Identification and In Silico Prediction of Anticoagulant Peptides from the Enzymatic Hydrolysates of Mytilus edulis Proteins

Abstract
:1. Introduction
2. Results and Discussion
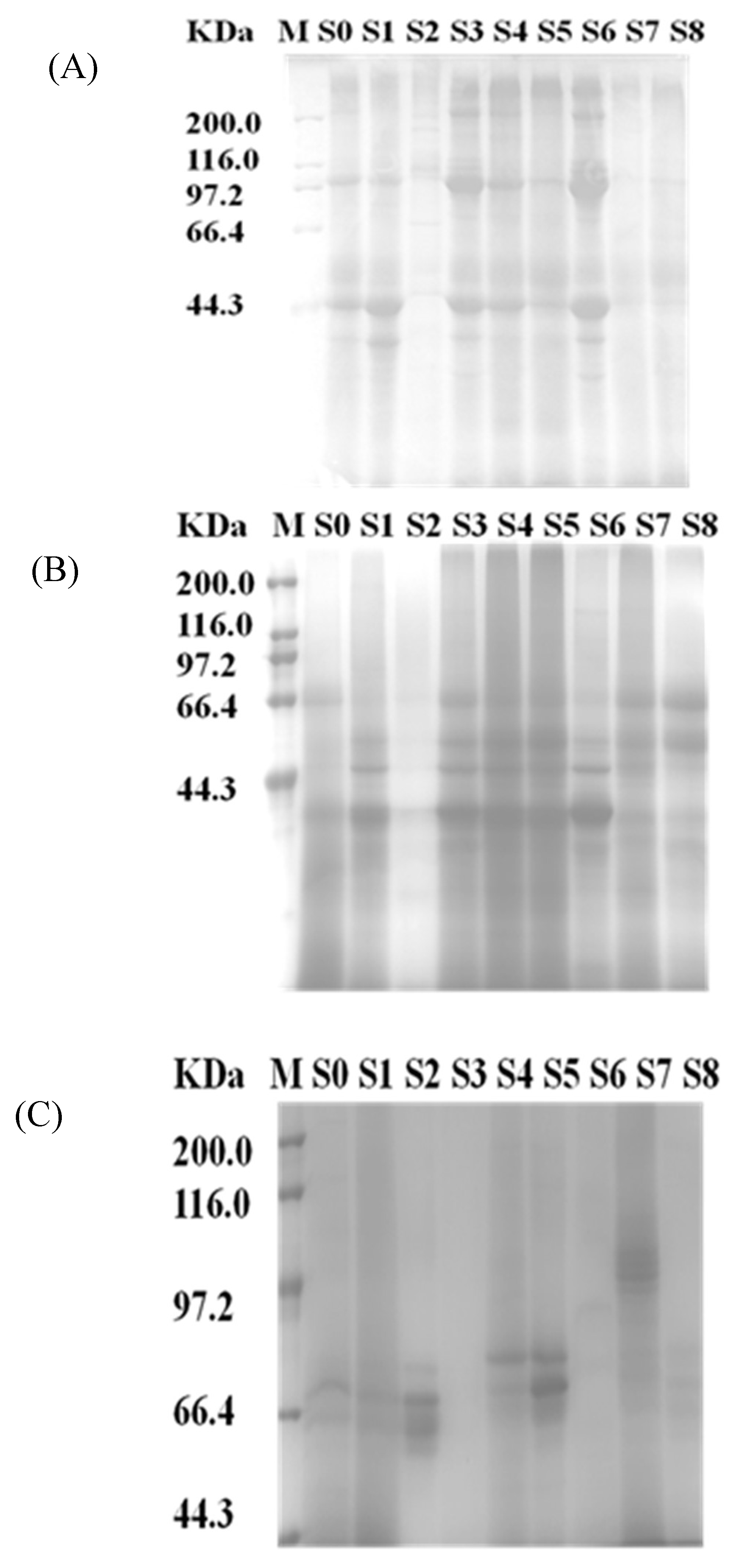
2.1. Distribution of Proteins from Different Tissues
2.2. Identification of the Peptides in Hydrolysates
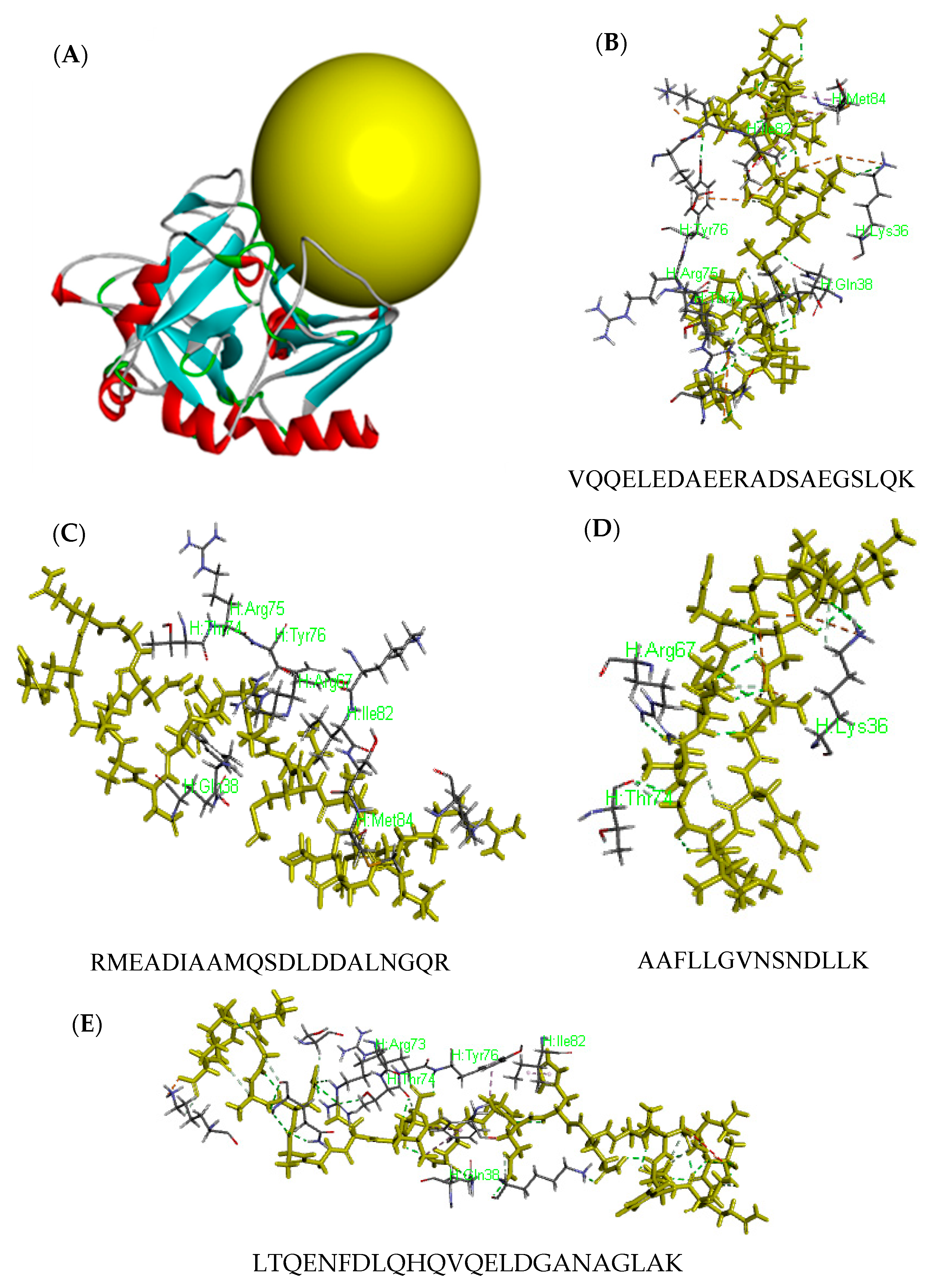
2.3. Activity Prediction and Molecular Docking of Peptides
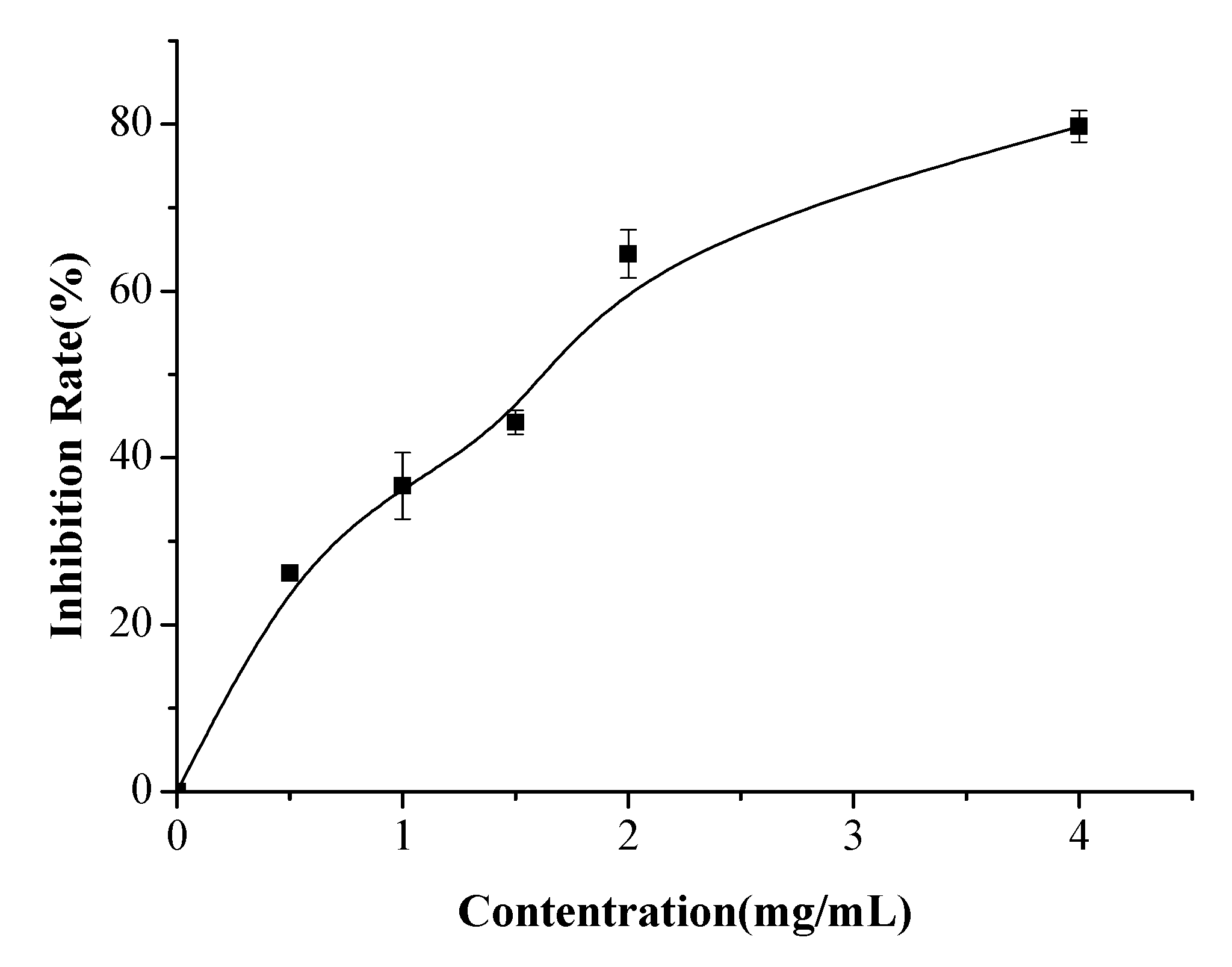
2.4. Anticoagulant Activity Determination
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Split of Mytilus edulis Organisms
3.3. SDS-PAGE
3.4. Enzymatic Hydrolysis by Trypsin
3.5. Peptide Identification by UPLC-Q-TOF-MS/MS
3.6. Activity Prediction by Peptide Ranker
3.7. Molecular Docking
3.8. Determination of Antithrombotic Peptides
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Lafarga, T.; Aluko, R.E.; Rai, D.K.; Oconnor, P.; Hayes, M. Identification of bioactive peptides from a papain hydrolysate of bovine serum albumin and assessment of an antihypertensive effect in spontaneously hypertensive rats. Food Res. Int. 2016, 81, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Je, J.Y.; Park, P.J.; Byun, H.G.; Jung, W.K.; Kim, S.K. Angiotensin I converting enzyme (ACE) inhibitory peptide derived from the sauce of fermented blue mussel, Mytilus edulis. Bioresour. Technol. 2005, 96, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zheng, W.W.; Li, J.; Wang, L.C.; Wu, H.; Wang, X.Z.; Shi, L. Rapid identification of bioactive peptides with antioxidant activity from the enzymatic hydrolysate of Mactraveneriformis by UHPLC–Q-TOF mass spectrometry. Food Chem. 2015, 167, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Switzar, L.; Giera, M.; Niessen, W.M.A. Protein digestion: An overview of the available techniques and recent developments. J. Proteome Res. 2013, 12, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ahn, C.B.; Je, J.Y. Anti-inflammatory action of high molecular weight Mytilus edulis hydrolysates fraction in LPS-induced RAW264.7 macrophage via NF-κB and MAPK pathways. Food Chem. 2016, 202, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Hernroth, B.; Baden, S.; Tassidis, H.; Hornaeus, K.; Guillemant, J.; Bergstrom, L.; Bergquist, J. Impact of ocean acidification on antimicrobial activity in gills of the blue mussel (Mytilus edulis). Fish Shellfish Immunol. 2016, 55, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Gosset, C.C.; Nascimento, J.D.; Augé, M.; Bierbe, N. Evidence for adaptation from standing genetic variation on an antimicrobial peptide gene in the mussel Mytilus edulis. Mol. Ecol. 2014, 23, 3000–3012. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.; Xu, Y.F. Purification and characterization of a novel antioxidant peptide derived from blue mussel (Mytilus edulis) protein hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Ahn, S.Y.; Je, J.Y. Antioxidant and Anti-Inflammatory Activities of Protein Hydrolysates from Mytilus edulis and Ultrafiltration Membrane Fractions. J. Food Biochem. 2014, 38, 460–468. [Google Scholar] [CrossRef]
- Jung, W.K.; Kim, S.K. Isolation and characterization of an anticoagulant oligopeptide from blue mussel, Mytilus edulis. Food Chem. 2009, 117, 687–692. [Google Scholar] [CrossRef]
- Goulas, S.; Gatos, D.; Barlo, K. Convergent solid-phase synthesis of hirudin. J. Pept. Sci. 2006, 12, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Davie, E.W.; Fujikawa, K.; Kisiel, W. The coagulation cascade: Initiation, maintenance, and regulation. Biochemistry 1991, 30, 10363–10370. [Google Scholar] [CrossRef] [PubMed]
- Luchtman-Jones, L.; Broze, G.J. The current status of coagulation. Ann. Med. 1995, 27, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, V.; Sturlese, M.; Cuzzolin, A.; Moro, S. Exploring Protein-Peptide Recognition Pathways Using a Supervised Molecular Dynamics Approach. Structure 2017, 25, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Hamzeh-Mivehroud, M.; Farajzadeh, M.; Dastmalchi, S. Identification of novel peptides against TNF-α using phage display technique and in silico modeling of their modes of binding. Eur. J. Pharm. Sci. 2017, 96, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Garciamora, P.; Martinmartinez, M.; Bonache, M.A.; Gonzalezmuniz, R.; Penas, E.; Frias, J.; Martinezuillaluenga, C. Identification, functional gastrointestinal stability and molecular docking studies of Lentil peptides with dual antioxidant and Angiotensin I converting enzyme inhibitory activities. Food Chem. 2017, 221, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.Y.; Du, J.J.; Jia, J.Q.; Kuang, C. Production of ACE inhibitory peptides from sweet sorghum grain protein using alcalase: Hydrolysis kinetic, purification and molecular docking study. Food Chem. 2016, 199, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.L.; Feng, T.L.; Wang, Z.Y.; Qiao, M.L.; Shahidi, F.; Lu, W.H.; Du, M. Sequence analysis and molecular docking of antithrombotic peptides from casein hydrolysate by trypsin digestion. J. Funct. Foods 2017, 32, 313–323. [Google Scholar] [CrossRef]
- Hao, S.X.; Hou, X.; Wei, L.; Li, J.; Li, Z.H.; Wang, X.T. Extraction and identification of the pigment in the adductor muscle scar of Pacific Oyster Crassostrea gigas. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, S.; Pan, S. Effect of water-soluble chitosan in combination with glutathione on the quality of pen shell adductor muscles. Int. J. Biol. Macromol. 2015, 72, 1250–1253. [Google Scholar] [CrossRef] [PubMed]
- Neumann, T.; Junker, H.D.; Keil, O.; Burkert, K.; Ottleben, H.; Gamer, J.; Sekul, R.; Deppr, H.A.; Feurer, D.; Tomandl, G. Discovery of thrombin inhibitor fragments from chemical microarray screening. Lett. Drug Des. Discov. 2005, 2, 590–594. [Google Scholar] [CrossRef]
- Bode, W. Structure and interaction modes of thrombin. Blood Cells Mol. Dis. 2006, 36, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Biela, A.; Khayat, M.; Tan, H.; Kong, J.; Heine, A.; Hangauer, D.; Klebe, G. Impact of ligand and protein desolvation on ligand binding to the S1 pocket of thrombin. J. Mol. Biol. 2012, 418, 350–366. [Google Scholar] [CrossRef] [PubMed]
- Kjeldahl, J. Neue method zur bestimmung des stickstoffs in organischen körpern. Fresenius J. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Lynch, J.M.; Barbano, D.M. Kjeldahl nitrogen analysis as a reference method for protein determination in dairy products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar] [PubMed]
- Marbaix, H.; Budinger, D.; Dieu, M.; Fumiere, O.; Gillard, N.; Delahaut, P.; Mauro, S.; Raes, M. Identification of proteins and peptide biomarkers for detecting banned Processed Animal Proteins (PAPs) in meat and bone meal by mass spectrometry. J. Agric. Food Chem. 2016, 64, 2405–2414. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Liceaga, A.M. Effect of Microwave Treatments on Antioxidant Activity and Antigenicity of Fish Frame Protein Hydrolysates. Food Bioprocess Technol. 2017, 10, 582–591. [Google Scholar] [CrossRef]
- Hörnaeus, K.; Guillemant, J.; Mi, J.; Hernroth, B.; Bergquit, J.; Lind, B.S. Mass spectrometry data from a quantitative analysis of protein expression in gills of immuno-challenged blue mussels (Mytilus edulis). Data Brief 2016, 8, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Khaket, T.P.; Singh, J. Potential of plant’s dipeptidyl peptidase I & II homologs in generation of ace inhibitory peptides. Int. J. Pept. Res. Ther. 2017, 23, 81–90. [Google Scholar]
- Krieg, R.C.; Dong, Y.; Schwamborn, Y.; Knueche, R. Protein quantification and its tolerance for different interfering reagents using the BCA-method with regard to 2D SDS-PAGE. J. Biochem. Biophys. Methods 2005, 65, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Li, W.; Ullah, K.; Hasan, M.; Linna, G.; Awan, U.; Zhang, Y.Q.; Batool, S.; Deng, H.L. Study of rat hypothalamic proteome by HPLC/ESI ion trap and HPLC/ESI-Q-TOF MS. Proteomics 2013, 13, 2455–2468. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.T.; Tu, M.L.; Qiao, M.L.; Fan, F.J.; Chen, H.; Song, W.; Du, M. Thrombin inhibitory peptides derived from Mytilus edulis proteins: Identification, molecular docking and in silico prediction of toxicity. Eur. Food Res. Technol. 2018, 244, 207–217. [Google Scholar] [CrossRef]
- Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Samperi, R.; Ventura, S.; Chiozzi, R.Z.; Laganà, A. Identification of potential bioactive peptides generated by simulated gastrointestinal digestion of soybean seeds and soy milk proteins. J. Food Compos. Anal. 2015, 44, 205–213. [Google Scholar] [CrossRef]
- Rawendra, R.D.S.; Chang, C.I.; Chen, H.H.; Huang, T.; Hsu, J. A novel angiotensin converting enzyme inhibitory peptide derived from proteolytic digest of Chinese soft-shelled turtle egg white proteins. J. Proteom 2013, 94, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.S. Towards the improved discovery and design of functional peptides: Common features of diverse classes permit generalized prediction of bioactivity. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.Y.; Jia, J.Q.; Yan, H.; Du, J.J.; Gui, Z.Z. A novel angiotensin-I converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zhang, L.W.; Han, X.; Cheng, D.Y. Novel angiotensin I-converting enzyme inhibitory peptides from protease hydrolysates of Qula casein: Quantitative structure-activity relationship modeling and molecular docking study. J. Funct. Foods 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Yang, W.G.; Wang, Z.; Xu, S.Y. A new method for determination of antithrombotic activity of egg white protein hydrolysate by microplatereader. Chin. Chem. Lett. 2007, 18, 449–451. [Google Scholar] [CrossRef]
- Zhang, S.B.; Wang, Z.; Xu, S.Y. Antioxidant and antithrombotic activities of rapeseed peptides. J. Am. Oil Chem. Soc. 2008, 85, 521–527. [Google Scholar] [CrossRef]
- Charles, R.S.T.; Padmanabhan, K.; Arni, R.V.; Padmanabhan, K.P.; Tulinsky, A. Structure of tick anticoagulant peptide at 1.6 Å resolution complexed with bovine pancreatic trypsin inhibitor. Protein Sci. 2000, 9, 265–272. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Length of Peptide | Molecular Weight | Tissues | Peptide Ranker |
|---|---|---|---|---|
| GPAGIIGLIGPK | 12 | 857.4555 | S1, S2, S3 | 0.83 |
| GPIGPAGGKGPTGPK | 15 | 1289.7138 | S2 | 0.83 |
| NDFDKDFFK | 9 | 1174.5336 | S6 | 0.78 |
| AGLQFPVGR | 9 | 569.3174 | S0, S3, S4, S5, S6, S7, S8 | 0.76 |
| GYSAELFR | 8 | 792.3822 | S1 | 0.71 |
| SPNFTKPGK | 9 | 801.4345 | S1 | 0.7 |
| GPTGPQGLR | 9 | 881.4739 | S1, S2 | 0.68 |
| FPGQLNADLR | 10 | 1129.5894 | S3, S5 | 0.67 |
| LAVNMVPFPR | 10 | 1029.5439 | S0, S3, S4, S5, S6, S7, S8 | 0.67 |
| SYSAELFR | 8 | 971.4715 | S3, S4, S6 | 0.65 |
| TATSPFFK | 8 | 897.4602 | S3, S4, S7, S8 | 0.65 |
| KLAVNMVPFPR | 11 | 1270.7253 | S0 | 0.64 |
| AVFPSIVGRPR | 11 | 1197.7004 | S0, S1, S2, S3, S4, S5, S6, S7, S8 | 0.62 |
| LSHYAFSSLR | 10 | 1180.4689 | S5 | 0.62 |
| AAFLLGVNSNDLLK | 14 | 1473.8192 | S3, S6 | 0.59 |
| GIQGPEGELGPVGK | 14 | 1337.7210 | S2 | 0.58 |
| TLYGFGG | 7 | 713.3405 | S5 | 0.58 |
| WSYAPQSR | 8 | 993.4698 | S1, S3, S4, S6 | 0.58 |
| VQQELEDAEERADSAEGSLQK | 21 | 2331.0806 | S6 | 0.57 |
| AGFAGDDAPR | 10 | 975.4435 | S1, S2, S3, S4, S6, S7, S8 | 0.56 |
| SFVNDIFER | 9 | 824.5024 | S0 | 0.56 |
| NLNADVDSVRESLEEEQESKSDLQR | 25 | 2889.364 | S1 | 0.55 |
| SPNFGRPGNASK | 12 | 1230.6178 | S3, S6 | 0.55 |
| ALAADINLR | 9 | 956.5409 | S6 | 0.53 |
| RMEADIAAMQSDLDDALNGQR | 21 | 2319.064 | S1, S6 | 0.5 |
| Peptide | Length of Peptide | Tissue(s) | -CDOCKERENERGY |
|---|---|---|---|
| LTQENFDLQHQVQELDGANAGLAK | 24 | S1 | 311.73 |
| VQQELEDAEERADSAEGSLQK | 21 | S6 | 303.38 |
| TLADLQKEEDKVNHLNK | 17 | S1 | 282.78 |
| MEKENALDRAEQLEQK | 16 | S1 | 272.87 |
| KKLEQDINELEMALDTSNR | 19 | S1 | 270.74 |
| SIQTENDLDNTQTQLQDVQAK | 21 | S6 | 264.94 |
| AKLESTLDEMEDNLER | 16 | S1 | 255.701 |
| ITIQQELEDARSLLEHAER | 19 | S1, S2, S3, S6 | 254.26 |
| LADELRQEQDNYKNAESLR | 19 | S6 | 251.11 |
| KLEQDINELEMALDTSNR | 18 | S1 | 249.98 |
| HQEALNDLTDQLEHMGK | 17 | S1, S2, S3, S4, S6 | 241.79 |
| RRHQEALNDLTDQLEHMGK | 20 | S1, S6 | 238.67 |
| NRLQGELDDLLIEVER | 16 | S1 | 237.02 |
| VKELQTEIDTAHTEAR | 16 | S1 | 236.54 |
| DLEETTLQHEAQVSSLR | 17 | S6 | 236.102 |
| MIEEAEDVASITMNKYR | 17 | S1, S6 | 235.72 |
| RHQEALNDLTDQLEHMGK | 18 | S1, S2, S3, S4 S6 | 232.22 |
| WIAEEADKKYEEAAR | 15 | S1, S2, S3, S6 | 231.80 |
| AAVLEYLAAEVLELAGNAAR | 20 | S0 | 231.28 |
| LLDEEDAASELEGLKK | 16 | S1 | 230.16 |
| NQLIIEIDSLQAMNDGLQK | 19 | S2, S3, S6 | 228.63 |
| LADELRQEQDNYK | 13 | S2, S3, S6 | 227.66 |
| QNLQVQLAAIQSDYDNLNAR | 20 | S6 | 227.36 |
| VIDLEEQLTVVGANIK | 16 | S1, S6 | 226.86 |
| NLAEEIHELTEQLSEGGR | 18 | S6 | 226.02 |
| AAEERADRLQAEVNR | 15 | S2, S3, S6 | 225.98 |
| IRELEDSLDSEREMR | 15 | S1, S4 | 225.89 |
| IRDLENELEADQRR | 14 | S2, S3, S4, S6 | 225.40 |
| MIEEAEDVASITMNK | 15 | S6 | 223.61 |
| ELEDSLDSEREMR | 13 | S2, S3, S6 | 222.18 |
| RMEADIAAMQSDLDDALNGQR | 21 | S1, S6 | 220.49 |
| ENALDRAEQLEQK | 13 | S1, S6 | 220.44 |
| EVDRLEDELLTEK | 13 | S6 | 219.35 |
| EITVRLEEAEAFAQR | 15 | S6 | 217.95 |
| TFDREGQGYISGAEMR | 16 | S1 | 217.67 |
| KLAITEVDLERAEAR | 15 | S1 | 214.16 |
| ATQEVVEELEGVKR | 14 | S1, S2, S3 | 213.487 |
| LTEVQLQVTALTNDKR | 16 | S2, S3, S6 | 213.42 |
| LQGELDDLLIEVER | 14 | S1 | 213.06 |
| AVFVDLEPTVVDEVR | 15 | S5 | 212.988 |
| VQFNLKDYQSSANVKHAVDK | 20 | S4 | 212.98 |
| IRDLENELEADQR | 13 | S1, S2, S3, S4, S6 | 212.26 |
| HQGVMVGMGQKDSYVGDEAQSKR | 23 | S1 | 212.01 |
| ELQTEIDTAHTEAR | 14 | S1 | 211.61 |
| LEDAMGTSTTVSEVSR | 16 | S6 | 211.59 |
| TLQGEMAQQDEQISK | 15 | S1 | 211.51 |
| IAIIITDGKPTDINATQR | 18 | S2, S3, S4 | 211.34 |
| SGVLVRPK | 8 | S0, S8 | 210.81 |
| MSADSKIDALEGSNSR | 16 | S1, S2, S3, S6 | 209.52 |
| DLENELEADQRR | 12 | S2, S3, S6 | 209.289 |
| AQYEETSDTIEALRR | 15 | S1 | 209.038 |
| DLYANTVLSGGTTMFPGIADR | 21 | S6 | 207.485 |
| QLDDTRNQLSVSER | 14 | S6 | 206.48 |
| LTGELEDLGIDVER | 14 | S6 | 206.25 |
| QIAEHEQEIQSLTR | 14 | S6 | 205.14 |
| ELDDVQSQLSHSMK | 14 | S1 | 204.91 |
| QLEDAEHTIGSLTK | 14 | S1 | 204.72 |
| ELEGELDSEQRR | 12 | S6 | 203.33 |
| LAEAEQAAEAANAK | 14 | S1 | 202.27 |
| IRELEDSLDSER | 12 | S2, S3, S4 | 202.17 |
| ALDSMQASLEAEAK | 14 | S2, S3, S6 | 202.096 |
| QVAELTSITDQLTMK | 15 | S6 | 201.63 |
| INELAAQVSSAQAQKR | 16 | S1 | 201.27 |
| DKSALTSQLEEAKR | 14 | S1 | 201.151 |
| KNAENELGEVTVR | 13 | S0, S2, S3, S4, S6 | 200.88 |
| LLSGVTIAQGGVLPNIQAVLLPK | 23 | S0 | 200.55 |
| AKIEDDYNSLQK | 12 | S1, S6 | 198.5 |
| SYYDTSREENDIRR | 14 | S6 | 197.19 |
| SYELPDGQVITIGNER | 16 | S0,S1, S2, S3, S4, S6, S7, S8 | 196.76 |
| VTDLQSELENAQK | 13 | S2, S3, S6 | 196.65 |
| DLENELEADQR | 11 | S6 | 196.13 |
| LDLAGRDLTDYLMK | 14 | S1, S2, S3, S4, S6 | 191.19 |
| KVGINYQPPTVVPGGDLAK | 19 | S0 | 190.22 |
| DIEDLETTLAK | 11 | S1 | 189.759 |
| SALYEDTFIPEVIRPR | 16 | S2, S3, S6 | 188.36 |
| LEDDQSLIAQLQR | 13 | S6 | 188.31 |
| YEEESENASSLR | 12 | S2, S3, S4, S6 | 188.24 |
| MSATFIGNSTAIQELFKR | 18 | S5 | 184.40 |
| NAENELGEVTVR | 12 | S6 | 183.23 |
| AMSIMNSFVNDIFER | 15 | S0, S2, S3, S4, S5, S8 | 178.809 |
| DSYVGDEAQSKR | 12 | S1, S2, S3, S4 | 177.848 |
| SALTSQLEEAKR | 12 | S1 | 177.63 |
| DSYVGDEAQSK | 11 | S1, S2, S3 | 177.572 |
| KRITIQQELEDAR | 13 | S2, S3, S6 | 177.35 |
| KAQSLIDEAEQR | 12 | S2, S3, S4 | 177.22 |
| AQSLIDEAEQR | 11 | S4 | 176.453 |
| ETVQASDEDRR | 11 | S6 | 176.32 |
| QLENENAALQK | 11 | S2, S3 | 175.83 |
| KMEGENSEMKR | 11 | S1 | 174.67 |
| QEYDESGPSIVHR | 13 | S6 | 174.00 |
| LTDEQVDDIIR | 11 | S6 | 173.52 |
| ATQEAVEDLER | 11 | S2, S3 | 173.43 |
| KLAITEVDLER | 11 | S6 | 173.15 |
| SKLQSEVTEINR | 12 | S1 | 172.71 |
| ELEDSLDSER | 12 | S3, S6 | 172.60 |
| ITIQQELEDAR | 11 | S2, S3, S6 | 171.94 |
| LTDMIDKLQSK | 11 | S1 | 171.79 |
| AAFLLGVNSNDLLK | 14 | S2, S3, S6 | 171.476 |
| SLENTIAELQHK | 12 | S2, S3, S6 | 170.94 |
| ENKNLADEIR | 12 | S1 | 170.32 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, M.; Tu, M.; Chen, H.; Mao, F.; Yu, C.; Du, M. Identification and In Silico Prediction of Anticoagulant Peptides from the Enzymatic Hydrolysates of Mytilus edulis Proteins. Int. J. Mol. Sci. 2018, 19, 2100. https://doi.org/10.3390/ijms19072100
Qiao M, Tu M, Chen H, Mao F, Yu C, Du M. Identification and In Silico Prediction of Anticoagulant Peptides from the Enzymatic Hydrolysates of Mytilus edulis Proteins. International Journal of Molecular Sciences. 2018; 19(7):2100. https://doi.org/10.3390/ijms19072100
Chicago/Turabian StyleQiao, Meiling, Maolin Tu, Hui Chen, Fengjiao Mao, Cuiping Yu, and Ming Du. 2018. "Identification and In Silico Prediction of Anticoagulant Peptides from the Enzymatic Hydrolysates of Mytilus edulis Proteins" International Journal of Molecular Sciences 19, no. 7: 2100. https://doi.org/10.3390/ijms19072100