Selected Molecular Mechanisms Involved in the Parasite–Host System Hymenolepis diminuta–Rattus norvegicus

 , , , , and
, , , , and 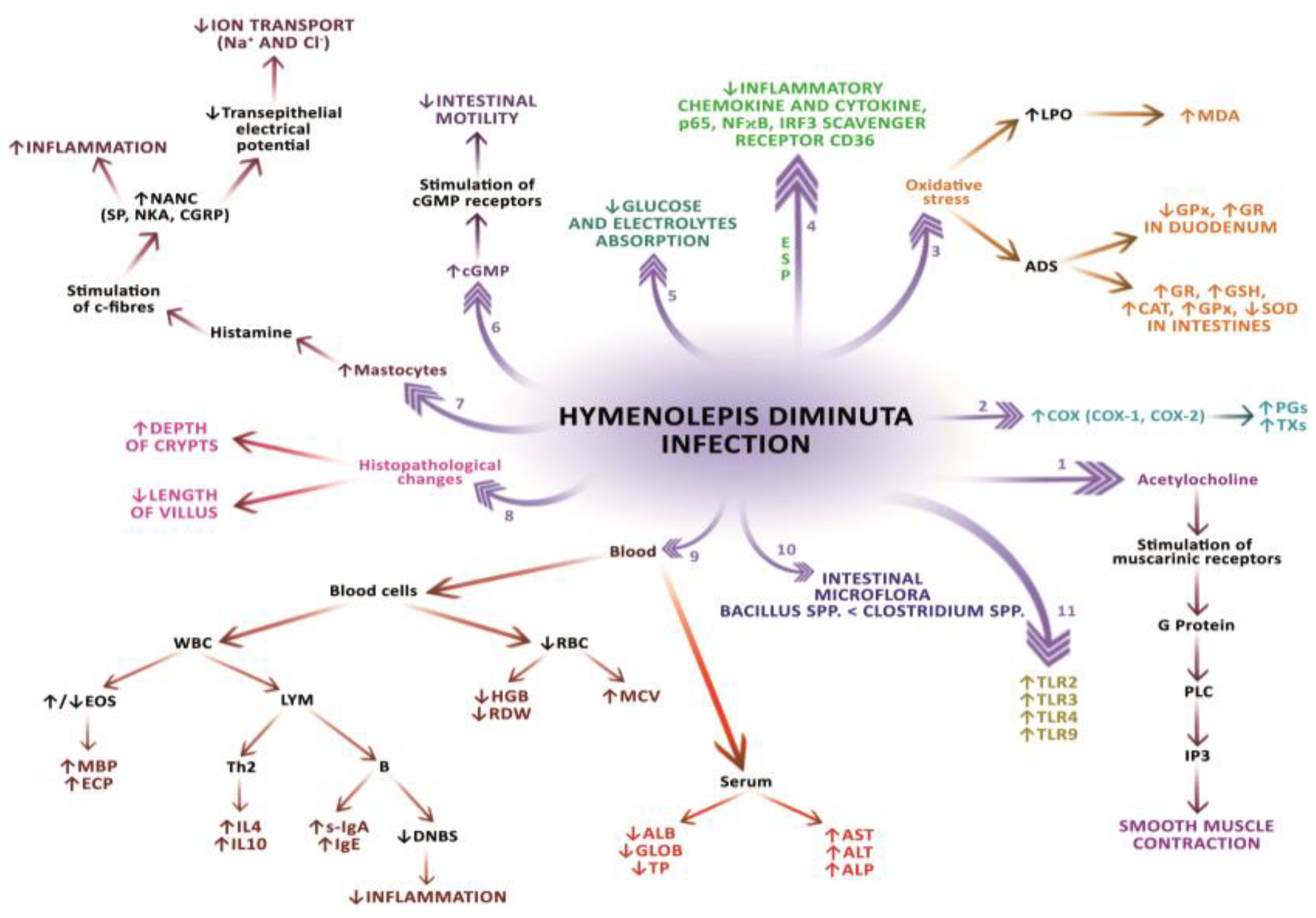{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Rat Tapeworm Hymenolepis diminuta Rudolphi, 1819
2.1. Morphological and Histomorphological Changes in the Host’s Digestive Tract
2.2. Pathophysiological and (Sub-)Cellular Changes in the Host’s Digestive Tract
2.3. Influence of Gastrointestinal Flora on the Course of H. Diminuta Infection
2.4. Changes in Ion Transport in the Host’s Digestive Tract
2.5. Morphological and Biochemical Changes in the Host’s Blood
2.6. Changes in the Host’s Antioxidant Enzymes Activity
2.7. Changes in Cyclooxygenase Activity and Development of Inflammatory Reaction
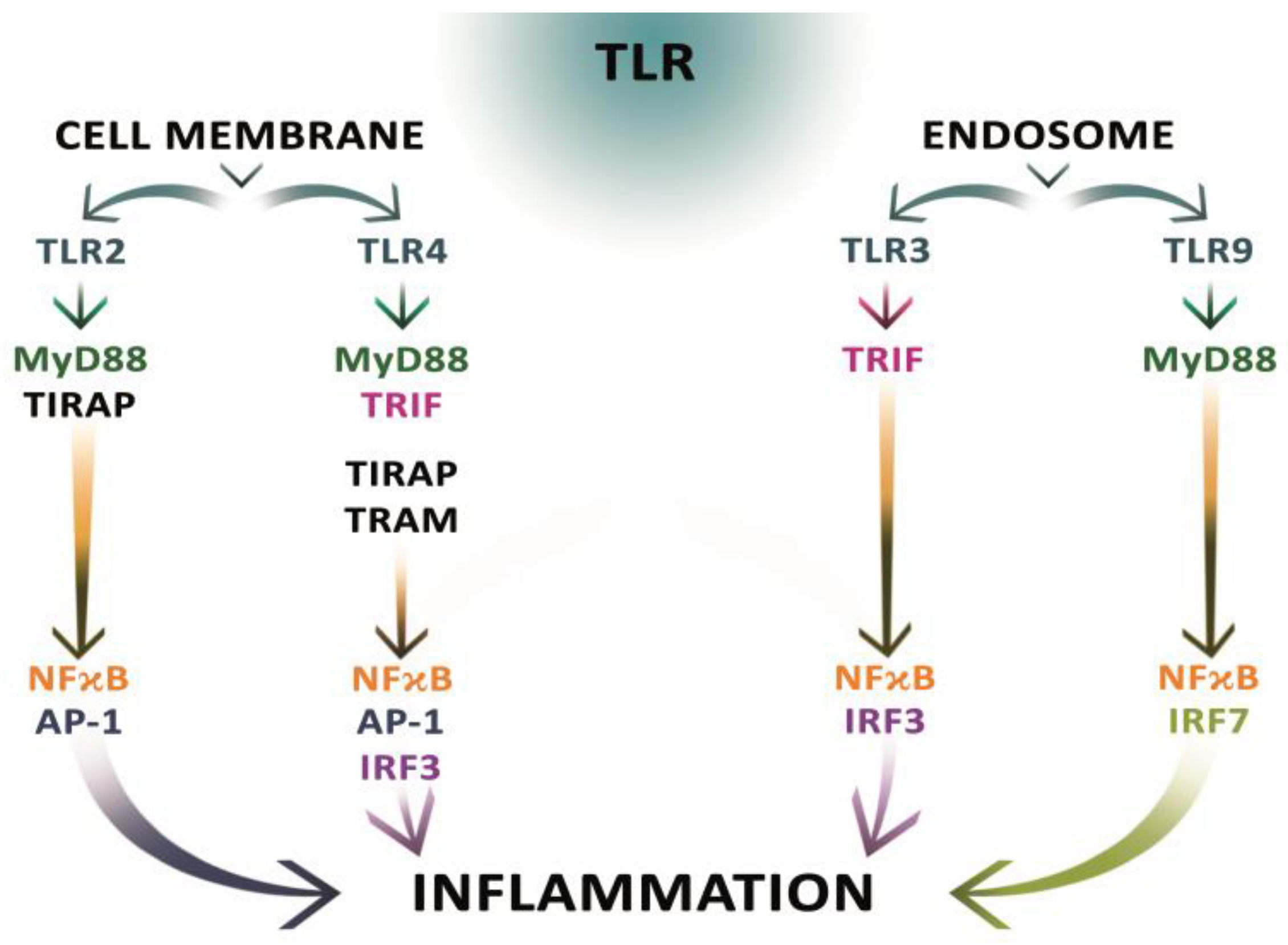
2.8. Changes in the Expression of Toll-Like Receptors and the Mechanisms of Immune Response
3. Conclusions and Further Research Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pojmańska, T.; Niewiadomska, K. Parasites—Inconvenient element in the structure of ecosystem food web. Kosmos 2010, 59, 99–110. [Google Scholar]
- Hadaś, E.; Derda, M. Parasites are still dangerous. Probl. Hig. Epidemiol. 2014, 95, 6–13. [Google Scholar]
- Przyjałkowski, Z. Studies on the settlement of Aspicularis tetraptera Nitzsch 1821 (Nematoda, Oxyuridae) in non-bacterial mice. Acta Parasitol. Pol. 1972, 20, 389–395. [Google Scholar]
- Vythilingam, I. Plasmodium knowlesi and Wuchereria bancrofti: Their vectors and challenges for the future. Front Physiol. 2012, 3, 115. [Google Scholar] [CrossRef] [PubMed]
- Zammarchi, L.; Strohneyer, M.; Bartalesi, F.; Bruno, E.; Munoz, J.; Buonfrate, D.; Nicoletti, A.; Garcia, H.H.; Pozio, E.; Bartoloni, A. Epidemiology and management of cysticercosis and Taenia solium taeniasis in Europe, Systematic review 1990–2011. PLoS ONE 2013, 8, e69537. [Google Scholar] [CrossRef]
- Wędrychowicz, H. Evasion of host immunity by parasites. Kosmos 2005, 54, 39–48. [Google Scholar]
- Nazimek, K.; Bryniarski, K. The biological activity of macrophages in health and disease. Postepy Hig. Med. Dosw. 2012, 66, 507–520. [Google Scholar] [CrossRef]
- Niścigórska, J. Liver in infectious diseases. Post. Nauk. Med. 2000, 1, 27–30. [Google Scholar]
- Przybyła, A.; Nowak, M.; Krzowska-Firych, J. Selected parasitic diseases of the liver. Hepatol. Pol. 2014, 14, 115–122. [Google Scholar]
- Rutkowska, J. Regulation of parasite-host immune response interactions on the cellular level. Now Lek. 2007, 76, 251–255. [Google Scholar]
- Bień, J.; Sałamatin, R.; Sulima, A.; Savijoki, K.; Conn, D.B.; Nareaho, A.; Młocicki, D. Mass spectrometry analysis of the excretory-secretory (E-S) products of the model cestode Hymenolepis diminuta reveals their immunogenic properties and the presence of New E-S proteins in cestodes. Acta Parasitol. 2016, 61, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.J.; Wang, A.; Catarino, M.E.D.; Ball, L.; Phan, V.C.; MacDonald, J.A.; McKay, D.M. Extracts of the rat tapeworm, Hymenolepis diminuta, suppres macrophage activation in vitro and alleviate chemically induced colitis in mice. Infect. Immun. 2010, 78, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites: Masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.S.; Taylor, M.D.; Balic, A.; Finney, C.A.; Lamb, J.R.; Maizels, R.M. Suppression of allergic airway inflammation by helminth-induced regulatory T cells. J. Exp. Med. 2005, 202, 1199–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwinell, M.B.; Wise, R.M.; Bas, P.; Oaks, J.A. Hymenolepis diminuta: Mucosal mastocytosis and intestinal smooth muscle hypertrophy occur in tapeworm-infected rats. Exp. Parasitol. 1998, 89, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.; Singh, S.M.; Kataria, M.; Somvanshi, R. Clinicopathological studies on spontaneous Hymenolepis diminuta infection in wild and laboratory rats. Braz. J. Vet. Pathol. 2011, 4, 103–111. [Google Scholar]
- Ishih, A.; Uchikawa, R. Immunoglobulin E and mast cells responses are related to worm biomass but not expulsion of Hymenolepis diminuta during low dose infection in rats. Parasite Immunol. 2000, 22, 561–566. [Google Scholar] [CrossRef] [PubMed]
- McKey, D.M.; Halton, D.W.; Johnston, C.F.; Shaw, C.; Fairweather, I.; Buchanan, K.D. Hymenolepis diminuta: Changes in the levels of certain intestinal regulatory peptides in infected C57 mice. Exp. Parasitol. 1991, 73, 15–26. [Google Scholar] [CrossRef]
- Read, C.P.; Kilejian, A.Z. Circadian migratory behavior of a cestode symbiote in the rat host. J. Parasitol. 1969, 55, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Reardon, C.; Sanchez, A.; Hogaboam, C.M.; McKay, D.M. Tapeworm infection reduces epithelial ion transport abnormalities in murine dextran sulfate sodium-induced colitis. Infect. Immun. 2001, 69, 4417–4423. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.L.; Roberts, L.S. In vivo effects of putative crowding factors on development of Hymenolepis diminuta. J. Parasitol. 1991, 77, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, C.A.; Andreassen, J. Inhibition of growth of a tapeworm Hymenolepis diminuta in its normal host (rat). Int. J. Parasitol. 1991, 21, 47–55. [Google Scholar] [CrossRef]
- Stradowski, M. Effects of inbreeding in Hymenolepis diminuta (Cestoda). Acta Parasitol. 1994, 39, 146–149. [Google Scholar]
- Zavras, E.T.; Roberts, L.S. Developmental physiology of cestodes: Cyclic nucleotides and the identity of putative crowding factors in Hymenolepis diminuta. J. Parasitol. 1985, 71, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, J.; Bennet-Jenkins, E.M.; Bryant, C. Immunology and biochemistry of Hymenolepis diminuta. Adv. Parasitol. 1999, 42, 243–275. [Google Scholar]
- Malmberg, M.; Andreassen, J.; Malmberg, G. Hymenolepis diminuta: A comparison between young developing and small, destrobilated worms in the rat intestine. Z. Parasitenkd 1985, 71, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Sulima, A.; Bień, J.; Savijoki, K.; Nareaho, A.; Sałamatin, R.; Conn, D.B.; Młocicki, D. Identification of immunogenic proteins of the cysticercoid of Hymenolepis diminuta. Parasit. Vectors 2017, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.W.; Barley, A.J. Beetle-to-beetle transmission and dispersal of Hymenolepis diminuta (Cestoda) eggs via the feces of Tenebrio molitor. J. Parasitol. 1999, 85, 384–385. [Google Scholar] [CrossRef] [PubMed]
- Makki, M.S.; Mowlavi, G.; Shahbazi, F.; Abai, M.R.; Najafi, F.; Hosseini-Farash, B.R.; Teimoori, S.; Hasanpour, H.; Naddaf, S.R. Identification of Hymenolepis diminuta Cysticercoid Larvae in Tribolium castaneum (Coleoptera: Tenebrionidae) Beetles from Iran. J. Arthropod Borne Dis. 2017, 11, 338–343. [Google Scholar] [PubMed]
- Kołodziej, P.; Rzymowska, J.; Stępień-Rukasz, H.; Lorencowicz, R.; Lucińska, M.; Dzióbek, M. Analysis of a child infected with Hymenolepis diminuta in Poland. Ann. Agric. Environ. Med. 2014, 21, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Mowlavi, G.H.; Mobedi, I.; Mamishi, S.; Rezaeian, M.; Haghi Ashtiani, M.T.; Kashi, M. Hymenolepis diminuta (Rodolphi, 1819) infection in a child from Iran. Iran. J. Publ. Health 2008, 37, 120–122. [Google Scholar]
- Patamia, I.; Cappello, E.; Castellano-Chiodo, D.; Greco, F.; Nigro, L.; Cacopardo, B. A human case of Hymenolepis diminuta in child from Eastern Sicily. Korean J. Parasitol. 2010, 48, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Rohela, M.; Ngui, R.; Li, Y.A.L.; Kalaichelvan, B.; Wan Hafiz, W.I.; Mohd Redzuan, A.N. A case report of Hymenolepis diminuta infection in a Malaysian child. Trop. Biomed. 2012, 29, 224–230. [Google Scholar] [PubMed]
- Tena, D.; Perez Simon, M.; Gimeno, C.; Perez Pomata, M.T.; Illescas, S.; Amondarin, I.; Gonzales, A.; Dominguez, J.; Bisquert, J. Human infection with Hymenolepis diminuta: Case report from Spain. J. Clin. Microbiol. 1998, 36, 2375–2376. [Google Scholar] [PubMed]
- Fal, W.; Czaplicka, H. Effect of experimental hymenolepiasis on various tissue reactions in rats. Wiad. Parazytol. 1991, 37, 331–342. [Google Scholar] [PubMed]
- Hopkins, C.A.; Subramanian, G.; Stallard, H. The development of Hymenolepis diminuta in primary and secondary infections in mice. Parasitology 1972, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Read, C.P. Longevity of the tapeworm, Hymenolepis diminuta. J. Parasitol. 1967, 53, 1055–1056. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.G.; Turton, J.A. Antibody response to tapeworm (Hymenolepis diminuta) in the rat. Nature 1973, 246, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Chappell, L.H.; Pike, A.W. Loss of Hymenolepis diminuta from the rat. Int. J. Parasitol. 1976, 6, 333–339. [Google Scholar] [CrossRef]
- Andreassen, J. Immunity to adult cestodes: Basic knowledge and vaccination problems. A review. Parassitologia 1991, 33, 45–53. [Google Scholar] [PubMed]
- Featherston, D.W.; Wakelin, D.; Lammas, D.A. Inflammatory responses in the intestine during tapeworm infections. Mucosal mast cells and mucosal mast cell proteases in Sprague-Dawley rats infected with Hymenolepis diminuta. Int. J. Parasitol. 1992, 22, 961–966. [Google Scholar] [CrossRef]
- Hesselberg, C.A.; Andreassen, J. Some influences of population density on Hymenolepis diminuta in rats. Parasitology 1975, 71, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Hoque, T.; Bhogal, M.; Webb, R.A. Validation of intestinal controls for gene expression analysis in the intestine of rats infected with Hymenolepis diminuta. Parasitol Int. 2007, 56, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Kolasa, A. Histopathological changes in small and large intestines during hymenolepidosis in rats. Folia Biol. Krakow 2012, 60, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Holland, C. Scanning electron microscopic studies of the mucosa of rats infected with Hymenolepis diminuta. J. Helminth 1984, 58, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Starke, W.A.; Oaks, J.A. Ileal mucosal mast cell, eosinophil, and goblet cell populations during Hymenolepis diminuta infection of the rat. J Parasitol. 2001, 87, 1222–1225. [Google Scholar] [CrossRef]
- Firlotte, W.R. A survey of the parasites of the brown Norway rat. Can. J. Comp. Med. Vet. Sci. 1948, 12, 187–191. [Google Scholar] [PubMed]
- Goswami, R.; Somvanshi, R.; Singh, S.M.; Singh, S. A preliminary survey on incidence of helminthic and protozoal diseases in rats. Indian J. Vet. Pathol. 2009, 33, 90–92. [Google Scholar]
- Sadjjadi, S.M.; Massoud, J. Helminth parasites of wild rodents in Khuzestan Province, South-West of Iran. J. Vet. Parasitol. 1999, 13, 55–56. [Google Scholar]
- Waugh, C.A.; Lindo, J.F.; Foronda, P.; Angeles-Santana, M.; Lorenzo-Morales, J.; Robinson, R.D. Population distributin and zoonotic potential of gastrointestinal helminthes of wild rats Rattus rattus and Rattus norvegicus from Jamaica. J. Parasitol. 2006, 92, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Webster, J.P.; Macdonald, D.W. Parasites of wild brown rats (Rattus norvegicus) on UK farms. Parasitology 1995, 111, 247–255. [Google Scholar] [CrossRef]
- Hindsbo, O.; Andreassen, J.; Ruitenberg, J. Immunological and histopathological reactions of the rat against the tapeworm Hymenolepis diminuta and the effects anti-thymocyte serum. Parasite Immunol. 1982, 4, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Dudzińska, E. Influence of the intestinal micro biota on the development of irritable bowel syndrome. Med. Środow 2016, 19, 70–76. [Google Scholar]
- Hart, A.; Kamm, M.A. Review article: Mechanisms of initiation and perpetuation of gut inflammation by stress. Aliment. Pharmacol. Ther. 2002, 16, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L. Prostaglandin biology in inflammatory bowel disease. Gastroenterol. Clin. N. Am. 2001, 30, 971–980. [Google Scholar] [CrossRef]
- Rocha, S.M.; Cristovao, A.C.; Campos, F.L.; Fonseca, C.P.; Baltazar, G. Astrocyte—Derived GNDF is a potent inhibitor of microglial activation. Neurobiol. Dis. 2012, 47, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Meir, M.; Flemming, S.; Burkard, N.; Wagner, J.; Germer, C.T.; Schlegel, N. The glial cell-line devired neurotrophic factor: A novel regulator of intestinal barier function in health and disease. An. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G1118–G1123. [Google Scholar] [CrossRef] [PubMed]
- Starke-Buzetti, W.A.; Oaks, J.A. Increased glial-derived neurotrophic factor in the small intestine of rats infected with the tapeworm, Hymenolepis diminuta. Int. J. Exp. Pathol. 2008, 89, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Malbon, C.C. G proteins indevelopment. Nat. Rev. Mol. Cell Biol. 2005, 6, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Bikopoulos, G.J.; Hoque, T.; Webb, R.A. Infection with the cestode Hymenolepis diminuta induces changes in acetylcholine metabolism and muscarinic receptor mRNA expression in the rat jejunum. Parasitol. Res. 2006, 99, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Zeiss, M.M.; Harris, N.L. Interactions between the intestinal microbiome and helminth parasites. Parasie Immunol. 2016, 38, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Berrilli, F.; Di Care, D.; Cavallero, S.; D’Amelio, S. Interactions between parasites and microbial communities in the human gut. Front Cell Infect. Microbiol. 2012, 2, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przyjałkowski, Z. Effect of intestinal bacteria on the host response in Trichinella spiralis infection. Acta Parasitol. Pol. 1978, 25, 287–292. [Google Scholar]
- Stefański, W. Bacterial flora as one of the factors influencing the settling of parasites in the intestines of their hosts. Acta Parasitol. Pol. 1965, 12, 1–6. [Google Scholar]
- Houser, B.B.; Burns, W.C. Experimental infection of gnotobiotic Tenebrio molitor and white rats with Hymenolepis diminuta (Cestoda: Cyclophyllidea). J. Parasitol. 1968, 54, 69–73. [Google Scholar] [CrossRef] [PubMed]
- McKenney, E.A.; Williamson, L.; Yoder, A.D.; Rawls, J.F.; Bilbo, S.D.; Parker, W. Alteration of the rat cecal microbiome during colonization with the helminth Hymenolepis diminuta. Gut. Microbes. 2015, 69, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Wegener Parfreg, L.; Jirku, M.; Sima, R.; Jalorecka, M.; Sak, B.; Grigore, K.; Jirku Pomajbikova, K. A benign helminth alters the host immune system and the gut microbiota in a rat model system. PLoS ONE 2017, 12, e0182205. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Goderska, K.; Grajek, K.; Grajek, W. The in vitro models of gastrointestinal tract to study bioavailability of nutriments. Zywn Nauk Technol. J. A 2006, 1, 30–45. [Google Scholar]
- Evers, B.M. Small Intestine. In Chirurgia Sabistona; Popiela, T., Ed.; Urban & Partner: Bratislava, Slovakia, 2013; pp. 165–187. [Google Scholar]
- Kosik-Bogacka, D.; Baranowska-Bosiacka, I.; Salamatin, R. Hymenolepis diminuta: Effect of infection on ion transport in colon and blood picture of rats. Exp. Parasitol. 2010, 124, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Kolasa, A.; Baranowska-Bosiacka, I.; Marchlewicz, M. Hymenolepis diminuta: The effects of infection on transepithelial ion transport and tight junctions in rat intestines. Exp. Parasitol. 2011, 127, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Furnes, J.B.; Kunze, W.A.; Clere, N. Nutrient tasting and signaling mechanisms in the gut. II. The intestine as a sensory organ: Neutral, endocrine and immune responses. Am. J. Physiol. 1999, 277, 922–928. [Google Scholar] [CrossRef]
- Schultheiss, G.; Hennig, B.; Schunack, W.; Prinz, G.; Diener, M. Histamine-induced ion secretion across rat distal colon: In volvement of histamine H1 and H2 receptors. Eur. J. Pharmacol. 2006, 546, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, N.P.; Brownfield, M.S.; DeVente, J.; Bass, P.; Oaks, J.A. cGMP secreted from the tapeworm Hymenolepis diminuta is a signal molecule to the host intestine. J. Parasitol. 2001, 94, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Kunzelman, K.; Mall, M. Electrolyte transport in the mammalian colon: Mechanisms and implications for disease. Physiol. Rev. 2002, 82, 245–289. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Kołodziejczyk, L. Ion transport in the colon of rats experimentally infected with liver fluke (Fasciola hepatica). Folia Biol. 2004, 52, 243–246. [Google Scholar] [CrossRef]
- O’Malley, K.E.; Sloan, T.; Joyce, P.; Baird, A.W. Type 1 hypersensitivity reactions in intestinal mucosae from rats infected with Fasciola hepatica. Parasite Immunol. 1993, 15, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Palmer, J.M.; Cooke, H.J. Neuroimmune regulation of colonic secretion in guinea pigs. Am. J. Physiol. 1991, 260, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Podesta, R.B.; Mettrick, D.F. The effect of bicarbonate and acidification on water and electrolyte absorpyion by the intestine of normal and infected (Hymenolepis diminuta: Cestoda) rats. Am. J. Dig. Dis. 1974, 19, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Gill, N.; Shaikh, A.A.; Khan, M.M.; Memon, M.S. Influence on intestinal cestodes on the blood picture of brown rats (Rattus norvegicus) of Hyderabad Sindh, Pakistan, Pak. J. Biol. Sci. 2007, 10, 4479–4484. [Google Scholar] [CrossRef]
- Sommerfelt, I.E.; Santillan, G.; Mira, G.; Ribicich, M.; De Torres, R. Toxocara canis infection in a pig model: Immunological, haematological and blood biochemistry responses. J. Helminthol. 2006, 80, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Weller, P.F. Eosinophilia in travelers. Med. Clin. N. Am. 1992, 76, 1413–1432. [Google Scholar] [CrossRef]
- Habior, A. Acute hepatic failure. Post. Nauk. Med. 2014, 1, 24–30. [Google Scholar]
- Mathur, C.S.; Johnson, S. Blood profile of house rats naturally infected with Vampirolepis fraternal (Cestoda). Indian J. Helminthol. 1989, 4, 14–16. [Google Scholar]
- Moshin, M.; Rahmam, M.M.; Das, P.M.; Fazul Haque, A.K.M. Haematological observations in cattle naturally infected with Fasciola gigantic. Bangladesh Vet. 1991, 8, 31–34. [Google Scholar] [CrossRef]
- Jarret, E.E.; Miller, H.R. Production and activities of IgE in helminth infection. Prog. Allergy 1982, 31, 178–233. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Protection Against Oxidants in Biological Systems: The Superoxide Theory of Oxygen Toxicity. In Free Radical in Biology and Medicine; Clarendon Press: Oxford, UK, 1989; pp. 86–123. [Google Scholar]
- Ohshima, H.; Bartsch, H. Chronic infections and inflammatory processes as cancer risk factors: Possible role of nitric oxide in carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar] [CrossRef]
- Esterbauer, H.; Puhl, H.; Dieber-Rotheneder, M.; Waeg, G.; Rabl, H. Effect of antioxidants on oxidative modification of LDL. Ann. Med. 1991, 23, 573–581. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.J.; Bittelmeyer, A.M.; Raess, B.U. Irreversible inhibition of plasma membrane (Ca2+ + Mg2+)-ATPase and Ca2+ transport by 4-OH-2, 3-trans-nonenal. Arch. Biochem. Biophys. 1999, 361, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Czajka, A. Reactive oxygen species and mechanisms of body protection. Now Lek 2006, 75, 582–586. [Google Scholar]
- Selvan, J.P.; Aranganathan, S.; Gopalan, R.; Nalini, N. Chemopreventive efficacy of pronyl-lysine on lipid peroxidation and antioxidant status in rat colon carcinogenesis. Fund. Clin. Pharmacol. 2009, 23, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Niwa, A.; Miyazato, T. Reactive oxygen intermediates from eosinophils in mice infected with Hymenolepis nana. Parasite Immunol. 1996, 18, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Menzies, M.; Reverter, A.; Andronicos, N.; Hunt, P.; Windon, R.; Ingham, A. Nematode challenge induces differential expression of oxidant, antioxidant and mucous genes down the longitudinal axis of the sheep gut. Parasite Immunol. 2010, 32, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Baranowska-Bosiacka, I.; Noceń, I.; Jakubowska, K.; Chlubek, D. Hymenolepis diminuta: Activity of anti-oxidant enzymes in defferent parts of rat gastrointestinal tract. Exp. Parasitol. 2011, 128, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Skrzycki, M.; Majewska, M.; Podsiad, M.; Czeczot, H.; Salamatin, R.; Twarowska, J.; Grytner-Zięcina, B. Hymenolepis diminuta: Experimental studies on the antioksidant system with short and long term infection periods in the rats. Exp. Parasitol. 2011, 129, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Sulima, A.; Savijoki, K.; Bień, J.; Nareako, A.; Sałamatin, R.; Conn, D.R.; Młocicki, D. Comparative proteomic analysis of Hymenolepis diminuta cysticercoid and adult sta ges. Front. Microbiol. 2017, 8, 2672. [Google Scholar] [CrossRef] [PubMed]
- Burdan, F.; Chałas, A.; Szumiło, J. Cyclooxygenase and prostanoids biological implications. Postepy Hig. Med. Dosw. 2006, 60, 129–141. [Google Scholar]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenase 1 and 2. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Baranowska-Bosiacka, I.; Kolasa-Wołosiuk, A.; Łanocha-Arendarczyk, N.; Gutowska, I.; Korbecki, J.; Namięta, H.; Rotter, I. The inflamatory effect of infection with Hymenolepis diminuta via the increased expression and activity of COX-1 and COX-2 in the rat jejunum and colon. Exp. Parasitol. 2016, 169, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kargman, S.; Charleson, S.; Cartwright, M.; Frank, J.; Riendean, D.; Mancini, I.; Evans, J.; O’Neill, G. Characterization of Prostaglandin G/H Synthase 1 and 2 in rat, dog, monkey and human gastrointestinal tracts. Gastroenterology 1996, 111, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.K.; Abdel-Hafeez, E.H.; Kamal, A.M. Some studies on spontaneous Hymenolepis diminuta infection in laboratory rats. J. Egypt Soc. Parasitol. 2015, 45, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Gieryńska, M.; Kalinowska-Gacek, E. Immunological alert at the mucosal sites. Part II. Życie Wet. 2009, 84, 115–122. [Google Scholar]
- Garrett, W.S.; Gordon, J.I.; Glimcher, L.H. Homeostasis and inflammation in the intestine. Cell 2010, 140, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska-Gacek, E.; Gieryńska, M. Immunological alert at mucosal surfaces. Part I. Życie Weterynaryjne 2009, 84, 17–20. [Google Scholar]
- Sanz, Y.; De Palma, G. Gut microbiota and probiotics in modulation of epithelium and gut-associated lymphoid tissue function. Int. Rev. Immunol. 2009, 28, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennant, S.M.; Hartland, E.L.; Phumoonna, T.; Lyras, D.; Rood, J.I.; Robins-Browne, R.M.; van Driel, I.R. Influence of gastric acid on susceptibility to infection with ingested bacterial pathogens. Infect. Immun. 2008, 76, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Niedworok, M.; Sordyl, B.; Borecka, A.; Gawor, J.; Małecka-Panas, E. Estimation of eosinophilia, immunoglobulin E and eosinophilic cationic protein concentration during the treatment of toxocariasis. Wiad Parazytol 2008, 54, 225–230. [Google Scholar] [PubMed]
- Zawistowska-Denziak, A.; Basałaj, K.; Strojny, B.; Młocicki, D. New data on human macrophages polarization by Hymenolepis diminuta tapeworm—An in vitro study. Front. Immunol. 2017, 8, 148. [Google Scholar] [CrossRef] [PubMed]
- Aira, N.; Andersson, A.M.; Singh, S.K.; McKay, D.M.; Blomgran, R. Species dependent impact of helminth-derived antigens on human macrophages infected with Mycobacterium tuberculosis: Direct effect on the innate anti-mycobacterial response. PLoS Negl Trop. Dis. 2017, 11, e0005390. [Google Scholar] [CrossRef] [PubMed]
- Graepel, R.; Leung, G.; Wang, A.; Villemaire, M.; Jirik, F.R.; Sharkey, K.A.; McDougall, J.J.; McKay, D.M. Murine autoimmune arthritis is exaggerated by infection with the rat tapeworm, Hymenolepis diminuta. Int. J. Parasitol. 2013, 43, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Matisz, C.E.; Leung, G.; Reyes, J.L.; Wang, A.; Sharkey, K.A.; McKay, D.M. Adoptive transfer of helminth antigen-pulsed dendritic cells protects against the development of experimental colitis in mice. Eur. J. Immunol. 2015, 45, 3126–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melon, A.; Wang, A.; Phan, V.; McKay, D.M. Infection with Hymenolepis diminuta is more effective than daily corticosteroids in blocking chemically induced colitis in mice. J. Biomed. Biotechnol. 2010, 2010, 384523. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.L.; Fernando, M.R.; Lopes, F.; Leung, G.; Mancini, N.L.; Matisz, C.E.; Wang, A.; McKay, D.M. IL-22 restrains tapeworm-mediated protection against experimental colitis via regulation of IL-25 expression. PLoS Pathog. 2016, 12, e1005481. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.L.; Wang, A.; Fernando, M.R.; Graepel, R.; Leung, G.; van Rooijen, N.; Sigvardsson, M.; McKay, D.M. Splenic B cells from Hymenolepis diminuta-infected mice ameliorate colitis independent of T cells and via cooperation with macrophages. J. Immunol. 2015, 194, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Polińska, B.; Matowicka-Karna, J.; Kemona, H. The cytokines in inflammatory bowel disease. Postepy Hig. Med. Dosw. 2009, 63, 389–394. [Google Scholar]
- Wang, A.; McKay, D.M. Immune modulation by a high molecular weight fraction from the rat tapeworm Hymenolepis diminuta. Parasitology 2005, 130, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Lopes, F.; Shute, A.; Wang, A.; McKay, D.M. Young mice expel the tapeworm Hymenolepis diminuta and are protected from colitis by triggering a memory response with worm antigen. Am. J. Physiol. Gastrointest Liver Physiol. 2018, 314, G461–G470. [Google Scholar] [CrossRef] [PubMed]
- Smyth, K.; Morton, C.; Mathew, A.; Karuturi, S.; Haly, C.; Zhang, M.; Holzknecht, Z.E.; Swanson, C.; Lin, S.S.; Parker, W.; et al. Production and Use of Hymenolepis diminuta Cysticercoids as Anti-Inflammatory Therapeutics. J. Clin. Med. 2017, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.M.; Wallace, J.L. Acetic acid induced ulceration in rats is not affected by infection with Hymenolepis diminuta. J. Parasitol. 2009, 95, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Sauter, B.; Albert, M.L.; Francisco, L.; Larson, M.; Somersan, S.; Bhardwaj, N. Consequences of cell death: Exposure to necrotic tumor cells, but not primary tissue cells or apoptotic cells, induces the maturation of immunostimulatory dendritic cells. J. Exp. Med. 2000, 191, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Vabulas, R.M.; Ahmad-Nejad, P.; Ghose, S.; Kirschning, C.J.; Issels, R.D.; Wagner, H. HSP70 as endogenous stimulus of the Toll/interleukin-1 receptor signal pathway. J. Biol. Chem. 2002, 277, 15107–15112. [Google Scholar] [CrossRef] [PubMed]
- Pachówka, M.; Kluk, M.; Korczak-Kowalska, G. Role of Toll-like receptors (TLR) in induction and regulation of immune response. Postepy Biol. Komorki 2009, 36, 429–442. [Google Scholar]
- Ortega-Cava, C.F.; Ishihar, S.; Rumi, M.A.; Kawashima, K.; Ishimura, N.; Kazumori, H.; Udagawa, J.; Kadowaki, Y.; Kinoshita, Y. Strategic compartmentalization of Toll-like receptor 4 in the mouse gut. J. Immunol. 2003, 170, 3977–3985. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Hemmi, H.; Miyamoto, H.; Moriishi, K.; Tamura, S.; Takaku, H.; Akira, S.; Matsuura, Y. Involvement of the Toll-like receptor 9 signaling pathway in the induction of innate immunity by baculovirius. J. Virol. 2005, 79, 2847–2858. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Functions of Toll-like receptors: Lessons from KO mice. C. R. Biol. 2004, 327, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, E.; Zouain, C.S.; Vanhoutte, F.; Fontaine, J.; Pavelka, N.; Thieblemont, N.; Willems, F.; Ricciardi-Castagnoli, P.; Goldman, M.; Capron, M.; et al. Double-stranded RNAs from the helminth parasite Schistosoma activate TLR3 in dendritic cells. J. Biol. Chem. 2005, 280, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Ashkar, A.A.; Rosenthal, K.L. Toll-like receptor 9, CpG DNA and innate immunity. Curr. Mol. Med. 2002, 2, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Furrie, E.; Macfarlane, S.; Thomson, G.; MacFarlane, G.T. Toll-like receptors -2, -3, and -4 expression patterns on human colon and their regulation by mucosal-associated bacteria. Immunology 2005, 115, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C.A., Jr. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Passare, C.; Medzhitov, R. Toll-like receptors: Linking innate and adaptive immunity. Nat. Rev. Immunol. 2004, 6, 1382–1387. [Google Scholar] [CrossRef] [PubMed]
- Rauta, P.R.; Samanta, M.; Dash, H.R.; Nayak, B.; Das, S. Toll-like receptors (TLRs) in aquatic animals: Signaling pathways, expressions and immune responses. Immunol. Lett. 2014, 158, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Salamatin, R.; Jagodziński, P.P.; Wandurska-Nowak, E. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Exp. Parasitol. 2012, 130, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Czernomysy-Furowicz, D.; Lanocha, N.; Wandurska-Nowak, E.; Salamatin, R.; Jagodziński, P.P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Part II. Exp. Parasitol. 2013, 135, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Kroening, K.D.; Zimmerman, N.P.; Bass, P.; Oaks, J.A. Partial characterization of a tapeworm-secreted signal factor inducing sustained spike potentials in the smooth muscle of the rat small intestine. J. Parasitol. 2002, 88, 227–231. [Google Scholar] [CrossRef]
- Głobińska, A.; Kowalski, M.L. Natural immunological response to respiratory viruses—Intracellular signalling pathways. Alerg. Astma Immun. 2012, 17, 66–76. [Google Scholar]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Baranowska-Bosiacka, I.; Lanocha, N.; Wandurska-Nowak, E.; Gutowska, I.; Salamatin, R.; Jagodziński, P.P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes and protein (TLR3 and TLR9) in the small and large intestines of rats. Exp. Parasitol. 2014, 145, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Czerkies, M.; Kwiatkowska, K. Toll-like receptors and their contribution to innate immunity: Focus on TLR4 activation by lipopolysaccharide. Postepy Biol. Komorki 2013, 40, 39–64. [Google Scholar] [CrossRef]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapczuk, P.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Gutowska, I.; Kupnicka, P.; Chlubek, D.; Baranowska-Bosiacka, I. Selected Molecular Mechanisms Involved in the Parasite–Host System Hymenolepis diminuta–Rattus norvegicus. Int. J. Mol. Sci. 2018, 19, 2435. https://doi.org/10.3390/ijms19082435
Kapczuk P, Kosik-Bogacka D, Łanocha-Arendarczyk N, Gutowska I, Kupnicka P, Chlubek D, Baranowska-Bosiacka I. Selected Molecular Mechanisms Involved in the Parasite–Host System Hymenolepis diminuta–Rattus norvegicus. International Journal of Molecular Sciences. 2018; 19(8):2435. https://doi.org/10.3390/ijms19082435
Chicago/Turabian StyleKapczuk, Patrycja, Danuta Kosik-Bogacka, Natalia Łanocha-Arendarczyk, Izabela Gutowska, Patrycja Kupnicka, Dariusz Chlubek, and Irena Baranowska-Bosiacka. 2018. "Selected Molecular Mechanisms Involved in the Parasite–Host System Hymenolepis diminuta–Rattus norvegicus" International Journal of Molecular Sciences 19, no. 8: 2435. https://doi.org/10.3390/ijms19082435