Age-Related Changes in MicroRNA in the Rat Pituitary and Potential Role in GH Regulation

Abstract
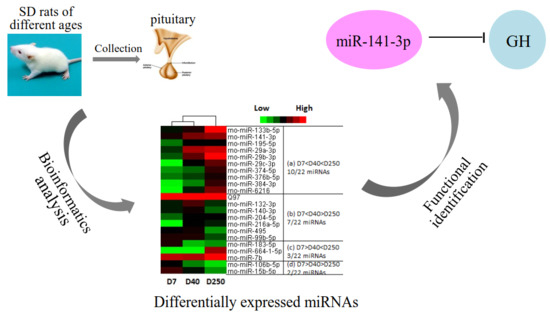
:
1. Introduction
2. Results
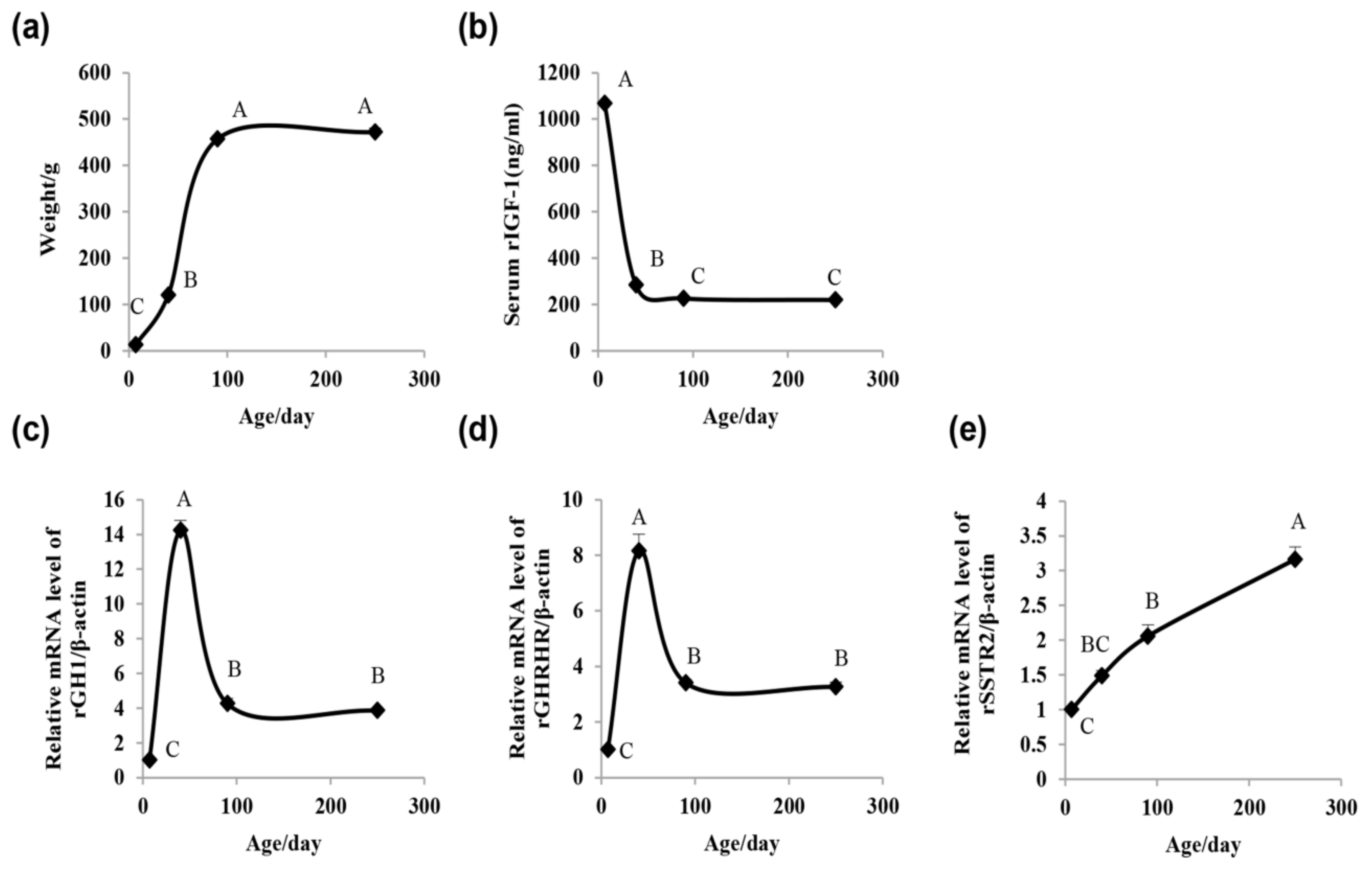
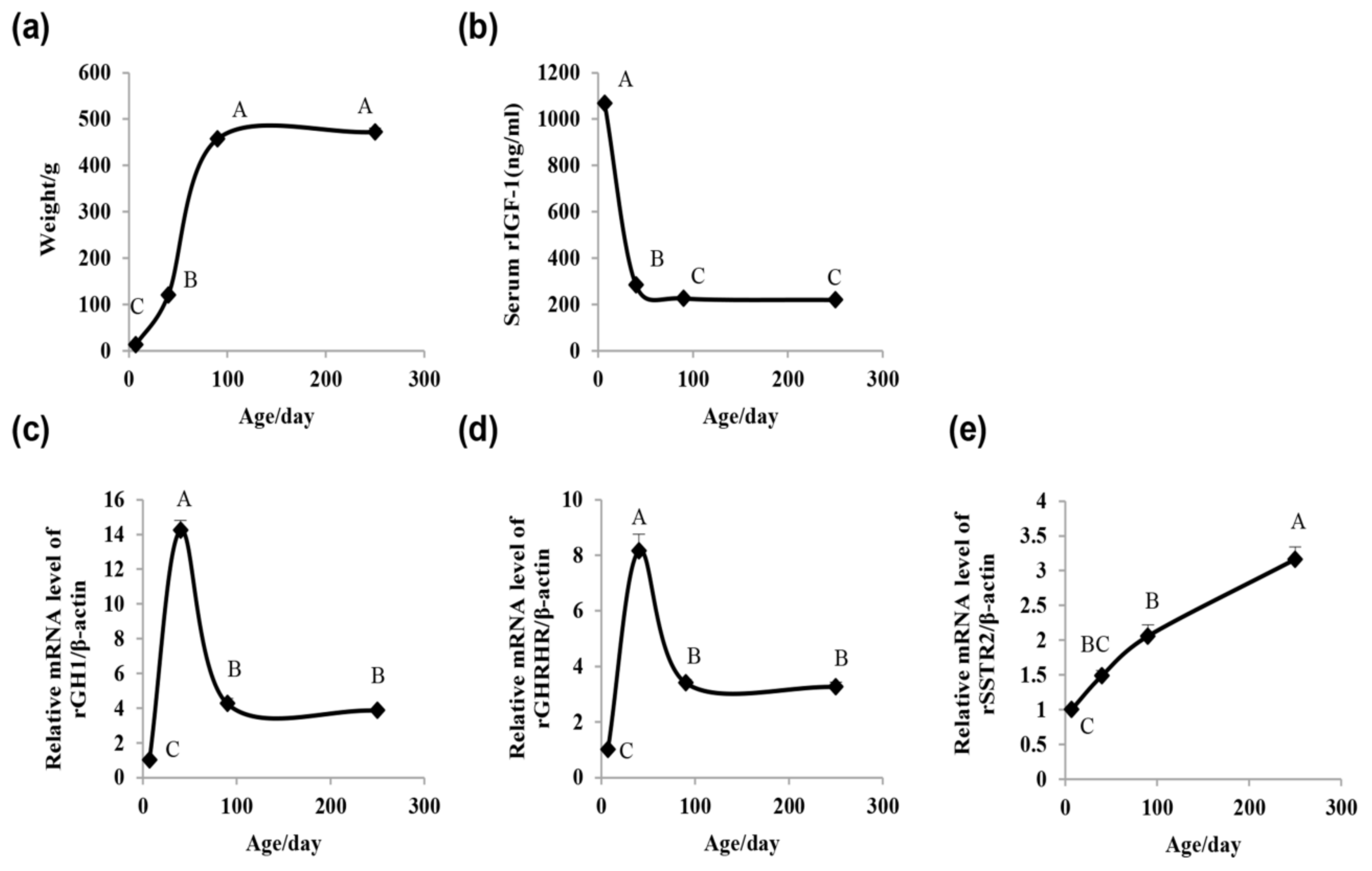
2.1. Rat Growth Curve, Serum IGF-1 Concentration, and Pituitary GH1, Growth Hormone-Releasing Hormone Receptor(GHRHR), and Somatostatin Receptor 2 (SSTR2) mRNA Expression Assay
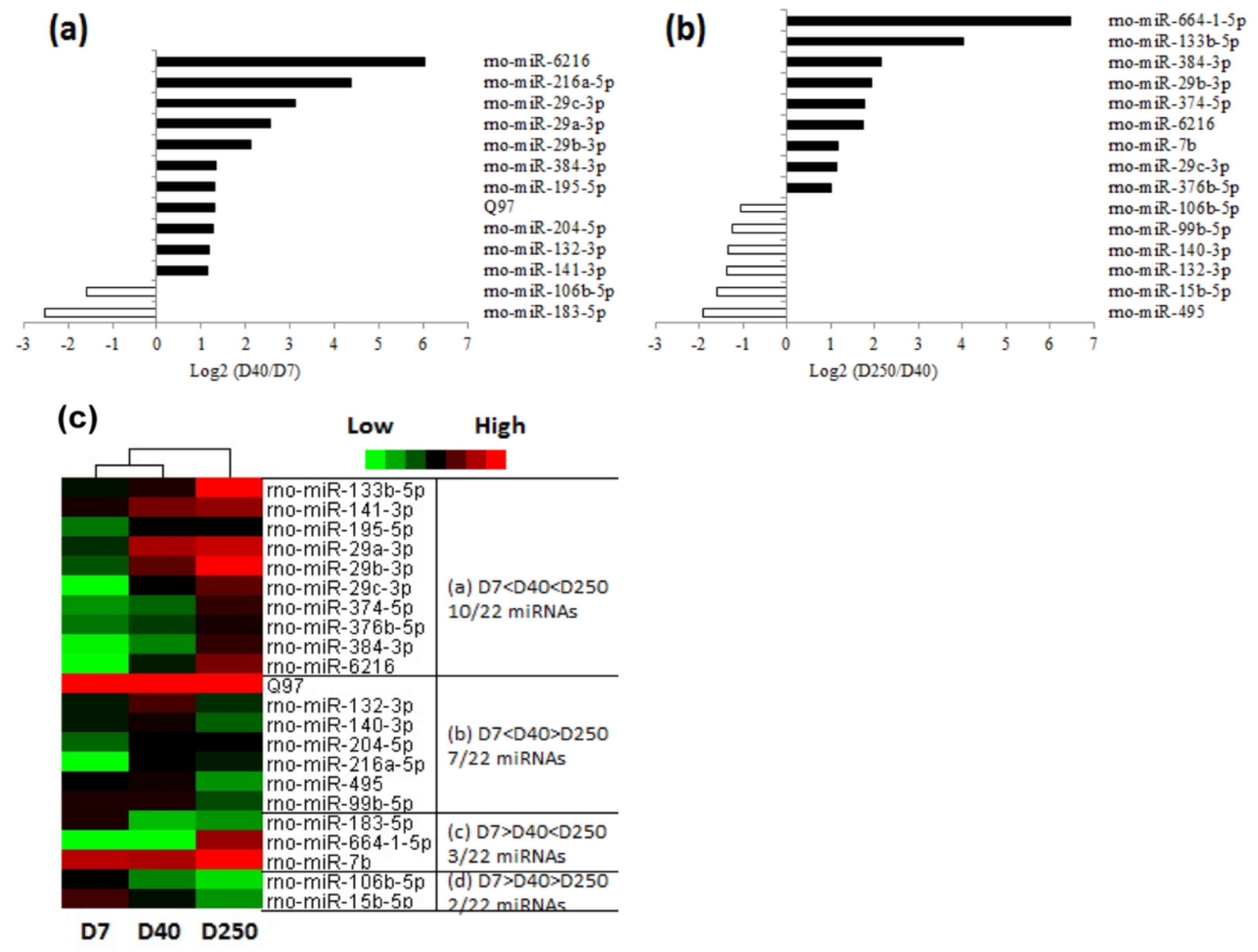
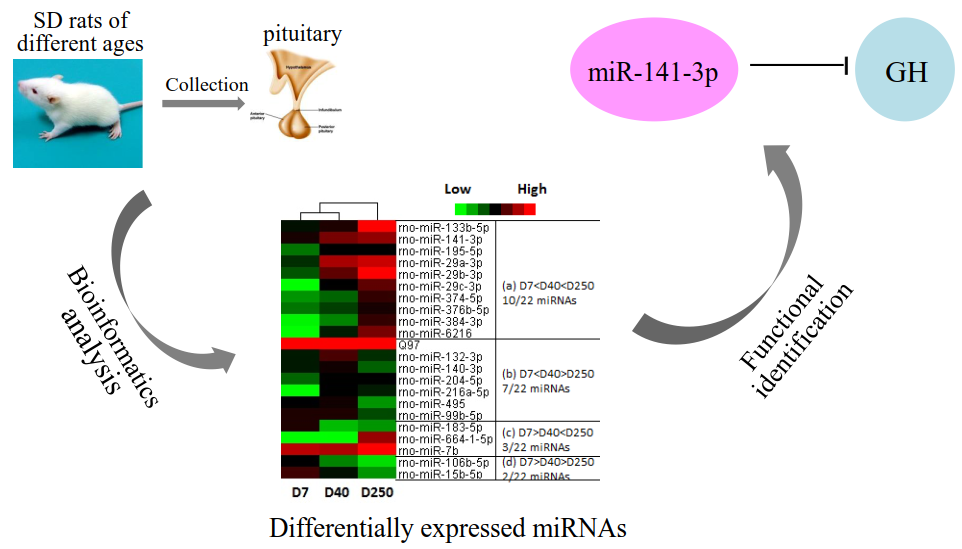
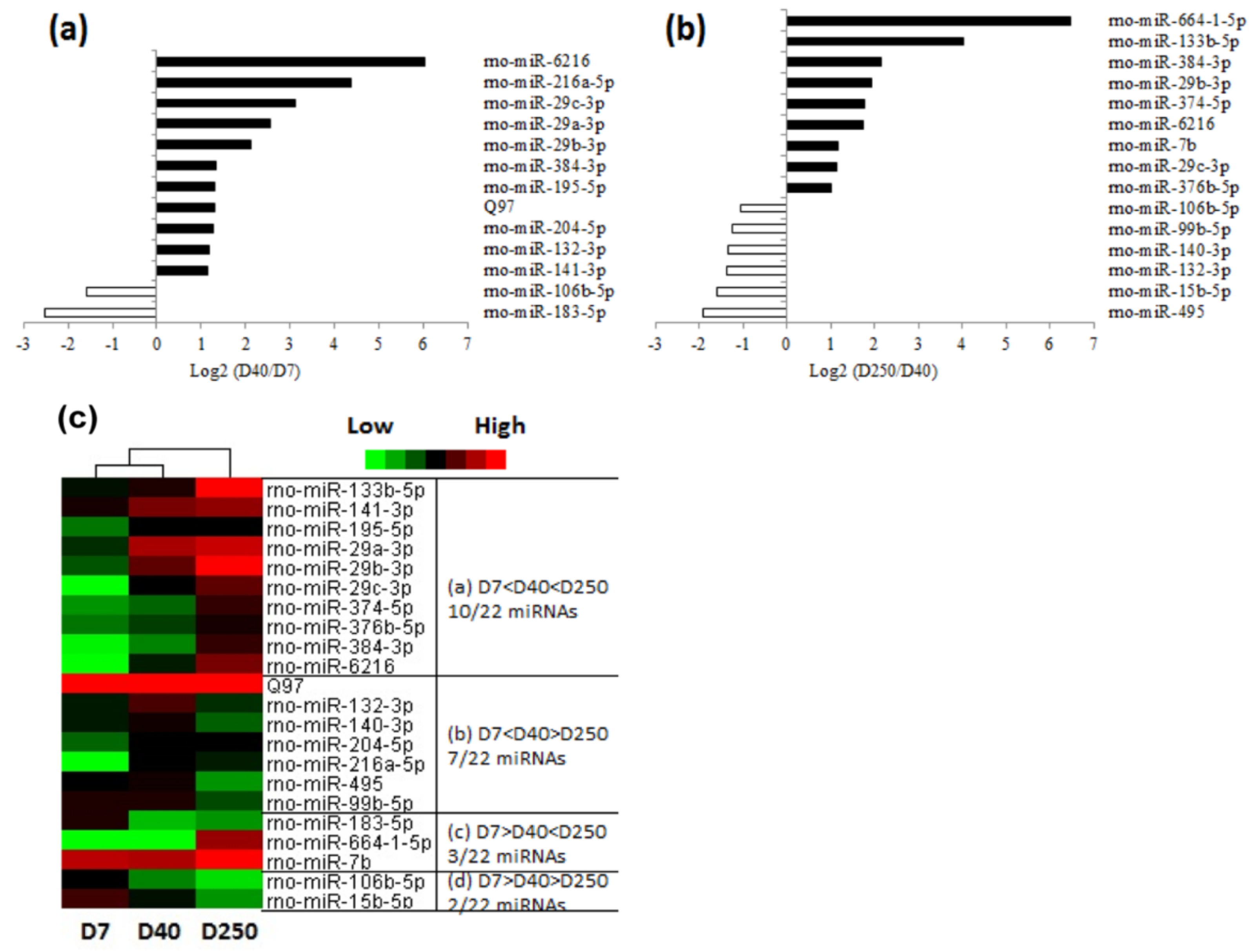
2.2. Dynamic Expression of miRNAs in the Pituitary
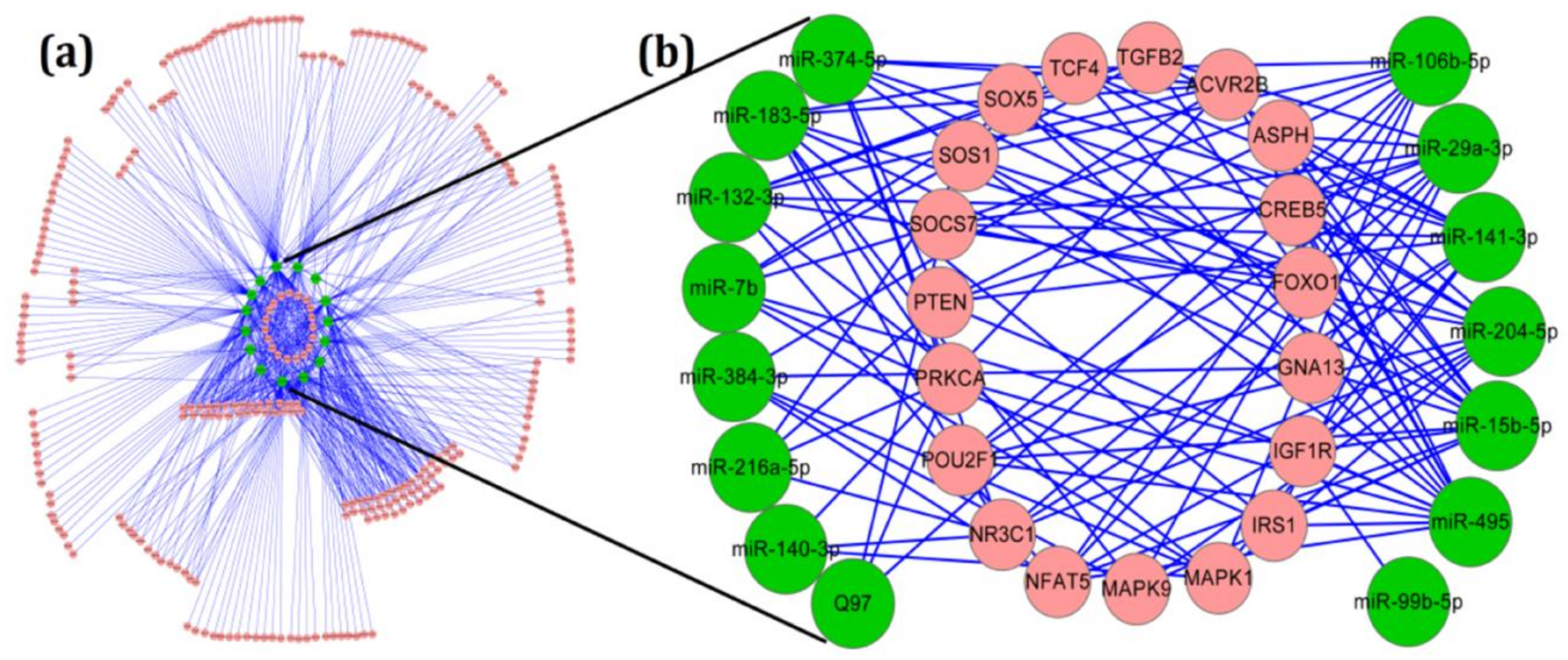
2.3. Target and Pathway Analysis of Differentially Expressed miRNAs
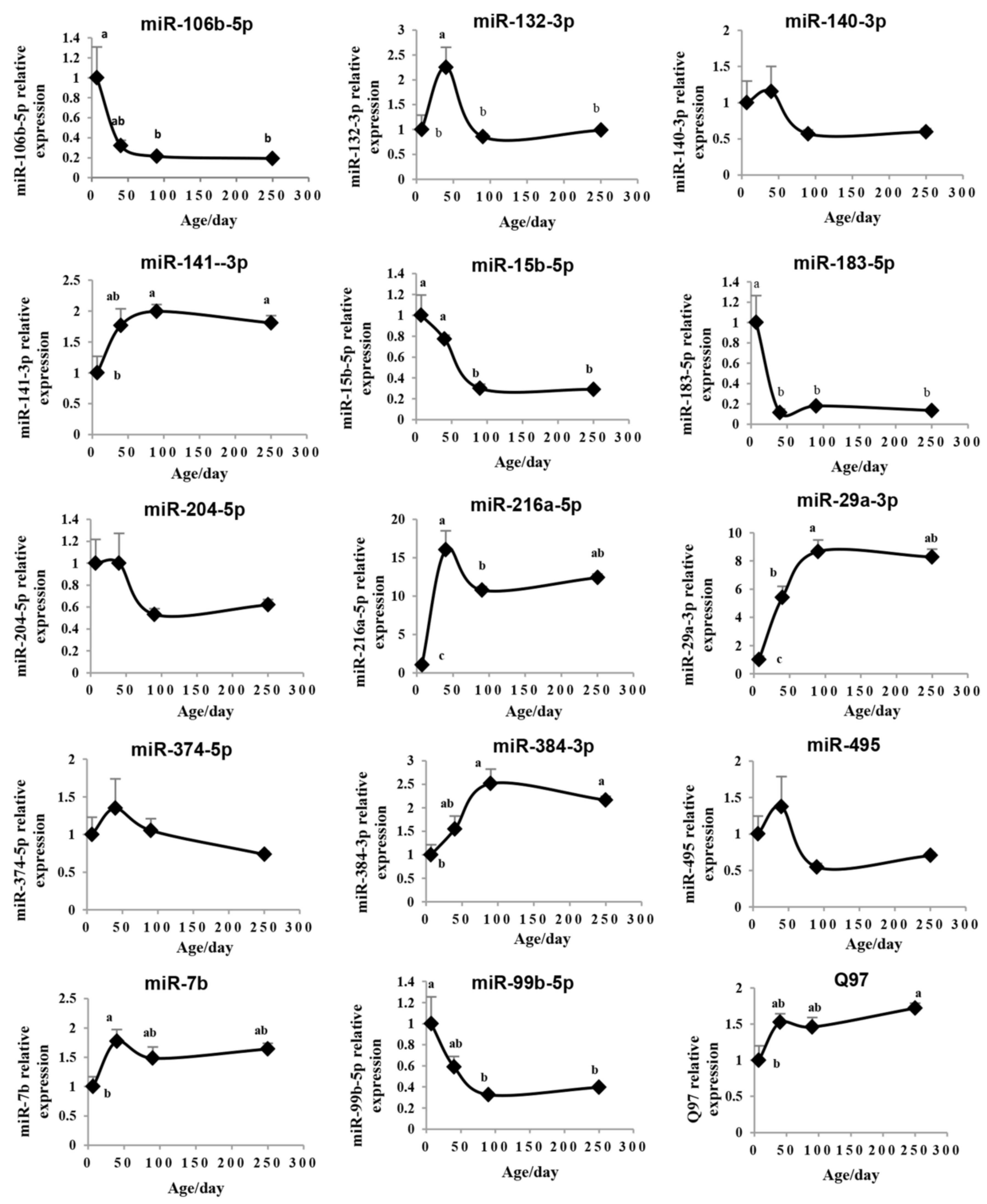
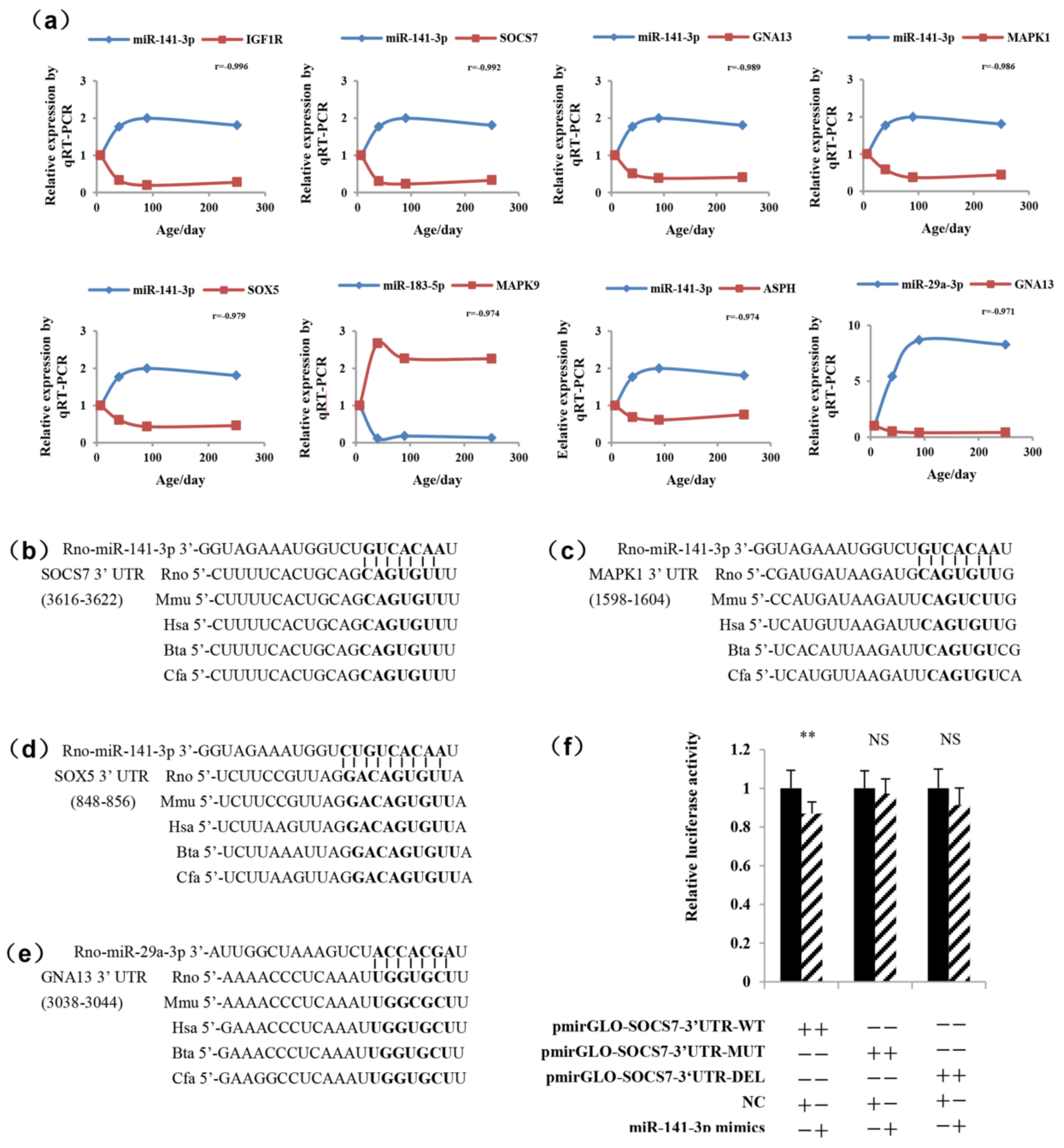
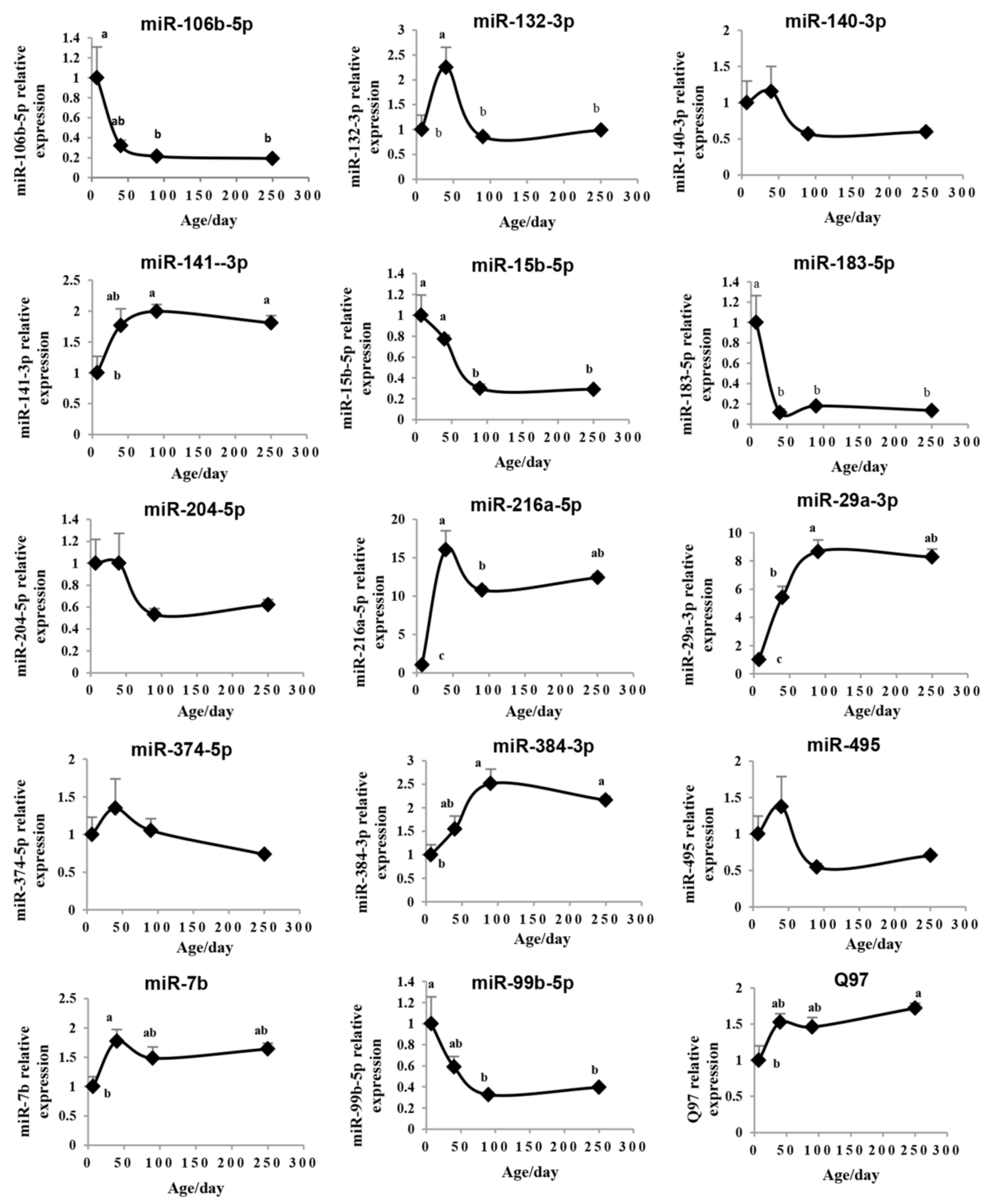
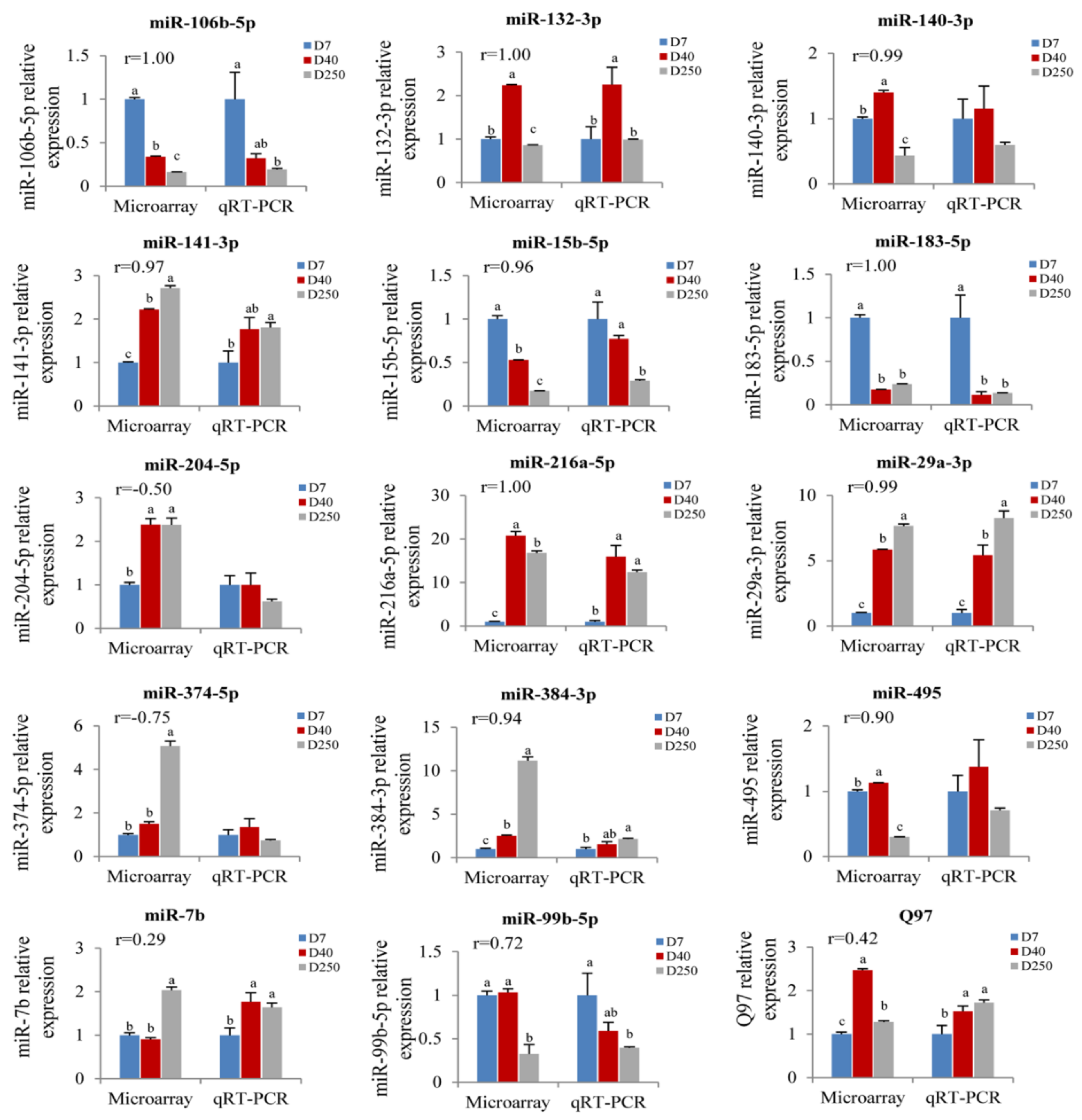
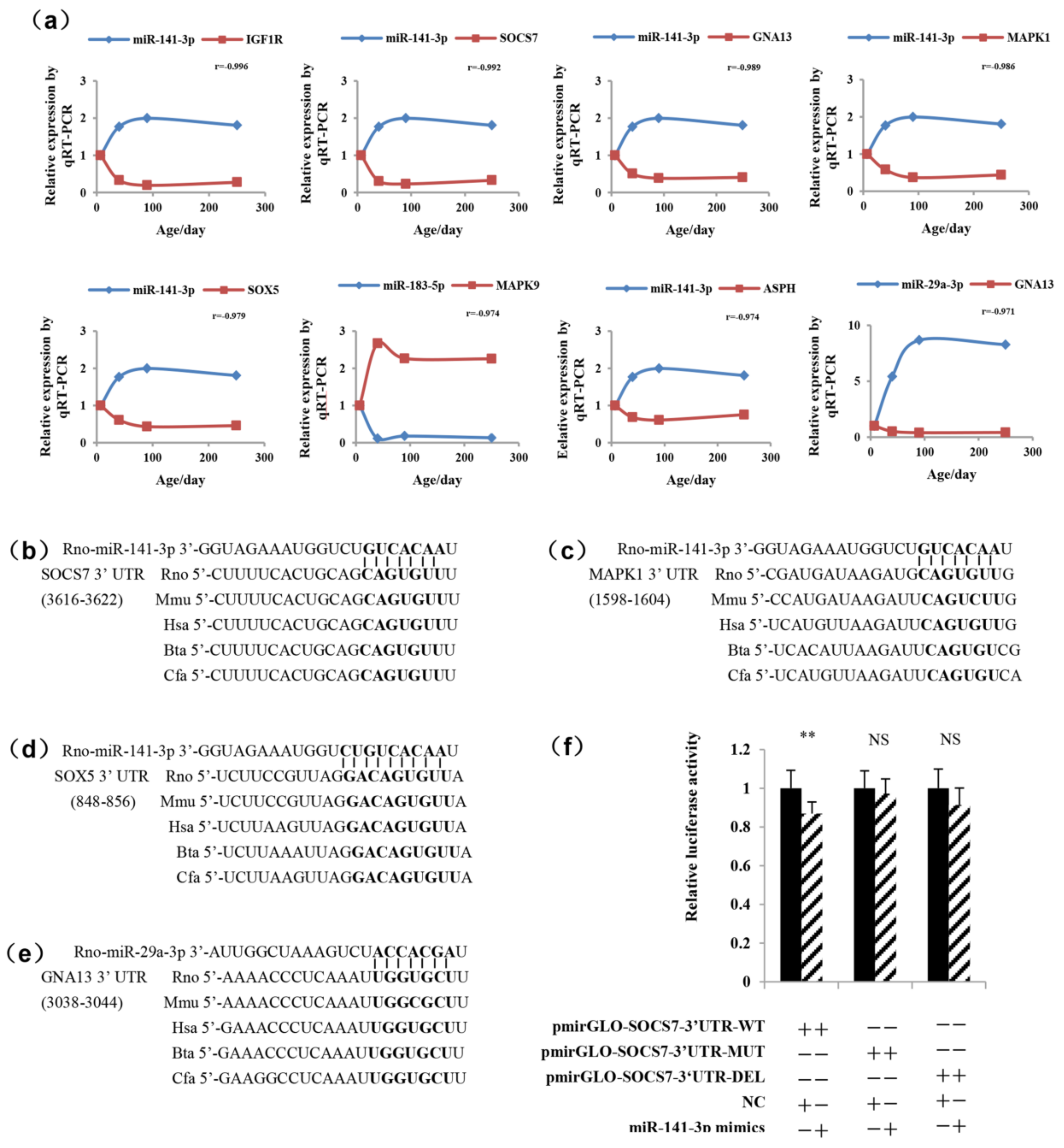
2.4. Validation of the Pituitary miRNAs via qRT-PCR
2.5. Determination of the Functional miRNA Target Pairs
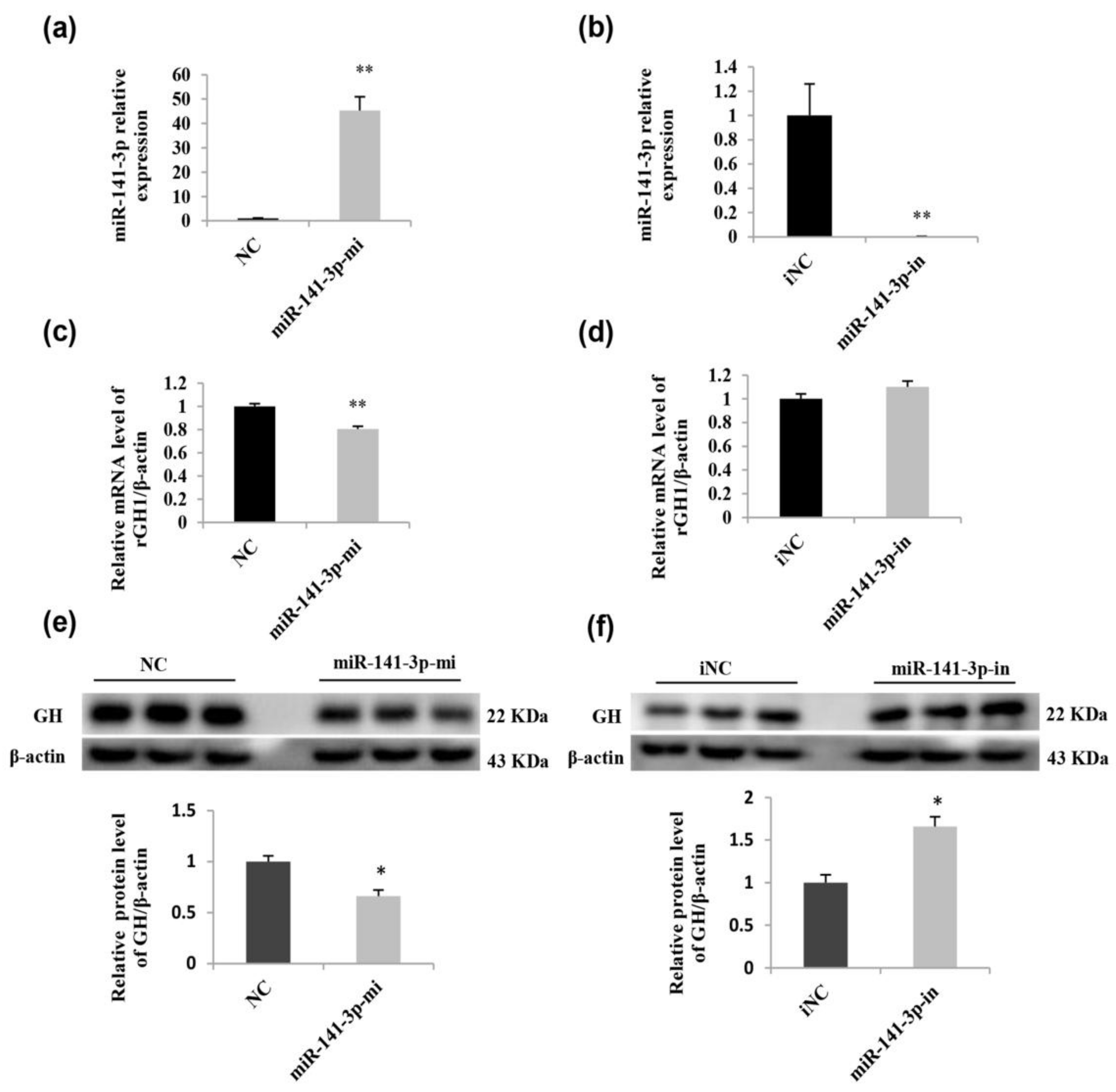
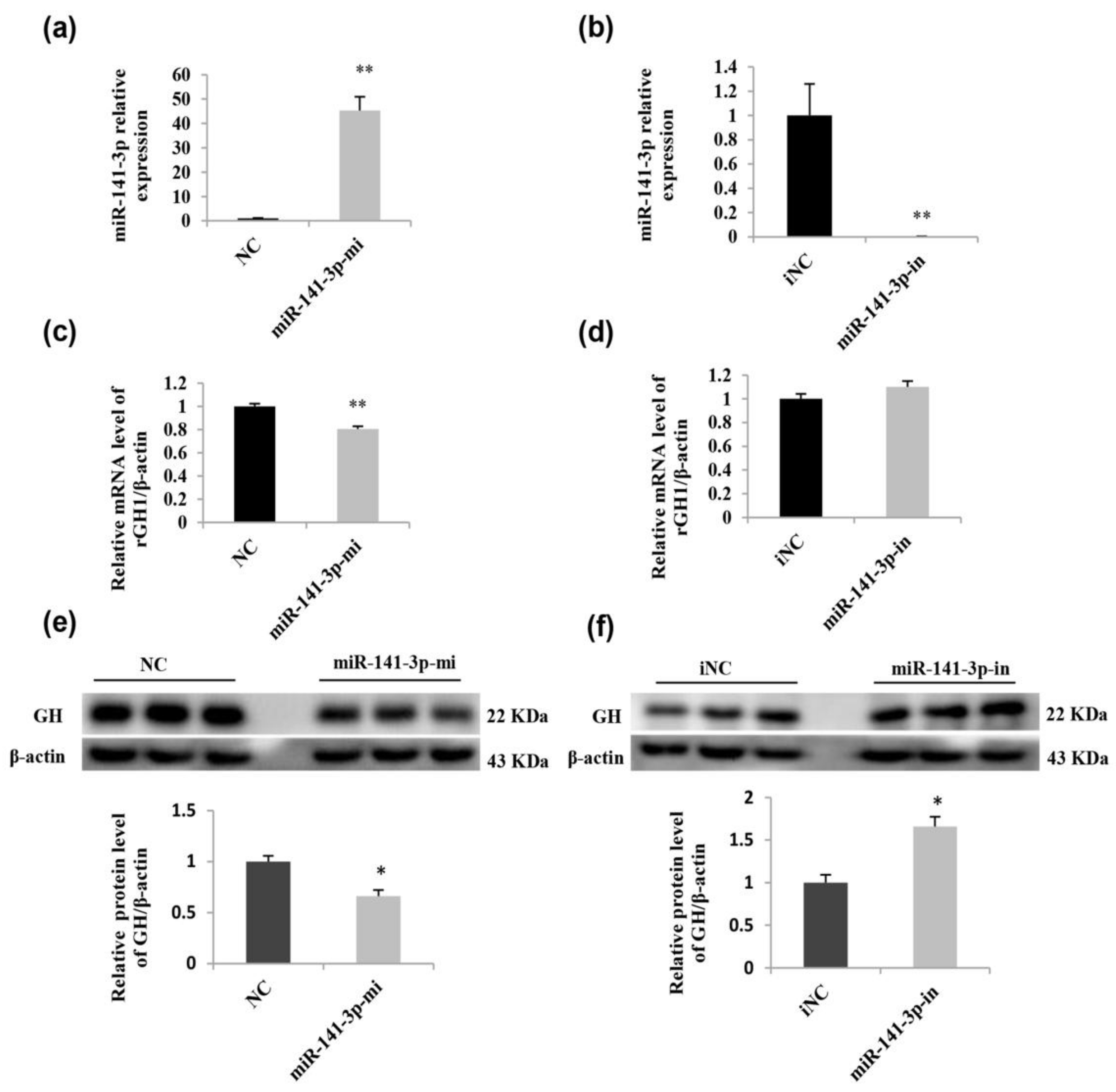
2.6. miR-141-3p Negatively Regulates Rat Pituitary GH Expression
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Preparation
4.2. Serum IGF-1 Concentration Assay
4.3. RNA Extraction and Quantitative Real-Time PCR Analysis
4.4. MicroRNA Microarray Assay
4.5. Tissue-Specific miRNA Target Prediction and Exploration of the Involved Pathways
4.6. Cell Lines and Cell Transfection
4.7. Western Blot Analysis
4.8. Luciferase Reporter Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GH | Growth hormone |
| miRNA | MicroRNA |
| mRNA | Messenger RNA |
| RT-PCR | Reverse transcriptase quantitative polymerase chain reaction |
| 3′UTR | 3′ Untranslated region |
| GHRHR | Growth hormone-releasing hormone receptor |
| IPA | Ingenuity pathway analysis |
| SSTR2 | Somatostatin receptor 2 |
References
- Yeung, C.M.; Chan, C.B.; Leung, P.S.; Cheng, C.H. Cells of the anterior pituitary. Int. J. Biochem. Cell Boil. 2006, 38, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Gleiberman, A.S.; Rosenfeld, M.G. Molecular physiology of pituitary development: Signaling and transcriptional networks. Physiol. Rev. 2007, 87, 933–963. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, O.G.; Lindahl, A.; Nilsson, A.; Isgaard, J. Mechanism of the stimulatory effect of growth hormone on longitudinal bone growth. Endocr. Rev. 1987, 8, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Salomon, F.; Cuneo, R.C.; Hesp, R.; Sönksen, P.H. The effects of treatment with recombinant human growth hormone on body composition and metabolism in adults with growth hormone deficiency. N. Engl. J. Med. 1989, 321, 1797–1803. [Google Scholar] [CrossRef] [PubMed]
- Binnerts, A.; Swart, G.R.; Wilson, J.H.; Hoogerbrugge, N.; Pols, H.A.; Birkenhager, J.C.; Lamberts, S.W. The effect of growth hormone administration in growth hormone deficient adults on bone, protein, carbohydrate and lipid homeostasis, as well as on body composition. Clin. Endocrinol. 1992, 37, 79–87. [Google Scholar] [CrossRef]
- Robert, C.T.; Brown, A.L.; Graham, D.E.; Seelig, S.; Berry, S.; Gabbay, K.H.; Rechler, M.M. Growth hormone regulates the abundance of insulin-like growth factor 1 RNA in adult rat liver. J. Biol. Chem. 1986, 261, 10025–10028. [Google Scholar]
- Bichell, D.P.; Kikuchi, K.; Rotwein, P. Growth hormone rapidly activates insulin-like growth factor 1 gene transcription in vivo. Mol. Endocrinol. 1992, 6, 1899–1908. [Google Scholar] [PubMed]
- Domené, H.; Krishnamurthi, K.; Eshet, R.; Gilad, I.; Laron, Z.; Koch, I.; Stannard, B.; Cassorla, F.; Roberts, C.T., Jr. Growth hormone (GH) stimulates insulin-like growth factor-1 (IGF-1) and IGF-1-binding protein-3, but not GH receptor gene expression in livers of juvenile rats. Endocrinology 1993, 133, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Houston, B.; O’Neill, I.E. Insulin and growth hormone act synergistically to stimulate insulin-like growth factor-1 production by cultured chicken hepatocytes. J. Endocrinol. 1991, 128, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Steger, R.W.; Bartke, A.; Cecim, M. Premature ageing in transgenic mice expressing different growth hormone genes. J. Reprod. Fertil. 1993, 46, 61–75. [Google Scholar]
- Sonntag, W.E.; Lynch, C.; Thornton, P.; Khan, A.; Bennett, S.; Ingram, R. The effects of growth hormone and IGF-1 deficiency on cerebrovascular and brain ageing. J. Anat. 2000, 197, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Flurkey, K.; Papaconstantinou, J.; Miller, R.A.; Harrison, D.E. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proc. Natl. Acad. Sci. USA 2001, 98, 6736–6741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, C.S.; Ramsey, M.M.; Sonntag, W.E. A critical analysis of the role of growth hormone and IGF-1 in aging and lifespan. Trends Genet. 2002, 18, 295–301. [Google Scholar] [CrossRef]
- Corpas, E.; Harman, S.M.; Blackman, M.R. Human growth hormone and human aging. Endocr. Rev. 1993, 14, 20–39. [Google Scholar] [CrossRef] [PubMed]
- Lamberts, S.W.; Aw, V.D.B.; Aj, V.D.L. The endocrinology of aging. Science 1997, 278, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceda, G.P.; Dall’Aglio, E.; Maggio, M.; Lauretani, F.; Bandinelli, S.; Falzoi, C.; Grimaldi, W.; Ceresini, G.; Corradi, F.; Ferrucci, L. Clinical implications of the reduced activity of the GH-IGF-1 axis in older men. J. Endocrinol. Investig. 2005, 28, 96–100. [Google Scholar]
- Ghigo, E.; Arvat, E.; Gianotti, L.; Ramunni, J.; Divito, L.; Maccagno, B.; Grottoli, S.; Camanni, F. Human aging and the GH-IGF-1 axis. J. Pediatr. Endocrinol. Metab. 1996, 9, 271–278. [Google Scholar] [PubMed]
- Troen, B.R. The biology of aging. Mt. Sinai J. Med. 2003, 70, 3–22. [Google Scholar] [PubMed]
- Chen, L.H.; Chiou, G.Y.; Chen, Y.W.; Li, H.Y.; Chiou, S.H. MicroRNA and aging: A novel modulator in regulating the aging network. Ageing Res. Rev. 2010, 9, S59–S66. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bushati, N.; Cohen, S.M. MicroRNA functions. Annu. Rev. Cell Dev. Boil. 2007, 23, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Florez, S.; Gutierrezhartmann, A.; Martin, J.F.; Amendt, B.A. MicroRNAs regulate pituitary development, and microRNA 26b specifically targets lymphoid enhancer factor 1 (Lef-1), which modulates pituitary transcription factor 1 (Pit-1) expression. J. Biol. Chem. 2010, 285, 34718–34728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Lin, J.K.; Chen, J.; Liu, X.F.; Liu, J.L.; Luo, H.S.; Li, Y.Q.; Cui, S. MicroRNA 375 mediates the signaling pathway of corticotropin-releasing factor (CRF) regulating pro-opiomelanocortin (POMC) expression by targeting mitogen-activated protein kinase 8. J. Biol. Chem. 2013, 288, 10361–10373. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.J.; Li, N.; Liang, R.; Sarojini, H.; Jin, A.; Masternak, M.M.; Bartke, A.; Wang, E. MicroRNA regulation in ames dwarf mouse liver may contribute to delayed aging. Aging Cell 2010, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Azhar, G.; Wei, J.Y. The expression of microRNA and microRNA clusters in the aging heart. PLoS ONE 2012, 7, e34688. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Boopathi, E.; Addya, S.; Phillips, B.; Rigoutsos, I.; Penn, R.B.; Rattan, S. Aging-associated changes in microRNA expression profile of internal anal sphincter smooth muscle: Role of microRNA-133a. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G964–G973. [Google Scholar] [CrossRef] [PubMed]
- Danka Mohammed, C.P.; Park, J.S.; Nam, H.G.; Kim, K. Micrornas in brain aging. Mech. Ageing Dev. 2017, 168, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cai, Z.; Wei, S.; Zhou, H.; Zhou, H.; Jiang, X.; Xu, N. MicroRNA expression profiling of the porcine developing hypothalamus and pituitary tissue. Int. J. Mol. Sci. 2013, 14, 20326–20339. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kohane, I.S. Correction: Tissue and process specific microRNA–mRNA co-expression in mammalian development and malignancy. PLoS ONE 2009, 4, e5436. [Google Scholar] [CrossRef]
- Li, H.; Xi, Q.; Xiong, Y.; Xiao, C.; Qi, Q.; Lin, Y.; Gang, S.; Wang, S.; Wang, L.; Ping, G. A comprehensive expression profile of microRNAs in porcine pituitary. PLoS ONE 2011, 6, e24883. [Google Scholar] [CrossRef] [PubMed]
- Sivapragasam, M.; Rotondo, F.; Lloyd, R.V.; Scheithauer, B.W.; Cusimano, M.; Syro, L.V.; Kovacs, K. MicroRNAs in the human pituitary. Endocr. Pathol. 2011, 22, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Han, D.X.; Dai, L.S.; Gao, Y.; Ding, Y.; Yu, X.F.; Chen, J.; Jiang, H.; Chen, C.Z.; Zhang, J.B. A comprehensive expression profile of microRNAs in rat’s pituitary. Int. J. Clin. Exp. Med. 2015, 8, 13289–13295. [Google Scholar] [PubMed]
- Rieutort, M. Pituitary content and plasma levels of growth hormone in foetal and weanling rats. J. Endocrinol. 1974, 60, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, W.E.; Steger, R.W.; Forman, L.J.; Meites, J. Decreased pulsatile release of growth hormone in old male rats. Endocrinology 1980, 107, 1875–1879. [Google Scholar] [CrossRef] [PubMed]
- Iruthayanathan, M.; Zhou, Y.H.; Childs, G.V. Dehydroepiandrosterone restoration of growth hormone gene expression in aging female rats, in vivo and in vitro: Evidence for actions via estrogen receptors. Endocrinology 2005, 146, 5176–5187. [Google Scholar] [CrossRef] [PubMed]
- Birge, C.A.; Peake, G.T.; Mariz, I.K.; Daughaday, W.H. Radioimmunoassayable growth hormone in the rat pituitary gland: Effects of age, sex and hormonal state. Endocrinology 1967, 81, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.O.; Eden, S.; Isaksson, O. Sexual dimorphism in the control of growth hormone secretion. Endocr. Rev. 1985, 6, 128–150. [Google Scholar] [CrossRef] [PubMed]
- Silvia, G.P.; Chowen, J.A.; García, S.L.M.; Jesús, A. Ontogeny of pituitary transcription factor-1 (PIT-1), growth hormone (GH) and prolactin (PRL) mRNA levels in male and female rats and the differential expression of PIT-1 in lactotrophs and somatotrophs. J. Neuroendocrinol. 2010, 8, 211–225. [Google Scholar]
- Reed, D.K.; Korytko, A.I.; Hipkin, R.W.; Wehrenberg, W.B.; Schonbrunn, A.; Cuttler, L. Pituitary somatostatin receptor (sst)1-5 expression during rat development: Age-dependent expression of sst2. Endocrinology 1999, 140, 4739–4744. [Google Scholar] [CrossRef] [PubMed]
- Korytko, A.I.; Zeitler, P.; Cuttler, L. Developmental regulation of pituitary growth hormone-releasing hormone receptor gene expression in the rat. Endocrinology 1996, 137, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Boehm, M.; Slack, F. A developmental timing microRNA and its target regulate life span in C. elegans. Science 2005, 310, 1954–1957. [Google Scholar] [CrossRef] [PubMed]
- Maes, O.C.; An, J.; Sarojini, H.; Wang, E. Murine microRNAs implicated in liver functions and aging process. Mech. Ageing Dev. 2008, 129, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Miska, E.A.; Alvarezsaavedra, E.; Townsend, M.; Yoshii, A.; Rakic, P.; Constantinepaton, M.; Horvitz, H.R. Microarray analysis of microRNA expression in the developing mammalian brain. Genome Biol. 2004, 5, R68. [Google Scholar] [CrossRef] [PubMed]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Sun, Y.; Wu, J.; Yan, S.; Deng, Z.; Wang, J.; Liao, S.; Yin, D.; Li, G. Discovering novel microRNAs and age-related nonlinear changes in rat brains using deep sequencing. Neurobiol. Aging 2015, 36, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Capri, M.; Olivieri, F.; Lanzarini, C.; Remondini, D.; Borelli, V.; Lazzarini, R.; Graciotti, L.; Albertini, M.C.; Bellavista, E.; Santoro, A. Identification of miR-31-5p, miR-141-3p, miR-200c-3p, and GLT1 as human liver aging markers sensitive to donor–recipient age-mismatch in transplants. Aging Cell 2017, 16, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Rajewsky, N. MicroRNA target predictions in animals. Nat. Genet. 2006, 38, S8. [Google Scholar] [CrossRef] [PubMed]
- Tombol, Z.; Szabo, P.M.; Molnar, V.; Wiener, Z.; Tolgyesi, G.; Horanyi, J.; Riesz, P.; Reismann, P.; Patocs, A.; Liko, I.; et al. Integrative molecular bioinformatics study of human adrenocortical tumors: MicroRNA, tissue-specific target prediction, and pathway analysis. Endocr. Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Niu, P.D.; Perezsanchez, J.; Le, B.P. Development of a protein binding assay for teleost insulin-like growth factor (IGF)-like: Relationships between growth hormone (GH) and IGF-like in the blood of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 1993, 11, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.L.; Liang, A.X.; Mao, D.G.; Han, L.; Liu, X.B.; Zhang, J.; Yang, L.G. Effect of genetic adjuvants on immune respondance, growth and hormone levels in somatostatin DNA vaccination-induced hu lambs. Vaccine 2010, 28, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Marsh, E.E.; Lin, Z.; Yin, P.; Milad, M.; Chakravarti, D.; Bulun, S.E. Differential expression of microRNA species in human uterine leiomyoma versus normal myometrium. Fertil. Steril. 2008, 89, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, F.; Xu, J.; Ye, F.; Shen, Y.; Zhou, J.; Lu, W.; Wan, X.; Ma, D.; Xie, X. Progressive miRNA expression profiles in cervical carcinogenesis and identification of HPV-related target genes for miR-29. J. Pathol. 2011, 224, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Botstein D: Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed]
- Vergoulis, T.; Vlachos, I.S.; Alexiou, P.; Georgakilas, G.; Maragkakis, M.; Reczko, M.; Gerangelos, S.; Koziris, N.; Dalamagas, T.; Hatzigeorgiou, A.G. Tarbase 6.0: Capturing the exponential growth of miRNA targets with experimental support. Nucleic Acids Res. 2012, 40, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Ambs, S.; Prueitt, R.L.; Yi, M.; Hudson, R.S.; Howe, T.M.; Petrocca, F.; Wallace, T.A.; Liu, C.G.; Volinia, S.; Calin, G.A. Genomic profiling of microRNA and messenger RNA reveals deregulated microRNA expression in prostate cancer. Cancer Res. 2008, 68, 6162–6170. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Baek, M.; Gusev, Y.; Brackett, D.J.; Nuovo, G.J.; Schmittgen, T.D. Systematic evaluation of microRNA processing patterns in tissues, cell lines, and tumors. RNA 2008, 14, 35–42. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Count of miRNAs |
|---|---|
| Calcium Signaling | 15 |
| Cyclic adenosine monophosphate (cAMP)-mediated Signaling | 15 |
| Externally regulated kinases/ mitogen-activated protein kinase (ERK/MAPK) Signaling | 15 |
| Glucocorticoid Receptor Signaling | 15 |
| Gonadotropin-releasing hormone (GNRH) Signaling | 15 |
| Growth Hormone Signaling | 15 |
| Insulin-like growth factor 1 (IGF-1) Signaling | 15 |
| Insulin Receptor Signaling | 15 |
| Protein Kinase A Signaling | 15 |
| Retinoic acid receptor (RAR) Activation | 15 |
| Wnt/β-catenin Signaling | 15 |
| Corticotropin Releasing Hormone Signaling | 14 |
| cAMP-response element binding protein (CREB) Signaling in Neurons | 14 |
| G-Protein Coupled Receptor Signaling | 14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Qi, Q.; Chen, T.; Luo, J.; Xi, Q.; Jiang, Q.; Sun, J.; Zhang, Y. Age-Related Changes in MicroRNA in the Rat Pituitary and Potential Role in GH Regulation. Int. J. Mol. Sci. 2018, 19, 2058. https://doi.org/10.3390/ijms19072058
Zhang H, Qi Q, Chen T, Luo J, Xi Q, Jiang Q, Sun J, Zhang Y. Age-Related Changes in MicroRNA in the Rat Pituitary and Potential Role in GH Regulation. International Journal of Molecular Sciences. 2018; 19(7):2058. https://doi.org/10.3390/ijms19072058
Chicago/Turabian StyleZhang, Haojie, Qien Qi, Ting Chen, Junyi Luo, Qianyun Xi, Qingyan Jiang, Jiajie Sun, and Yongliang Zhang. 2018. "Age-Related Changes in MicroRNA in the Rat Pituitary and Potential Role in GH Regulation" International Journal of Molecular Sciences 19, no. 7: 2058. https://doi.org/10.3390/ijms19072058