Effect of N- and C-Terminal Amino Acids on the Interfacial Binding Properties of Phospholipase D from Vibrio parahaemolyticus

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
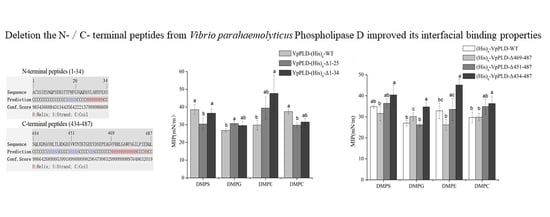
2.1. Analysis of the Primary and Secondary Structures of VpPLD-WT and Its N- and C-Terminal Sequences
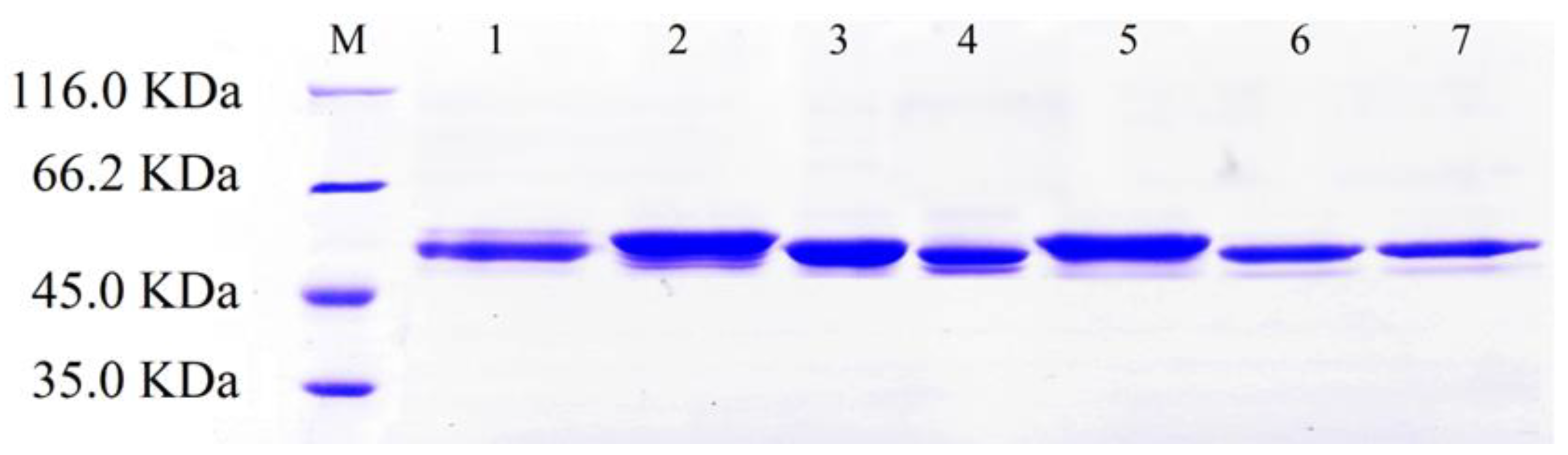
2.2. Mutants Construction and Purification
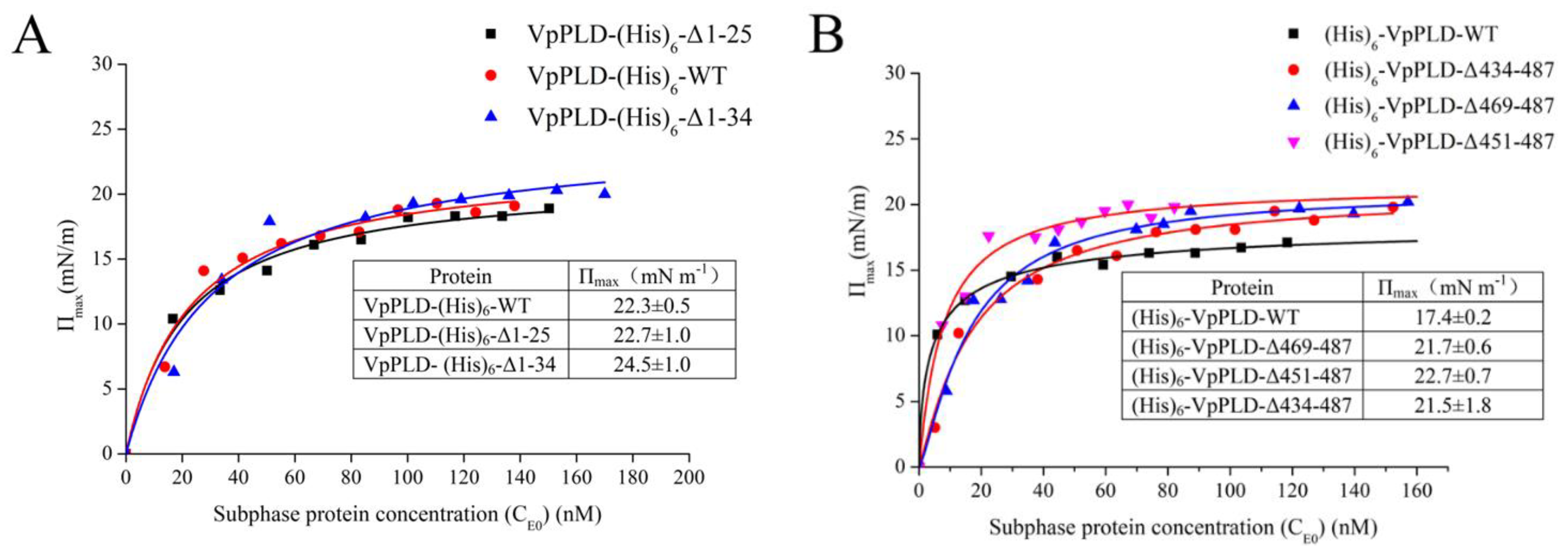
2.3. Binding of VpPLD-WT and Its N- and C-Terminal Truncated Mutants to an Air–Water Interface
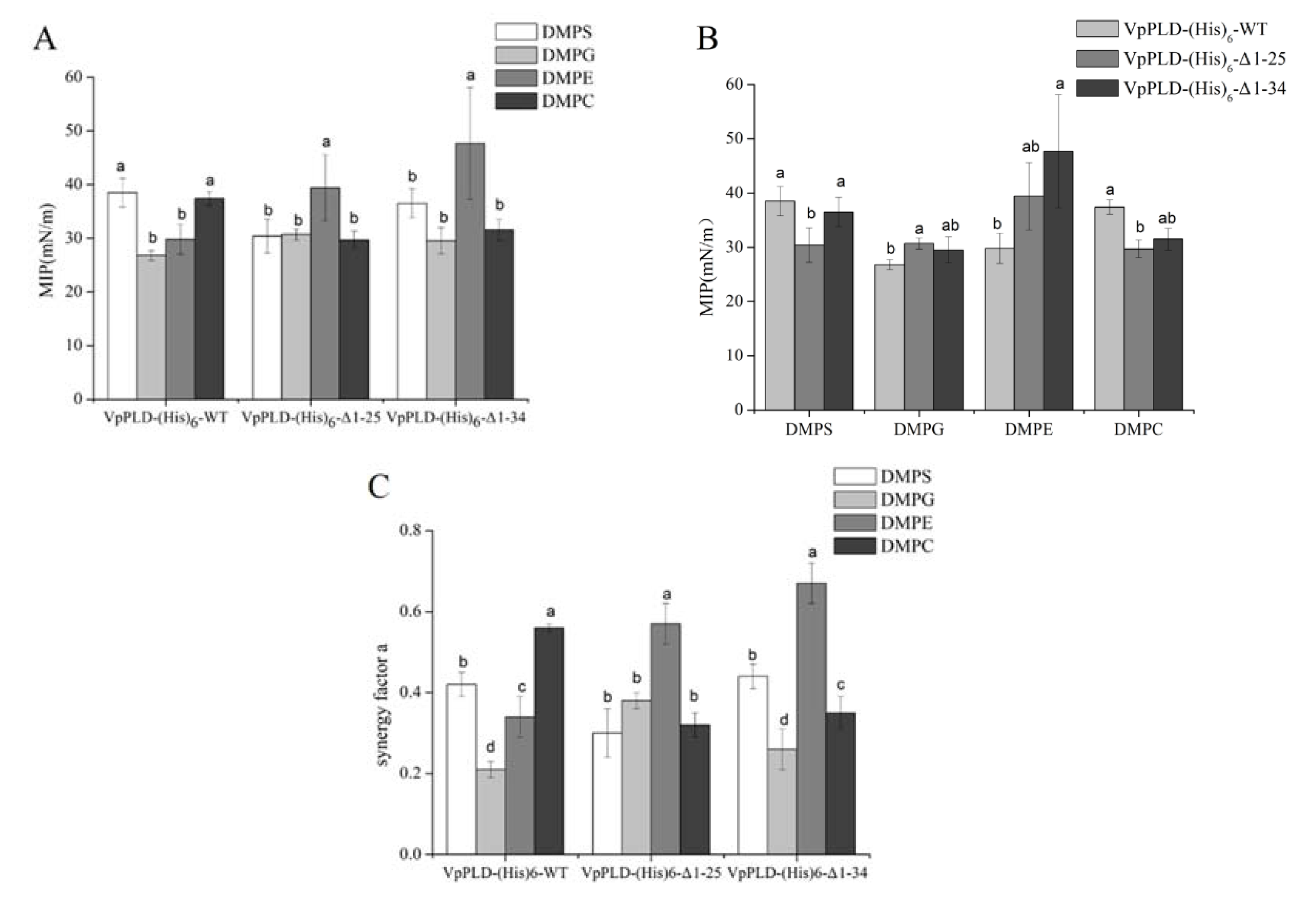
2.4. Binding of VpPLD-(His)6-WT and Its N-Terminal Truncated Mutants to Different Phospholipid Monolayers
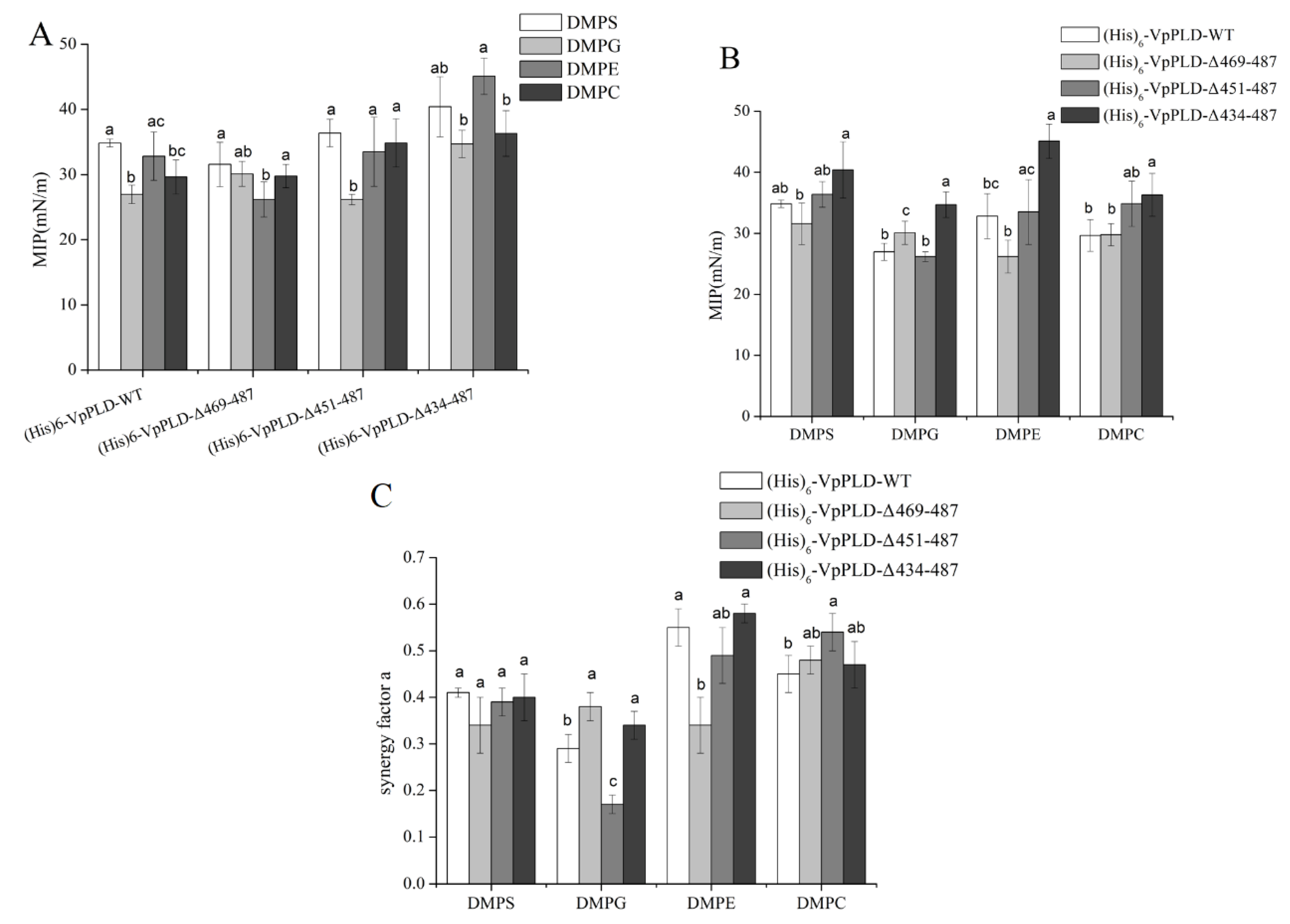
2.5. Extent of the Binding of (His)6-WT and Its C-Terminal Truncated Mutants to Different Phospholipid Monolayers
3. Materials and Methods
3.1. Materials
3.2. Total Gene Synthesis of VpPLD and Construction of the Expression Plasmid
3.3. Construction of Truncated Mutants of VpPLD
3.4. Recombinant Protein Expression and Protein Purification
3.5. Binding of VpPLD-WT and Truncated Mutants to an Air–Water Interface in the Absence of a Phospholipid Monolayer
3.6. Binding of the VpPLD-WT and Its Truncated Mutants to Various Phospholipid Monolayers
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mondal, S.; Khelashvili, G.; Weinstein, H. Not just an oil slick: How the energetics of protein-membrane interactions impacts the function and organization of transmembrane proteins. Biophys. J. 2014, 106, 2305–2316. [Google Scholar] [CrossRef] [PubMed]
- Laganowsky, A.; Reading, E.; Allison, T.M.; Ulmschneider, M.B.; Degiacomi, M.T.; Baldwin, A.J. Membrane proteins bind lipids selectively to modulate their structure and function. Nature 2014, 510, 172–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, T.J.; Arkin, I.T. Do more complex organisms have a greater proportion of membrane proteins in their genomes? Proteins 2000, 39, 417–420. [Google Scholar] [CrossRef]
- Mouritsen, O.G. Lipidology and lipidomics––Quo vadis? A new era for the physical chemistry of lipids. Phys. Chem. Chem. Phys. 2011, 13, 19195–19250. [Google Scholar] [CrossRef] [PubMed]
- Czogalla, A.; Grzybek, M.; Jones, W.; Coskun, U. Validity and applicability of membrane model systems for studying interactions of peripheral membrane proteins with lipids. Biochim. Biophys. Acta 2014, 8, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Brezesinski, G.; Mohwald, H. Langmuir monolayers to study interactions at model membrane surfaces. Adv. Colloid Interface Sci. 2003, 100–102, 563–584. [Google Scholar] [CrossRef]
- Brockman, H. Lipid monolayers: Why use half a membrane to characterize protein-membrane interactions? Curr. Opin. Struct. Biol. 1999, 9, 438–443. [Google Scholar] [CrossRef]
- Boucher, J.; Trudel, E.; Methot, M.; Desmeules, P.; Salesse, C. Organization, structure and activity of proteins in monolayers. Colloids Surf. B 2007, 58, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Maget-Dana, R. The monolayer technique: A potent tool for studying the interfacial properties of antimicrobial and membrane-lytic peptides and their interactions with lipid membranes. Biochim. Biophys. Acta 1999, 1462, 109–140. [Google Scholar] [CrossRef]
- Si-shen, F. Interpretation of mechanochemical properties of lipid bilayer vesicles from the equation of state or pressure–area measurement of the monolayer at the air–water or oil–water Interface. Langmuir 1999, 15, 998–1010. [Google Scholar]
- MacDonald, R.C.; Simon, S.A. Lipid monolayer states and their relationships to bilayers. Proc. Natl. Acad. Sci. USA 1987, 84, 4089–4093. [Google Scholar] [CrossRef] [PubMed]
- Lhor, M.; Bernier, S.C.; Horchani, H.; Bussières, S.; Cantin, L.; Desbat, B.; Salesse, C. Comparison between the behavior of different hydrophobic peptides allowing membrane anchoring of proteins. Adv. Colloid Interface Sci. 2014, 207, 223–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, M.C.; Sparks, C.E. Properties of apolipiproteins at the air–water interface. Ann. N. Y. Acad. Sci. 1980, 348, 122–137. [Google Scholar] [CrossRef]
- Calvez, P.; Demers, E.; Boisselier, E.; Salesse, C. Analysis of the contribution of saturated and polyunsaturated phospholipid monolayers to the binding of proteins. Langmuir 2011, 27, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Calvez, P.; Bussieres, S.; Demers, E.; Salesse, C. Parameters modulating the maximum insertion pressure of proteins and peptides in lipid monolayers. Biochimie 2009, 91, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Bénarouche, A.; Point, V.; Parsiegla, G.; Carrière, F.; Cavalier, J.F. New insights into the pH-dependent interfacial adsorption of dog gastric lipase using the monolayer technique. Colloids Surf. B Biointerfaces 2013, 111, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Exton, J.H. Phospholipase D structure, regulation and function. Rev. Physiol. Biochem. Pharmacol. 2002, 144, 1–94. [Google Scholar] [PubMed]
- Wang, X. Lipid signaling. Curr. Opin. Plant Biol. 2004, 7, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Kuppe, K.; Kerth, A.; Blume, A.; Ulbrich-Hofmann, R. Calcium-induced membrane microdomains trigger plant Phospholipase D activity. Chem. Biol. Chem. 2008, 9, 2853–2859. [Google Scholar] [CrossRef] [PubMed]
- Damnjanović, J.; Iwasaki, Y. Phospholipase D as a catalyst: Application in phospholipid synthesis, molecular structure and protein engineering. J. Biosci. Bioeng. 2013, 116, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Chahinian, H.; Bezzine, S.; Ferrato, F.; Ivanova, M.; Perez, B.; Lowe, M.; Carrière, F. Theβ5′ loop of the Pancreatic Lipase C2-like domain plays a critical role in the lipase-lipid interactions. Biochemistry 2002, 41, 13725–13735. [Google Scholar] [CrossRef] [PubMed]
- Bussieres, S.; Cantin, L.; Desbat, B.; Salesse, C. Binding of a truncated form of Lecithin: Retinol Acyltransferase and its N- and C-terminal peptides to lipid monolayers. Langmuir 2012, 28, 3516–3523. [Google Scholar] [CrossRef] [PubMed]
- Sayari, A.; Frikha, F.; Miled, N. N-terminal peptide of Rhizopus oryzae lipase is important for its catalytic properties. FEBS Lett. 2005, 579, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Rahier, R.; Noiriel, A.; Abousalham, A. Functional Characterization of the N-Terminal C2 Domain from Arabidopsis thaliana Phospholipase Dα and Dβ. BioMed Res. Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Horchani, H.; Fendri, A.; Louati, H.; Sayari, A.; Gargouri, Y.; Verger, R. Purification, biochemical and kinetic properties of recombinant Staphylococcus aureus lipase. Methods Mol. Biol. 2012, 861, 267–282. [Google Scholar] [PubMed]
- Sayari, A.; Mosbah, H.; Verger, R.; Gargouri, Y. The N-terminal His-tag affects the enantio selectivity of staphylococcal lipases: A monolayer study. J. Colloid Interface Sci. 2007, 1, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. Signal P 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 60, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- I-TASSER Server. Available online: http://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 12 April 2018).
- Proteinprediction. Available online: https://http://www.predictprotein.org (accessed on 8 April 2018).
- SSPro. Available online: http://scratch.proteomics.ics.uci.edu (accessed on 10 March 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids Number | Amino Acid Sequence 1 | Proportion of Amino Acids | ||
|---|---|---|---|---|
| Hydrophobic | Polar | Charged | ||
| 1–25 | ACSSIESNQPSEKSTTFHFGYQQDS | 24% (6/25) | 60% (15/25) | 16% (4/25) |
| 26–34 | VLAHYFEAY | 56% (5/9) | 22% (2/9) | 22% (2/9) |
| 434–450 | SQLKDNAYRLTLRDGDI | 35% (6/17) | 30% (5/17) | 35% (6/17) |
| 451–468 | VWTNTKTGEEYDSEPEAG | 33% (6/18) | 33% (6/18) | 33% (6/18) |
| 469–487 | VFRRLGAWFSGILPIEDQL | 68% (13/19) | 11% (2/19) | 21% (4/19) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Wei, R.; Abousalham, A.; Chen, W.; Yang, B.; Wang, Y. Effect of N- and C-Terminal Amino Acids on the Interfacial Binding Properties of Phospholipase D from Vibrio parahaemolyticus. Int. J. Mol. Sci. 2018, 19, 2447. https://doi.org/10.3390/ijms19082447
Wang F, Wei R, Abousalham A, Chen W, Yang B, Wang Y. Effect of N- and C-Terminal Amino Acids on the Interfacial Binding Properties of Phospholipase D from Vibrio parahaemolyticus. International Journal of Molecular Sciences. 2018; 19(8):2447. https://doi.org/10.3390/ijms19082447
Chicago/Turabian StyleWang, Fanghua, Ruixia Wei, Abdelkarim Abousalham, Wuchong Chen, Bo Yang, and Yonghua Wang. 2018. "Effect of N- and C-Terminal Amino Acids on the Interfacial Binding Properties of Phospholipase D from Vibrio parahaemolyticus" International Journal of Molecular Sciences 19, no. 8: 2447. https://doi.org/10.3390/ijms19082447