Methyl Parathion Masks Withdrawal from Physical Dependence on Morphine

Department of Pharmacology & Toxicology, University of Mississippi Medical Center, Jackson, MS, 39216, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2002, 3(10), 1073-1081; https://doi.org/10.3390/i3101073
Submission received: 7 June 2002
/
Accepted: 31 October 2002
/
Published: 31 October 2002
(This article belongs to the Special Issue Recent Advances in Environmental Health Research: Health Disparities, Toxicology and Carcinogenesis. Part II)
Abstract
:The cholinergic system has been proposed to participate in the development of dependence on opioids. The present study examined effects of dermal pretreatment with methyl parathion (MP), an acetylcholinesterase inhibitor, on the development of physical dependence on morphine. Opioid dependence was induced by continuous intracerebroventri-cular (i.c.v.) infusion of morphine (26 nmol/μl/h) for 3 days in adult male Sprague-Dawley rats. Each rat received two doses of MP, 12.5 mg/kg, dermally, initially, 3 days prior to initiation of i.c.v. morphine infusion and again on the first day of infusion. Withdrawal was precipitated after 3 days of infusion by administering an opioid antagonist, naloxone (48 nmol/5 μl, i.c.v.). Twelve of 23 MP-treated rats exhibited signs of acetylcholinesterase inhibitor intoxication (mild tremors) and showed reduced spontaneous locomotor activity (tested by an open field test), prior to naloxone. The brain cholinesterase activity in these 12 rats was 13% of levels in control rats. Eleven rats that did not show toxic signs, exhibited cholinesterase activities that were 20% of control (not significant versus toxic group). The group that showed signs of MP intoxication exhibited a significantly lower incidence of opioid withdrawal jumping, rearing and wet dog shakes compared with the non-toxic group. No differences between quantal withdrawal signs (ptosis, penis-licking, and vocalization) were noted between the two groups. The results suggest that toxic inhibition of acetylcholin-esterase non-specifically reduces locomotor activity and may obscure certain behavioral signs of withdrawal from opioid dependence. This indicates that caution should be used in interpreting a direct involvement of acetylcholinesterase inhibition in preventing opioid dependence.
Introduction
Chronic treatment with opioid drugs, such as morphine, leads to development of physical dependence, which is characterized by a continuous need for drug in order to avoid physical discomfort (withdrawal symptoms). The central cholinergic system has been suggested to participate in the action of opioids. As an inhibitory neuromodulator, a single dose of opioid agonists has been shown to decrease the release of and turnover rate for acetylcholine in several brain regions, including cortex, hippocampus, nucleus accumbens and globus pallidus [1,2,3,4,5]. However, after chronic opioid treatment, tolerance develops to the inhibitory effects of opioids and an increase in acetylcholine output occurs during precipitated withdrawal [6,7], suggesting a role for brain cholinergic mechanisms in opioid dependence and withdrawal [8,9].
Recently, activation of the central cholinergic system by administration of an acetylcholinesterase inhibitor, diisopropylfluorophosphate, has been reported to reduce the expression of withdrawal-induced increases in blood pressure, as well as the expression of certain withdrawal symptoms [10]. Anti-acetylcholinesterase agents elicit their effects through inhibition of acetylcholinesterase, thereby causing marked accumulation of acetylcholine in central and peripheral synapses and resulting in overstimulation of autonomic organs, skeletal muscle, and cholinergic receptors in the central nervous system [11]. Both acute and chronic treatments with an acetylcholinesterase inhibitor are known to result in depression of spontaneous locomotor activity and neuromuscular coordination [12]. This side effect of acetylcholinesterase inhibition may overshadow the expression of opioid withdrawal symptoms.
In the present study, the effects of pretreatment with an acetylcholinesterase inhibitor, methyl parathion (O-O-dimethyl-O-p-nitrophenyl phosphorothioate, MP), on the development and expression of morphine withdrawal were examined. The behavioral changes were correlated the effects with MP-induced changes in spontaneous locomotor activity and blood cholinesterase activity. Methyl parathion is an organophophorous compound that may only be used lawfully as an insecticide on agricultural crops and which has gained notoriety recently as a consequence of its illegal use as a pesticide in private homes and other buildings across several states. As of 1996, approximately 18,000 people have been affected as a result of direct contact by the illegal spraying of methyl parathion. However, there are limited data documenting the impact of exposure to methyl parathion on behavioral changes.
2. Materials and methods
2.1. Animals
Adult male Sprague-Dawley rats (Harlan Sprague-Dawley, Inc., Indianapolis, IN) weighing 250-275 grams were purchased and housed in groups of 3 or 4 animals in a cage. They were maintained under standard Laboratory Animal Facility conditions of 21 ± 2 °C and a 12-12 hour light-dark cycle with free access to food and water for a week before surgery. The experimental protocol was reviewed and approved by the University of Mississippi Medical Center Institutional Animal Care and Use Committee.
2.2. Induction of morphine dependence
Opioid dependence was induced by continuous intracerebroventricular (i.c.v.) infusion of morphine for 3 days. Rats were anesthetized with halothane (2% in oxygen) and then were placed in a stereotaxic apparatus. A stainless steel guide cannula (21 gauge, 10 mm long) was implanted into the right lateral cerebral ventricle (0.5 mm posterior, 1.3 mm lateral to bregma, 4.5 mm ventral) [13]. The presence of cerebrospinal fluid in the guide cannula was utilized as verification of proper placement. Dental acrylic cement (Lang Dental MFG Co., Wheeling, IL) was applied to the surface of the skull, and a protective aluminum cap was placed around the cannula. A stylet (26 gauge stainless steel tubing) was placed into the guide cannula to maintain cannula patency during the post-surgical recovery period. Housed individually, animals were allowed at least a week to recover from surgery.
Animals were rendered physically dependent by i.c.v. infusion of morphine sulfate (26 nmol morphine base/μl/h) for 3 days through osmotic minipumps (Alzet 2001, Alza Corp., Palo Alto, CA). Both the infusion period and dose paradigm have been well documented by previous studies to produce robust physical dependence [14]. Before introduction into the pump, the morphine solution was passed through 0.2 μm sterile Acrodisk filters (Gelman Science, Ann Arbor, MI). The minipumps were primed overnight at 35 °C in sterile saline so that the nominal flow rate (1 μl/h) was attained. Under halothane anesthesia, osmotic minipumps were implanted subcutaneously between the scapulae. A 4-cm piece of Tygon tubing (0.38 mm inner diameter, Cole-Palmer, Chicago, IL) was used to connect the outlet of the minipump to a piece of ‘L’-shaped stainless steel injector tubing (26 gauge, 15 mm long), which was placed into the i.c.v. guide cannula.
2.3. Pretreatment with MP
Rats (n=23) received two separate applications of MP, 12.5 mg/kg (~ 20% of the dermal LD50), dermally, the first at 3 days prior to initiation of i.c.v. morphine infusion and again on the first day of infusion. Rats were lightly anesthetized with halothane, and a portion of the back between the scapulae was shaved. MP (12.5 mg/kg) was applied to the shaven area in 100 μl ethanol. Control rats (n=12) were treated with ethanol vehicle.
2.4. Behavioral assessment
After 3 days of i.c.v. infusion with morphine, the effect of methyl parathion on spontaneous locomotor activity was tested by an open field test. The test was performed in a cardboard box (50 x 50 x 46 cm high) with clear Plexiglas on the inner surface. The floor of the box was divided into 25 equal squares. An animal was placed gently in the box and the number of squares crossed with all paws during the next 2 minutes was recorded. A 90% alcohol solution and soapy water were used to clean the inner surface of the box between trials to remove interfering odors left by the previous animal.
After the open field tests, the connecting tube between the i.c.v. cannula and the outlet of the minipump was severed and the implanted i.c.v. cannula flushed of any remaining morphine by injection of 5 μl of sterile saline through the cannula. Two hours following termination of the infusion, naloxone (48 μmol/5 μl) was injected i.c.v. by means of a hand-held microliter syringe and withdrawal behaviors were observed. The absolute frequency of 5 episodic behaviors was scored based on multiples of 5 incidents (0 = no incidents; 1 = 1-5 incidents; 2 = 5-10 incidents; and 3 = >11 incidents). Behaviors scored in this manner included: jumping (escape behaviors), rearing, wet-dog shakes, teeth-chattering, and scratching. Three behaviors (ptosis, vocalization, and penis-licking) cannot be defined in discrete episodes, and were recorded in an all-or-none manner. These withdrawal signs were scored during a 30-min period after naloxone injection. Behavioral observations occurred following placement of each animal individually into a transparent plastic cage containing bedding.
2.5. Brain cholinesterase assay
After the behavioral tests, rats were sacrificed by decapitation. The frontal cortex was dissected and homogenated for cholinesterase assays. Total cholinesterase activity was measured using a modified version [15] of the traditional spectrophotometric assay described by Ellman et al. [16]. Cholinesterase activity was calculated on the basis of a standard curve generated using increasing concentrations of glutathione and expressed as nmol acetylthiocholine iodide hydrolyzed per min after normalization to protein concentration. Protein concentrations were determined as described by Bradford [17].
2.6. Statistics
Data from open field tests and scores of opioid behaviors defined in discrete episodes (jumping, rearing, wet-dog shakes, teeth-chattering, and scratching) were analyzed by the nonparametric Kruskal-Wallis and Dunn’s tests. Quantal (all-or-none, including ptosis, vocalization, and penis licking) behavioral data were analyzed by the chi-square test. Brain cholinesterase activity was analyzed by one-way ANOVA, followed by the Student-Newman-Keuls test. Significance was set at P < 0.05.
3. Results
3.1. MP-induced toxic signs and suppression of spontaneous locomotor activity
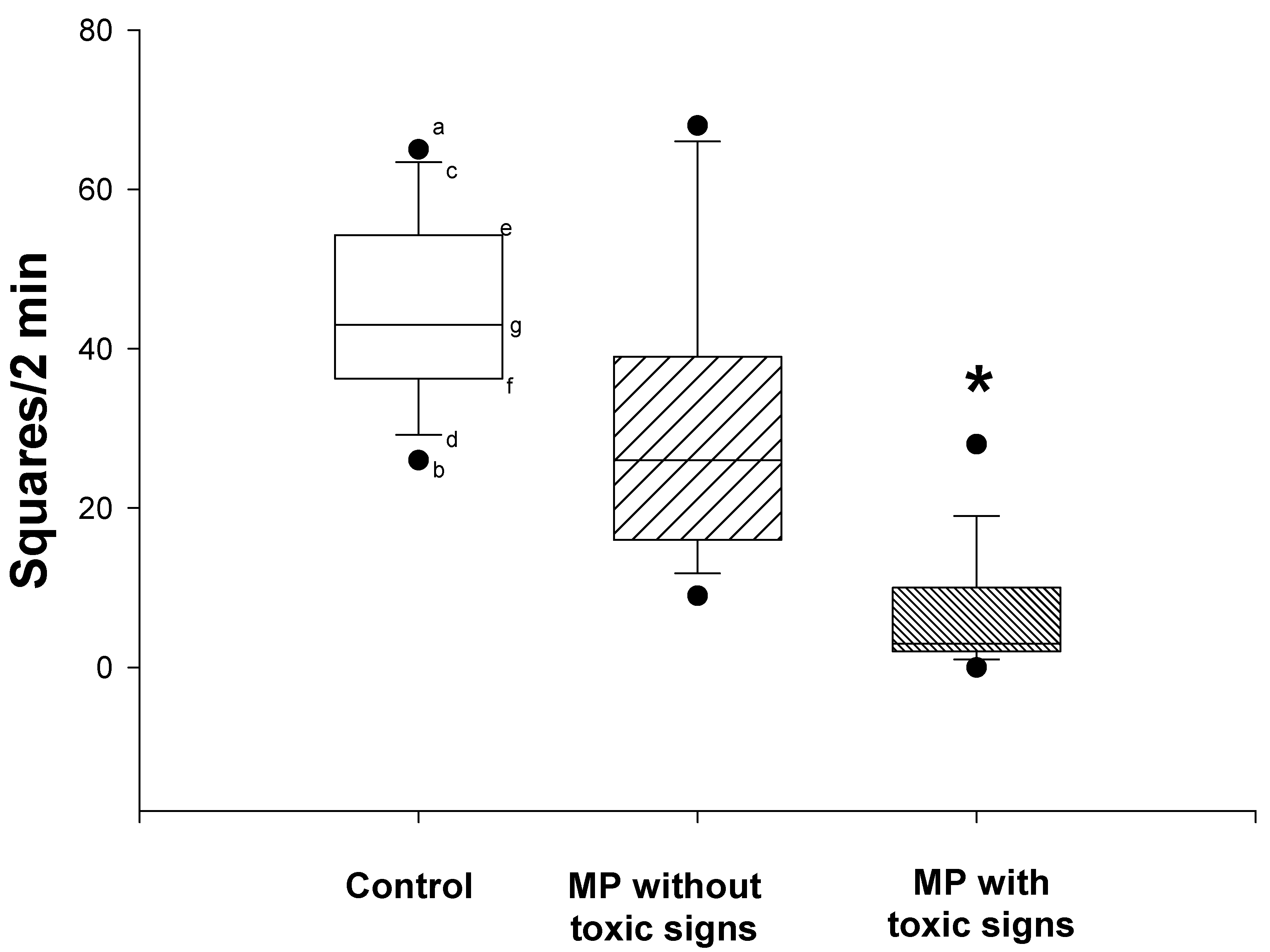
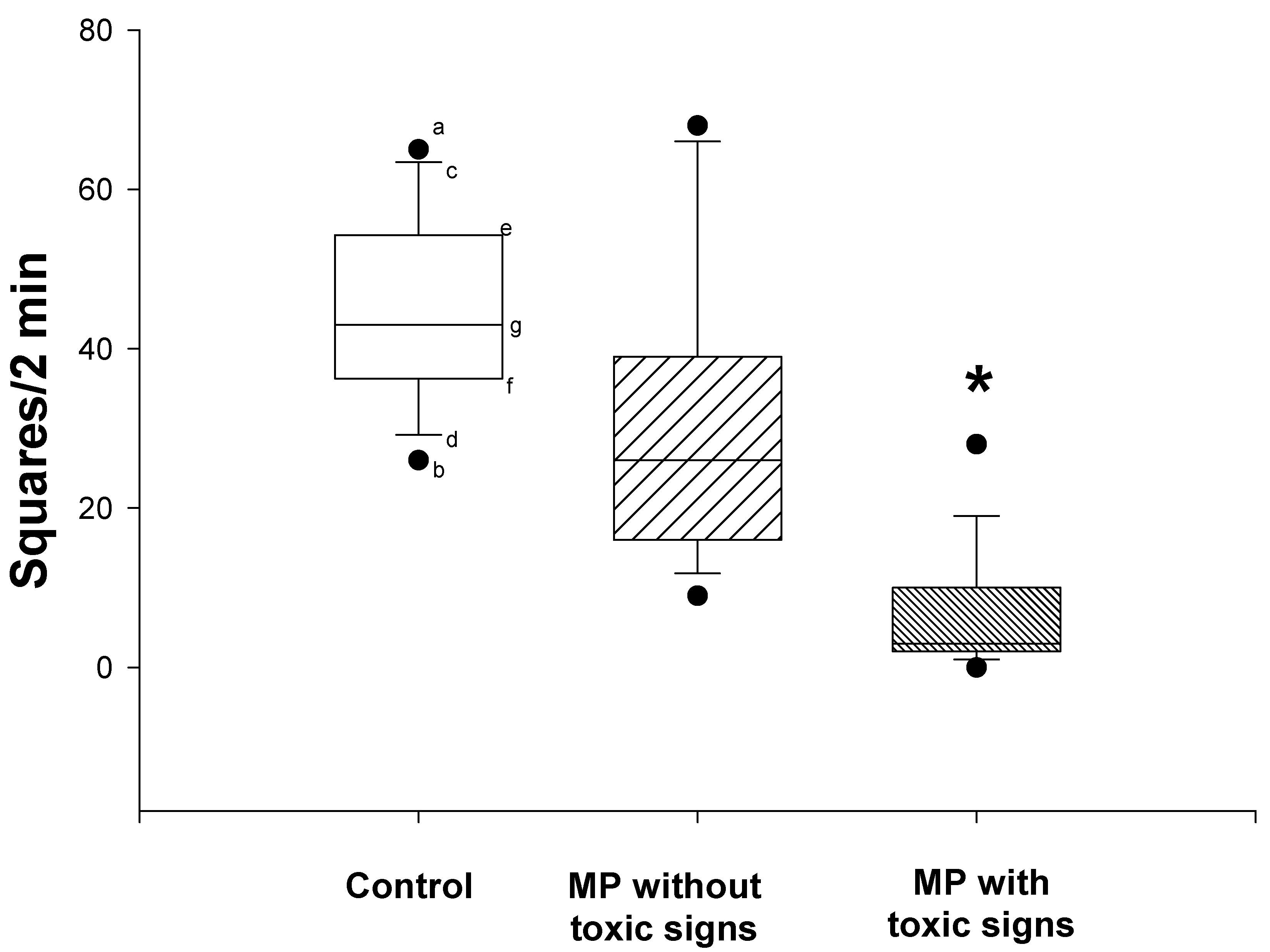
Twenty-three rats received two doses of MP, 12.5 mg/kg (dermal), 3 days prior to initiation of i.c.v. morphine infusion and again on the first day of infusion. Although all 23 rats received identical doses of MP, differences could be discerned in the apparent degree of intoxication. Thus, when examined 6 days after the first dose of MP, 12 of the 23 showed signs of acetylcholinesterase inhibitor intoxication, i.e., slight tremor and lacrimation, while the remaining 11 rats showed no signs of toxicity. Spontaneous locomotor activities, which were assessed by the open field test (Fig. 1), were significantly reduced in the rats that showed toxic signs when compared to either the control rats or the MP-treated rats that did not show toxic signs (nonparametric Kruskal-Wallis and Dunn’s tests; P<0.05). There was no significant difference in the spontaneous locomotor activity between the control rats and the MP-treated rats that did not show toxic signs.
Figure 1.
Effects of dermal administration of MP on spontaneous locomotor activity (open field tests). Open field tests were performed prior to termination of the morphine infusion. Data are expressed as the number of squares crossed by the rats within 2 minutes. Symbols represent: the median (g), 25th(e), 75th(f), 10th(c), and 90th(d) percentiles, maximum (a) and minimum (b) of 11-12 rats. *Significantly different from both control and non-toxic groups, P<0.05.
Figure 1.
Effects of dermal administration of MP on spontaneous locomotor activity (open field tests). Open field tests were performed prior to termination of the morphine infusion. Data are expressed as the number of squares crossed by the rats within 2 minutes. Symbols represent: the median (g), 25th(e), 75th(f), 10th(c), and 90th(d) percentiles, maximum (a) and minimum (b) of 11-12 rats. *Significantly different from both control and non-toxic groups, P<0.05.
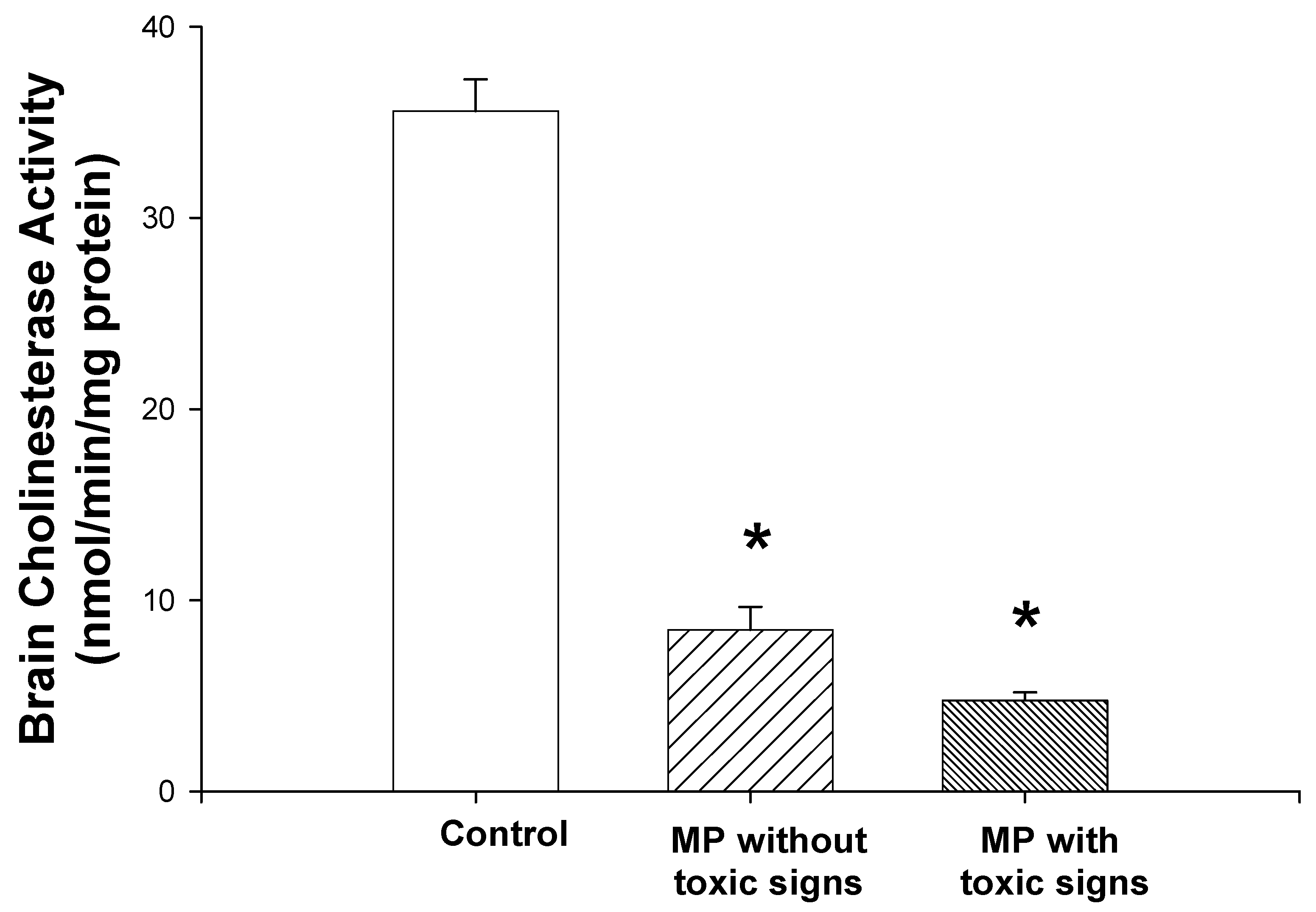
3.2. Brain cholinesterase activity
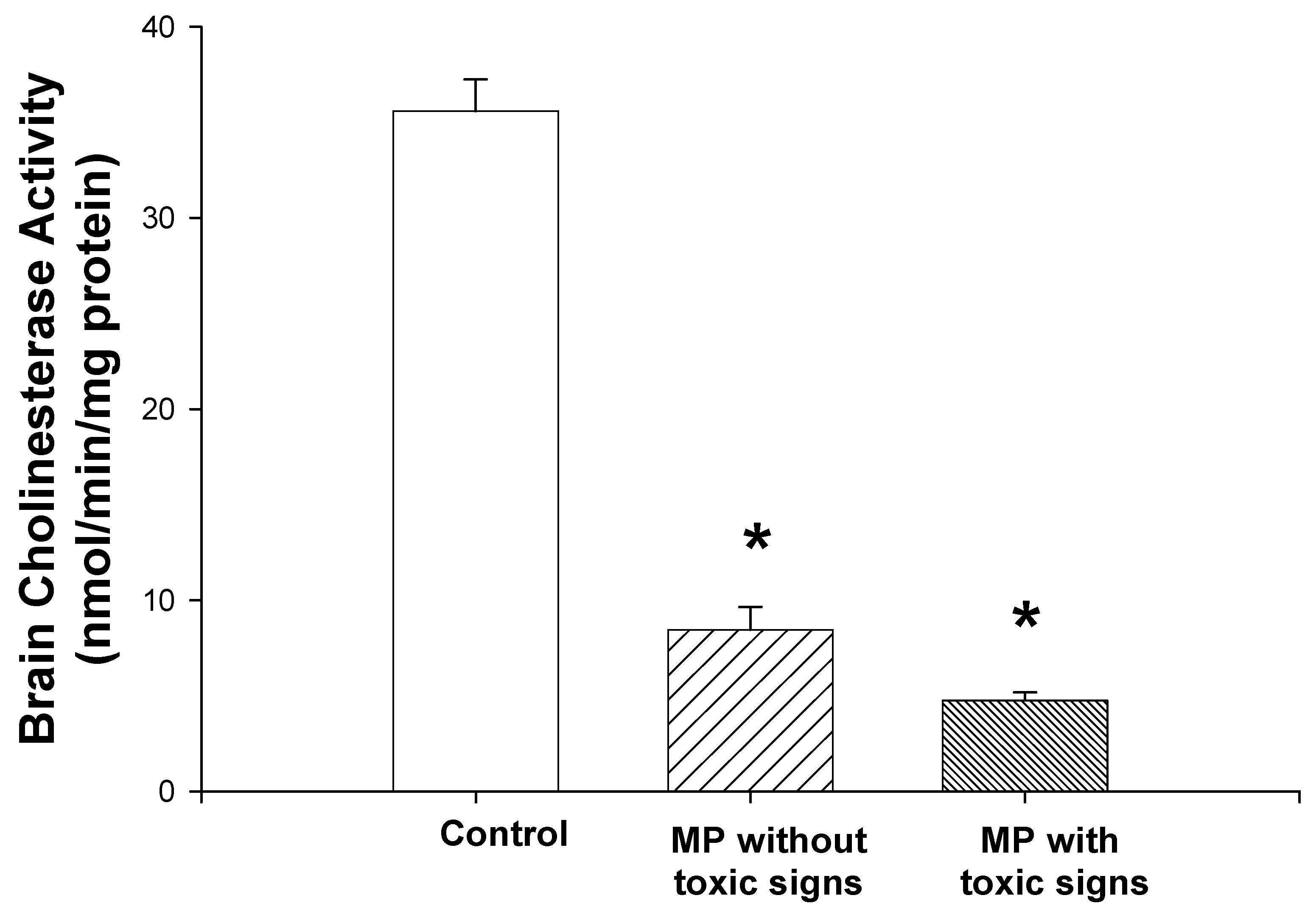
Brain cholinesterase activity was significantly decreased in both groups of MP-treated rats (one-way ANOVA and Student-Newman-Keuls tests, P<0.01; Fig. 2). Brain cholinesterase activity of the rats that showed toxic signs was 13% of the levels in control rats. The rats that did not show toxic signs, exhibited cholinesterase activities that were 20% of control. However, there was no significant difference in the cholinesterase activity between the rats with toxic signs and those without toxic signs.
Figure 2.
Effects of dermal administration of MP on brain cortical cholinesterase activities. Rats received two doses of MP, 12.5 mg/kg, dermally, 3 days prior to initiation of i.c.v. morphine infusion and again on the first day of infusion. Brain cholinesterase assays were performed as described in Materials and Methods. Reductions of 79.8 % (non-toxic group) and 87.1 % (toxic group) from the control group cholinesterase activities were noted. Data are presented as mean values + S.E.M. *Significantly different from the control group, P<0.05. There was not a significant difference between the values from the toxic group and those from the non-toxic group.
Figure 2.
Effects of dermal administration of MP on brain cortical cholinesterase activities. Rats received two doses of MP, 12.5 mg/kg, dermally, 3 days prior to initiation of i.c.v. morphine infusion and again on the first day of infusion. Brain cholinesterase assays were performed as described in Materials and Methods. Reductions of 79.8 % (non-toxic group) and 87.1 % (toxic group) from the control group cholinesterase activities were noted. Data are presented as mean values + S.E.M. *Significantly different from the control group, P<0.05. There was not a significant difference between the values from the toxic group and those from the non-toxic group.
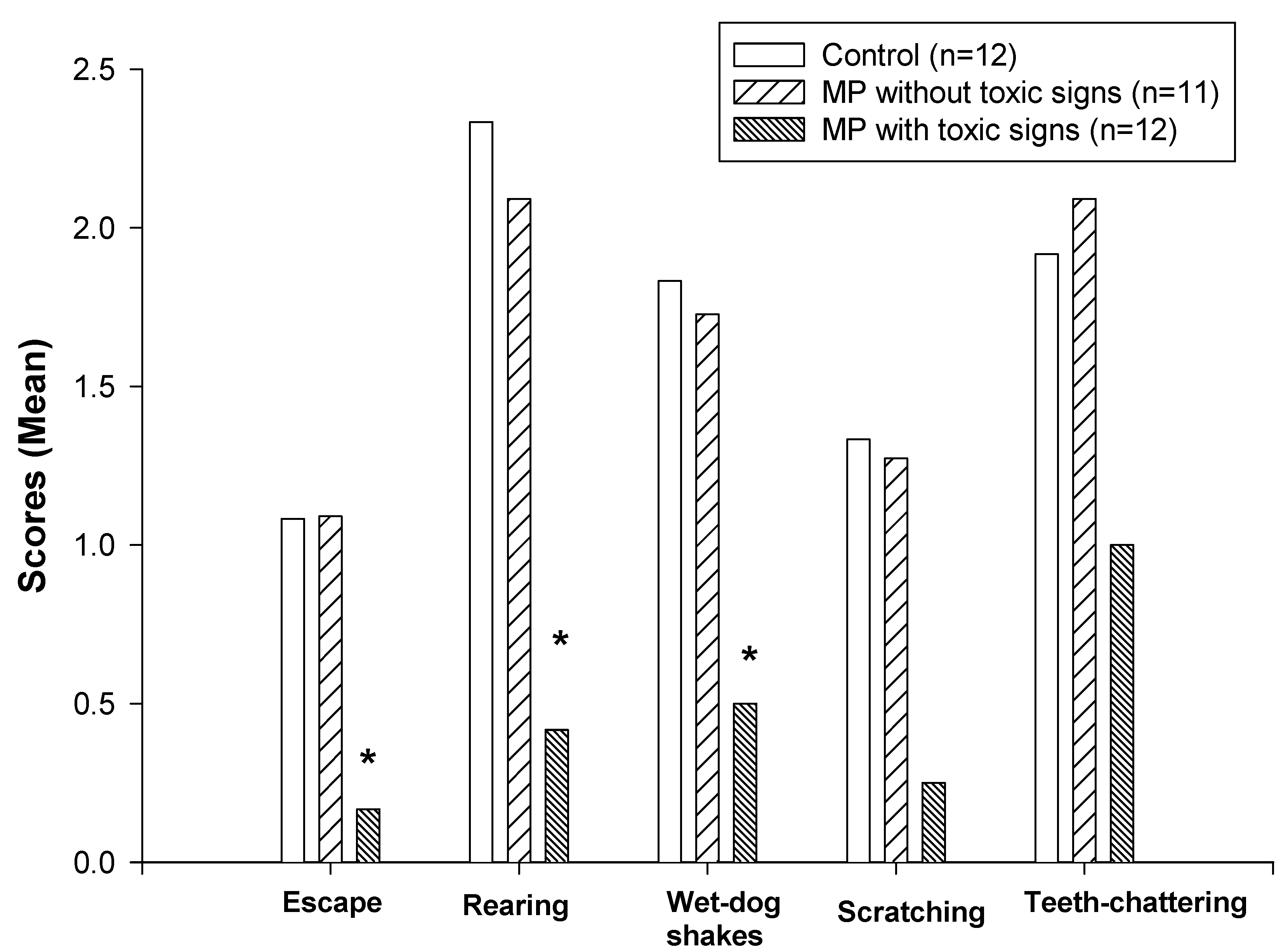
3.3. Naloxone-precipitated withdrawal from morphine
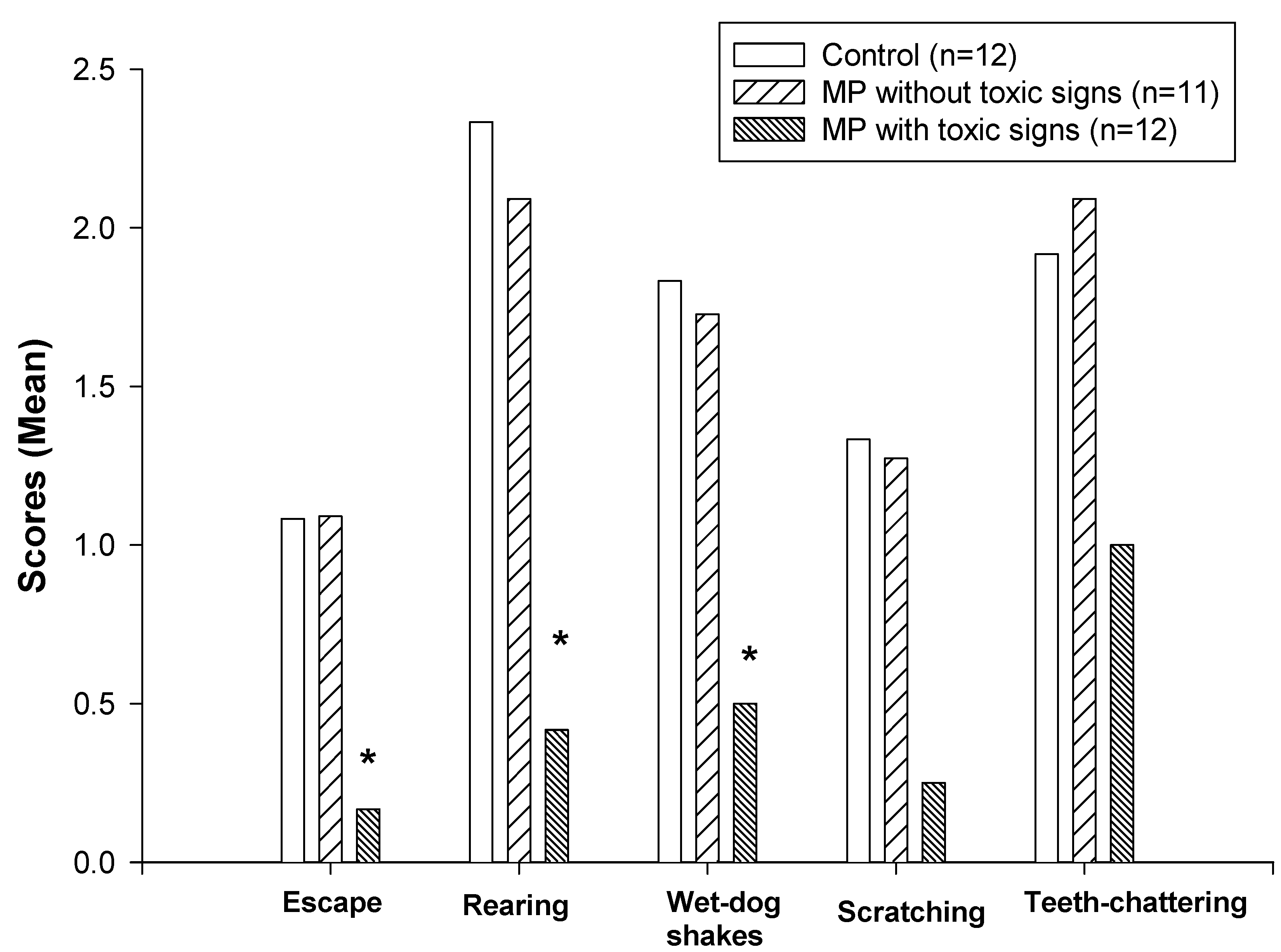
The effects of pretreatment with MP on naloxone-precipitated withdrawal behaviors are shown in Figure 3 and Table 1. The group that showed signs of MP intoxication and reduction of spontaneous locomotor activity exhibited a significantly lower incidence of opioid withdrawal behaviors (nonparametric Kruskal-Wallis and Dunn’s tests, P<0.05) including jumping, rearing and wet-dog shakes compared with the control group (Fig. 3). However, there was no significant decrease in the intensity of withdrawal behaviors in the MP-treated rats that did not show toxic signs. No differences in quantal withdrawal signs (ptosis, penis-licking, and vocalization) were noted among the groups (chi-square test).
4. Discussion
Brain cholinergic system has been proposed to be involved in opioid dependence and withdrawal, especially in the autonomic symptoms associated with opioid withdrawal [9]. During withdrawal, cholinergic activity is enhanced [6,7] and blockade of cholinergic activity by atropine or hemicholinium-3, a drug that depletes endogenous acetylcholine, has been shown to attenuate certain withdrawal behaviors [18]. In opposition to the enhanced activity during withdrawal, the function of the cholinergic system is inhibited by acute opioids [1,2,3,4,5]. Anticholinesterase agents, which augment cholinergic tone and were hypothesized to compensate the inhibitory effect of acute opioids, when co-administered with morphine, have been shown to attenuate certain withdrawal behaviors including escape attempts and body shakes [10]. However, systemic administration of acetylcholinesterase inhibitors causes accumulation of acetylcholine at synaptic terminals and stimulation, followed by depression of skeletal muscle, resulting in muscular weakness [11]. In the present study, the effects of pretreatment with an anticholinesterase agent, MP, on the expression of opioid withdrawal symptoms were examined and correlated with the changes in rat locomotor function. Dermal administration of MP (12.5 mg/kg), 3 days prior to initiation of i.c.v. morphine infusion and on the first day of infusion, resulted in an 83% inhibition of brain cholinesterase activity, which is comparable to the cholinesterase inhibition noted in other studies [10]. No significant reduction in the intensity of opioid withdrawal behaviors was noted in the MP-treated rats in which neither toxic signs nor a reduction in spontaneous locomotor activities was observed. However, a decreased intensity of opioid withdrawal behaviors was observed in the rats that exhibited both toxic signs and a reduction of spontaneous locomotor activity. The attenuated behaviors included jumping, rearing and wet-dog shakes, all of which are related to motor function in the rat. The side effects of acetylcholinesterase inhibitors make it difficult to conclude that these drugs specifically prevent the withdrawal behaviors. The possibility that non-specific motor depression induced by anticholinesterase intoxification overshadows the expression of certain withdrawal behaviors must be considered.
Figure 3.
Effects of dermal administration of MP on naloxone-precipitated withdrawal behaviors. Withdrawal behaviors were scored during a 30-minute period following naloxone injection. Data are presented as mean values only since the incidence of these behaviors are not normally distributed. *Significantly different from both the control and non-toxic groups, P<0.05.
Figure 3.
Effects of dermal administration of MP on naloxone-precipitated withdrawal behaviors. Withdrawal behaviors were scored during a 30-minute period following naloxone injection. Data are presented as mean values only since the incidence of these behaviors are not normally distributed. *Significantly different from both the control and non-toxic groups, P<0.05.

{kind=link}
{kind=link}
{kind=link}
| Control (N = 12) | MP with Toxic Signs (N = 12) | MP without Toxic Signs (N = 11) | |
|---|---|---|---|
| Vocalization | 5/12 | 6/12 | 4/11 |
| Penis licking | 7/12 | 7/12 | 5/11 |
| Ptosis | 10/12 | 11/12 | 9/11 |
* Data are presented as fractions (the number of rats showing positive signs over the total number of rats tested). There were no significant differences among the groups.
In conclusion, our results suggest that toxic inhibition of acetylcholinesterase non-specifically reduces locomotor activity and may obscure certain behavioral signs of withdrawal from opioid dependence. This indicates that caution should be used in interpreting a direct involvement of acetylcholinesterase inhibition in preventing opioid dependence.
Acknowledgements
This work was supported by grant RO6-CCR419466-01.
References and Notes
- Costa, E.; Cheney, D.L.; Racagni, G.; Zsilla, G. An analysis at synaptic level of the morphine action in striatum and N. accumbens: dopamine and acetylcholine interactions. Life Sci. 1975, 17(1), 1–8. [Google Scholar] [CrossRef] [PubMed]
- Moroni, F.; Cheney, D.L.; Costa, E. The turnover rate of acetylcholine in brain nuclei of rats injected intraventricularly and intraseptally with alpha and beta-endorphin. Neuropharmacology 1978, 17(3), 191–61. [Google Scholar] [CrossRef] [PubMed]
- Mulder, A.H.; Wardeh, G.; Hogenboom, F.; Frankhuyzen, A.L. Kappa- and delta-opioid receptor agonists differentially inhibit striatal dopamine and acetylcholine release. Nature 1984, 308(5956), 278–80. [Google Scholar] [CrossRef] [PubMed]
- Ennis, C.; Wyllie, M.G. Evidence for functionally distinct mu receptors modulating acetylcholine release. Neuropeptides 1984, 5(1-3), 109–12. [Google Scholar] [CrossRef] [PubMed]
- Lapchak, P.A.; Araujo, D. M.; Collier, B. Regulation of endogenous acetylcholine release from mammalian brain slices by opiate receptors: hippocampus, striatum and cerebral cortex of guinea-pig and rat. Neuroscience 1989, 31(2), 313–25. [Google Scholar] [CrossRef] [PubMed]
- Domino, E.F.; Wilson, A.E. Letter: Enhanced utilization of brain acetylcholine during morphine withdrawal in the rat. Nature 1973, 243(5405), 285–6. [Google Scholar] [CrossRef] [PubMed]
- Casamenti, F.; Pedata, F.; Corradetti, R.; Pepeu, G. Acetylcholine input from the cerebral cortex, choline uptake and muscarinic receptors in morphine-dependent, freely-moving rats. Neuropharmacology 1980, 19(7), 597–605. [Google Scholar] [CrossRef] [PubMed]
- Redmond, D.E.; Krystal, J.H. Multiple mechanisms of withdrawal from opioid drugs. Ann. Rev. Neurosci. 1984, 7, 443–78. [Google Scholar] [CrossRef] [PubMed]
- Buccafusco, J.J. Neuropharmacologic and behavioral actions of clonidine: interactions with central neurotransmitters. Intern. Rev. Neurobiol. 1992, 33, 55–107. [Google Scholar]
- Buccafusco, J.J.; Zhang, L.C.; Shuster, L.C.; Jonnala, R.R.; Gattu, M. Prevention of precipitated withdrawal symptoms by activating central cholinergic systems during a dependence-producing schedule of morphine in rats. Brain Res. 2000, 852(1), 76–83. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P. Anticholinesterase agents. In Goodman and Gillman’s The Pharmacological Basis of TherapeuticsHardman, J.G., Limbird, L.E., Molinoff, P.B., Ruddon, R.W., Gilman, A.G., Eds.; McGraw-Hill: New York, 9th ed.; 1996; chapter 8. [Google Scholar]
- Zhu, H.; Rockhold, R.W.; Baker, R.C.; Kramer, R.E.; Ho, I.K. Effects of repeated or single dermal administration of methyl parathion on behavior and cholinesterase activity in rats. J. Biomed. Sci. 2001, 8, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic CoordinatesAcademic Press: Orlando, 2nd ed.; 1986. [Google Scholar]
- Zhu, H.; Ho, I.K. NMDA-R1 antisense oligonucleotide attenuates withdrawal signs from morphine. Eur. J. Pharm. 1998, 352(2-3), 151–6. [Google Scholar] [CrossRef]
- Nostrandt, A. C.; Duncan, J.A.; Padilla, S. A modified spectrophotometric method appropriate for measuring cholinesterase activity in tissue from carbaryl-treated animals. Fund. Appl. Toxicol. 1993, 21(2), 196–203. [Google Scholar] [CrossRef]
- Ellman, G.C.; Courtney, K.O.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–54. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.C.; Buccafusco, J.J. Spinal cholinergic neurons and the expression of morphine withdrawal symptoms in the rat. J. Neurosci. 1987, 7(3), 621–8. [Google Scholar] [PubMed] [Green Version]
© 2002 by MDPI (http://www.mdpi.org).
Share and Cite
MDPI and ACS Style
Zhu, H.; Ho, I.K.; Kramer, R.E.; Baker, R.C.; Rockhold, R.W. Methyl Parathion Masks Withdrawal from Physical Dependence on Morphine. Int. J. Mol. Sci. 2002, 3, 1073-1081. https://doi.org/10.3390/i3101073
AMA Style
Zhu H, Ho IK, Kramer RE, Baker RC, Rockhold RW. Methyl Parathion Masks Withdrawal from Physical Dependence on Morphine. International Journal of Molecular Sciences. 2002; 3(10):1073-1081. https://doi.org/10.3390/i3101073
Chicago/Turabian StyleZhu, Hong, Ing K. Ho, Robert E. Kramer, Rodney C. Baker, and Robin W. Rockhold. 2002. "Methyl Parathion Masks Withdrawal from Physical Dependence on Morphine" International Journal of Molecular Sciences 3, no. 10: 1073-1081. https://doi.org/10.3390/i3101073