Implication of Membrane Androgen Receptor (ZIP9) in Cell Senescence in Regressed Testes of the Bank Vole

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
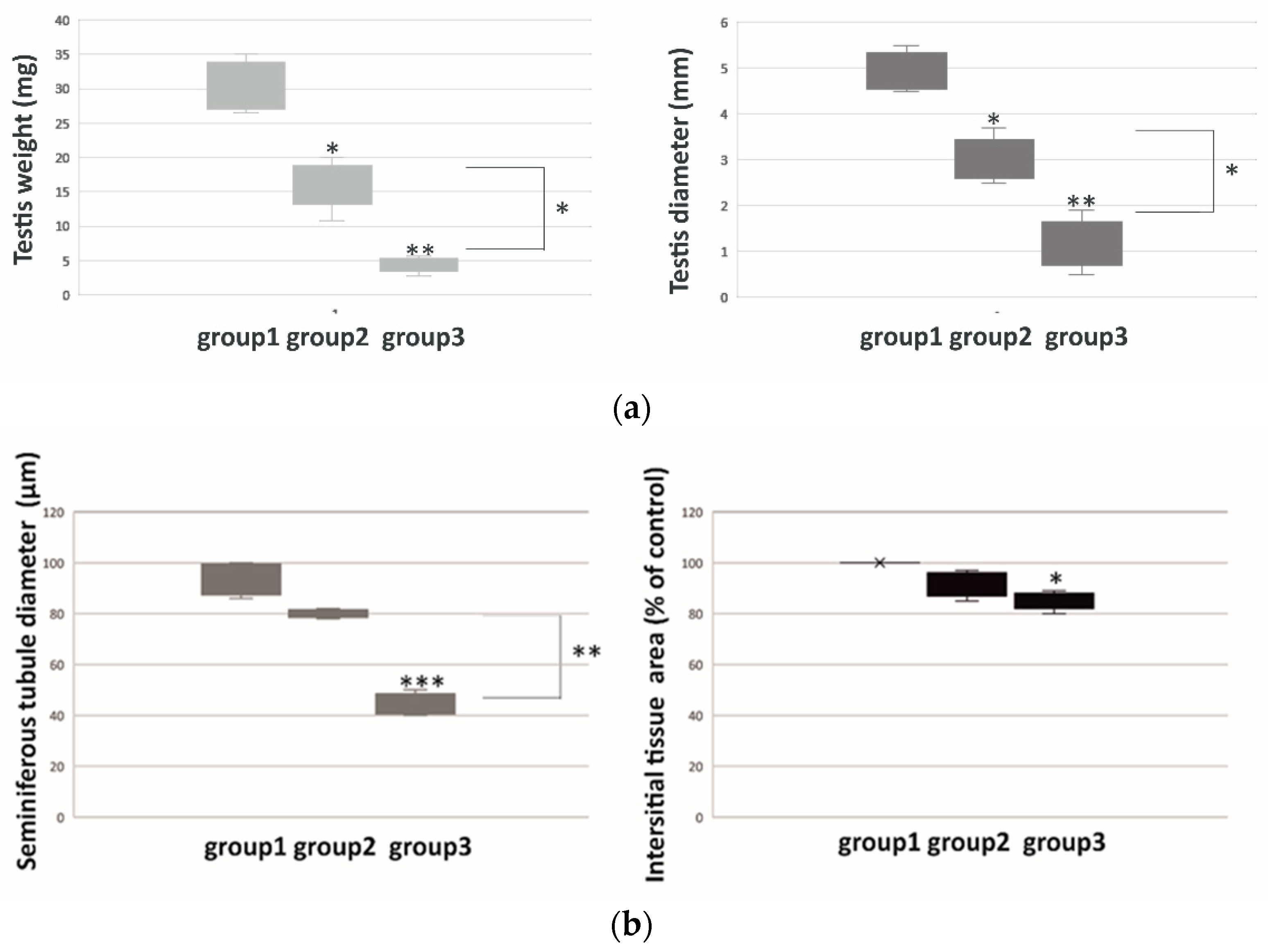
2.1. Weight, Diameter, Morphometry and Morphology of Bank Vole Regressed Testes
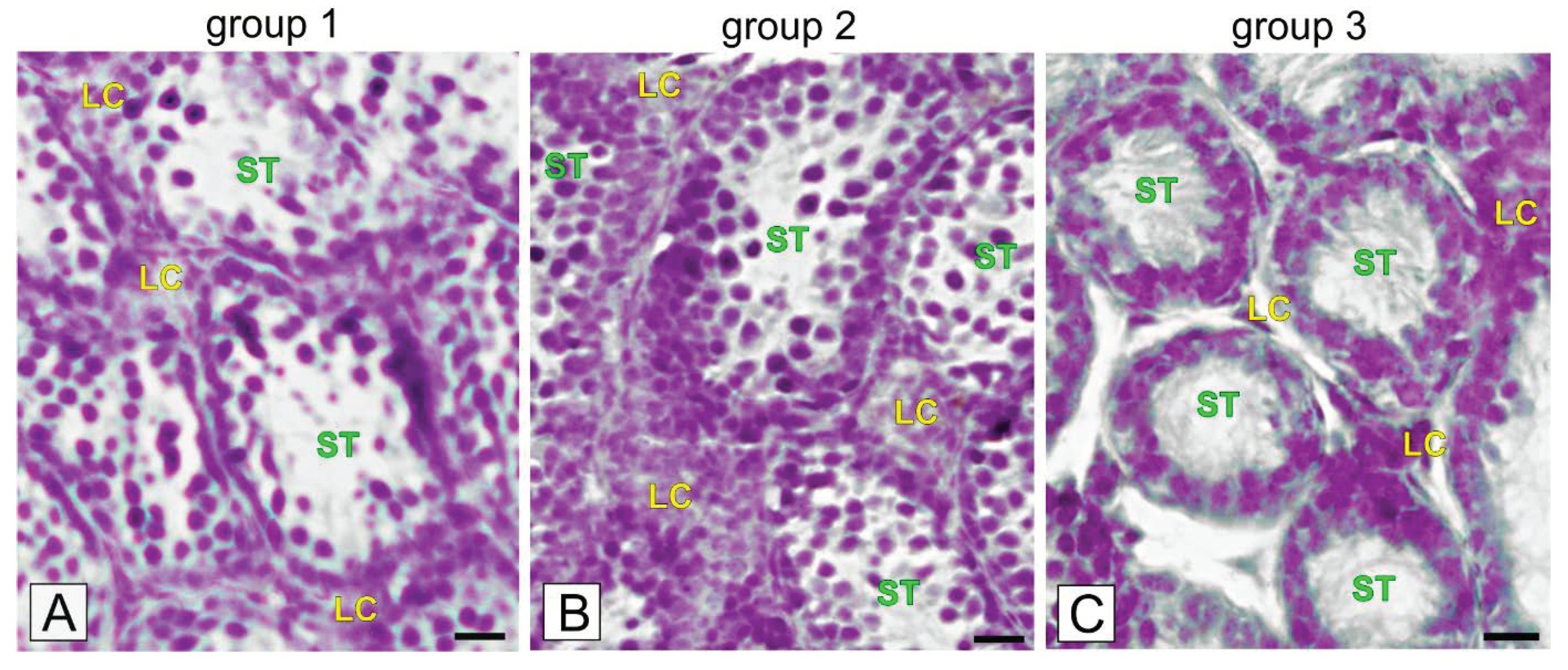
2.2. Localization of p16 and ZIP9 Receptor in Bank Vole Regressed Testes
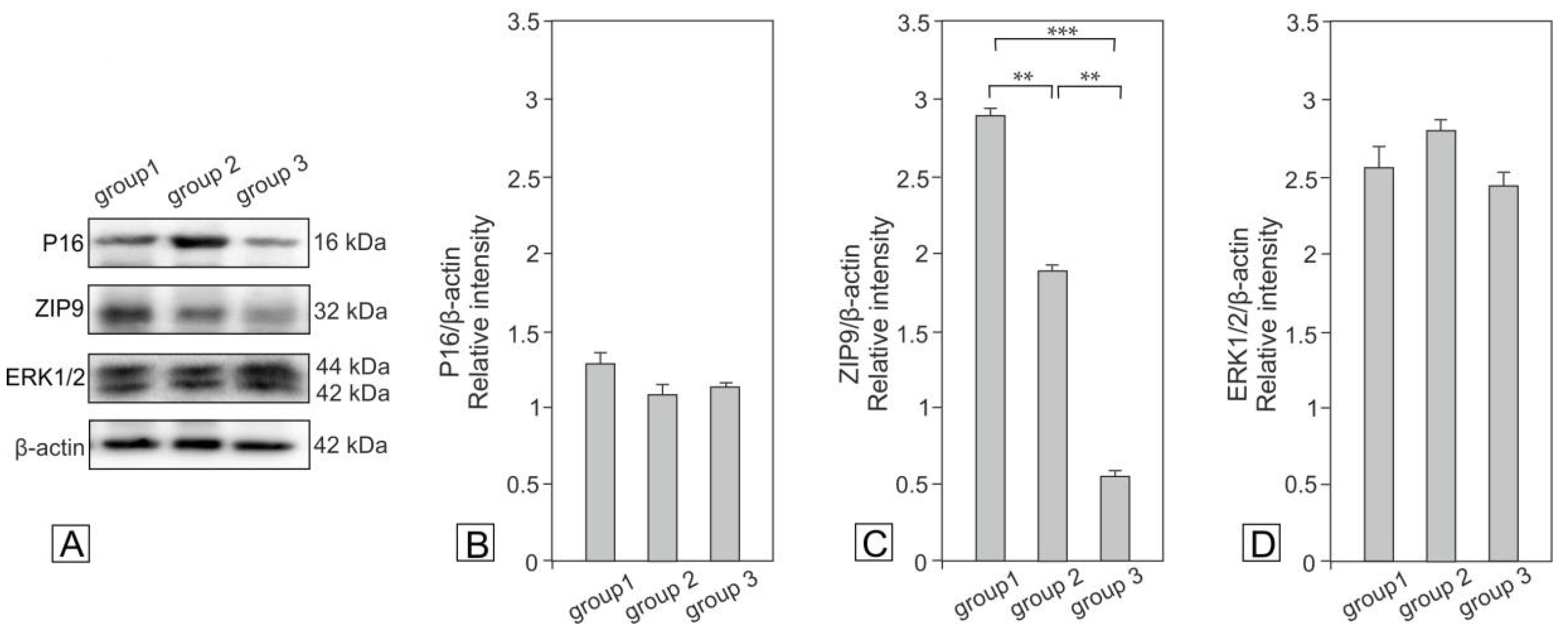
2.3. Expression of p16, ZIP9 and ERK1/2 in Bank Vole Regressed Testes
2.4. Concentration of cAMP and Testosterone in Bank Vole Regressed Testes
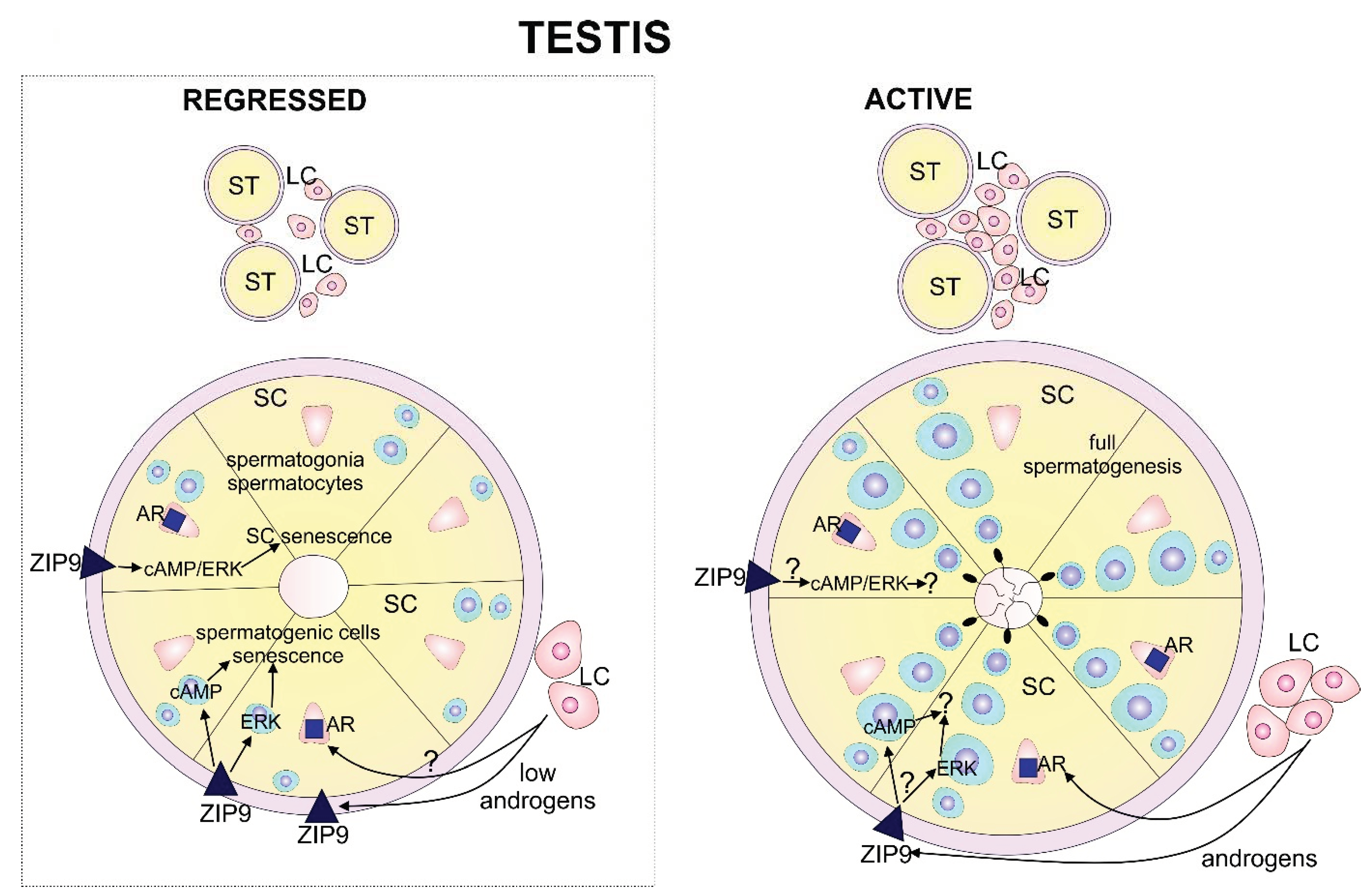
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Morphometry
4.3. Western Blot Analysis
4.4. Immunohistochemistry
4.5. Measurement of cAMP Concentration
4.6. Measurement of Testosterone Level
4.7. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bex, F.J.; Bartke, A. Testicular LH binding in the hamster: Modification by photoperiod and prolactin. Endocrinology 1977, 100, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Welch, D.; Gordon, I.J. The influence of vegetation pattern on the grazing of heather moorland by red deer and sheep. II. The impact on heather. J. Appl. Ecol. 1995, 32, 177–186. [Google Scholar] [CrossRef]
- Bilinska, B.; Schmalz-Fraczek, B.; Kotula, M.; Carreau, S. Photoperiod-dependent Capability of Androgen Aromatization and the Role of Estrogens in the Bank Vole Testis Visualized by Means of Immunohistochemistry. Mol. Cell Endocrinol. 2001, 178, 189–198. [Google Scholar] [CrossRef]
- Mano, T.; Tsubota, T. Reproductive Characteristics of Brown Bears on the Oshima Peninsula, Hokkaido, Japan. J. Mammal. 2002, 83, 026–1034. [Google Scholar] [CrossRef] [Green Version]
- Dadhich, R.K.; Barrionuevo, F.J.; Real, F.M.; Lupiañez, D.G.; Ortega, E.; Burgos, M.; Jiménez, R. Identification of live germ-cell desquamation as a major mechanism of seasonal testis regression in mammals: A study in the Iberian mole (Talpa occidentalis). Biol. Reprod. 2013, 18, 101. [Google Scholar] [CrossRef] [PubMed]
- Luaces, J.P.; Rossi, L.F.; Sciurano, R.B.; Rebuzzini, P.; Merico, V.; Zuccotti, M.; Merani, M.S.; Garagna, S. Loss of Sertoli-germ cell adhesion determines the rapid germ cell elimination during the seasonal regression of the seminiferous epithelium of the large hairy armadillo Chaetophractus villosus. Biol. Reprod. 2014, 6, 48. [Google Scholar] [CrossRef]
- Jiménez, R.; Burgos, M.; Barrionuevo, F.J. Circannual Testis Changes in Seasonally Breeding Mammals. Sex. Dev. 2015, 9, 205–215. [Google Scholar] [CrossRef]
- Speakman, J.R. The physiological costs of reproduction in small mammals. Philos. Trans. R Soc. Lond. B Biol. Sci. 2008, 27, 375–398. [Google Scholar] [CrossRef] [Green Version]
- Klemme, I.; Eccard, J.A.; Ylönen, H. Do Female Bank Voles (Clethrionomys glareolus) Mate Multiply to Improve on Previous Mates? Behav. Ecol. Sociobiol. 2006, 60, 415–421. [Google Scholar] [CrossRef]
- Torre, I.; Arrizabalaga, A. Habitat preferences of the bank vole Myodes glareolus in a Mediterranean mountain range. Acta Theriol. 2008, 53, 241–250. [Google Scholar] [CrossRef]
- Dadhich, R.K.; Real, F.M.; Zurita, F.; Barrionuevo, F.J.; Burgos, M.; Jiménez, R. Role of Apoptosis and Cell Proliferation in the Testicular Dynamics of Seasonal Breeding Mammals: A Study in the Iberian Mole, Talpa occidentalis. Biol. Reprod. 2010, 83, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Neaves, W.B. Changes in testicular leydig cells and in plasma testosterone levels among seasonally breeding rock hyrax. Biol. Reprod. 1973, 4, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha Hikim, A.P.; Bartke, A.; Russell, L.D. Morphometric studies on hamster testes in gonadally active and inactive states: Light microscope findings. Biol. Reprod. 1988, 39, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayed, A.E.; Hifny, A.; Abou-Elmagd, A.; Wrobel, K.H. Seasonal changes in the intertubular tissue of the camel testis (Camelus dromedarius). Ann. Anat. 1995, 177, 199–212. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Serrano, M. Cellular senescence: From physiology to pathology. Nat. Rev. Mol. Cell Biol. 2014, 15, 482–496. [Google Scholar] [CrossRef]
- Seco-Rovira, V.; Beltrán-Frutos, E.; Ferrer, C.; Saez, F.J.; Madrid, J.F.; Canteras, M.; Pastor, L.M. Testicular histomorphometry and the proliferative and apoptotic activities of the seminiferous epithelium in Syrian hamster (Mesocricetus auratus) during regression owing to short photoperiod. Andrology 2015, 3, 598–610. [Google Scholar] [CrossRef]
- Blottner, S.; Schön, J.; Roelants, H. Apoptosis is not the cause of seasonal testicular involution in roe deer. Cell Tissue Res. 2007, 327, 615–624. [Google Scholar] [CrossRef]
- Dadhich, R.K.; Barrionuevo, F.J.; Lupiañez, D.G.; Real, F.M.; Burgos, M.; Jiménez, R. Expression of genes controlling testicular development in adult testis of the seasonally breeding Iberian mole. Sex. Dev. 2011, 5, 77–88. [Google Scholar] [CrossRef]
- Ellis, G.B.; Turek, F.W. Time course of the photoperiod-induced change in sensitivity of the hypothalamic-pituitary axis to testosterone feedback in castrated male hamsters. Endocrinology 1979, 104, 625–630. [Google Scholar] [CrossRef]
- Meyer, S.L.; Goodman, R.L. Separate neural systems mediate the steroid-dependent and steroid-independent suppression of tonic luteinizing hormone secretion in the anestrous ewe. Biol. Reprod. 1986, 35, 562–571. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Schosserer, M.; Grillari, J.; Wolfrum, C.; Scheideler, M. Age-Induced Changes in White, Brite, and Brown Adipose Depots: A Mini-Review. Gerontology 2018, 64, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Bulldan, A.; Dietze, R.; Shihan, M.; Scheiner-Bobis, G. Non-classical testosterone signaling mediated through ZIP9 stimulates claudin expression and tight junction formation in Sertoli cells. Cell Signal. 2016, 28, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, L.; Smith, L.B. Androgen receptor roles in spermatogenesis and infertility. Best Practice Res. Clin. Endocrinol. Metab. 2015, 29, 595–605. [Google Scholar]
- Bilińska, B.; Słomczyńska, M.; Kmicikiewicz, I. Immunocytochemical demonstration of androgen receptors in Leydig cells of the bank vole (Clethrionomys glareolus, Schreber): An in vitro study. Acta Histochem. 1996, 98, 157–164. [Google Scholar] [CrossRef]
- Gancarczyk, M.; Paziewska-Hejmej, A.; Carreau, S.; Tabarowski, Z.; Bilinska, B. Dose- and photoperiod-dependent effects of 17beta-estradiol and the anti-estrogen ICI 182,780 on testicular structure, acceleration of spermatogenesis, and aromatase immunoexpression in immature bank voles. Acta Histochem. 2004, 106, 269–278. [Google Scholar] [CrossRef]
- Hejmej, A.; Kotula-Balak, M.; Galas, J.; Bilińska, B. Effects of 4-tert-octylphenol on the testes and seminal vesicles in adult male bank voles. Reprod Toxicol. 2011, 31, 95–105. [Google Scholar] [CrossRef]
- Kotula-Balak, M.; Hejmej, A.; Lydka, M.; Cierpich, A.; Bilinska, B. Detection of aromatase, androgen, and estrogen receptors in bank vole spermatozoa. Theriogenology 2012, 15, 385–392. [Google Scholar] [CrossRef]
- Pawlicki, P.; Milon, A.; Zarzycka, M.; Galas, J.; Tworzydlo, W.; Kaminska, A.; Pardyak, L.; Lesniak, K.; Pacwa, A.; Bilinska, B.; et al. Does signaling of estrogen-related receptors affect structure and function of bank vole Leydig cells? J. Physiol. Pharmacol. 2017, 68, 459–476. [Google Scholar]
- Milon, A.; Knapczyk-Stwora, K.; Pawlicki, P.; Duliban, M.; Gorowska-Wojtowicz, E.; Kotula-Balak, M.; Bilinska, B. Effect of estrogen-related receptor silencing on miRNA protein machinery expression, global methylation, and deacetylation in bank vole (Myodes glareolus) and mouse tumor Leydig cells. Theriogenology 2019, 139, 178–190. [Google Scholar] [CrossRef]
- Milon, A.; Pawlicki, P.; Rak, A.; Mlyczynska, E.; Płachno, B.J.; Tworzydlo, W.; Gorowska-Wojtowicz, E.; Bilinska, B.; Kotula-Balak, M. Telocytes are localized to testis of the bank vole (Myodes glareolus) and are affected by lighting conditions and G-coupled membrane estrogen receptor (GPER) signaling. Gen. Comp. Endocrinol. 2019, 15, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A.; Klemcke HGAmador, A.; Van Sickle, M. Photoperiod and regulation of gonadotropin receptors. Ann. N. Y. Acad. Sci. 1982, 383, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Gravis, C.J. Testicular Involution Following Optic Enucleation: The Leydig Cell. Cell Tissue Res. 1977, 19, 303–313. [Google Scholar] [CrossRef]
- Desjardins, C.; Ewing, L.L.; Johnson, B.H. Effects of light deprivation upon the spermatogenic and steroidogenic elements of hamster testes. Endocrinology 1971, 89, 791–800. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. The Physiological Function of Melatonin in Plants. Plant. Signal. Behav. 2006, 1, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Kruczek, M.; Styrna, J. Semen Quantity and Quality Correlate With Bank Vole Males’ Social Status. Behav. Processes 2009, 82, 279–285. [Google Scholar] [CrossRef]
- Chen, H.J.; Reiter, R.J. The combination of twice daily luteinizing hormone-releasing factor administration and renal pituitary homografts restores normal reproductive organ size in male hamsters with pineal-mediated gonadal atrophy. Endocrinology 1980, 106, 1382–1385. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A.; Goldman, B.D.; Bex, F.J.; Kelch, R.P.; Smith, M.S.; Dalterio, S.; Doherty, P.C. Effects of prolactin on testicular regression and recrudescence in the golden hamster. Endocrinology 1980, 106, 167–172. [Google Scholar] [CrossRef]
- Muñoz, E.; Fogal, T.; Dominguez, S.; Scardapane, L.; Guzmán, J.; Piezzi, R.S. Seasonal changes of the Leydig cells of viscacha (Lagostomus maximus maximus). A light and electron microscopy study. Tissue Cell 1997, 29, 119–128. [Google Scholar] [CrossRef]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell Actions 2016, 6, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Li, G.; Liu, L.; Feng, L.; Wang, X.; Jin, H. Regulation and function of mitophagy in development and cancer. Autophagy 2013, 1, 1720–1736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, L.; Yun, Q.; Hong, X. Mitochondrial cAMP signaling. Cell Mol. Life Sci. 2016, 73, 4577–4590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holstein, A.F.; Eckmann, C. Multinucleated spermatocytes and spermatids in human seminiferous tubules. Andrologia 1986, 8, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Contreras, H.; Ronco, A.M. Leydig Cell Heterogeneity as Judged by Quantitative Cytochemistry of 3 Beta-Hydroxysteroid Dehydrogenase Activity in Individual Rat Leydig Cells. J. Steroid. Biochem. Mol. Biol. 1994, 51, 73–79. [Google Scholar] [CrossRef]
- Yokoyama, C.; Chigi, Y.; Baba, T.; Ohshitanai, A.; Harada, Y.; Takahashi, F.; Morohashi, K.-I. Three populations of adult Leydig cells in mouse testes revealed by a novel mouse HSD3B1-specific rat monoclonal antibody. Biochem. Biophys. Res. Commun. 2019, 511, 916–920. [Google Scholar] [CrossRef]
- Akingbemi, B.T. Estrogen regulation of testicular function. Reprod. Biol. Endocrinol. 2005, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Gould, M.L.; Hurst, P.R.; Nicholson, H.D. The effects of oestrogen receptors alpha and beta on testicular cell number and steroidogenesis in mice. Reproduction 2007, 134, 271–279. [Google Scholar] [CrossRef]
- Verhoeven, G.; Willems, A.; Denolet, E.; Swinnen, J.V.; De Gendt, K. Androgens and spermatogenesis: Lessons from transgenic mouse models. Philos Trans. R Soc. Lond. B Biol. Sci. 2010, 27, 1537–1556. [Google Scholar] [CrossRef] [Green Version]
- Bhanmeechao, C.; Srisuwatanasagul, S.; Ponglowhapan, S. Age-related changes in interstitial fibrosis and germ cell degeneration of the canine testis. Reprod. Domest. Anim. 2018, 53, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Kotula-Balak, M.; Hejmej, A.; Kopera, I.; Lydka, M.; Bilinska, B. Prenatal and neonatal exposure to flutamide affects function of Leydig cells in adult boar. Domest. Anim. Endocrinol. 2012, 42, 142–145. [Google Scholar] [CrossRef]
- Sterbis, J.; E-Nunu, T. Leydig Cell Hyperplasia in the Setting of Klinefelter Syndrome. BMJ Case Rep. 2015, 2015, bcr2015209805. [Google Scholar] [CrossRef] [PubMed]
- Kamińska, A.; Pardyak, L.; Marek, S.; Wróbel, K.; Kotula-Balak, M.; Bilińska, B.; Hejmej, A. Notch signaling regulates nuclear androgen receptor AR and membrane androgen receptor ZIP9 in mouse Sertoli cells. Andrology 2020, 8, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Kamińska, A.; Marek, S.; Pardyak, L.; Brzoskiwinia, M.; Bilińska, B.; Hejmej, A. Crosstalk between androgen signaling and Notch pathway in Sertoli cells. Int. J. Mol. Sci. 2020. submitted. [Google Scholar]
- Martínez-Hernández, J.; Seco-Rovira, V.; Beltrán-Frutos, E.; Ferrer, C.; Serrano-Sánchez, M.I.; Pastor, L.M. Proliferation, apoptosis, and number of Sertoli cells in the Syrian hamster during recrudescence after exposure to short photoperiod. Biol. Reprod. 2020, 102, 588–597. [Google Scholar] [CrossRef]
- Thomas, P.; Converse, A.; Berg, H.A. ZIP9, a novel membrane androgen receptor and zinc transporter protein. Gen. Comp. Endocrinol. 2018, 257, 130–136. [Google Scholar] [CrossRef]
- Matsuura, W.; Yamazaki, T.; Yamaguchi-Iwai, Y.; Masuda, S.; Nagao, M.; Andrews, G.K.; Kambe, T. SLC39A9 (ZIP9) Regulates Zinc Homeostasis in the Secretory Pathway: Characterization of the ZIP Subfamily I Protein in Vertebrate Cells. Biosci. Biotechnol. Biochem. 2009, 7, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- Yamasaki, S.; Sakata-Sogawa, K.; Hasegawa, A.; Suzuki, T.; Kabu, K.; Sato, E.; Kurosaki, T.; Yamashita, S.; Tokunaga, M.; Nishida, K.; et al. Zinc is a novel intracellular second messenger. J. Cell Biol. 2007, 21, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Ramisz, G.; Turek, W.; Chmurska-Gasowska, M.; Rak, A.; Pietsch-Fulbiszewska, A.; Galuszka, A.; Kotula-Balak, M.; Tarasiuk, K. Senescence and adiponectin signaling—studies in canine testis. Ann. Anat. 2020. accepted. [Google Scholar]
- Lustofin, K.; Niedbala, P.; Pawlicki, P.; Tuz, R.; Płachno, B.J.; Profaska-Szymik, M.; Galuszka, A.; Stolarczyk, P.; Gorowska-Wojtowicz, E.; Kotula-Balak, M. Senescent cells in rabbit, nutria and chinchilla testes. Animal Reprod. Sci. 2020. submitted. [Google Scholar]
- Reiter, R.J.; Johnson, L.Y. Elevated pituitary LH and depressed pituitary prolactin levels in female hamsters with pineal-induced gonadal atrophy and the effects of chronic treatment with synthetic LRF. Neuroendocrinology 1974, 14, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, A.H.; Rice, C.D.; Rahman, M.S.; Dong, J.; Thomas, P. Identification and characterization of membrane androgen receptors in the ZIP9 zinc transporter subfamily: I. Discovery in female atlantic croaker and evidence ZIP9 mediates testosterone-induced apoptosis of ovarian follicle cells. Endocrinology 2014, 155, 4237–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.P.; Wolfgang, C.D.; Hai, T. Analysis of ATF3, a transcription factor induced by physiological stresses and modulated by gadd153/Chop10. Mol. Cell Biol. 1996, 16, 1157–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, R.T., 3rd; Zhang, H.T. Senescent-induced dysregulation of cAMP/CREB signaling and correlations with cognitive decline. Brain Res. 2013, 21, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Nan, G. The extracellular signal-regulated kinase 1/2 pathway in neurological diseases: A potential therapeutic target. Int. J. Mol. Med. 2017, 39, 1338–1346. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Shi, B.; Zheng, H.; Min, L.; Yang, J.; Li, X.; Liao, X.; Huang, W.; Zhang, M.; Xu, S.; et al. Senescence-associated secretory factors induced by cisplatin in melanoma cells promote non-senescent melanoma cell growth through activation of the ERK1/2-RSK1 pathway. Cell Death Dis. 2018, 15, 260. [Google Scholar] [CrossRef]
- Buscà, R.; Pouysségur, J.; Lenormand, P. ERK1 and ERK2 Map Kinases: Specific Roles or Functional Redundancy? Fron. Cell Dev. Biol. 2016, 8, 53. [Google Scholar]
- Zou, J.; Lei, T.; Guo, P.; Yu, J.; Xu, Q.; Luo, Y.; Ke, R.; Huang, D. Mechanisms shaping the role of ERK1/2 in cellular senescence. Mol. Med. Rep. 2019, 19, 759–770. [Google Scholar]
- Roediger, J.; Hessenkemper, W.; Bartsch, S.; Manvelyan, M.; Huettner, S.S.; Liehr, T.; Esmaeili, M.; Foller, S.; Petersen, I.; Grimm, M.O.; et al. Supraphysiological androgen levels induce cellular senescence in human prostate cancer cells through the Src-Akt pathway. Mol. Cancer 2014, 12, 214. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, P.; Schweizer, M.T.; Lucas, J.M.; Coleman, I.; Nyquist, M.D.; Frank, S.B.; Tharakan, R.; Mostaghel, E.; Luo, J.; Pritchard, C.C.; et al. Supraphysiological androgens suppress prostate cancer growth through androgen receptor-mediated DNA damage. J. Clin. Investig. 2019, 16, 4245–4260. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.N.; Su, R.N.; Yang, B.Y.; Yang, K.X.; Yang, L.F.; Yan, Y.; Chen, Z.G. Potential Role of Cellular Senescence in Asthma. Front Cell. Dev. Biol. 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Gravina, G.L.; Marampon, F.; Sanità, P.; Festuccia, C.; Forcella, C.; Scarsella, L.; Jitariuc, A.; Vetuschi, A.; Sferra, R.; Colapietro, A.; et al. Episode-like pulse testosterone supplementation induces tumor senescence and growth arrest down-modulating androgen receptor through modulation of p-ERK1/2, pARser81 and CDK1 signaling: Biological implications for men treated with testosterone replacement therapy. Oncotarget 2017, 30, 113792–113806. [Google Scholar]
- Schmid, N.; Flenkenthaler, F.; Stöckl, J.B.; Dietrich, K.G.; Köhn, F.M.; Schwarzer, J.U.; Kunz, L.; Luckner, M.; Wanner, G.; Arnold, G.J.; et al. Insights into replicative senescence of human testicular peritubular cells. Sci. Rep. 2019, 21, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldakowski, L.; Taylor, J.R.E. Oxidative damage and antioxidant defense are assay and tissue-dependent both in captive and wild-caught bank voles (Myodes glareolus) before and after reproduction. Ecol. Evol. 2018, 8, 7543–7552. [Google Scholar] [CrossRef] [Green Version]
- Duliban, M.; Gorowska-Wojtowicz, E.; Tworzydlo, W.; Rak, A.; Brzoskwinia, M.; Krakowska, I.; Wolski, J.K.; Kotula-Balak, M.; Płachno, B.J.; Bilinska, B. Interstitial Leydig Cell Tumorigenesis-Leptin and Adiponectin Signaling in Relation to Aromatase. Int. J. Mol. Sci. 2020, 21, 3649. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host Species | Vendor | Dilution |
|---|---|---|---|
| ZIP9 | Rabbit | Sigma-Aldrich cat.no. SAB3500599 | 1:200 (IHC) 1:1500 (WB) |
| p16 | Rabbit | Abcam cat.no. ab151303 | 1:50 (IHC) 1:1000 (WB) |
| γH2AX | Mouse | Abcam cat.no. ab11174 | 1:250 (IHC) 1:1000 (WB) |
| ERK ½ | rabbit | Abcam cat. no. ab17942 | 1:1500 (WB) |
| β-actin | Mouse | Sigma–Aldrich cat. no. A2228 | 1:3000 (WB) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Profaska-Szymik, M.; Galuszka, A.; Korzekwa, A.J.; Hejmej, A.; Gorowska-Wojtowicz, E.; Pawlicki, P.; Kotula-Balak, M.; Tarasiuk, K.; Tuz, R. Implication of Membrane Androgen Receptor (ZIP9) in Cell Senescence in Regressed Testes of the Bank Vole. Int. J. Mol. Sci. 2020, 21, 6888. https://doi.org/10.3390/ijms21186888
Profaska-Szymik M, Galuszka A, Korzekwa AJ, Hejmej A, Gorowska-Wojtowicz E, Pawlicki P, Kotula-Balak M, Tarasiuk K, Tuz R. Implication of Membrane Androgen Receptor (ZIP9) in Cell Senescence in Regressed Testes of the Bank Vole. International Journal of Molecular Sciences. 2020; 21(18):6888. https://doi.org/10.3390/ijms21186888
Chicago/Turabian StyleProfaska-Szymik, Magdalena, Anna Galuszka, Anna J. Korzekwa, Anna Hejmej, Ewelina Gorowska-Wojtowicz, Piotr Pawlicki, Małgorzata Kotula-Balak, Kazimierz Tarasiuk, and Ryszard Tuz. 2020. "Implication of Membrane Androgen Receptor (ZIP9) in Cell Senescence in Regressed Testes of the Bank Vole" International Journal of Molecular Sciences 21, no. 18: 6888. https://doi.org/10.3390/ijms21186888