Impact of Long-Term RF-EMF on Oxidative Stress and Neuroinflammation in Aging Brains of C57BL/6 Mice

 ,
,
Abstract
:1. Introduction
2. Results
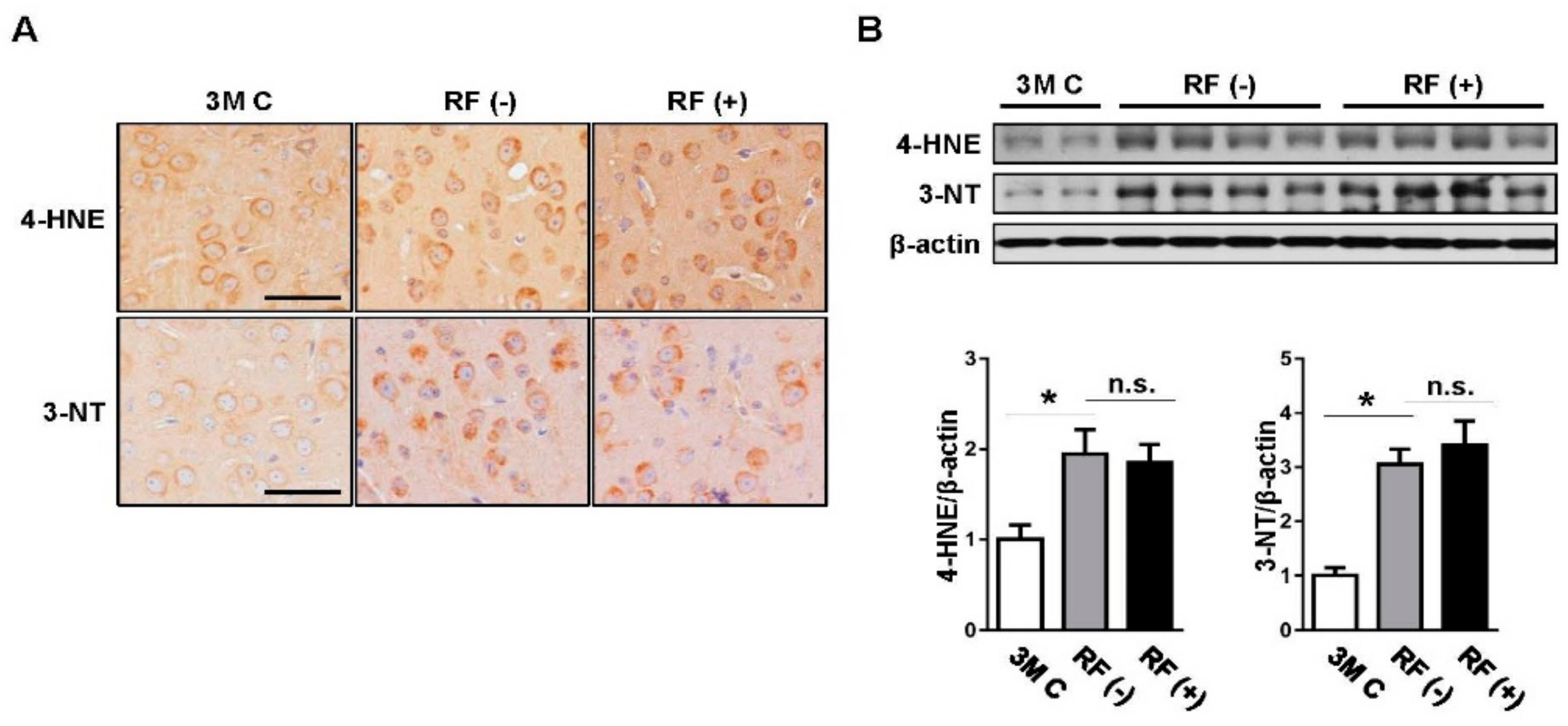
2.1. Effect of Long-Term RF Exposure on Age-Induced Oxidative Stress Markers in the Mouse Brain
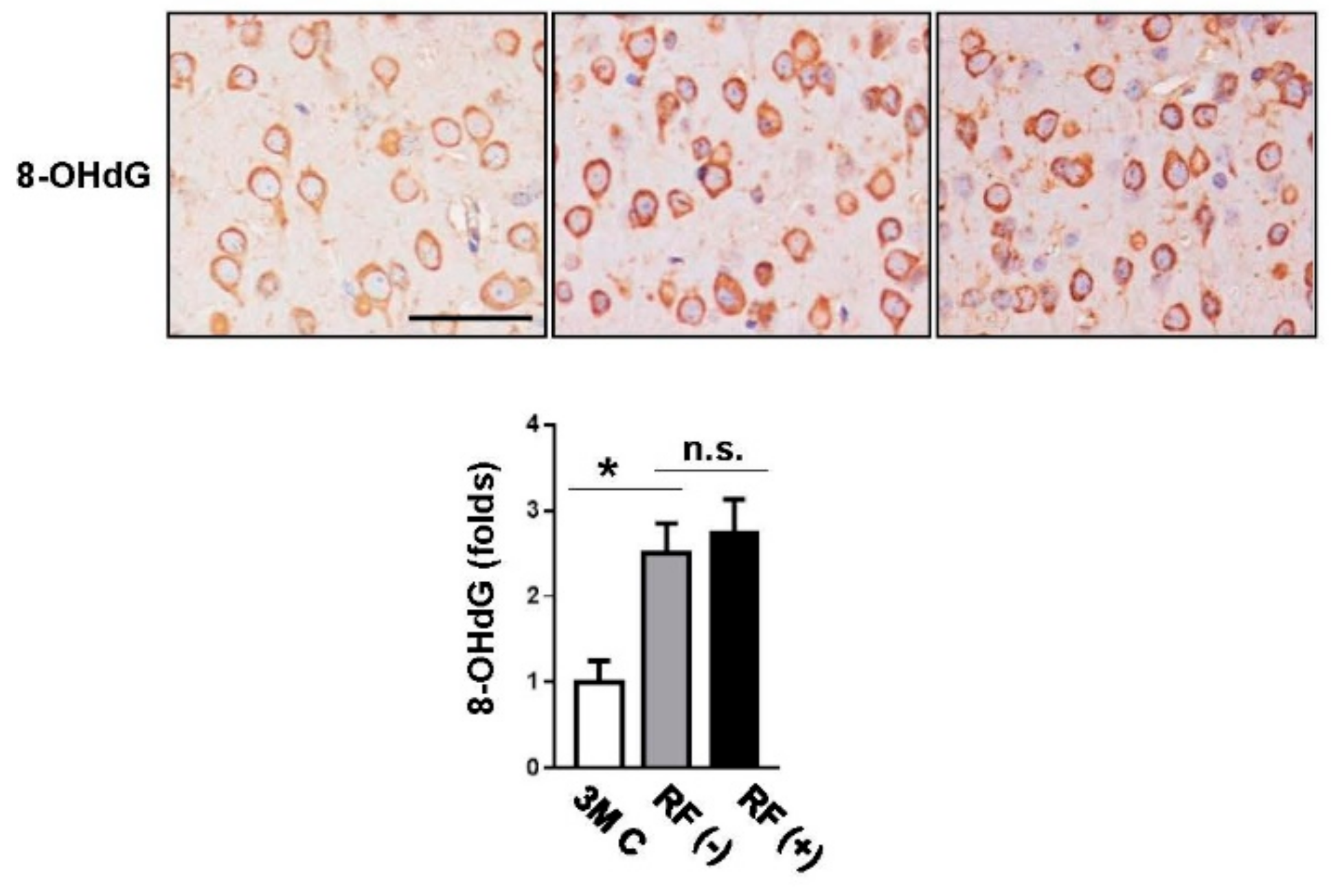
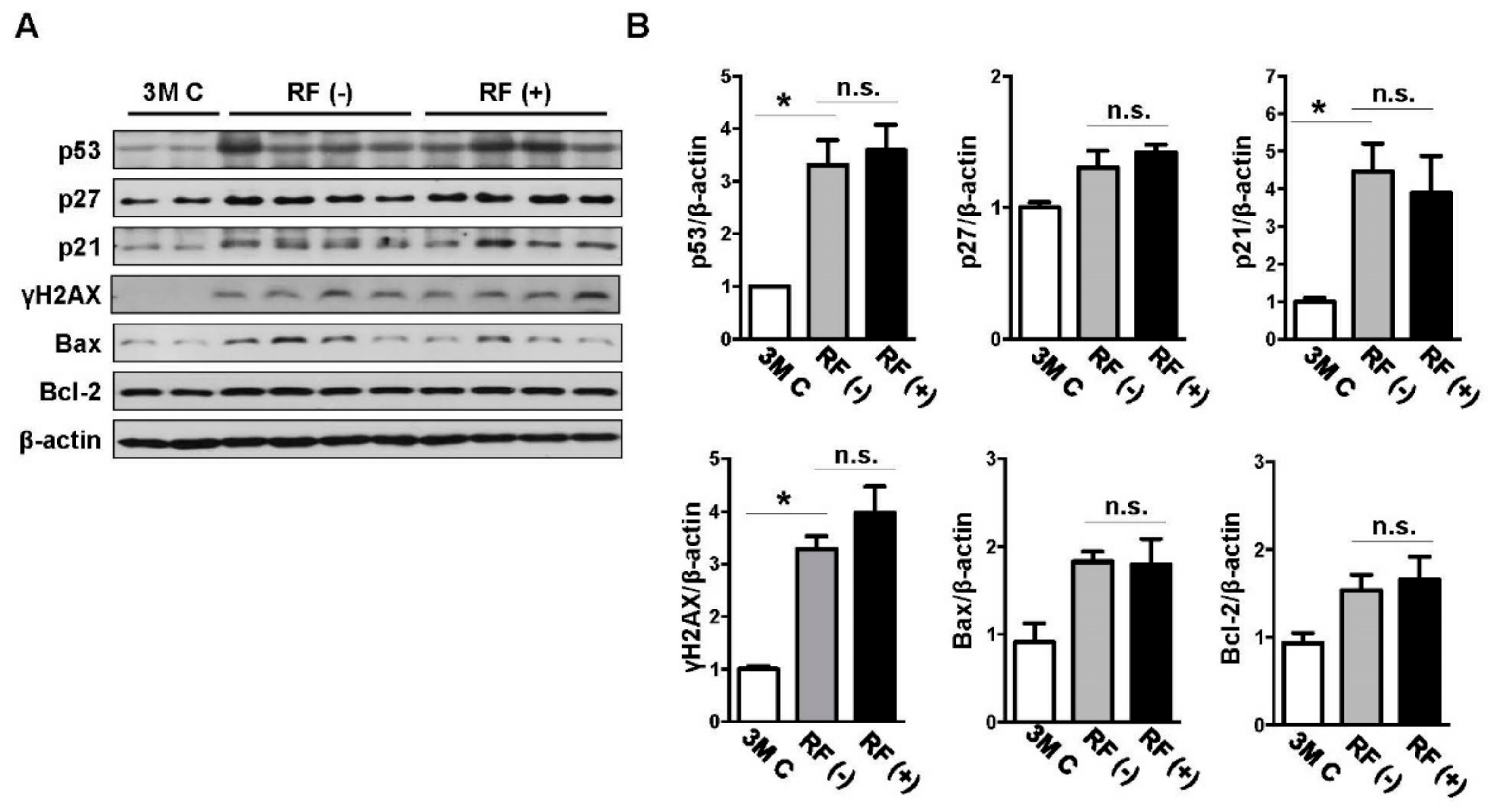
2.2. Effect of Long-Term RF Exposure on Age-Induced DNA Damage
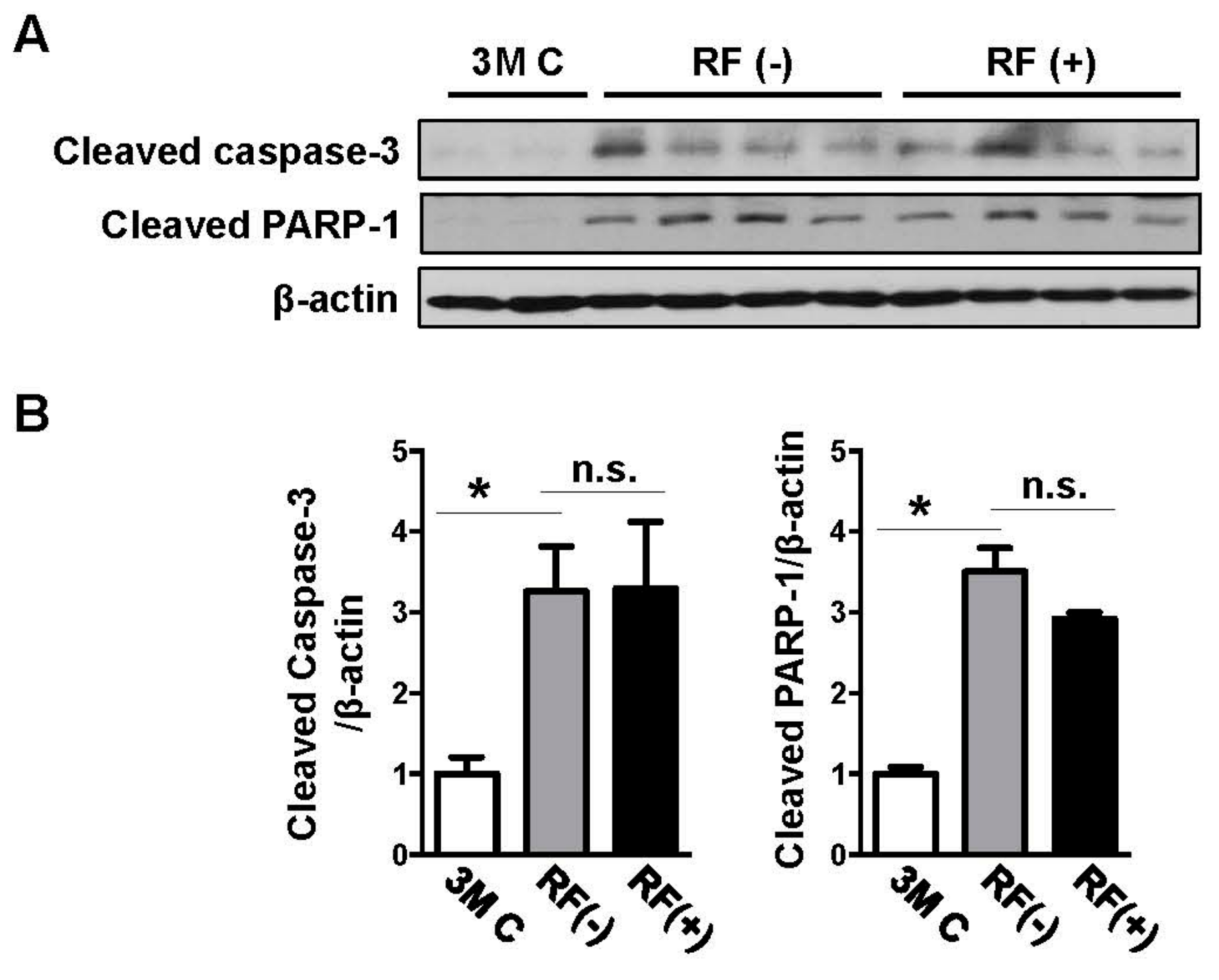
2.3. Effects of Long-Term RF Exposure on Cleaved Caspase-3 and PARP-1 Expression in the Mouse Brain
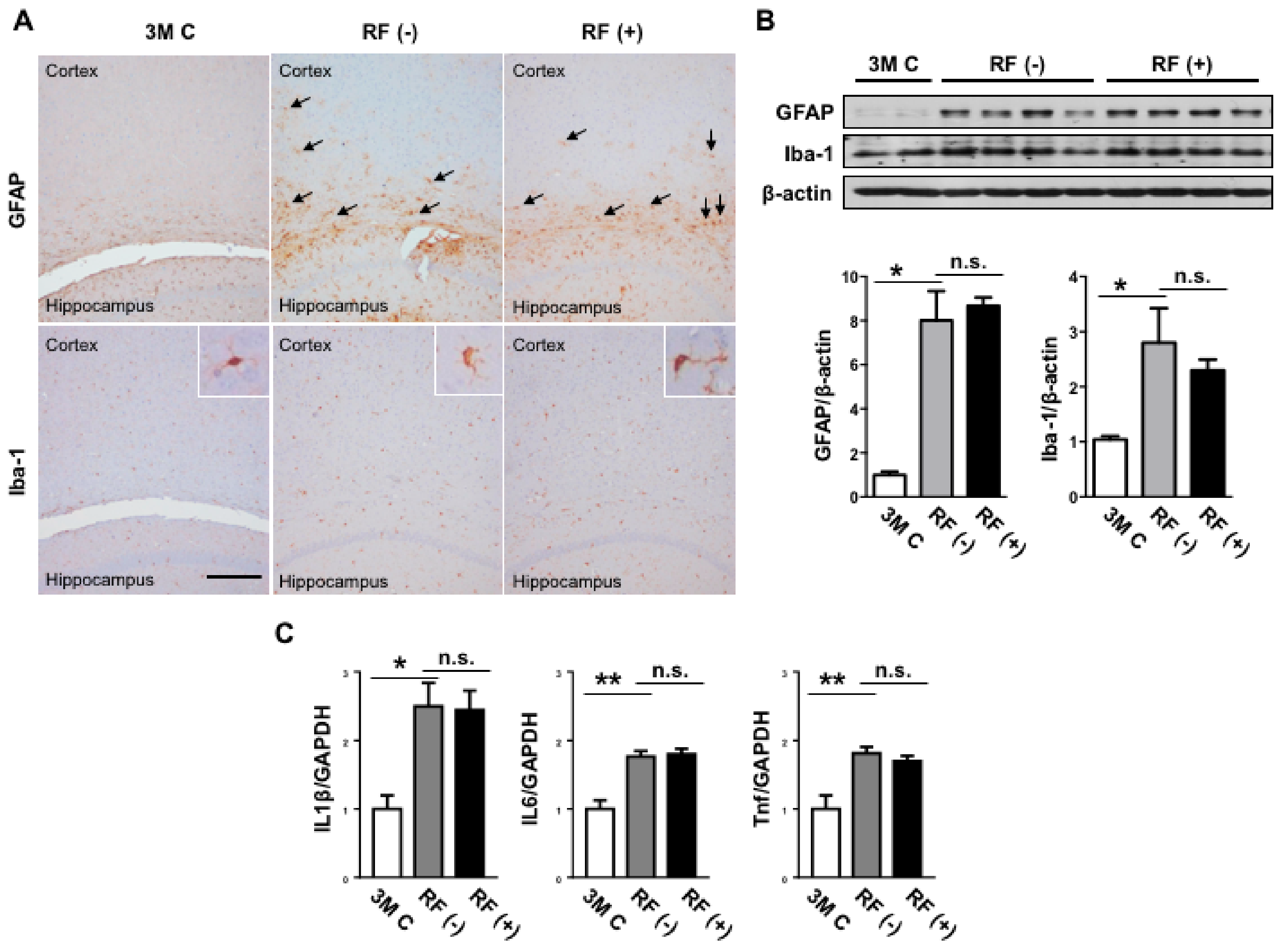
2.4. Effect of Long-Term RF Exposure on GFAP and Iba-1 in the Mouse Brain
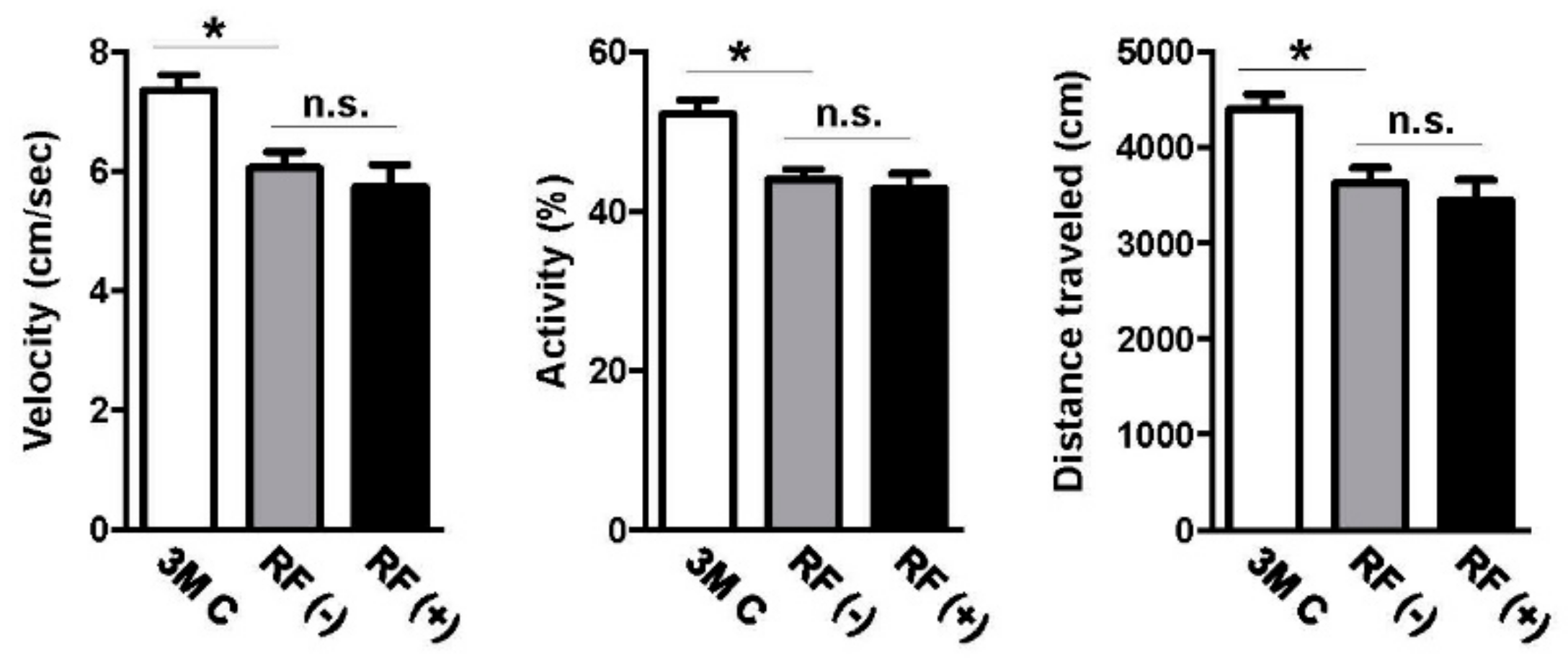
2.5. Effect of Long-Term RF Exposure on Locomotor Activity
3. Discussion
4. Materials and Methods
4.1. Animals
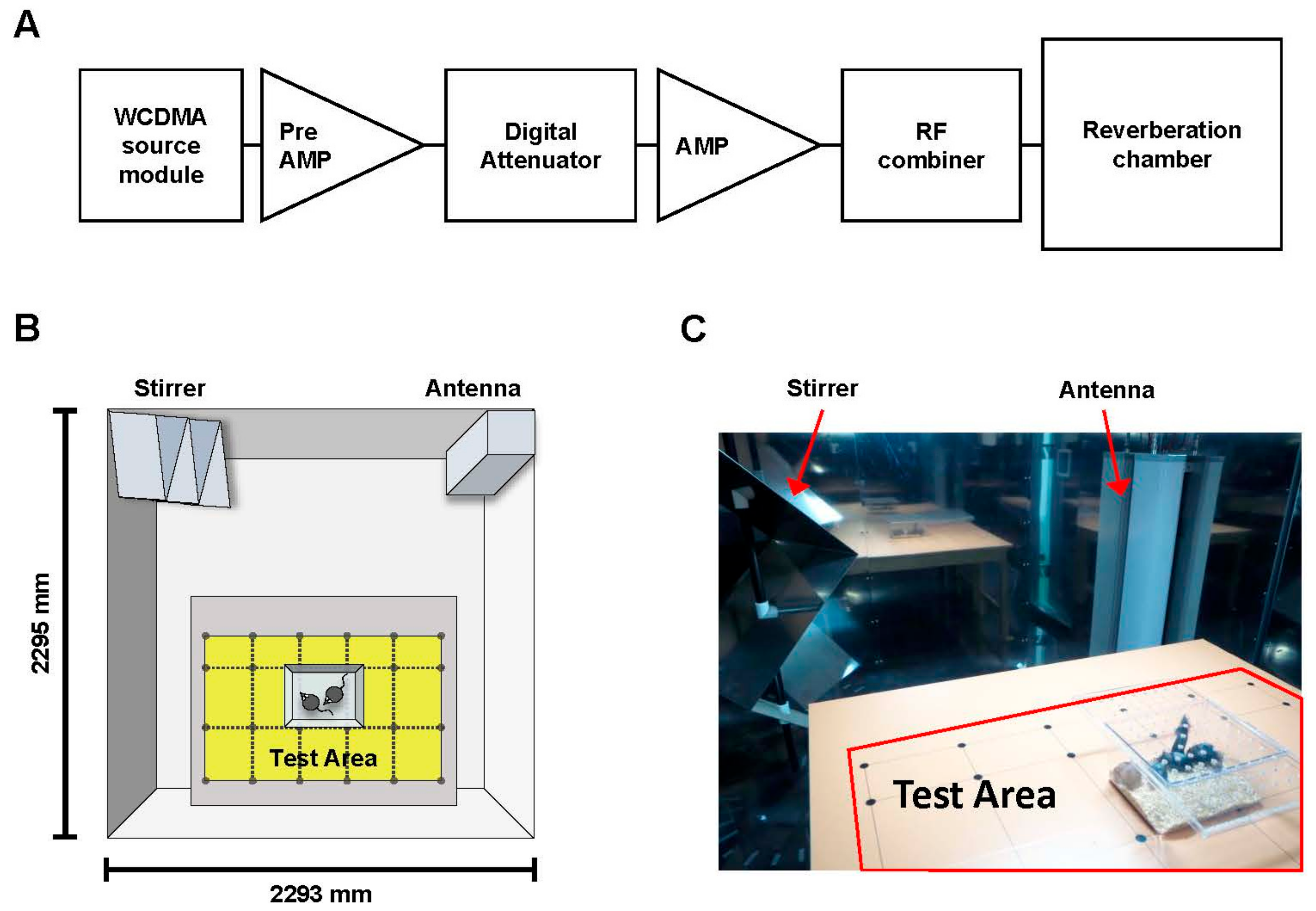
4.2. RF Exposure System
4.3. Sample Preparation
4.4. Immunohistochemistry
4.5. Western Blotting
4.6. Quantitative Real-Time PCR (qRT-PCR)
4.7. Open Field Test
4.8. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mhatre, M.; Floyd, R.A.; Hensley, K. Oxidative stress and neuroinflammation in Alzheimer’s disease and amyotrophic lateral sclerosis: Common links and potential therapeutic targets. J. Alzheimer’s Dis. 2004, 6, 147–157. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xu, Y. p53, oxidative stress, and aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-M.; Zhou, H.; Hong, J.-S. Oxidative stress, neuroinflammation, and neurodegeneration. In Neuroinflammation and Neurodegeneration; Peterson, P.K., Toborek, M., Eds.; Springer: New York, NY, USA, 2014. [Google Scholar]
- Dei, R.; Takeda, A.; Niwa, H.; Li, M.; Nakagomi, Y.; Watanabe, M.; Inagaki, T.; Washimi, Y.; Yasuda, Y.; Horie, K.; et al. Lipid peroxidation and advanced glycation end products in the brain in normal aging and in Alzheimer’s disease. Acta Neuropathol. 2002, 104, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Gemma, C.; Mesches, M.H.; Sepesi, B.; Choo, K.; Holmes, D.B.; Bickford, P.C. Diets enriched in foods with high antioxidant activity reverse age-induced decreases in cerebellar beta-adrenergic function and increases in proinflammatory cytokines. J. Neurosci. 2002, 22, 6114–6120. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, K. 4-hydroxynonenal and neurodegenerative diseases. Mol. Asp. Med. 2003, 24, 293–303. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.M.; Chung, Y.H.; Kim, M.J.; Lee, E.Y.; Kim, E.G.; Cha, C.I. Age-related changes in the distribution of nitrotyrosine in the cerebral cortex and hippocampus of rats. Brain Res. 2002, 931, 194–199. [Google Scholar] [CrossRef]
- Sloane, J.A.; Hollander, W.; Moss, M.B.; Rosene, D.L.; Abraham, C.R. Increased microglial activation and protein nitration in white matter of the aging monkey. Neurobiol. Aging 1999, 20, 395–405. [Google Scholar] [CrossRef]
- Tohgi, H.; Abe, T.; Yamazaki, K.; Murata, T.; Ishizaki, E.; Isobe, C. Remarkable increase in cerebrospinal fluid 3-nitrotyrosine in patients with sporadic amyotrophic lateral sclerosis. Ann. Neurol. 1999, 46, 129–131. [Google Scholar] [CrossRef]
- Agarwal, S.; Sohal, R.S. Aging and protein oxidative damage. Mech. Ageing Dev. 1994, 75, 11–19. [Google Scholar] [CrossRef]
- Richter, C.; Park, J.W.; Ames, B.N. Normal oxidative damage to mitochondrial and nuclear DNA is extensive. Proc. Natl. Acad. Sci. USA 1988, 85, 6465–6467. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, P.; MacGarvey, U.; Kaufman, A.E.; Koontz, D.; Shoffner, J.M.; Wallace, D.C.; Beal, M.F. Oxidative damage to mitochondrial DNA shows marked age-dependent increases in human brain. Ann. Neurol. 1993, 34, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T. Ageing, neurodegeneration and brain rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, K.; Wu, W.; Wang, K.; Ni, S.; Ye, P.; Yu, Y.; Ye, J.; Sun, L. Electromagnetic noise inhibits radiofrequency radiation-induced DNA damage and reactive oxygen species increase in human lens epithelial cells. Mol. Vis. 2008, 14, 964–969. [Google Scholar] [PubMed]
- Xu, S.; Zhou, Z.; Zhang, L.; Yu, Z.; Zhang, W.; Wang, Y.; Wang, X.; Li, M.; Chen, Y.; Chen, C.; et al. Exposure to 1800 MHz radiofrequency radiation induces oxidative damage to mitochondrial DNA in primary cultured neurons. Brain Res. 2010, 1311, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kesari, K.K.; Behari, J.; Kumar, S. Mutagenic response of 2.45 GHz radiation exposure on rat brain. Int. J. Radiat. Biol. 2010, 86, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Stronati, L.; Testa, A.; Moquet, J.; Edwards, A.; Cordelli, E.; Villani, P.; Marino, C.; Fresegna, A.M.; Appolloni, M.; Lloyd, D. 935 MHz cellular phone radiation. An in vitro study of genotoxicity in human lymphocytes. Int. J. Radiat. Biol. 2006, 82, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Malyapa, R.S.; Ahern, E.W.; Bi, C.; Straube, W.L.; LaRegina, M.; Pickard, W.F.; Roti Roti, J.L. DNA damage in rat brain cells after in vivo exposure to 2450 MHz electromagnetic radiation and various methods of euthanasia. Radiat. Res. 1998, 149, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, P.; Ernst, H.; Huuskonen, H.; Komulainen, H.; Kumlin, T.; Maki-Paakkanen, J.; Puranen, L.; Juutilainen, J. No effects of radiofrequency radiation on 3-chloro-4-(dichloromethyl)-5-hydroxy-2(5H)-furanone-induced tumorigenesis in female Wistar rats. Radiat. Res. 2006, 166, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Misa Agustino, M.J.; Leiro, J.M.; Jorge Mora, M.T.; Rodriguez-Gonzalez, J.A.; Jorge Barreiro, F.J.; Ares-Pena, F.J.; Lopez-Martin, E. Electromagnetic fields at 2.45 GHz trigger changes in heat shock proteins 90 and 70 without altering apoptotic activity in rat thyroid gland. Biol. Open 2012, 1, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.S.; He, G.L.; Hao, Y.T.; Xiao, Y.; Chen, C.H.; Zhang, G.B.; Yu, Z.P. Exposure to 2.45 GHz electromagnetic fields elicits an HSP-related stress response in rat hippocampus. Brain Res. Bull. 2012, 88, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Watilliaux, A.; Edeline, J.M.; Leveque, P.; Jay, T.M.; Mallat, M. Effect of exposure to 1800 MHz electromagnetic fields on heat shock proteins and glial cells in the brain of developing rats. Neurotox. Res. 2011, 20, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Ammari, M.; Gamez, C.; Lecomte, A.; Sakly, M.; Abdelmelek, H.; de Seze, R. GFAP expression in the rat brain following sub-chronic exposure to a 900 MHz electromagnetic field signal. Int. J. Radiat. Biol. 2010, 86, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Finnie, J.W.; Cai, Z.; Manavis, J.; Helps, S.; Blumbergs, P.C. Microglial activation as a measure of stress in mouse brains exposed acutely (60 minutes) and long-term (2 years) to mobile telephone radiofrequency fields. Pathology 2010, 42, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Bodera, P.; Stankiewicz, W.; Antkowiak, B.; Paluch, M.; Kieliszek, J.; Sobiech, J.; Niemcewicz, M. Influence of electromagnetic field (1800 MHz) on lipid peroxidation in brain, blood, liver and kidney in rats. Int. J. Occup. Med. Environ. Health 2015, 28, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Ragy, M.M. Effect of exposure and withdrawal of 900-MHz-electromagnetic waves on brain, kidney and liver oxidative stress and some biochemical parameters in male rats. Electromagn. Biol. Med. 2015, 34, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Dasdag, S.; Akdag, M.Z. The link between radiofrequencies emitted from wireless technologies and oxidative stress. J. Chem. Neuroanat. 2016, 75, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Tai, J.L.; Li, G.Q.; Zhang, Z.W.; Xue, J.H.; Liu, H.S.; Zhu, H.; Cheng, J.D.; Liu, Y.L.; Li, A.M.; et al. Exposure to 1950-MHz TD-SCDMA electromagnetic fields affects the apoptosis of astrocytes via caspase-3-dependent pathway. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Tohidi, F.Z.; Bahreyni-Toosi, M.H.; Azimian, H.; Khademi, S.; Fardid, R.; Anani Sarab, G. The gene expression level of p53 and p21 in mouse brain exposed to radiofrequency field. Int. J. Radiat. Res. 2015, 13, 337–343. [Google Scholar]
- Akdag, M.Z.; Dasdag, S.; Canturk, F.; Karabulut, D.; Caner, Y.; Adalier, N. Does prolonged radiofrequency radiation emitted from Wi-Fi devices induce DNA damage in various tissues of rats? J. Chem. Neuroanat. 2016, 75, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, F.; Dasdag, S.; Akdag, M.Z.; Kilinc, N. Whole-body exposure of radiation emitted from 900 MHz mobile phones does not seem to affect the levels of anti-apoptotic bcl-2 protein. Electromagn. Biol. Med. 2008, 27, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Lynch, A.M.; Murphy, K.J.; Deighan, B.F.; O’Reilly, J.A.; Gun’ko, Y.K.; Cowley, T.R.; Gonzalez-Reyes, R.E.; Lynch, M.A. The impact of glial activation in the aging brain. Aging Dis. 2010, 1, 262–278. [Google Scholar] [PubMed]
- Luo, X.G.; Ding, J.Q.; Chen, S.D. Microglia in the aging brain: Relevance to neurodegeneration. Mol. Neurodegener. 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arellano, J.J.; Parpura, V.; Zorec, R.; Verkhratsky, A. Astrocytes in physiological aging and Alzheimer’s disease. Neuroscience 2016, 323, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Mausset-Bonnefont, A.L.; Hirbec, H.; Bonnefont, X.; Privat, A.; Vignon, J.; de Seze, R. Acute exposure to GSM 900-MHz electromagnetic fields induces glial reactivity and biochemical modifications in the rat brain. Neurobiol. Dis. 2004, 17, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Ammari, M.; Brillaud, E.; Gamez, C.; Lecomte, A.; Sakly, M.; Abdelmelek, H.; de Seze, R. Effect of a chronic GSM 900 MHz exposure on glia in the rat brain. Biomed. Pharmacother. 2008, 62, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; He, M.; Zhang, Y.; Xu, S.; Zhang, L.; He, Y.; Chen, C.; Liu, C.; Pi, H.; Yu, Z.; et al. Differential pro-inflammatory responses of astrocytes and microglia involve STAT3 activation in response to 1800 MHz radiofrequency fields. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Bouji, M.; Lecomte, A.; Gamez, C.; Blazy, K.; Villegier, A.S. Neurobiological effects of repeated radiofrequency exposures in male senescent rats. Biogerontology 2016, 17, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Saikhedkar, N.; Bhatnagar, M.; Jain, A.; Sukhwal, P.; Sharma, C.; Jaiswal, N. Effects of mobile phone radiation (900 MHz radiofrequency) on structure and functions of rat brain. Neurol. Res. 2014, 36, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, D.; Jay, T.; Edeline, J.M. Head-only exposure to GSM 900-MHz electromagnetic fields does not alter rat’s memory in spatial and non-spatial tasks. Behav. Brain Res. 2003, 145, 51–61. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tsurita, G.; Ueno, S.; Watanabe, S.; Wake, K.; Taki, M.; Nagawa, H. 1439 MHz pulsed TDMA fields affect performance of rats in a T-maze task only when body temperature is elevated. Bioelectromagnetics 2003, 24, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Jeong, Y.J.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. 1950 MHz radiofrequency electromagnetic fields do not aggravate memory deficits in 5xFAD mice. Bioelectromagnetics 2016, 37, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendash, G.W.; Sanchez-Ramos, J.; Mori, T.; Mamcarz, M.; Lin, X.; Runfeldt, M.; Wang, L.; Zhang, G.; Sava, V.; Tan, J.; et al. Electromagnetic field treatment protects against and reverses cognitive impairment in Alzheimer’s disease mice. J. Alzheimer’s Dis. 2010, 19, 191–210. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Jeong, Y.J.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. The effect of sub-chronic whole-body exposure to a 1950 MHz electromagnetic field on the hippocampus in the mouse brain. JEES 2015, 15, 151–157. [Google Scholar]
- Klose, M.; Grote, K.; Spathmann, O.; Streckert, J.; Clemens, M.; Hansen, V.W.; Lerchl, A. Effects of early-onset radiofrequency electromagnetic field exposure (GSM 900 MHz) on behavior and memory in rats. Radiat. Res. 2014, 182, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.B.; Choi, H.D.; Kim, B.C.; Pack, J.K.; Kim, N.; Lee, Y.S. Effects of simultaneous combined exposure to CDMA and WCDMA electromagnetic fields on serum hormone levels in rats. J. Radiat. Res. 2013, 54, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.A.; Ahn, H.S.; Lee, Y.S.; Gye, M.C. Effect of a 20 kHz sawtooth magnetic field exposure on the estrous cycle in mice. J. Microbiol. Biotechnol. 2007, 17, 398–402. [Google Scholar] [PubMed]
- Aydin, M.; Cevik, A.; Kandemir, F.M.; Yuksel, M.; Apaydin, A.M. Evaluation of hormonal change, biochemical parameters, and histopathological status of uterus in rats exposed to 50-Hz electromagnetic field. Toxicol. Ind. Health 2009, 25, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Hata, K.; Yamaguchi, H.; Tsurita, G.; Wake, K.; Watanabe, S.; Taki, M.; Ueno, S.; Nagawa, H. Short-term exposure to a 1439-MHz TDMA signal exerts no estrogenic effect in rats. Bioelectromagnetics 2010, 31, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Van Deventer, E.; van Rongen, E.; Saunders, R. WHO research agenda for radiofrequency fields. Bioelectromagnetics 2011, 32, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jin, Y.B.; Kim, T.H.; Pack, J.K.; Kim, N.; Choi, H.D.; Lee, J.S.; Lee, Y.S. The effects of simultaneous combined exposure to CDMA and WCDMA electromagnetic fields on rat testicular function. Bioelectromagnetics 2012, 33, 356–364. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence | Product Size (bp) | Source Sequence ID |
|---|---|---|---|
| IL-1β | F: 5′-ACCTTTTGACAGTGATGAGAA-3′ R: 5′-GCTGCTGCGAGATTTGA-3′ | 128 | NM_008361.4 |
| IL-6 | F: 5′-CCTTCCCTACTTCACAAGTC-3′ R: 5′-TTTTCTGCAAGTGCATCATC-3′ | 187 | NM_001314054.1 |
| TNF | F: 5′-TGGGTTGTACCTTGTCTACT-3′ R: 5′-TGGTATGAGATAGCAAATCGG-3′ | 102 | NM_013693.3 |
| GAPDH | F: 5′-CAAGAAGGTGGTGAAGCAGG-3′ R: 5′-AGGTGGAAGAGTGGGAGTTG-3′ | 110 | NM_008084.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.J.; Son, Y.; Han, N.-K.; Choi, H.-D.; Pack, J.-K.; Kim, N.; Lee, Y.-S.; Lee, H.-J. Impact of Long-Term RF-EMF on Oxidative Stress and Neuroinflammation in Aging Brains of C57BL/6 Mice. Int. J. Mol. Sci. 2018, 19, 2103. https://doi.org/10.3390/ijms19072103
Jeong YJ, Son Y, Han N-K, Choi H-D, Pack J-K, Kim N, Lee Y-S, Lee H-J. Impact of Long-Term RF-EMF on Oxidative Stress and Neuroinflammation in Aging Brains of C57BL/6 Mice. International Journal of Molecular Sciences. 2018; 19(7):2103. https://doi.org/10.3390/ijms19072103
Chicago/Turabian StyleJeong, Ye Ji, Yeonghoon Son, Na-Kyung Han, Hyung-Do Choi, Jeong-Ki Pack, Nam Kim, Yun-Sil Lee, and Hae-June Lee. 2018. "Impact of Long-Term RF-EMF on Oxidative Stress and Neuroinflammation in Aging Brains of C57BL/6 Mice" International Journal of Molecular Sciences 19, no. 7: 2103. https://doi.org/10.3390/ijms19072103