.; Kobori, L.; Vereczkey, L. Interspecies differences in acetaminophen sensitivity of human, rat, and mouse primary hepatocytes. Toxicol. In Vitro 2008, 22, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Hadi, M.; Dragovic, S.; van Swelm, R.; Herpers, B.; van de Water, B.; Russel, F.G.M.; Commandeur, J.N.M.; Groothuis, G.M.M. AMAP, the alleged non-toxic isomer of acetaminophen, is toxic in rat and human liver. Arch. Toxicol. 2013, 87, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Jetten, M.J.A.; Claessen, S.M.; Dejong, C.H.C.; Lahoz, A.; Castell, J.V.; van Delft, J.H.M.; Kleinjans, J.C.S. Interindividual variation in response to xenobiotic exposure established in precision-cut human liver slices. Toxicology 2014, 323, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Vickers, A.E.M.; Fisher, R.L. Evaluation of drug-induced injury and human response in precision-cut tissue slices. Xenobiotica 2013, 43, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Granitzny, A.; Knebel, J.; Schaudien, D.; Braun, A.; Steinberg, P.; Dasenbrock, C.; Hansen, T. Maintenance of high quality rat precision cut liver slices during culture to study hepatotoxic responses: Acetaminophen as a model compound. Toxicol. In Vitro 2017, 42, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Tokito, Y.; Dekura, E.; Kawai, Y. Technical Report: Application of Rat Precision-cut Liver Slices for Toxicity Assessment In Vitro. J. Toxicol. Pathol. 1999, 12, 71. [Google Scholar] [CrossRef]
- Matouskova, P.; Bartikova, H.; Bousova, I.; Hanusova, V.; Szotakova, B.; Skalova, L. Reference Genes for Real-Time PCR Quantification of Messenger RNAs and MicroRNAs in Mouse Model of Obesity. PLoS ONE 2014, 9, e86033. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matouskova, P. microRNAs and Reference Gene Methodology. In Handbook of Nutrition, Diet, and Epigenetics; Vinood, P., Victor, P., Eds.; Springer: Basel, Switzerland, 2017. [Google Scholar]
- Wu, C.; Zhang, J.; Cao, X.; Yang, Q.; Xia, D. Effect of Mir-122 on Human Cholangiocarcinoma Proliferation, Invasion, and Apoptosis Through P53 Expression. Med. Sci. Monit. 2016, 22, 2685–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.N.; Han, C.; Zhang, J.Q.; Song, K.; Wang, Y.; Wu, T. Deletion of Mir155 Prevents Fas-Induced Liver Injury through Up-Regulation of Mcl-1. Am. J. Pathol. 2015, 185, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.M.; Gilchrist, D.S.; Nijjar, J.; Araldi, E.; Ramirez, C.M.; Lavery, C.A.; Fernandez-Hernando, C.; McInnes, I.B.; Kurowska-Stolarska, M. MiR-155 Has a Protective Role in the Development of Non-Alcoholic Hepatosteatosis in Mice. PLoS ONE 2013, 8, e72324. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.L.; Jia, J.S.; Du, T.; Li, W.; Wang, X.Y.; Wei, J.Q.; Lin, X.; Zeng, H.; Yao, L.P.; Chen, X.B.; et al. Overexpression of miR-155 in the Liver of Transgenic Mice Alters the Expression Profiling of Hepatic Genes Associated with Lipid Metabolism. PLoS ONE 2015, 10, e0118417. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Csak, T.; Saha, B.; Zatsiorsky, J.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. The pro-inflammatory effects of miR-155 promote liver fibrosis and alcohol-induced steatohepatitis. J. Hepatol. 2016, 64, 1378–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.Z.; Guo, G.F.; Yin, C.X.; Jiang, C.; Wang, F.; Qiu, H.J.; Chen, X.X.; Rong, R.M.; Zhang, B.; Xia, L.P. Gamma-glutamyl transpeptidase level is a novel adverse prognostic indicator in human metastatic colorectal cancer. Colorectal Dis. 2013, 15, e443–e452. [Google Scholar] [CrossRef] [PubMed]
- Ciombor, K.K.; Goff, L.W. Current therapy and future directions in biliary tract malignancies. Curr. Treat. Opt. Oncol. 2013, 14, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Hadi, M.; Westra, I.M.; Starokozhko, V.; Dragovic, S.; Merema, M.T.; Groothuis, G.M. Human precision-cut liver slices as an ex vivo model to study idiosyncratic drug-induced liver injury. Chem. Res. Toxicol. 2013, 26, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Han, C.Y.; Kim, J.Y.; Cho, S.S.; Kim, Y.S.; Koo, J.H.; Lee, J.M.; Lim, S.C.; Kang, K.W.; Kim, J.S.; et al. Gα(12) overexpression induced by miR-16 dysregulation contributes to liver fibrosis by promoting autophagy in hepatic stellate cells. J. Hepatol. 2018, 68, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Song, J.N.; Bai, Z.G.; Han, W.; Zhang, J.; Meng, H.; Bi, J.T.; Ma, X.M.; Han, S.W.; Zhang, Z.T. Identification of Suitable Reference Genes for qPCR Analysis of Serum microRNA in Gastric Cancer Patients. Dig. Dis. Sci. 2012, 57, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Rinnerthaler, G.; Hackl, H.; Gampenrieder, S.P.; Hamacher, F.; Hufnagl, C.; Hauser-Kronberger, C.; Zehentmayr, F.; Fastner, G.; Sedlmayer, F.; Mlineritsch, B.; et al. miR-16-5p Is a Stably-Expressed Housekeeping MicroRNA in Breast Cancer Tissues from Primary Tumors and from Metastatic Sites. Int. J. Mol. Sci. 2016, 17, E156. [Google Scholar] [CrossRef] [PubMed]
- Das, M.K.; Andreassen, R.; Haugen, T.B.; Furu, K. Identification of Endogenous Controls for Use in miRNA Quantification in Human Cancer Cell Lines. Cancer Genom. Proteom. 2016, 13, 63–68. [Google Scholar]
- Wang, X.R.; Liao, Z.J.; Bai, Z.M.; He, Y.; Duan, J.; Wei, L.Y. MiR-93-5p Promotes Cell Proliferation through Down-Regulating PPARGC1A in Hepatocellular Carcinoma Cells by Bioinformatics Analysis and Experimental Verification. Genes 2018, 9, E51. [Google Scholar] [CrossRef] [PubMed]
- De Conti, A.; Ortega, J.F.; Tryndyak, V.; Dreval, K.; Moreno, F.S.; Rusyn, I.; Beland, F.A.; Pogribny, I.P. MicroRNA deregulation in nonalcoholic steatohepatitis-associated liver carcinogenesis. Oncotarget 2017, 8, 88517–88528. [Google Scholar] [CrossRef] [PubMed]
- Lanford, R.E.; Hildebrandt-Eriksen, E.S.; Petri, A.; Persson, R.; Lindow, M.; Munk, M.E.; Kauppinen, S.; Orum, H. Therapeutic Silencing of MicroRNA-122 in Primates with Chronic Hepatitis C Virus Infection. Science 2010, 327, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.G.; Wang, C.M.; Tian, C.; Wang, Y.; Li, L.; Sun, W.S.; Li, R.F.; Liu, Y.G. miR-122 inhibits viral replication and cell proliferation in hepatitis B virus-related hepatocellular carcinoma and targets NDRG3. Oncol. Rep. 2011, 26, 1281–1286. [Google Scholar] [PubMed]
- Coulouarn, C.; Factor, V.M.; Andersen, J.B.; Durkin, M.E.; Thorgeirsson, S.S. Loss of miR-122 expression in liver cancer correlates with suppression of the hepatic phenotype and gain of metastatic properties. Oncogene 2009, 28, 3526–3536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.; Han, C.; Zhang, J.Q.; Lu, D.D.; Dash, S.; Feitelson, M.; Lim, K.; Wu, T. Epigenetic Regulation of MicroRNA-122 by Peroxisome Proliferator Activated Receptor-gamma and Hepatitis B Virus X Protein in Hepatocellular Carcinoma Cells. Hepatology 2013, 58, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Chen, H.; Gao, S.S.; Qiu, S.L.; Zhou, H.; Yu, M.X.; Tu, J.C. Differential expression of plasma microRNA-125b in hepatitis B virus-related liver diseases and diagnostic potential for hepatitis B virus-induced hepatocellular carcinoma. Hepatol. Res. 2017, 47, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Giray, B.G.; Emekdas, G.; Tezcan, S.; Ulger, M.; Serin, M.S.; Sezgin, O.; Altintas, E.; Tiftik, E.N. Profiles of serum microRNAs; miR-125b-5p and miR223-3p serve as novel biomarkers for HBV-positive hepatocellular carcinoma. Mol. Biol. Rep. 2014, 41, 4513–4519. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhu, Y.H.; Han, H.; Zhang, Q.; Cui, K.S.; Shen, H.S.; Zhang, J.X.; Yan, J.; Prochownik, E.; Li, Y.J. MicroRNA-148a deficiency promotes hepatic lipid metabolism and hepatocarcinogenesis in mice. Cell Death Dis. 2017, 8, e2916. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wen, H.Q.; Jing, L.; Yang, Y.J.; Wang, W.J.; Liang, X.; Nan, K.J.; Yao, Y.; Tian, T. MicroRNA-155-5p promotes hepatocellular carcinoma progression by suppressing PTEN through the PI3K/Akt pathway. Cancer Sci. 2017, 108, 620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Liu, G.; Tang, W. MicroRNA-182-5p Ameliorates Liver Ischemia-Reperfusion Injury by Suppressing Toll-Like Receptor 4. Transplant. Proc. 2016, 48, 2809–2814. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.W.; Shen, J.L.; Wang, C.; Yang, L.L.; Zhang, X.W. MicroRNA-182 downregulates metastasis suppressor 1 and contributes to metastasis of hepatocellular carcinoma. BMC Cancer 2012, 12, 227. [Google Scholar] [CrossRef] [PubMed]
- Assal, R.A.; Tayebi, H.M.E.; Hosny, K.A.; Esmat, G.; Abdelaziz, A.I. A pleiotropic effect of the single clustered hepatic metastamiRs miR-96-5p and miR-182-5p on insulin-like growth factor II, insulin-like growth factor-1 receptor and insulin-like growth factor-binding protein-3 in hepatocellular carcinoma. Mol. Med. Rep. 2015, 12, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Krattinger, R.; Bostrom, A.; Schioth, H.B.; Thasler, W.E.; Mwinyi, J.; Kullak-Ublick, G.A. microRNA-192 suppresses the expression of the farnesoid X receptor. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G1044–G1051. [Google Scholar] [CrossRef] [PubMed]
- Raitoharju, E.; Seppala, I.; Lyytikainen, L.P.; Viikari, J.; Ala-Korpela, M.; Soininen, P.; Kangas, A.J.; Waldenberger, M.; Klopp, N.; Illig, T.; et al. Blood hsa-miR-122-5p and hsa-miR-885-5p levels associate with fatty liver and related lipoprotein metabolism—The Young Finns Study. Sci. Rep. 2016, 6, 38262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vliegenthart, A.D.B.; Shaffer, J.M.; Clarke, J.I.; Peeters, L.E.J.; Caporali, A.; Bateman, D.N.; Wood, D.M.; Dargan, P.I.; Craig, D.G.; Moore, J.K.; et al. Comprehensive microRNA profiling in acetaminophen toxicity identifies novel circulating biomarkers for human liver and kidney injury. Sci. Rep. 2015, 5, 15501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, J.H.; Tian, Y.P.; Wen, X.Y.; Zhang, W.H.; Zhang, P.J.; Gao, J.; Run, W.; Tian, L.Y.; Jia, X.W.; Gao, Y.H. Serum microRNA characterization identifies miR-885-5p as a potential marker for detecting liver pathologies. Clin. Sci. 2011, 120, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Yin, J.; Yang, J.; Shen, W.Z.; Zhang, C.Y.; Mou, W.J.; Luo, J.H.; Yan, H.; Sun, P.Q.; Luo, Y.P.; et al. miR-885-5p suppresses hepatocellular carcinoma metastasis and inhibits Wnt/beta-catenin signaling pathway. Oncotarget 2016, 7, 75038–75051. [Google Scholar] [PubMed]
- Lou, G.; Ma, N.; Xu, Y.; Jiang, L.; Yang, J.; Wang, C.X.; Jiao, Y.F.; Gao, X. Differential distribution of U6 (RNU6-1) expression in human carcinoma tissues demonstrates the requirement for caution in the internal control gene selection for microRNA quantification. Int. J. Mol. Med. 2015, 36, 1400–1408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Sample | Gender (Age) | Reason of Surgery | Long-Term Pharmacotherapy |
|---|---|---|---|
| 1 | Male (69) | Colorectal carcinoma | Acetylsalicylic acid, nitrendipine |
| 2 | Female (73) | Colorectal carcinoma | Lercanidipine, furosemide, perindopril, nadroparin |
| 3 | Female (67) | Cholangiocellular carcinoma | Nadroparin |
| 4 | Female (65) | Colorectal carcinoma | Amlodipine |
| 5 | Female (57) | Colorectal carcinoma with liver metastasis | none |
| Gene | Primer Sequences 5′-3′ | Function |
|---|---|---|
| hsa-miR-16-5p | F: ACAGCCTAGCAGCACGTAAAT RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACC GCCAA | Modulation of expression alters hepatic stellate cells’ autophagy, reference gene [41,42,43] |
| hsa-miR-93-5p | F: GTCAATCAAAGTGCTGTTCGTG RT: GTCTCCTCTGGTGCAGG GTCCGAGGTATTCGCACCAGAGGAGAC | Aberrant expression probably plays a role in hepatoma development, reference gene [42,44,45,46] |
| hsa-miR-122-5p | F: AGACCTCCTGTGCAAATCTATG RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACC AAACA | Control of diverse aspects of hepatic function (lipid metabolism) or dysfunction (viral infection, hepatocarcinogenesis), potential liver injury biomarker [47,48,49,50] |
| hsa-miR-125b-5p | F: AGACCTCCTGTGCAAATCTATG RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACC AAACA | Tumor suppressor, potential diagnostic tool for hepatitis B virus-induced hepatocellular carcinoma [51,52] |
| hsa-miR-148a-3p | F: GAGAATTCAGTGCACTACAGA RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACA CAAAG | Downregulated in hepatocellular carcinoma, deficiency enhances hepatic steatosis [53] |
| hsa-miR-155-5p | F: GGCCCTTTAATGCTAATCGTGA RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACA CCCCT | Important role in immune and inflammatory processes, oncogenic [54,55] |
| hsa-miR-182-5p | F: GATCACTTTGGCAATGGTAGAAC RT: GTCTCCTCTGGTGCAGG GTCCGAGGTATTCGCACCAGAG GAGACAGTGTG | Attenuates liver ischemia-reperfusion injury, plays a role in hepatocellular carcinoma and its metastasis [56,57,58] |
| hsa-miR-192-3p | F: ACGTGTCTGCCAATTCCATAG RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACT ATTTA | Suppresses farnesoid X receptor expression in adenocarcinoma cell lines [59] |
| hsa-miR-885-5p | F: GAGACATCCATTAC CTACCC RT: GTCTCCTCTGGTGCAGGGTCC GAGGTATTCGCACCAGAGGAGACA GAGGC | Lipoprotein and lipid metabolism, suppression of hepatocellular carcinoma metastasis, potential liver injury biomarker [60,61,62,63] |
| snU6 | F: GCTCGCTTCGGCAGCACA RT: AACGCTTCACGAATTTGCGTG | Reference gene [64] |
| Universal | R: GAGGTATTCGCACCAGAGGA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zárybnický, T.; Matoušková, P.; Lancošová, B.; Šubrt, Z.; Skálová, L.; Boušová, I. Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen. Int. J. Mol. Sci. 2018, 19, 1805. https://doi.org/10.3390/ijms19061805
Zárybnický T, Matoušková P, Lancošová B, Šubrt Z, Skálová L, Boušová I. Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen. International Journal of Molecular Sciences. 2018; 19(6):1805. https://doi.org/10.3390/ijms19061805
Chicago/Turabian StyleZárybnický, Tomáš, Petra Matoušková, Bibiána Lancošová, Zdeněk Šubrt, Lenka Skálová, and Iva Boušová. 2018. "Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen" International Journal of Molecular Sciences 19, no. 6: 1805. https://doi.org/10.3390/ijms19061805