Yeast-Based Screen to Identify Natural Compounds with a Potential Therapeutic Effect in Hailey-Hailey Disease


Abstract
:
1. Introduction
2. Results
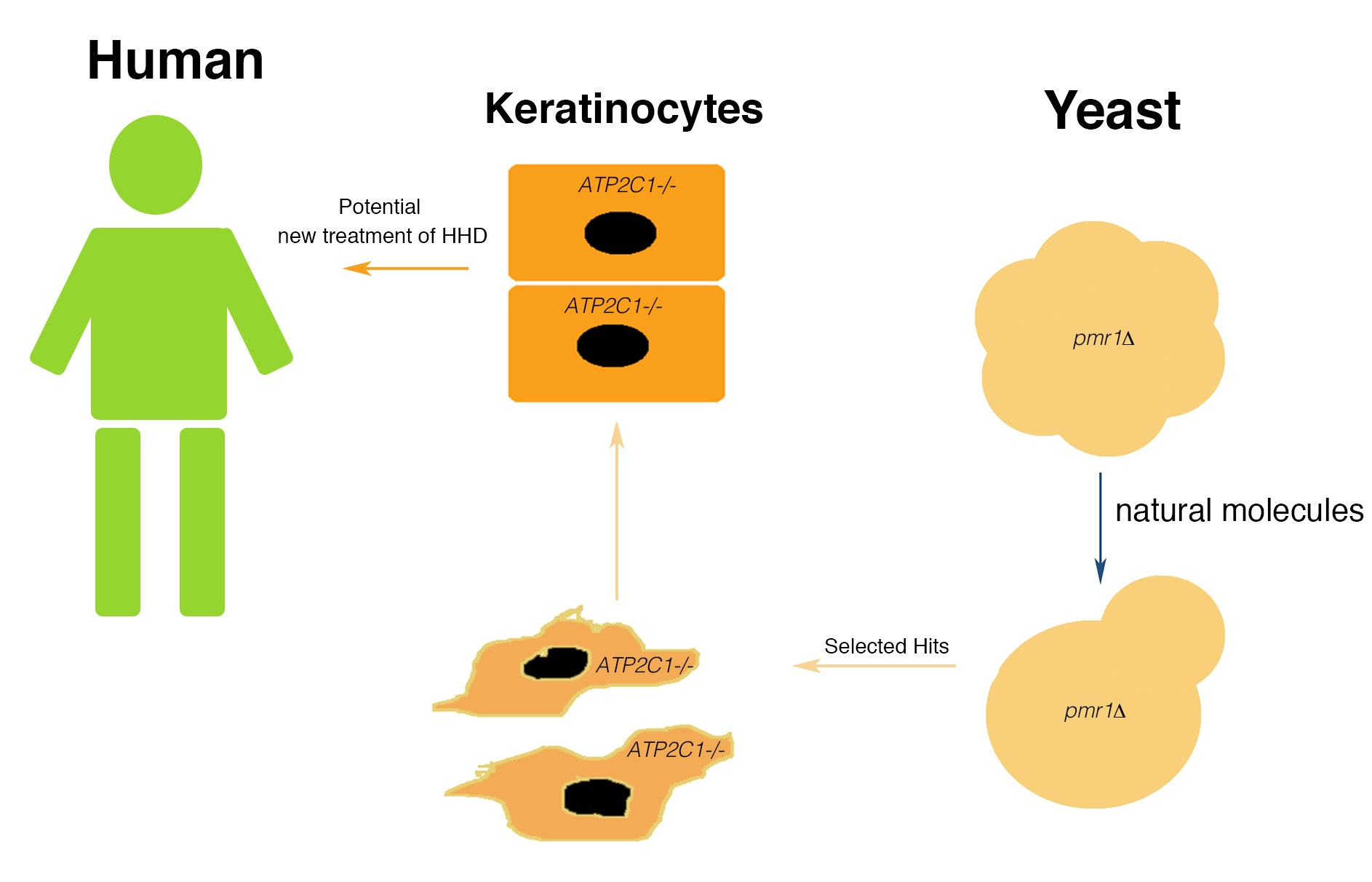
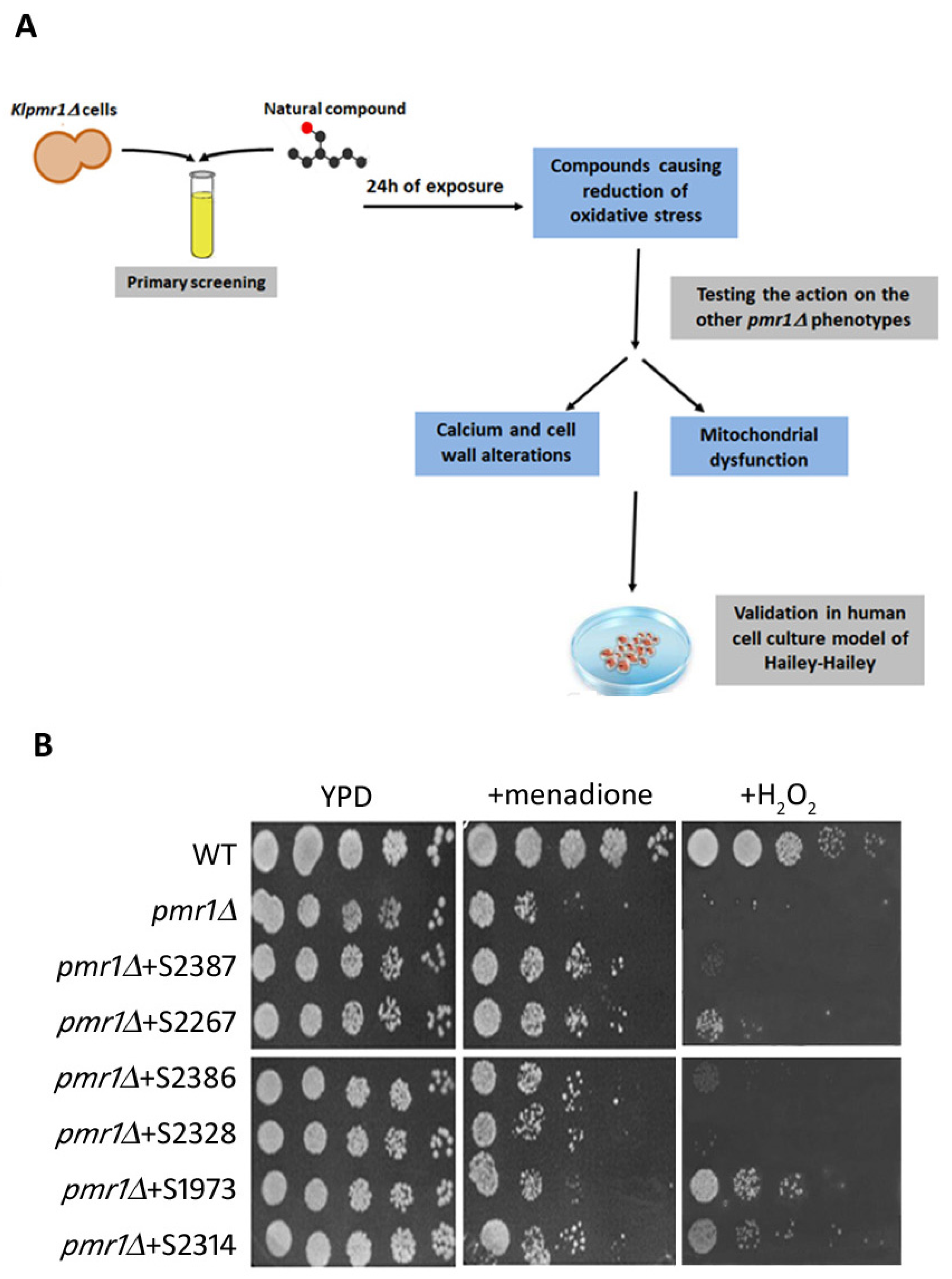
2.1. Primary Screen of Chemical Libraries Using KLPMR1-Based Assay
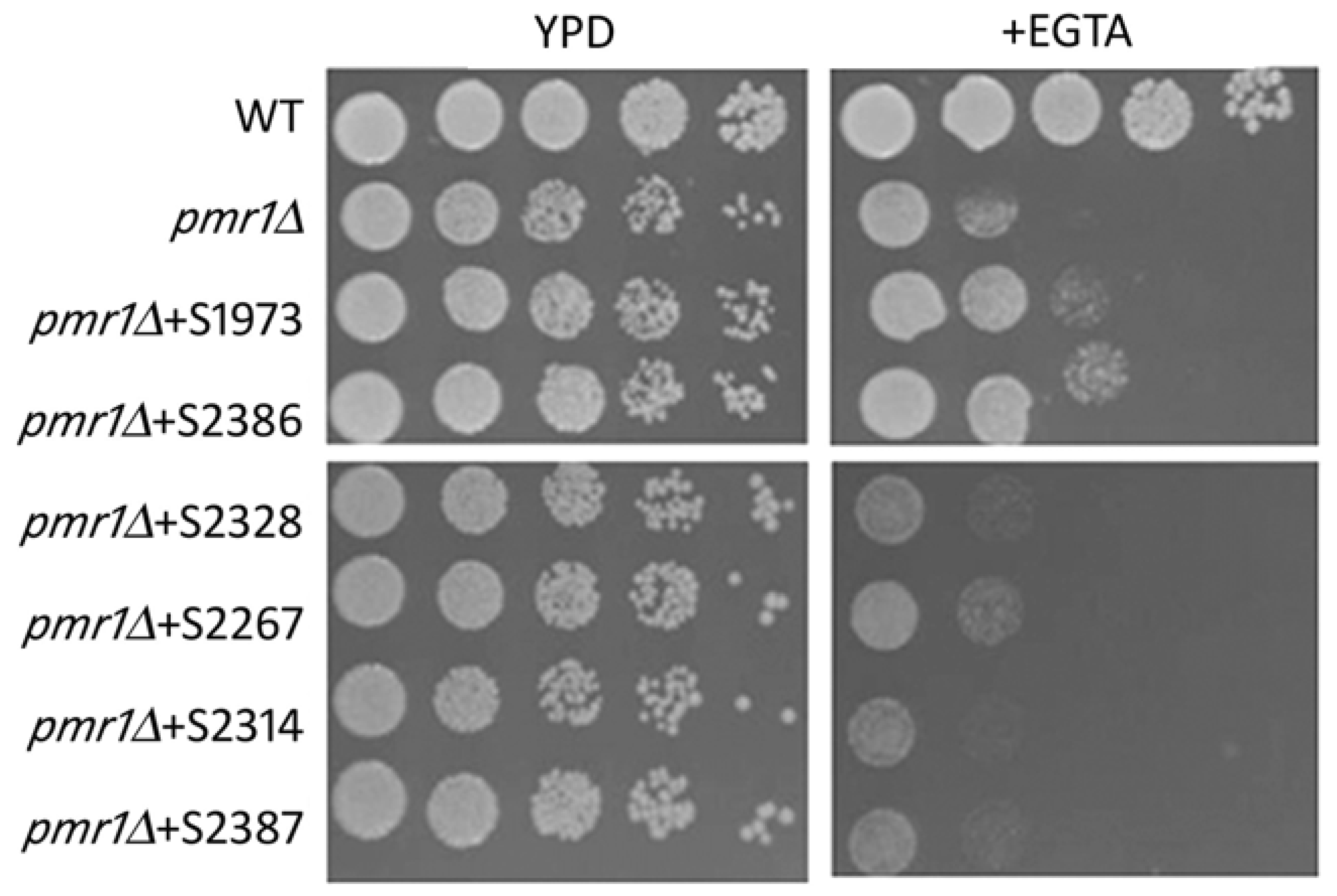
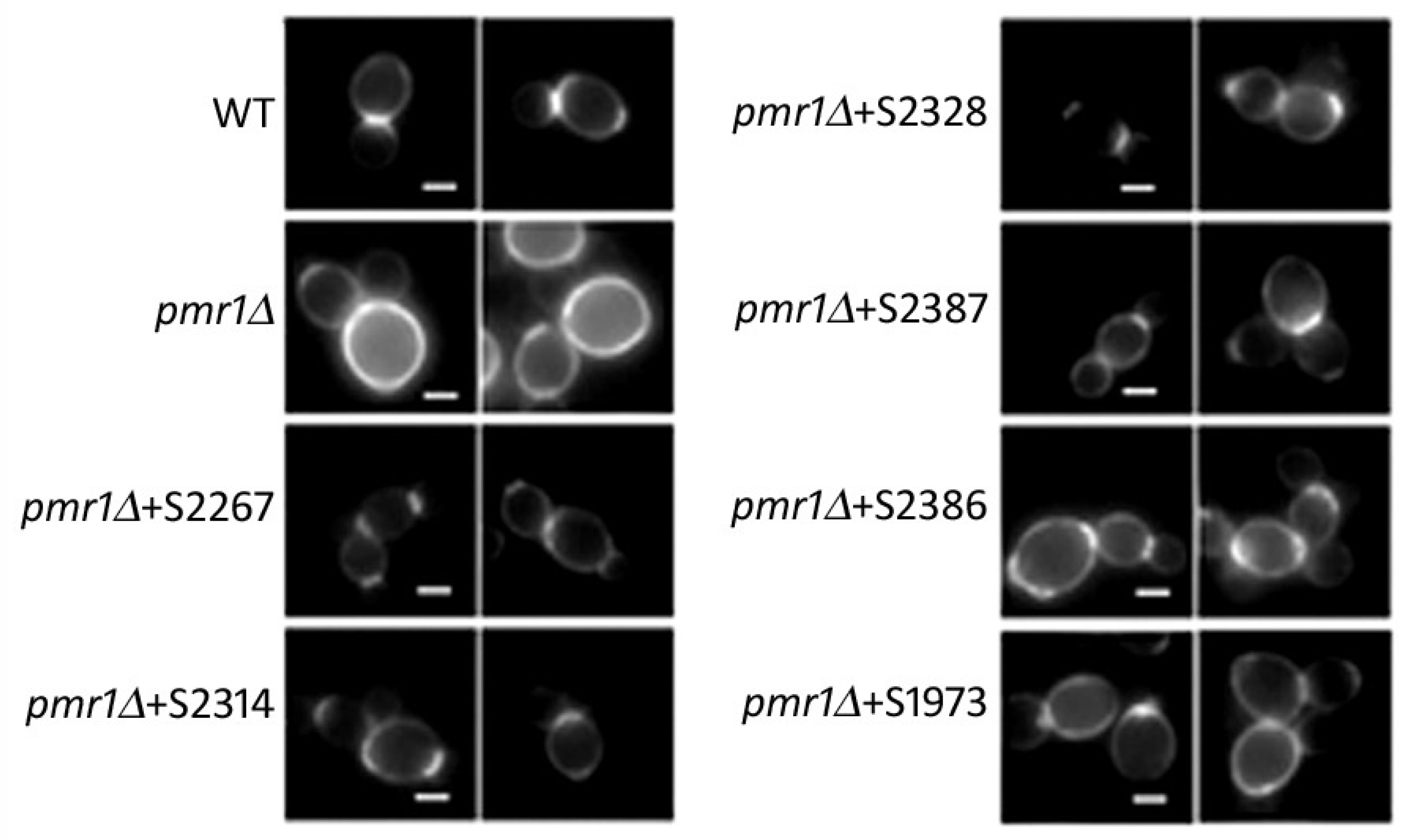
2.2. Yeast-Hits Rescue Multiple Defects in pmr1Δ Cells
2.3. Cell Wall Phenotype
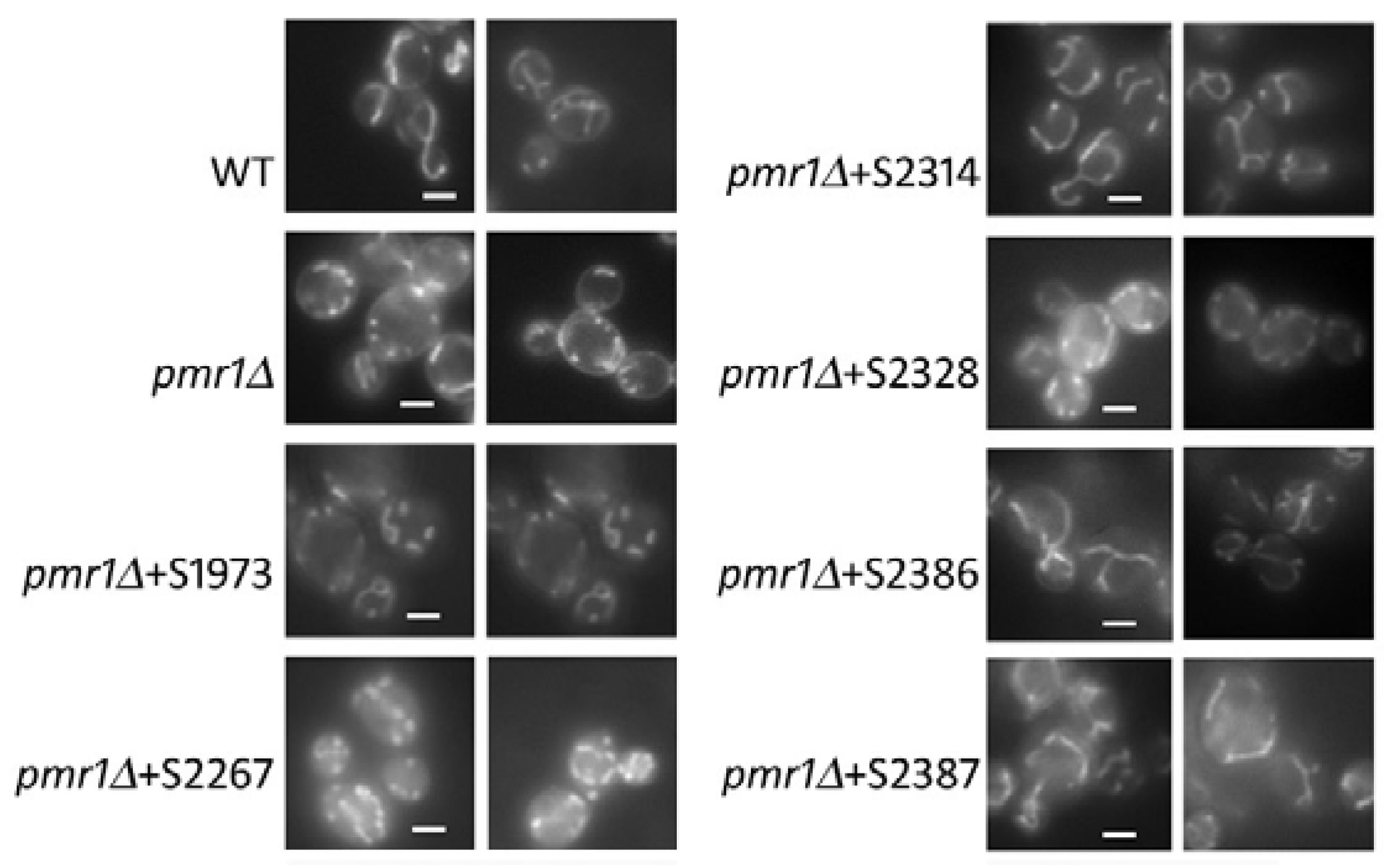
2.4. Mitochondrial Morphology
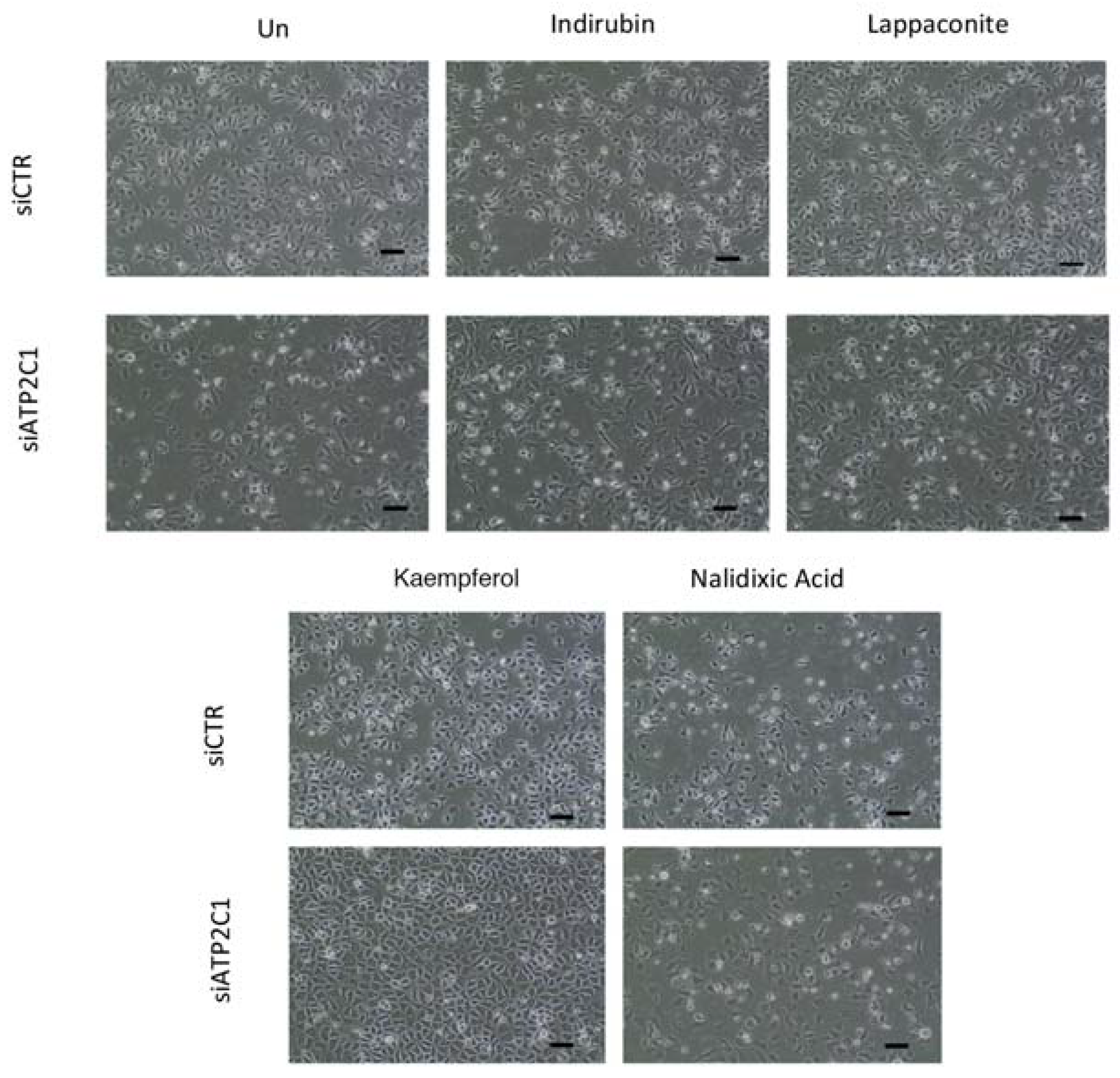
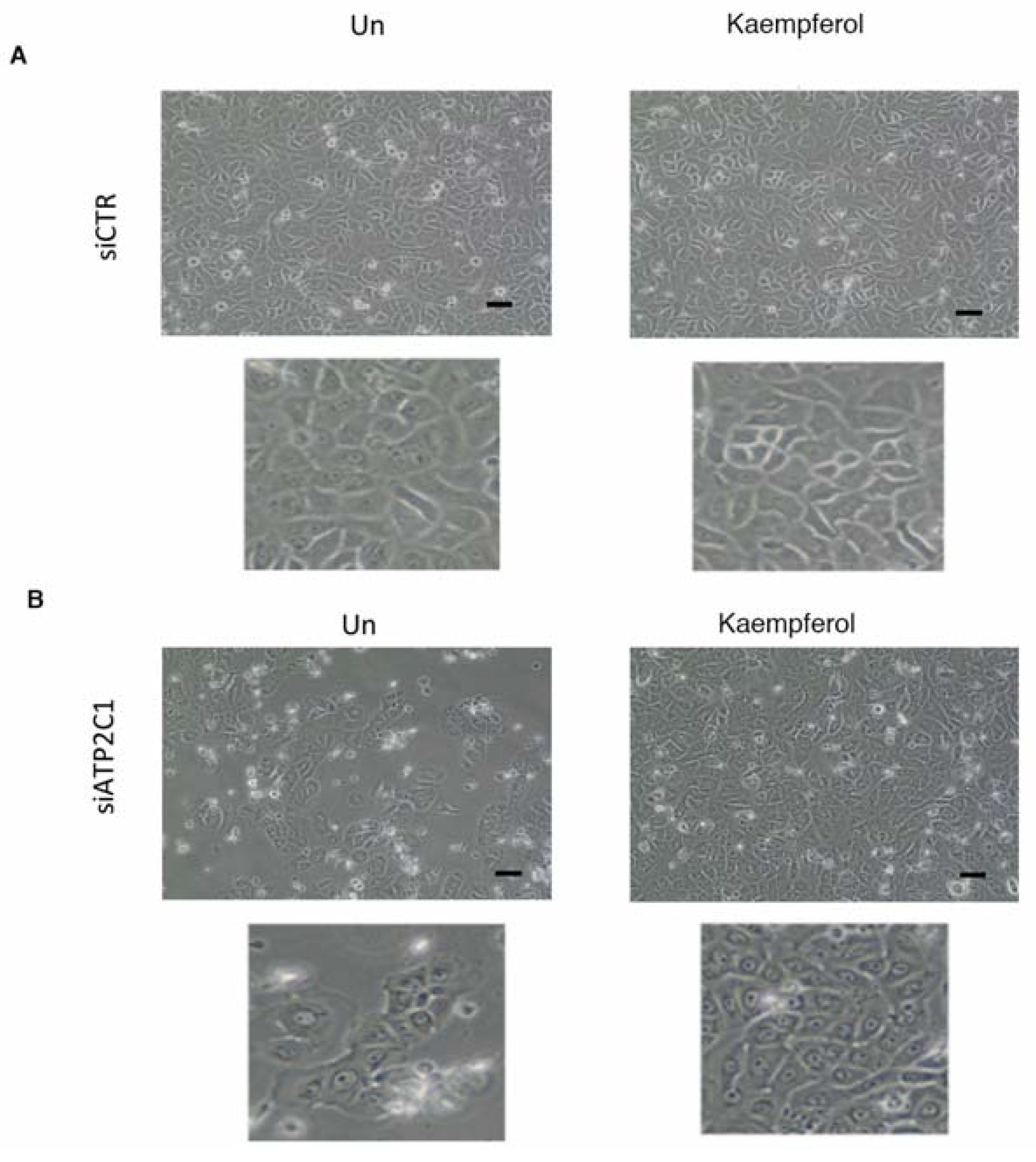
2.5. Drugs Active in the Yeast-Based Assay Were Also Active in Human Cells
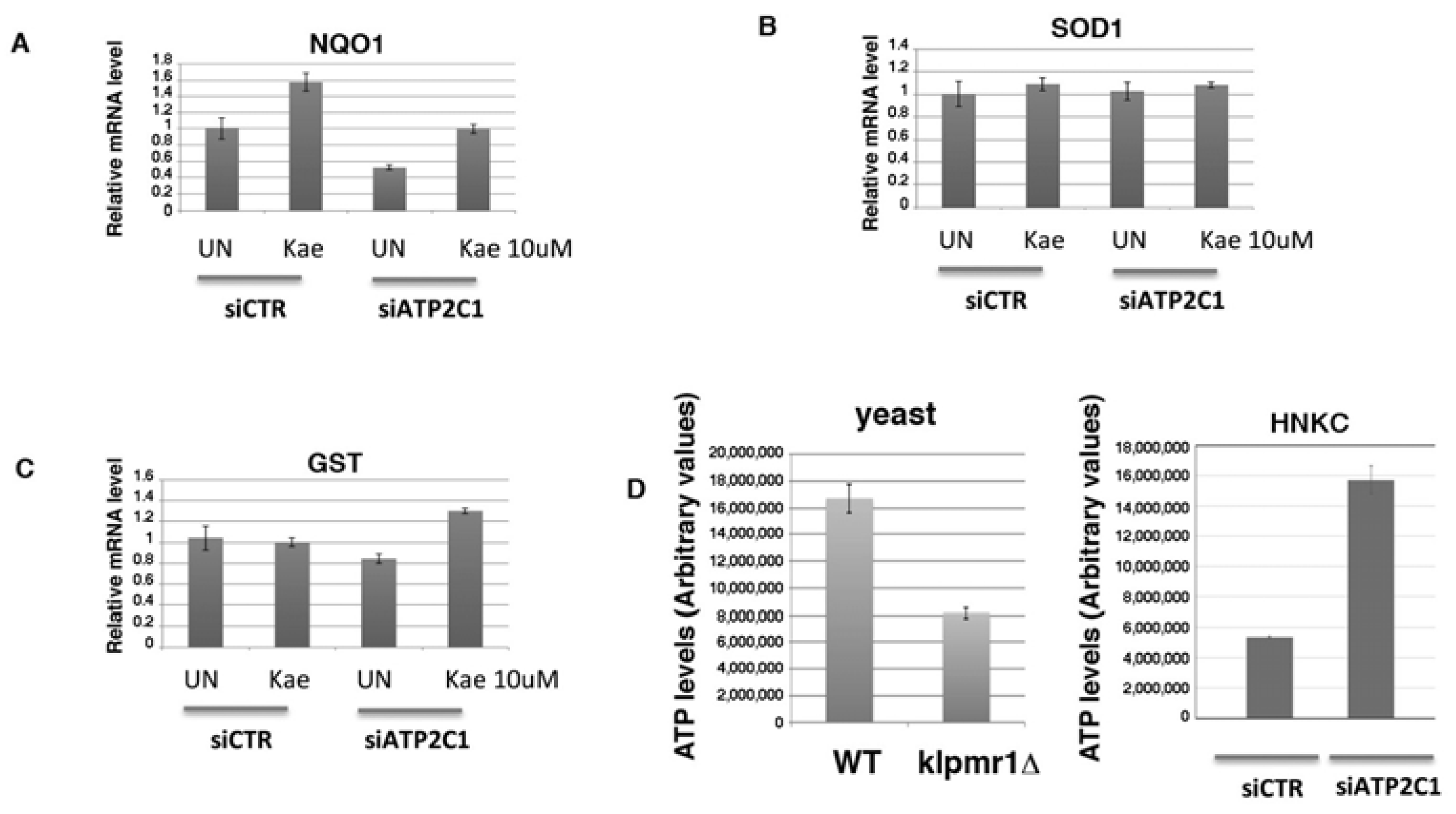
2.6. Potential Mechanism of Kaempferol against ATP2C1-Induced Oxidative-Stress through Regulation of Nuclear Factor Erythroid-2-Related Factor 2 Signaling
2.7. Increased Mitochondrial Activity as a Source of Oxidative Stress in ATP2C1-Defective Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Yeast Strains, Growth Conditions
4.2. Library Screen
4.3. Fluorescence Microscopy
4.4. Primary Human Keratinocytes
4.5. Cell Culture and Transfection
4.6. Reagents and Immunoblotting
4.7. RNA Analysis and Reverse Transcriptase-Polymerase Chain Reaction
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kellermayer, R. Hailey-Hailey disease as an orthodisease of PMR1 deficiency in Saccharomyces cerevisiae. FEBS Lett. 2005, 579, 2021–2025. [Google Scholar] [CrossRef] [PubMed]
- Raiko, L.; Siljamäki, E.; Mahoney, M.G.; Putaala, H.; Suominen, E.; Peltonen, J.; Peltonen, S. Hailey-Hailey disease and tight junctions: Claudins 1 and 4 are regulated by ATP2C1 gene encoding Ca2+/Mn2+ ATPase SPCA1 in cultured keratinocytes. Exp. Dermatol. 2012, 21, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Bonifas, J.M.; Beech, J.; Bench, G.; Shigihara, T.; Ogawa, H.; Ikeda, S.; Mauro, T.; Epstein, E.H., Jr. Mutations in ATP2C1, encoding a calcium pump, cause Hailey-Hailey disease. Nat. Genet. 2000, 24, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Sudbrak, R.; Brown, J.; Dobson-Stone, C.; Carter, S.; Ramser, J.; White, J.; Healy, E.; Dissanayake, M.; Larrègue, M.; Perrussel, M.; et al. Hailey-Hailey disease is caused by mutations in ATP2C1 encoding a novel Ca2+ pump. Hum. Mol. Genet. 2000, 9, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Wuytack, F.; Raeymaekers, L.; Missiaen, L. PMR1/SPCA Ca2+ pumps and the role of the Golgi apparatus as a Ca2+ store. Pflugers Arch. 2003, 446, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Dobson-Stone, C.; Fairclough, R.; Dunne, E.; Brown, J.; Monaco, A.P.; Hovnanian, A.; Dissanayake, M.; Munro, C.S.; Strachan, T.; Burge, S.; et al. Hailey-Hailey disease: Molecular and clinical characterization of novel mutations in the ATP2C1 gene. J. Investig. Dermatol. 2002, 118, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Cialfi, S.; Le Pera, L.; De Blasio, C.; Mariano, G.; Palermo, R.; Zonfrilli, A.; Uccelletti, D.; Palleschi, C.; Biolcati, G.; Barbieri, L.; et al. The loss of ATP2C1 impairs the DNA damage response and induces altered skin homeostasis: Consequences for epidermal biology in Hailey-Hailey disease. Sci. Rep. 2016, 6, 31567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, H.; Bray, F.N.; Cervantes, J.; Falto Aizpurua, L.A. Management of familial benign chronic pemphigus. Clin. Cosmet. Investig. Dermatol. 2016, 9, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Poblete-Gutiérrez, P.; Wiederholt, T.; König, A.; Jugert, F.K.; Marquardt, Y.; Rübben, A.; Merk, H.F.; Happle, R.; Frank, J. Allelic loss underlies type 2 segmental Hailey-Hailey disease, providing molecular confirmation of a novel genetic concept. J. Clin. Investig. 2004, 114, 1467–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, M.R.; Erdag, G.; Shada, A.L.; Williams, M.E.; Slingluff, C.L., Jr.; Patterson, J.W. Two patients with Hailey-Hailey disease, multiple primary melanomas, and other cancers. Arch. Dermatol. 2011, 147, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Okunade, G.W.; Miller, M.L.; Azhar, M.; Andringa, A.; Sanford, L.P.; Doetschman, T.; Prasad, V.; Shull, G.E. Loss of the Atp2c1 secretory pathway Ca2+-ATPase (SPCA1) in mice causes Golgi stress, apoptosis, and midgestational death in homozygous embryos and squamous cell tumors in adult heterozygotes. J. Biol. Chem. 2007, 282, 26517–26527. [Google Scholar] [CrossRef] [PubMed]
- Mager, W.H.; Winderickx, J. Yeast as a model for medical and medicinal research. Trends Pharmacol. Sci. 2005, 26, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Perocchi, F.; Mancera, E.; Steinmetz, L.M. Systematic screens for human disease genes, from yeast to human and back. Mol. Biosyst. 2008, 4, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; Chervitz, S.A.; Cherry, J.M. Yeast as a model organism. Science 1997, 277, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Foury, F. Human genetic diseases: A cross-talk between man and yeast. Gene 1997, 195, 1–10. [Google Scholar] [CrossRef]
- Rudolph, H.K.; Antebi, A.; Fink, G.R.; Buckley, C.M.; Dorman, T.E.; LeVitre, J.; Davidow, L.S.; Mao, J.I.; Moir, D.T. The yeast secretory pathway is perturbed by mutations in PMR1, a member of a Ca2+ ATPase family. Cell 1989, 58, 133–145. [Google Scholar] [CrossRef]
- Uccelletti, D.; Mancini, P.; Farina, F.; Morrone, S.; Palleschi, C. Inactivation of the KIPMR1 gene of Kluyveromyces lactis results in defective cell-wall morphogenesis. Microbiology 1999, 145 Pt 5, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Voisset, C.; Garcia-Rodriguez, N.; Birkmire, A.; Blondel, M.; Wellinger, R.E. Using yeast to model calcium-related diseases: Example of the Hailey-Hailey disease. Biochim. Biophys. Acta 2014, 1843, 2315–2321. [Google Scholar] [CrossRef] [PubMed]
- Szigeti, R.; Kellermayer, R. Autosomal-dominant calcium ATPase disorders. J. Investig. Dermatol. 2006, 126, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- Uccelletti, D.; Farina, F.; Pinton, P.; Goffrini, P.; Mancini, P.; Talora, C.; Rizzuto, R.; Palleschi, C. The Golgi Ca2+-ATPase KlPmr1p function is required for oxidative stress response by controlling the expression of the heat-shock element HSP60 in Kluyveromyces lactis. Mol. Biol. Cell 2005, 16, 4636–4647. [Google Scholar] [CrossRef] [PubMed]
- Ficociello, G.; Zanni, E.; Cialfi, S.; Aurizi, C.; Biolcati, G.; Palleschi, C.; Talora, C.; Uccelletti, D. Glutathione S-transferase circle minus-subunit as a phenotypic suppressor of pmr1 Delta strain, the Kluyveromyces lactis model for Hailey-Hailey disease. BBA Mol. Cell Res. 2016, 1863, 2650–2657. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Cialfi, S.; Oliviero, C.; Ceccarelli, S.; Marchese, C.; Barbieri, L.; Biolcati, G.; Uccelletti, D.; Palleschi, C.; Barboni, L.; De Bernardo, C.; et al. Complex multipathways alterations and oxidative stress are associated with Hailey-Hailey disease. Br. J. Dermatol. 2010, 162, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Damiens, E.; Baratte, B.; Marie, D.; Eisenbrand, G.; Meijer, L. Anti-mitotic properties of indirubin-3′-monoxime, a CDK/GSK-3 inhibitor: Induction of endoreplication following prophase arrest. Oncogene 2001, 20, 3786–3797. [Google Scholar] [CrossRef] [PubMed]
- Novotny, L.; Reichelova, V.; Balazova, E.; Ujhazy, V. Comparison of some biochemical parameters of arabinosylcytosine and cyclocytidine in L1210 murine leukemia cells. Neoplasma 1990, 37, 13–22. [Google Scholar] [PubMed]
- Murota, K.; Shimizu, S.; Miyamoto, S.; Izumi, T.; Obata, A.; Kikuchi, M.; Terao, J. Unique uptake and transport of isoflavone aglycones by human intestinal caco-2 cells: Comparison of isoflavonoids and flavonoids. J. Nutr. 2002, 132, 1956–1961. [Google Scholar] [CrossRef] [PubMed]
- Saeki, T.; Umezawa, H.; Tokieda-Fujishige, T.; Hori, M. Letter: Biological effects of azomycin (2-nitro-imidazole): Inhibition of ribonucleotide reductase. J. Antibiot. 1974, 27, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Manca, S.; Magrelli, A.; Cialfi, S.; Lefort, K.; Ambra, R.; Alimandi, M.; Biolcati, G.; Uccelletti, D.; Palleschi, C.; Screpanti, I.; et al. Oxidative stress activation of miR-125b is part of the molecular switch for Hailey-Hailey disease manifestation. Exp. Dermatol. 2011, 20, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Biolcati, G.; Aurizi, C.; Barbieri, L.; Cialfi, S.; Screpanti, I.; Talora, C. Efficacy of the melanocortin analogue Nle4-d-Phe7-alpha-melanocyte-stimulating hormone in the treatment of patients with Hailey-Hailey disease. Clin. Exp. Dermatol. 2014, 39, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D. Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug Metab. Rev. 2006, 38, 769–789. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.I.; Griffiths, E.J.; Rutter, G.A. Regulation of ATP production by mitochondrial Ca2+. Cell Calcium 2012, 52, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missiaen, L.; Dode, L.; Vanoevelen, J.; Raeymaekers, L.; Wuytack, F. Calcium in the Golgi apparatus. Cell Calcium 2007, 41, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Missiaen, L.; Raeymaekers, L.; Dode, L.; Vanoevelen, J.; Van Baelen, K.; Parys, J.B.; Callewaert, G.; De Smedt, H.; Segaert, S.; Wuytack, F. SPCA1 pumps and Hailey-Hailey disease. Biochem. Biophys. Res. Commun. 2004, 322, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.E.; Miller, M.L.; Prasad, V. Secretory pathway stress responses as possible mechanisms of disease involving Golgi Ca2+ pump dysfunction. BioFactors 2011, 37, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Szigeti, R.; Kellermayer, R. The forgotten yeast model of Hailey-Hailey disease. Int. J. Dermatol. 2015, 55, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, C.; Fink, G.R. Guide to Yeast Genetics and Molecular and Cell Biology, Part B; Academic Press: Boston, MA, USA, 2002. [Google Scholar]
- Cialfi, S.; Palermo, R.; Manca, S.; De Blasio, C.; Vargas Romero, P.; Checquolo, S.; Bellavia, D.; Uccelletti, D.; Saliola, M.; D’Alessandro, A.; et al. Loss of Notch1-dependent p21(Waf1/Cip1) expression influences the Notch1 outcome in tumorigenesis. Cell Cycle 2014, 13, 2046–2055. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemicals | Oxidative Stress Selection | EC50 (mM) |
|---|---|---|
| S1973 (Cyclocytidine) | H2O2 | 51.26 ± 1.19 |
| S2267 (Azomycin) | Menadione | 102.64 ± 0.50 |
| S2314 (Kaempferol) | H2O2 | 50.5 ± 0.44 |
| S2328 (Nalidixic acid) | Menadione | 50.13 ± 4.84 |
| S2386 (Indirubin) | Menadione | 48.56 ± 3.01 |
| S2387 (Lappaconite) | Menadione | 9.35 ± 0.42 |
| SyBr Green Assays | Sequence 5′-3′ |
| GST-M1 Fw | AGAGGAGAAGATTCGTGTGG |
| GST-M1 Rev | TGTTTCCTGCAAACCATGGC |
| GAPDH Fw | TGCACCACCAACTGCTTAG |
| GAPDH Rev | GAGGCAGGGATGATGTTC |
| Taqman Gene Expression Assays | Assay Reference Number |
| NFE2L2 (NRF2) | Hs00975961_g1 |
| GAPDH | Hs99999905_m1 |
| NQO1 | Hs02512143_s1 |
| SOD1 | Hs00533490_m1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ficociello, G.; Zonfrilli, A.; Cialfi, S.; Talora, C.; Uccelletti, D. Yeast-Based Screen to Identify Natural Compounds with a Potential Therapeutic Effect in Hailey-Hailey Disease. Int. J. Mol. Sci. 2018, 19, 1814. https://doi.org/10.3390/ijms19061814
Ficociello G, Zonfrilli A, Cialfi S, Talora C, Uccelletti D. Yeast-Based Screen to Identify Natural Compounds with a Potential Therapeutic Effect in Hailey-Hailey Disease. International Journal of Molecular Sciences. 2018; 19(6):1814. https://doi.org/10.3390/ijms19061814
Chicago/Turabian StyleFicociello, Graziella, Azzurra Zonfrilli, Samantha Cialfi, Claudio Talora, and Daniela Uccelletti. 2018. "Yeast-Based Screen to Identify Natural Compounds with a Potential Therapeutic Effect in Hailey-Hailey Disease" International Journal of Molecular Sciences 19, no. 6: 1814. https://doi.org/10.3390/ijms19061814