Cell Cycle Regulation by Ca2+-Activated K+ (BK) Channels Modulators in SH-SY5Y Neuroblastoma Cells

 , and
, and
Abstract
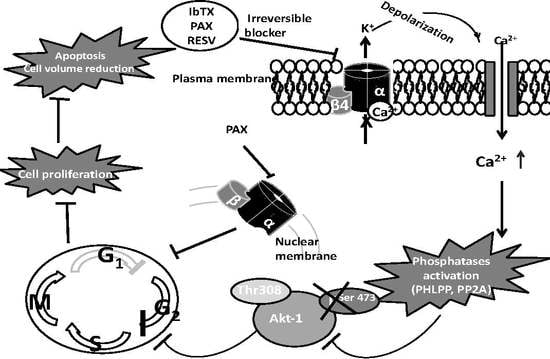
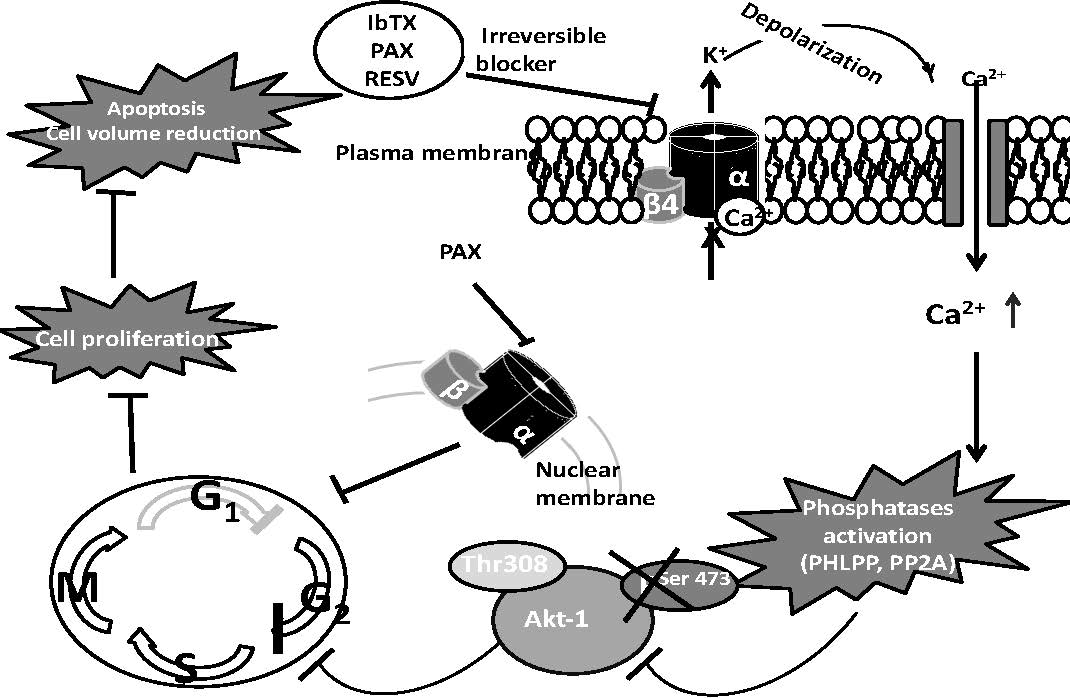
:
1. Introduction
2. Results
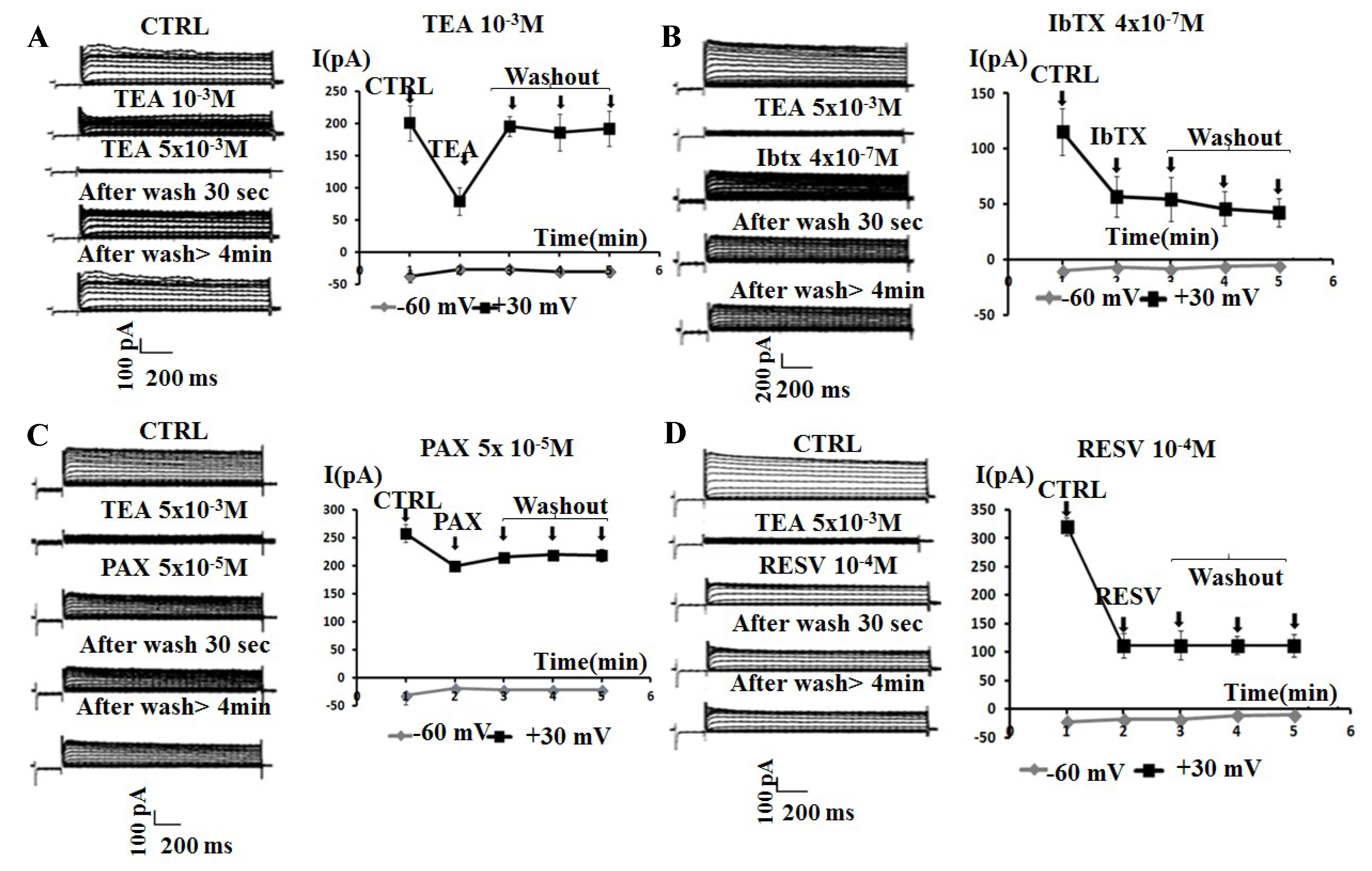
2.1. Effects of the Application of Kv/BK Channels Targeting Compounds on the Whole-Cell K+ Current of SH-SY5Y Cells
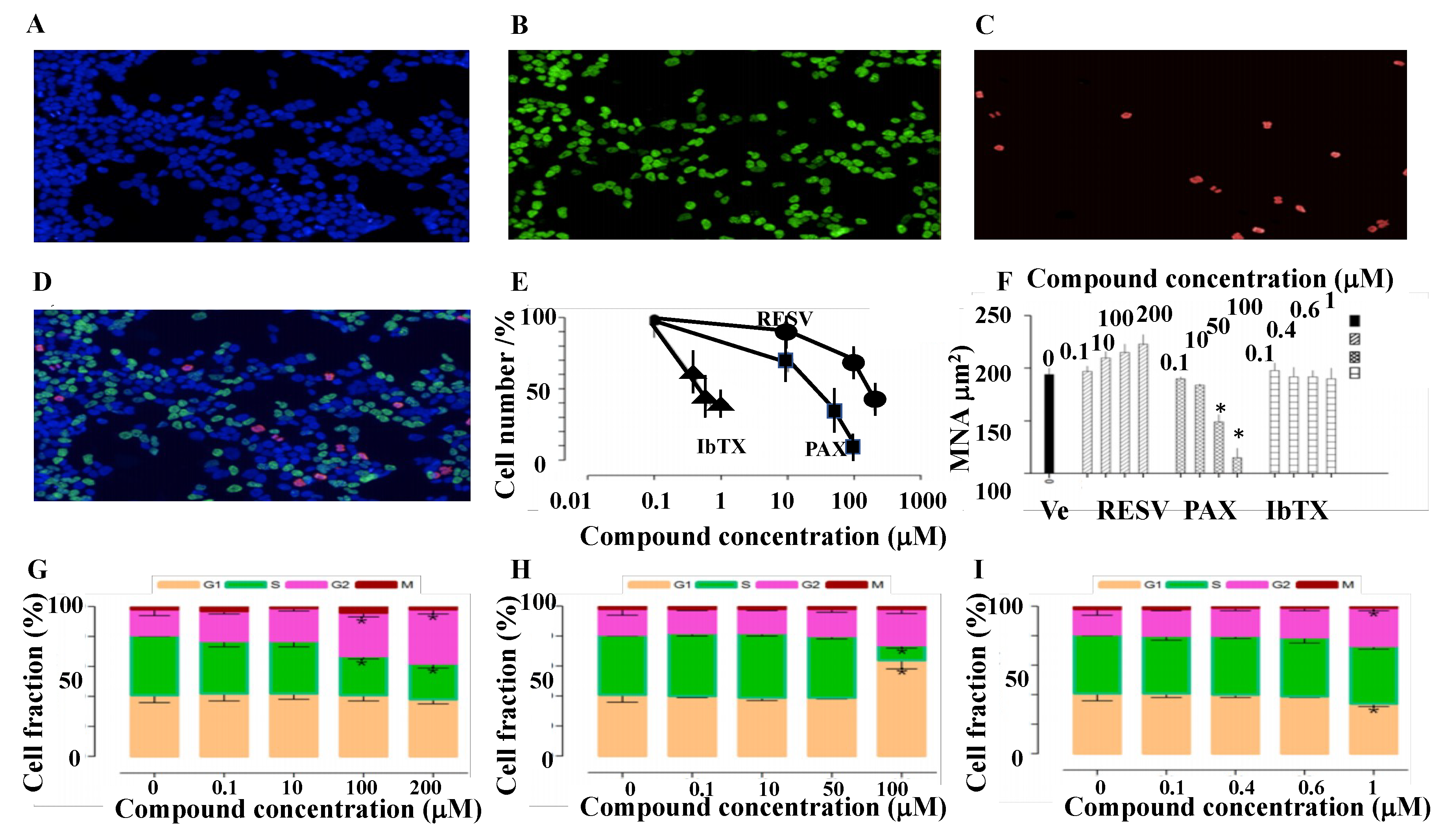
2.2. Cell Proliferation and Cell Cycle Progression in SH-SY5Y Cell Following 6 h Incubation of the Cells with Kv/BK Channel Targeting Drugs
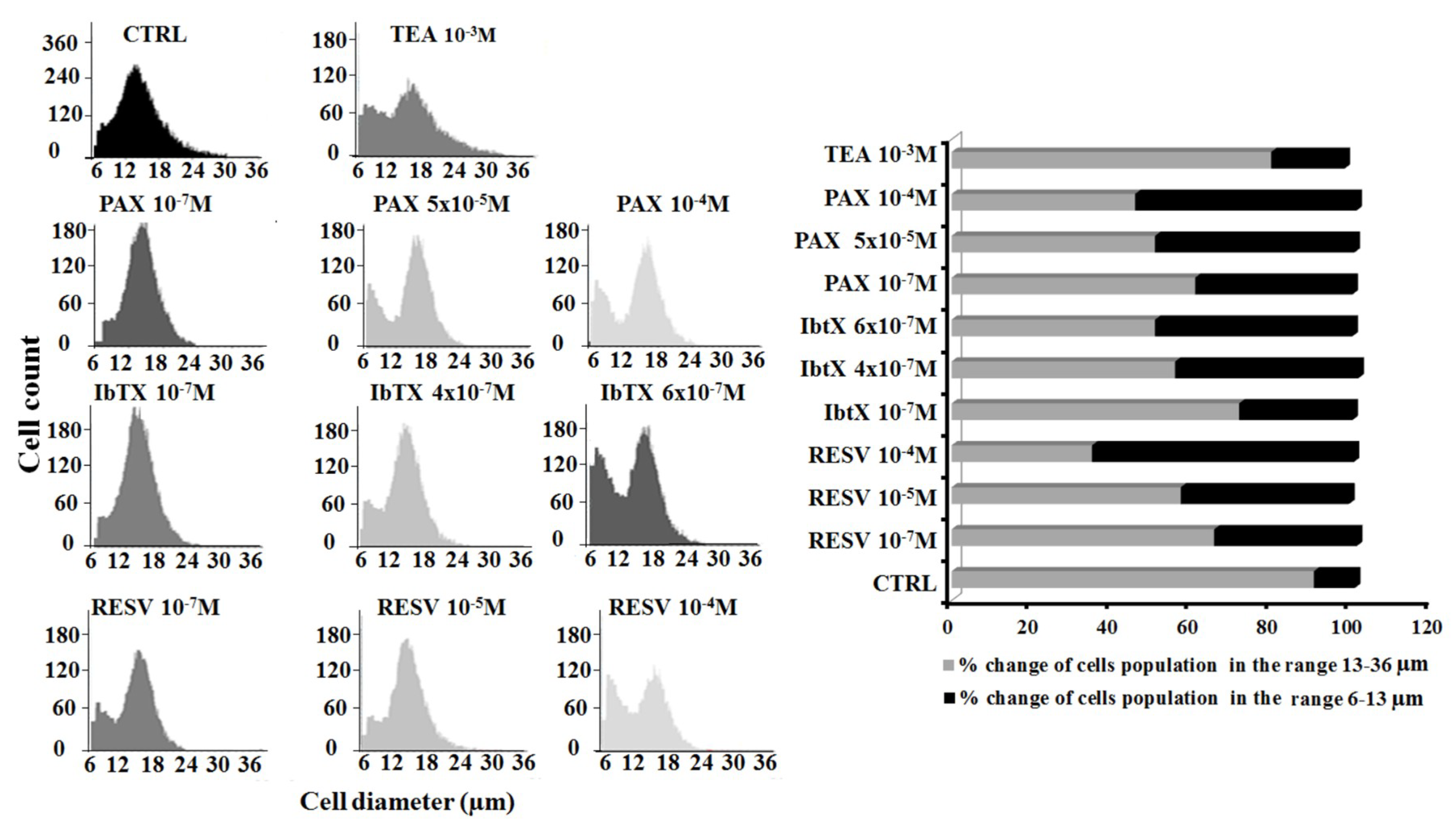
2.3. Cell Volume Changes Induced by Kv/BK Channels Targeting Compounds
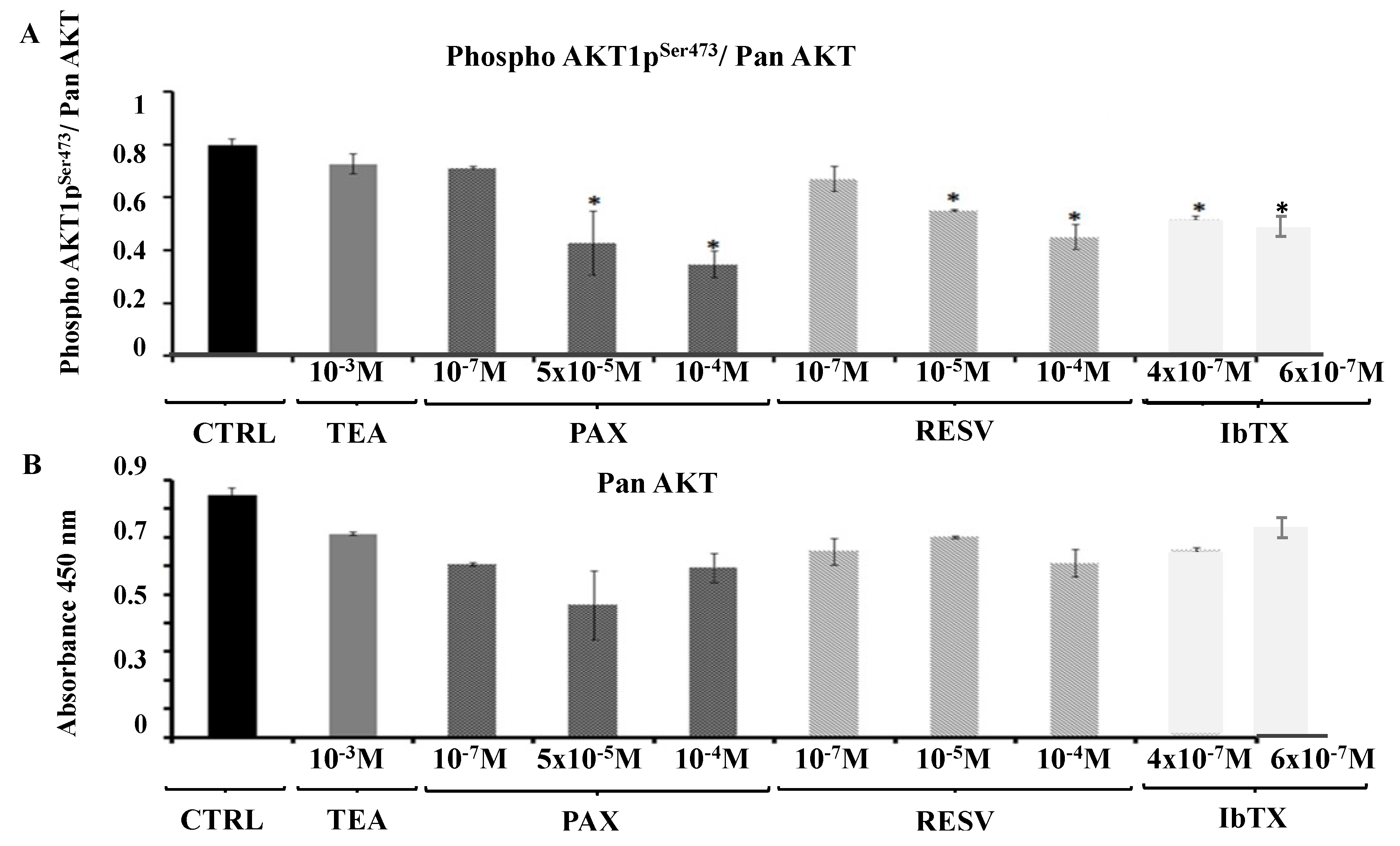
2.4. AKT1pser473a Putative Node in the Signaling Pathways Regulating Cell Proliferation in Response to the Modulation of the BK Channel Activity
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Patch Clamp Solutions
4.3. Whole-Cell K+ Current Recordings in the SH-SY5Y Cells
4.4. Cell Culture
4.5. High-Content Cell Cycle Analysis
4.6. Impedentiometric Cell Volume Assay
4.7. Phospo AKT (pSer473)/Pan AKT ELISA Assay
4.8. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roger, S.; Potier, M.; Vandier, C.; Le Guennec, J.Y.; Besson, P. Description and role in proliferation of iberiotoxin-sensitive currents in different human mammary epithelial normal and cancerous cells. Biochim. Biophys. Acta 2004, 1667, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeggerli, M.; Tian, Y.; Ruiz, C.; Wijker, B.; Sauter, G.; Obermann, E.; Guth, U.; Zlobec, I.; Sausbier, M.; Kunzelmann, K.; et al. Role of KCNMA1 in breast cancer. PLoS ONE 2012, 7, e41664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.Y.; Wright, C.M.; Kirschner, M.B.; Williams, M.; Sarun, K.H.; Sytnyk, V.; Leshchynska, I.; Edelman, J.J.; Vallely, M.P.; McCaughan, B.C.; et al. KCa1.1, a calcium-activated potassium channel subunit α 1, is targeted by miR-17-5p and modulates cell migration in malignant pleural mesothelioma. Mol. Cancer 2016, 15, 44. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Capriulo, R.; Conte Camerino, D. Involvement of KCa2+ channels in the local abnormalities and hyperkalemia following the ischemia-reperfusion injury of rat skeletal muscle. Neuromuscul. Disord. 2002, 12, 258–265. [Google Scholar] [CrossRef]
- Torres, Y.P.; Granados, S.T.; Latorre, R. Pharmacological consequences of the coexpression of BK channel alpha and auxiliary β subunits. Front. Physiol. 2014, 5, 383. [Google Scholar] [CrossRef] [PubMed]
- Imbrici, P.; Liantonio, A.; Camerino, G.M.; de Bellis, M.; Camerino, C.; Mele, A.; Giustino, A.; Pierno, S.; de Luca, A.; Tricarico, D.; et al. Therapeutic Approaches to Genetic Ion Channelopathies and Perspectives in Drug Discovery. Front. Pharmacol. 2016, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Dinardo, M.M.; Camerino, G.; Mele, A.; Latorre, R.; Conte Camerino, D.; Tricarico, D. Splicing of the rSlo gene affects the molecular composition and drug response of Ca2+-activated K+ channels in skeletal muscle. PLoS ONE 2012, 7, e40235. [Google Scholar] [CrossRef] [PubMed]
- Galvez, A.; Gimenez-Gallego, G.; Reuben, J.P.; Roy-Contancin, L.; Feigenbaum, P.; Kaczorowski, G.J.; Garcia, M.L. Purification and characterization of a unique, potent, peptidyl probe for the high conductance calcium-activated potassium channel from venom of the scorpion Buthus tamulus. J. Biol. Chem. 1990, 265, 11083–11090. [Google Scholar] [PubMed]
- Candia, S.; Garcia, M.L.; Latorre, R. Mode of action of iberiotoxin, a potent blocker of the large conductance Ca2+-activated K+ channel. Biophys. J. 1992, 63, 583–590. [Google Scholar] [CrossRef]
- MacKinnon, R.; Miller, C. Mechanism of charybdotoxin block of the high-conductance, Ca2+-activated K+ channel. J. Gen. Physiol. 1988, 91, 335–349. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, R.; Latorre, R.; Miller, C. Role of surface electrostatics in the operation of a high-conductance Ca2+-activated K+ channel. Biochemistry 1989, 28, 8092–8099. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, S.; Sun, P.; Pan, H.; Tian, C.; Zhang, L. Peptide toxins and small-molecule blockers of BK channels. Acta Pharmacol. Sin. 2016, 37, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaus, H.G.; McManus, O.B.; Lee, S.H.; Schmalhofer, W.A.; Garcia-Calvo, M.; Helms, L.M.; Sanchez, M.; Giangiacomo, K.; Reuben, J.P.; Smith, A.B. Tremorgenic indole alkaloids potently inhibit smooth muscle high-conductance calcium-activated potassium channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef] [PubMed]
- Imlach, W.L.; Finch, S.C.; Dunlop, J.; Dalziel, J.E. Structural determinants of lolitrems for inhibition of BK large conductance Ca2+-activated K+ channels. Eur. J. Pharmacol. 2009, 605, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lingle, C.J. Paxilline inhibits BK channels by an almost exclusively closed-channel block mechanism. J. Gen. Physiol. 2014, 144, 415–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, A.K.; Liu, X.; Sontheimer, H. Role for calcium-activated potassium channels (BK) in growth control of human malignant glioma cells. J. Neurosci. Res. 2004, 78, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatun, A.; Fujimoto, M.; Kito, H.; Niwa, S.; Suzuki, T.; Ohya, S. Down-Regulation of Ca2+-Activated K+ Channel KCa1.1 in Human Breast Cancer MDA-MB-453 Cells Treated with Vitamin D Receptor Agonists. Int. J. Mol. Sci. 2016, 17, 2083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Li, G.; Che, H.; Sun, H.Y.; Xiao, G.S.; Wang, Y.; Li, G.R. Effects of BKCa and Kir2.1 Channels on Cell Cycling Progression and Migration in Human Cardiac c-kit+ Progenitor Cells. PLoS ONE 2015, 10, e0138581. [Google Scholar] [CrossRef] [PubMed]
- Chittajallu, R.; Chen, Y.; Wang, H.; Yuan, X.; Ghiani, C.A.; Heckman, T.; McBain, C.J.; Gallo, V. Regulation of Kv1 subunit expression in oligodendrocyte progenitor cells and their role in G1/S phase progression of the cell cycle. Proc. Natl. Acad. Sci. USA 2002, 99, 2350–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, E.L.; Stuart, J.A. trans-Resveratrol as a neuroprotectant. Molecules 2010, 15, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Rieder, S.A.; Nagarkatti, P.; Nagarkatti, M. Multiple anti-inflammatory pathways triggered y resveratrol lead to amelioration of staphylococcal enterotoxin B-induced lung injury. Br. J. Pharmacol. 2012, 167, 1244–1258. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Chan, M.H.; Chen, L.; Wu, S.N.; Chen, H.H. Resveratrol attenuates cortical neuron activity: Roles of large conductance calcium-activated potassium channels and voltage-gated sodium channels. J. Biomed. Sci. 2016, 23, 47. [Google Scholar] [CrossRef] [PubMed]
- Curci, A.; Maqoud, F.; Mele, A.; Cetrone, M.; Angelelli, M.; Zizzo, N.; Tricarico, D. Antiproliferative effects of neuroprotective drugs targeting big Ca2+-activated K+ (BK) channel in the undifferentiated neuroblastoma cells. Curr. Top. Pharmacol. 2016, 20, 113–131. [Google Scholar]
- Carter, L.G.; D’Orazio, J.A.; Pearson, K.J. Resveratrol and cancer: Focus on in vivo evidence. Endocr. Relat. Cancer 2014, 21, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Athar, M.; Back, J.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Multiple molecular Targets of Resveratrol: Anti-carcinogenic Mechanisms. Arch. Biochem. Biophys. 2009, 486, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.M.; Hernandez, F.; Puerta, N.; de Angulo, G.; Webster, K.A.; Vanni, S. Resveratrol augments ER stress and the cytotoxic effects of glycolytic inhibition in neuroblastoma by downregulating AKT in a mechanism independent of SIRT1. Exp. Mol. Med. 2016, 48, e210-12. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolowski, B.; Orchard, S.; Harvey, M.; Sridhar, S.; Sakai, Y. Conserved BK Channel-Protein Interactions Reveal Signals Relevant to Cell Death and Survival. PLoS ONE 2011, 6, e28532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Park, S.J.; Chung, M.K.; Jung, K.H.; Choi, M.R.; Kim, Y.; Chai, Y.G.; Kim, S.J.; Park, K.S. High expression of large-conductance Ca2+-activated K+ channel in the CD133+ subpopulation of SH-SY5Y neuroblastoma cells. Biochem. Biophys. Res. Commun. 2010, 396, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Curci, A.; Mele, A.; Camerino, G.M.; Dinardo, M.M.; Tricarico, D. The large conductance Ca2+-activated K+ (BKCa) channel regulates cell proliferation in SH-SY5Y neuroblastoma cells by activating the staurosporine-sensitive protein kinases. Front. Physiol. 2014, 5, 476. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Brackenbury, W.J. Membrane potential and cancer progression. Front. Physiol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Giangiacomo, K.M.; Garcia, M.L.; McManus, O.B. Mechanism of iberiotoxin block of the large-conductance calcium-activated potassium channel from bovine aortic smooth muscle. Biochemistry 1992, 31, 6719–6727. [Google Scholar] [CrossRef] [PubMed]
- Meera, P.; Wallner, M.; Toro, L. A neuronal β subunit (KCNMB4) makes the large conductance, voltage- and Ca2+-activated K+ channel resistant to charybdotoxin and iberiotoxin. Proc. Natl. Acad. Sci. USA. 2000, 97, 5562–5567. [Google Scholar] [CrossRef] [PubMed]
- Li, H.F.; Chen, S.A.; Wu, S.N. Evidence for the stimulatory effect of resveratrol on Ca2+-activated K+ current in vascular endothelial cells. Cardiovasc. Res. 2000, 45, 1035–1045. [Google Scholar] [CrossRef]
- Tricarico, D.; Mele, A.; Conte Camerino, D. Carbonic anhydrase inhibitors ameliorate the symptoms of hypokalaemic periodic paralysis in rats by opening the muscular Ca2+-activated-K+channels. Neuromuscul. Disord. 2006, 16, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Lovaglio, S.; Mele, A.; Rotondo, G.; Mancinelli, E.; Meola, G.; Conte Camerino, D. Acetazolamide prevents vacuolar myopathy in skeletal muscle of K+-depleted rats. J Pharmacol 2008, 154, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Mele, A.; Liss, B.; Ashcroft, F.M.; Lundquist, A.L.; Desai, R.R.; George, A.L.J.; Conte Camerino, D. Reduced expression of Kir6.2/SUR2A subunits explains KATP deficiency in K+-depleted rats. Neuromuscul. Disord. 2008, 18, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Orsini, F.; Verotta, L.; Lecchi, M.; Restano, R.; Curia, G.; Redaelli, E.; Wanke, E. Resveratrol derivatives and their role as potassium channels modulators. J. Nat. Prod. 2004, 67, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.B.; Hu, G.Y. Trans-resveratrol, a red wine ingredient, inhibits voltage-activated potassium currents in rat hippocampal neurons. Brain Res. 2005, 1056, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Chi, T.C.; Chuang, L.M.; Su, M.J. Resveratrol enhances insulin secretion by blocking KATP and KV channels of β cells. Eur. J. Pharmacol. 2007, 568, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Li, M.; Hall, L.; Chen, S.; Sukur, S.; Lu, R.; Caputo, A.; Meredith, A.L.; Stefani, E.; Toro, L. MaxiK channel interactome reveals its interaction with GABA transporter 3 and heat shock protein 60 in the mammalian brain. Neuroscience 2016, 317, 76–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoose, S.A.; Duran, C.; Malik, I.; Eslamfam, S.; Shasserre, S.C.; Downing, S.S.; Hoover, E.M.; Dowd, K.E.; Smith, R.; Polymenis, M. Systematic analysis of cell cycle effects of common drugs leads to the discovery of a suppressive interaction between gemfibrozil and fluoxetine. PLoS ONE 2012, 7, e36503. [Google Scholar] [CrossRef] [PubMed]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stuhmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2014, 369, 20130094. [Google Scholar] [CrossRef] [PubMed]
- Dubois, J.M.; Rouzaire-Dubois, B. The influence of cell volume changes on tumour cell proliferation. Eur. Biophys. J. 2004, 33, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Dubois, J.M.; Rouzaire-Dubois, B. Roles of cell volume in molecular and cellular biology. Prog. Biophys. Mol. Biol. 2012, 108, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, J.I.; Segerstrom, L.; Orrego, A.; Elfman, L.; Henriksson, M.; Kagedal, B.; Eksborg, S.; Sveinbjornsson, B.; Kogner, P. Inhibitors of mammalian target of rapamycin downregulate MYCN protein expression and inhibit neuroblastoma growth in vitro and in vivo. Oncogene 2008, 27, 2910–2922. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.R.; Guo, S.; Minami, T.; Spokes, K.C.; Ueki, K.; Skurk, C.; Walsh, K.; Aird, W.C. Vascular endothelial growth factor activates PI3K/AKT/forkhead signaling in endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Ho, R.; Minturn, J.E.; Hishiki, T.; Zhao, H.; Wang, Q.; Cnaan, A.; Maris, J.; Evans, A.E.; Brodeur, G.M. Proliferation of human neuroblastomas mediated by the epidermal growth factor receptor. Cancer Res. 2005, 65, 9868–9875. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, H.J.; Xia, Y.Y.; Feng, Z.W. Insulin-like growth factor 1 protects human neuroblastoma cells SH-EP1 against MPP+-induced apoptosis by AKT/GSK-3β/JNK signaling. Apoptosis 2010, 15, 1470–1479. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Hong, S.I.; Kim, J.A.; Jung, Y.H.; Kim, S.Y.; Kim, H.C.; Lee, S.Y.; Jang, C.G. The neuroprotective effects of Lonicera japonica THUNB. against hydrogen peroxide-induced apoptosis via phosphorylation of MAPKs and PI3K/AKT in SH-SY5Y cells. Food Chem. Toxicol. 2011, 49, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Bayascas, J.R.; Alessi, D.R. Regulation of AKT/PKB Ser473 phosphorylation. Mol. Cell 2005, 18, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.P.; Liao, Y.; Xia, W.; Spohn, B.; Lee, M.H.; Hung, M.C. Cytoplasmic localization of p21Cip1/WAF1 by AKT-induced phosphorylation in HER-2/neu-overexpressing cells. Nat. Cell Biol. 2001, 3, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Viglietto, G.; Motti, M.L.; Bruni, P.; Melillo, R.M.; D’Alessio, A.; Califano, D.; Vinci, F.; Chiappetta, G.; Tsichlis, P.; Bellacosa, A.; et al. Cytoplasmic relocalization and inhibition of the cyclin-dependent kinase inhibitor p27(Kip1) by PKB/AKT-mediated phosphorylation in breast cancer. Nat. Med. 2002, 8, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.; Yakes, F.M.; Rojo, F.; Shin, N.Y.; Bakin, A.V.; Baselga, J.; Arteaga, C.L. PKB/AKT mediates cell-cycle progression by phosphorylation of p27(Kip1) at threonine 157 and modulation of its cellular localization. Nat. Med. 2002, 8, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zubovitz, J.; Petrocelli, T.; Kotchetkov, R.; Connor, M.K.; Han, K.; Lee, J.H.; Ciarallo, S.; Catzavelos, C.; Beniston, R.; et al. PKB/AKT phosphorylates p27, impairs nuclear import of p27 and opposes p27-mediated G1 arrest. Nat. Med. 2002, 8, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Niklasson, M.; Maddalo, G.; Sramkova, Z.; Mutlu, E.; Wee, S.; Sekyrova, P.; Schmidt, L.; Nicolas Fritz, N.; Dehnisch, I.; Kyriatzis, G.; et al. Membrane-Depolarizing Channel Blockers Induce Selective Glioma Cell Death by Impairing Nutrient Transport and Unfolded Protein/Amino Acid Responses. Cancer Res. 2017, 77, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Hung, M.C. Physiological regulation of AKT activity and stability. Am. J. Transl. Res. 2010, 2, 19–42. [Google Scholar] [PubMed]
- Duggal, S.; Jailkhani, N.; Midha, M.K.; Agrawal, N.; Rao, K.V.S.; Kumar, A. Defining the AKT1 interactome and its role in regulating the cell cycle. Sci. Rep. 2018, 8, 1303. [Google Scholar] [CrossRef] [PubMed]
- Mele, A.; Buttiglione, M.; Cannone, G.; Vitiello, F.; Conte Camerino, D.; Tricarico, D. Opening/blocking actions of pyruvate kinase antibodies on neuronal and muscular KATP channels. Pharmacol. Res. 2012, 66, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Mele, A.; Calzolaro, S.; Cannone, G.; Camerino, G.M.; Dinardo, M.M.; Latorre, R.; Conte Camerino, D. Emerging Role of Calcium-Activated Potassium Channel in the Regulation of Cell Viability Following Potassium Ions Challenge in HEK293 Cells and Pharmacological Modulation. PLoS ONE 2013, 8, e69551. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Barbieri, M.; Laghezza, A.; Tortorella, P.; Loiodice, F.; Conte Camerino, D. Dualistic actions of cromakalim and new potent 2H-1,4-benzoxazine derivatives on the native skeletal muscle KATP channel. Br. J. Pharmacol. 2003, 139, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Montanari, L.; Conte Camerino, D. Involvement of 3Na+/2K+ ATP-ase and Pi-3 kinase in the response of skeletal muscle ATP-sensitive K+ channels to insulin. Neuromuscul. Disord. 2003, 13, 712–719. [Google Scholar] [CrossRef]
- Massey, A.J. Multiparametric Cell Cycle Analysis Using the Operetta High-Content Imager and Harmony Software with PhenoLOGIC. PLoS ONE 2015, 10, e0134306. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, D.; Mele, A.; Camerino, G.M.; Bottinelli, R.; Brocca, L.; Frigeri, A.; Svelto, M.; George, A.L.J.; Conte Camerino, D. The KATP channel is a molecular sensor of atrophy in skeletal muscle. J. Physiol. 2010, 588, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Cetrone, M.; Mele, A.; Tricarico, D. Effects of the antidiabetic drugs on the age-related atrophy and sarcopenia associated with diabetes type II. Curr. Diabetes Rev. 2014, 10, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Mele, A.; Camerino, G.M.; Calzolaro, S.; Cannone, M.; Conte Camerino, D.; Tricarico, D. Dual response of the KATP channels to staurosporine: A novel role of SUR2B, SUR1 and Kir6.2 subunits in the regulation of the atrophy in different skeletal muscle phenotypes. Biochem. Pharmacol. 2014, 91, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Ernest, N.J.; Habela, C.W.; Sontheimer, H. Cytoplasmic condensation is both necessary and sufficient to induce apoptotic cell death. J. Cell Sci. 2008, 121, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs | Mechanism of BK Channel Modulation | Cell Cycle Phase | Nuclear/Cell Morphology | AKT1pser473 Phosporylation | Cell Proliferation |
|---|---|---|---|---|---|
| IbTX | Membrane impermeable; BK selective blocker; Open channel blocker; Not reversible action | G2 accumulation; G1 contraction; S not affected | No effects/ Diameter reduction | Dephosphorylation | Moderate reduction |
| PAX | Membrane permeable; BK selective blocker; Allosteric modulator and closed channel blocker; Not reversible action | G2 accumulation; S contraction; G1 accumulation | Nuclear area shrinking/ Diameter reduction | Marked dephosphorylation | Marked reduction |
| RESV | Membrane permeable; BK unselective modulator; Not reversible action | G2 accumulation; S contraction | Nuclear area enlargement/ Diameter reduction | Dephosphorylation | Moderate reduction |
| TEA | Kv/BK unselective blocker with reversible action | No effects | No Effects/No Effects | No effects | No effects |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maqoud, F.; Curci, A.; Scala, R.; Pannunzio, A.; Campanella, F.; Coluccia, M.; Passantino, G.; Zizzo, N.; Tricarico, D. Cell Cycle Regulation by Ca2+-Activated K+ (BK) Channels Modulators in SH-SY5Y Neuroblastoma Cells. Int. J. Mol. Sci. 2018, 19, 2442. https://doi.org/10.3390/ijms19082442
Maqoud F, Curci A, Scala R, Pannunzio A, Campanella F, Coluccia M, Passantino G, Zizzo N, Tricarico D. Cell Cycle Regulation by Ca2+-Activated K+ (BK) Channels Modulators in SH-SY5Y Neuroblastoma Cells. International Journal of Molecular Sciences. 2018; 19(8):2442. https://doi.org/10.3390/ijms19082442
Chicago/Turabian StyleMaqoud, Fatima, Angela Curci, Rosa Scala, Alessandra Pannunzio, Federica Campanella, Mauro Coluccia, Giuseppe Passantino, Nicola Zizzo, and Domenico Tricarico. 2018. "Cell Cycle Regulation by Ca2+-Activated K+ (BK) Channels Modulators in SH-SY5Y Neuroblastoma Cells" International Journal of Molecular Sciences 19, no. 8: 2442. https://doi.org/10.3390/ijms19082442